Introduction
Nasopharyngeal carcinoma (NPC) is a multifaceted
malignant tumor that originates from the nasopharynx epithelium
(1,2). Currently, radiotherapy combined with
chemotherapy is the main effective method for the treatment of NPC
(3). Despite significant progress
being made in the development of novel treatment methods for NPC,
its 5-year survival rate remains <40%, which is mainly due to
drug resistance (4,5). Therefore, it is vital to understand the
mechanism of cisplatin (DDP) resistance in NPC.
Long non-coding RNAs (lncRNAs) are a type of
non-coding RNA >200 nucleotides in length (6). Previous reports have revealed the
regulatory role of lncRNAs in multiple types of cancer (7–9). In
addition, lncRNAs have been demonstrated to exert crucial roles in
the chemoresistance of human cancer types, including NPC (10). For instance, lncRNA MAGI2-antisense
RNA 3 contributes to the DDP resistance of NPC by regulating the
microRNA (miRNA/miR)-218-5p/glycerophosphodiester phosphodiesterase
domain containing 5/SEC61 translocon subunit α1 axis (11). The lncRNA long intergenic non-protein
coding RNA 346 promotes DDP resistance in NPC cells by sponging
miR-342 (12). The knockdown of
deleted in lymphocytic leukemia 1 sensitizes NPC to DDP by
upregulating baculoviral IAP repeat containing 6 expression via
miR-381 inhibition (13). lncRNA
colon cancer associated transcript 1 regulates cytoplasmic
polyadenylation element binding protein 2 expression by
competitively binding to miR-181a, which results in the suppression
of NPC cell paclitaxel sensitivity (14). However, the role of homeobox
A11-antisense RNA (HOXA11-AS) in the DDP resistance of NPC remains
uncertain.
miRNAs are a class of small non-coding RNA 20–22
nucleotides in length that can regulate multiple biological
processes, including cell proliferation, metastasis,
differentiation, proliferation and apoptosis (15,16).
Previous studies have indicated that dysregulated miRNAs are
frequently involved in the progression of tumors via various
different mechanisms, including drug resistance (17,18). In
addition, several miRNAs have been reported to modulate the DDP
resistance of NPC. For example, miR-1278 suppresses the DDP
resistance of NPC cells by targeting autophagy-related 2B (19). The inhibition of miR-125b expression
facilitates DDP resistance and decreases DDP-induced apoptosis in
NPC (20). Nevertheless, to the best
of our knowledge, the involvement of miR-98 in DDP-resistant NPC
remains elusive.
The present study aimed to investigate the
biological role and molecular mechanism of HOXA11-AS in the
chemoresistance of NPC, which might provide therapeutic strategies
against the resistance of NPC to DDP.
Materials and methods
Patient samples
A total of 40 NPC tissues and adjacent normal
tissues (2 cm away from the tumor tissues) were collected from
patients (age range, 32–67 years; mean age, 56 years) who underwent
surgical resection surgery at The Shanghai Ninth People's Hospital
(Shanghai, China) between February 2017 and October 2019. Patients
who had recurrent disease within 6 months of completing primary
chemotherapy were classified as DDP-resistant (n=25). Patients with
recurrence beyond 6 months or without recurrence were classified as
DDP-sensitive (n=15). All patients who only had primary NPC were
included in the present study. Patients with NPC who had received
chemotherapy or radiotherapy prior to undergoing surgery were
excluded from the present study. All specimens were quickly frozen
in liquid nitrogen and stored at −80°C for subsequent use. The
present study was approved by the Ethics Committee of Shanghai
Ninth People's Hospital (Shanghai, China). All participants signed
the relevant informed consent forms. The classification of clinical
stage of patients with NPC was performed according to the American
Joint Committee on Cancer (AJCC) guidelines 6th Edition (21).
Cell culture
The NP-69 human normal nasopharyngeal epithelial
cell line (cat. no. BNCC338439), and the 5–8F (cat. no. BNCC353307)
and SUNE1 (cat. no. BNCC342441) human NPC cell lines were purchased
from BeNa Culture Collection (Beijing Beina Chunglian Biotechnology
Research Institute). 5–8F and SUNE1 cells were cultured in
RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific, Inc.)
containing streptomycin (100 U/ml) and 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) at 37°C in a humidified 5% CO2
environment. DDP-resistant NPC (5-8F/DDP and SUNE1/DDP) cell lines
were established by continuous exposure to progressively increasing
DDP (Sigma Aldrich; Merck KGaA) concentrations (0.5, 2, 4, 6 and 8
µg/ml) every 4 weeks. DDP-resistant NPC cells were cultured in
RPMI-1640 medium supplemented with 2 µg/ml DDP and 10% FBS at 37°C
in a humidified incubator with 5% CO2.
Cell transfection
Small interfering RNA (si) sequences specific for
HOXA11-AS (si-HOXA11-AS) or PBX3 (si-PBX3), scrambled control
sequences [si-negative control (NC)], miR-98 inhibitor, scrambled
inhibitor control (NC inhibitor), miR-98 mimics and scrambled mimic
negative control (NC mimics) were obtained from Sangon Biotech Co.,
Ltd. The sequences used were as follows: si-HOXA11-AS-1,
5′-CUACCAUCCCUGAGCCUUA-3′; si-PBX3, 5′-GCGAGCAUGUUUGTUGUUUU-3′;
si-NC, 5′-UCAAGUCCACGACGACUUUG-3′; miR-98 mimics,
5′-UGAGGUAGUAAGUUGUAUUGUU-3′; NC mimics,
5′-GUGUAACACGUCUAUACGCCCA-3′; miR-98 inhibitor,
5′-AACAAUACAACUUACUACCUC-3′; and NC inhibitor,
5′-ACUAUUGGAGAUUAGGUAUGUA-3′. To establish the role of HOXA11-AS or
PBX3, their corresponding overexpression plasmids were constructed
by cloning the full-length HOXA11-AS or PBX3 sequences into the
pcDNA3.1 plasmid vector (Invitrogen; Thermo Fisher Scientific,
Inc.). The final plasmids were referred to as pcDNA3.1-HOXA11-AS
(HOXA11-AS) or pcDNA3.1-PBX3 (PBX3). pcDNA3.1 was used as a
control. Cell were transfected with 50 nM miRNA mimic, 50 nM miRNA
inhibitor, 50 nM si-HOXA11-AS, 50 nM si-PBX3, 50 nM si-NC or 4.0 µg
pcDNA3.1-HOXA11-AS, 4.0 µg pcDNA3.1-PBX3 and 4.0 µg pcDNA3.1 using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C for 15 min according to the
manufacturer's protocol. After 48 h of transfection, the
transfected NPC cells were used for subsequent experiments.
Drug sensitivity assay
A Cell Counting Kit-8 (CCK-8) assay (Dojindo
Molecular Technologies, Inc.) was utilized to evaluate the
sensitivity of NPC cells to DDP. 5–8F/DDP and SUNE1/DDP cells
(1×104 cells/well) were plated into 96-well plates and
treated with DDP (10 µg) for 48 h. Subsequently, 10 µl CCK-8
solution was added to each well and incubated with the cells for 1
h. The absorbance was recorded using a microplate reader (BioTek
China) at 450 nm. DDP sensitivity was assessed using the
IC50.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from NPC tissues and cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). cDNA was obtained following the reverse
transcription of RNA using a PrimeScript RT Reagent kit (Takara
Bio, Inc.) according to the manufacturer's protocols. RT-qPCR was
conducted using the SYBR Green RT-qPCR Master mix (Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows:
Pre-denaturation at 95°C for 1 min, followed by 40 cycles of 95°C
for 15 sec, 60°C for 30 sec and 72°C for 30 sec. The data were
analyzed using the 2−ΔΔCq method (22). GAPDH or U6 was used as the internal
control for HOXA11-AS and PBX3 or miR-98, respectively. The primer
sequences were as follows: HOXA11-AS forward,
5′-ACGCTAGGCACCACTTTGTT-3′ and reverse, 5′-CCGGCTACTAGTCAGTGTGA-3′;
miR-98-5p forward, 5′-ACACTCCCUAUACAACUUAC-3′ and reverse,
5′-GGGAAAGUAGUGAGGCCTCAGA-3′; PBX3 forward,
5′-CGAGGCGCAAGCAAAGAAAC-3′ and reverse,
5′-TGCCAAAAGCATATTGTCCAGT-3′; GAPDH forward,
5′-CCCACTCCTCCACCTTTGAC-3′ and reverse,
5′-GGATCTCGCTCCTGGAAGATG-3′; and U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′.
TUNEL staining
5-8F/DDP and SUNE1/DDP cells were washed with PBS,
and fixed in 4% paraformaldehyde solution for 1 h at 4°C. The cells
were permeabilized in a solution containing 0.1% Triton X-100 for 2
min, and incubated in TUNEL reaction mixture for 1 h at 37°C.
Subsequently, the TUNEL-stained cells were washed with PBS 3 times
and counterstained with DAPI (2 µg/ml; Beyotime Institute of
Biotechnology) under antifade mounting medium (cat. no. P0126;
Beyotime Institute of Biotechnology) for 15 min at room
temperature. The number of apoptotic cells was counted in ≥5 fields
of view under a fluorescence microscope.
Bioinformatics prediction and dual
luciferase reporter assay
The downstream target genes of HOXA11-As and miR-98
were screened using the Starbase version 2.0 (http://starbase.sysu.edu.cn) and TargetScan version
7.2 (http://www.targetscan.org/vert_72/) websites,
respectively. The wild-type (WT) and mutant (Mut) sequences of
HOXA11-AS and PBX3 were cloned into the pmirGLO vector (Promega
Corporation). Subsequently, these constructed vectors were
co-transfected with NC mimics (5′-GUGUAACACGUCUAUACGCCCA-3′) and
miR-98 mimics (5′-UGAGGUAGUAAGUUGUAUUGUU-3′) from Sangon Biotech
Co., Ltd into 5–8F/DDP and SUNE1/DDP cells for 48 h using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
After 48 h, the relative luciferase activities were measured using
a Dual-Luciferase Reporter Assay System (Promega Corporation).
Firefly luciferase activity was normalized to Renilla
luciferase activity.
RNA immunoprecipitation (RIP)
The RIP assay was conducted to assess the
association between HOXA11-AS and miR-98 using the EZMagna RIP kit
(EMD Millipore). The cells were lysed with 100 µl RIP Lysis Buffer
(EMD Millipore) for 5 min at 4°C, the obtained cell lysate (100 µl)
was centrifuged at 40,000 × g at 4°C for 10 min and incubated with
50 µl A/G magnetic beads conjugated with argonaute 2 (Ago2; 5 µg;
cat. no. 2897; Cell Signaling Technology, Inc.) antibody and IgG (5
µg; cat. no. PP64B; EMD Millipore) antibody. Subsequently, the
beads were washed 3 times using RIP Wash Buffer. RNA was extracted
from the immunoprecipitation complex using 50 µg/ml Proteinase K
(cat. no. P2308; Sigma-Aldrich; Merck KGaA) at 55°C for 30 min to
digest the protein and the enrichment levels of HOXA11-AS and
miR-98 were measured using RT-qPCR as mentioned previously.
Western blot analysis
Total proteins were extracted from the treated cells
using RIPA lysis buffer (Beyotime Institute of Biotechnology). The
protein concentration was detected using a BCA protein quantitation
kit. The proteins (40 µg/lane) were separated by 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes. Following
blocking for 2 h with 5% skimmed dry milk at room temperature, the
membranes were incubated with antibodies against Bcl-2 (1:1,000;
cat. no. ab32124; Abcam), Bax (1:1,000; cat. no. ab32503; Abcam)
and GAPDH (1:1,000; cat. no. ab9485; Abcam) at 4°C overnight.
Subsequently, the membranes were incubated with horseradish
peroxidase-conjugated secondary antibody (1:1,000; cat. no.
ab205718; Abcam) for 1 h at 37°C. Finally, the protein expression
levels were examined using Pierce™ ECL A (Pierce; Thermo Fisher
Scientific, Inc.) and analyzed using Quantity One software version
4.6 (Bio-Rad Laboratories, Inc.).
Statistical analysis
All experiments were repeated in triplicate.
Statistical analysis was performed using SPSS version 16.0 (SPSS,
Inc.) and the results are presented as the mean ± standard
deviation. An unpaired Student's t-test was performed to determine
differences between two groups, and a paired Student's t-test was
used to determine differences between NPC tissues and adjacent
normal tissues. Differences among multiple groups were analyzed
using one-way ANOVA followed by Tukey's post hoc test. Kaplan-Meier
analysis and a log-rank test were used to assess the association
between HOXA11-AS expression and patient survival, and patients
were divided into high and low expression groups based on the mean
value of HOXA11-AS expression in patients with NPC. In addition,
the association between PBX3 expression and clinicopathological
characteristics of patients with NPC was analyzed using
χ2 tests, and the cut-off value (1.92) was the mean
expression of PBX3 in patients with NPC. P<0.05 was considered
to indicate a statistically significant difference.
Results
HOXA11-AS levels are increased in
DDP-resistant NPC tissues and cells
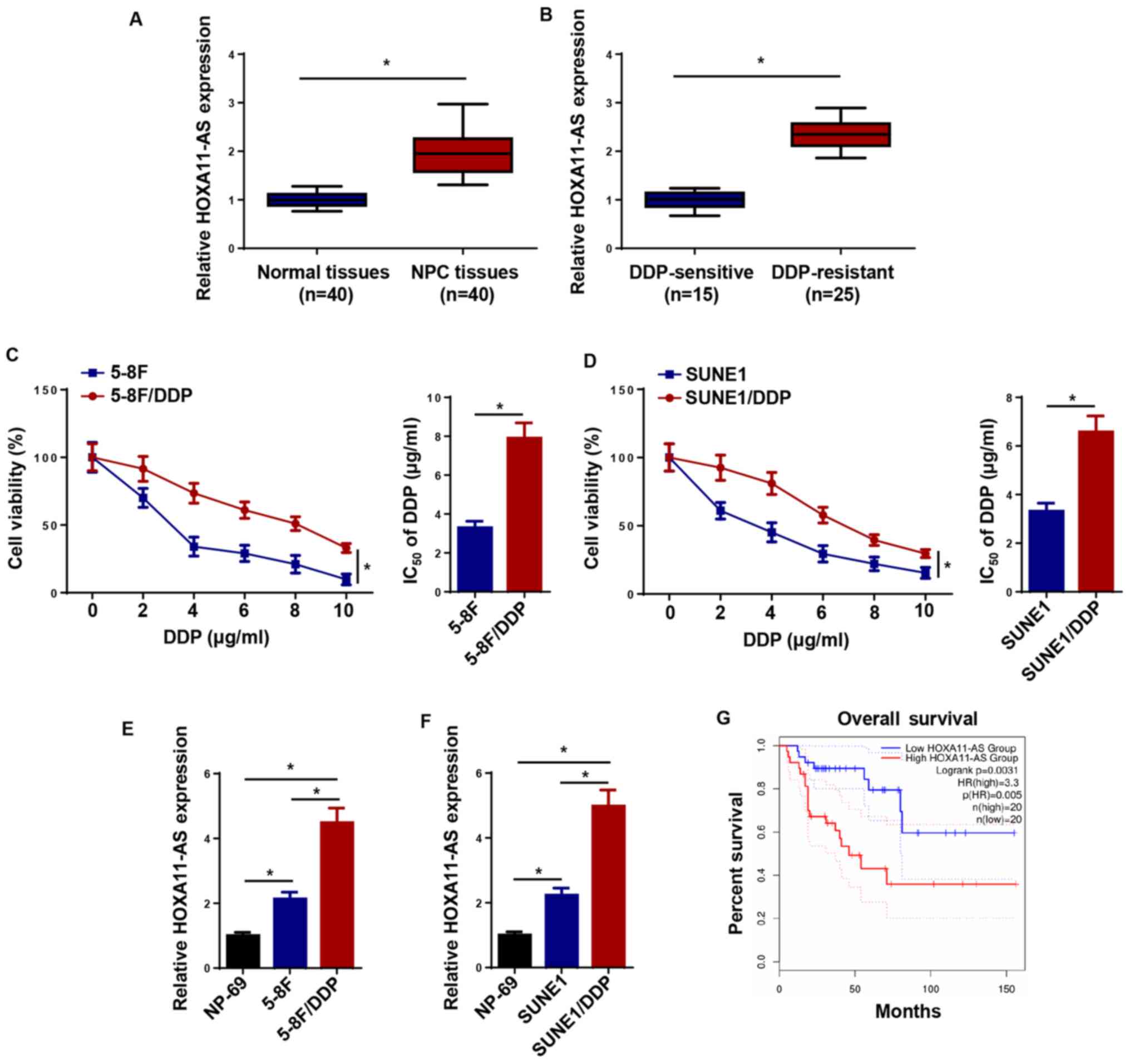
The expression levels of HOXA11-AS were increased in
NPC tissues compared with the levels in normal tissues (Fig. 1A). Furthermore, HOXA11-AS expression
was higher in DDP-resistant NPC tissues compared with that in
DDP-sensitive NPC tissues (Fig. 1B).
To determine whether DDP-resistant NPC cell lines (5-8F/DDP and
SUNE1/DDP) were successfully established, the IC50 of
DDP was measured using a CCK-8 assay. The results indicated that
5–8F/DDP and SUNE1/DDP cells displayed low sensitivity to DDP, as
demonstrated by the increased IC50 values (Fig. 1C and D). Furthermore, compared with
NP-69 cells, 5–8F and SUNE1 cells exhibited elevated expression
levels of HOXA11-AS, which were further upregulated in
DDP-resistant NPC cells (5-8F/DDP and SUNE1/DDP cells; Fig. 1E and F). In addition, patients with
high HOXA11-AS expression exhibited shorter survival times compared
with patients with low expression levels of HOXA11-AS (Fig. 1G). Overall, the data suggested that
HOXA11-AS upregulation may be involved in NPC DDP resistance.
Knockdown of HOXA11-AS attenuates DDP
resistance of NPC cells
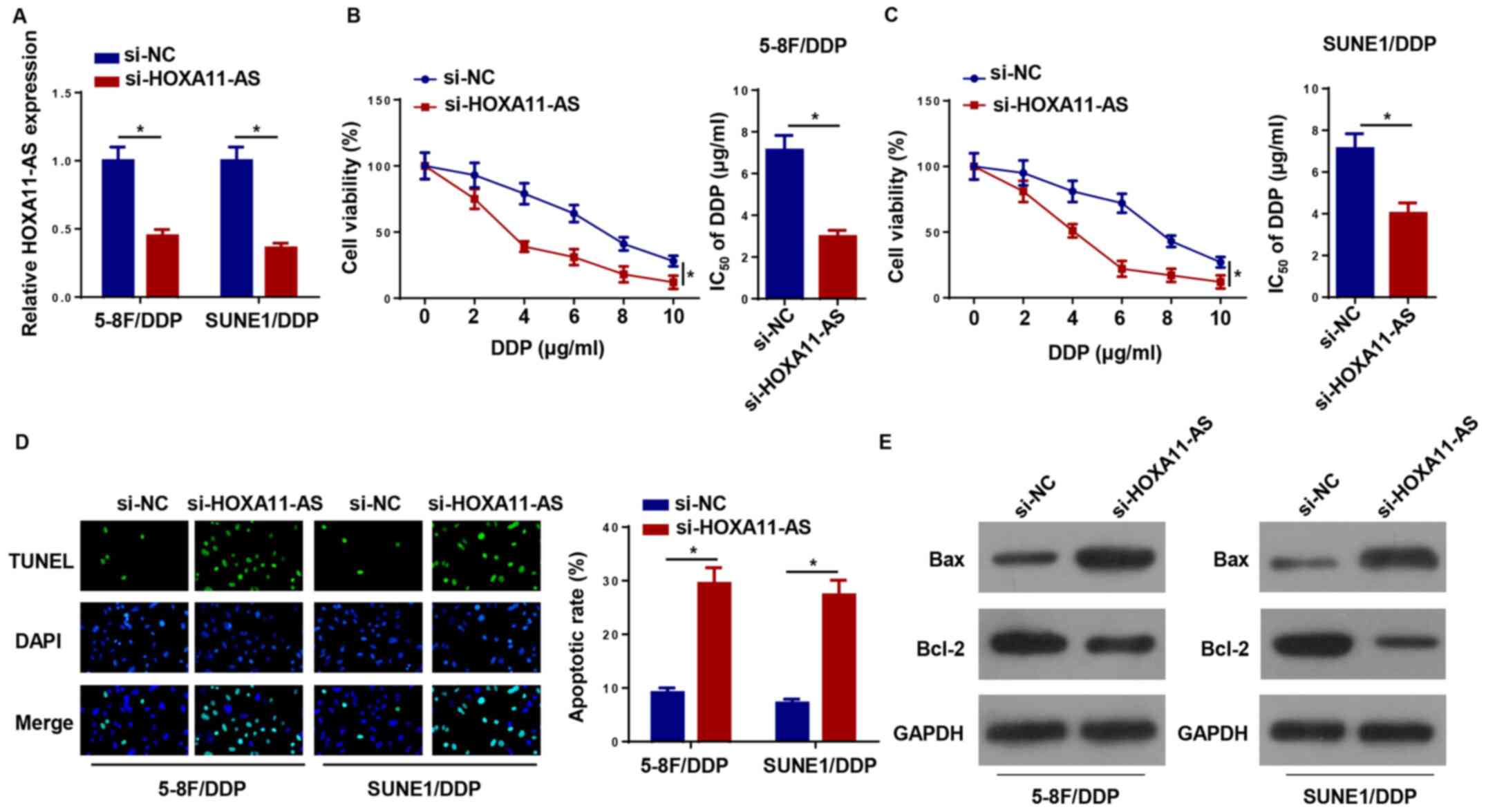
To further estimate the effect of HOXA11-AS on
DDP-resistant NPC cells, si-HOXA11-AS was transfected into 5–8F/DDP
and SUNE1/DDP cells to knockdown HOXA11-AS expression (Fig. 2A). Notably, HOXA11-AS knockdown
increased the DDP sensitivity of 5–8F/DDP and SUNE1/DDP cells
(Fig. 2B and C). Furthermore,
silencing of HOXA11-AS promoted the apoptosis of 5–8F/DDP and
SUNE1/DDP cells, which indicated that the downregulation of
HOXA11-AS increased the chemosensitivity of DDP-resistant NPC cells
(Fig. 2D). The expression levels of
apoptosis-associated proteins (Bax and Bcl-2) were assessed by
western blotting. HOXA11-AS depletion increased the protein
expression levels of Bax and inhibited Bcl-2 expression in
DDP-resistant NPC cells (Fig. 2E).
Collectively, the data indicated that HOXA11-AS-knockdown
suppressed DDP resistance in NPC cells.
HOXA11-AS acts as a molecular sponge
for miR-98
At present, multiple studies have suggested that
lncRNAs exert their biological function by interacting with
specific miRNAs (23,24). Therefore, StarBase version 2.0 was
used to uncover potential miRNAs, which could interact with
HOXA11-AS. The data demonstrated that miR-98 contained
complementary bases pairing with HOXA11-AS (Fig. 3A). RT-qPCR indicated that miR-98
expression was upregulated following miR-98 mimic transfection in
5–8F/DDP and SUNE1/DDP cells compared with in cells transfected
with NC mimics (Fig. 3B). Luciferase
reporter assays revealed that miR-98 mimics decreased the
luciferase activity of HOXA11-AS-WT, whereas this effect was not
noted for HOXA11-AS-Mut (Fig. 3C and
D). In addition, HOXA11-AS and miR-98 were highly enriched by
Ago2, while IgG was not significantly enriched (Fig. 3E and F). These results demonstrated
that HOXA11-AS could directly bind to miR-98. The HOXA11-AS
overexpression plasmid increased the expression levels of HOXA11-AS
in both 5–8F/DDP and SUNE1/DDP cells compared with cells
transfected with si-NC (Fig. 3G). To
investigate whether HOXA11-AS could regulate the expression levels
of miR-98, si-HOXA11-AS and HOXA11-AS overexpression plasmids were
transfected into 5–8F/DDP and SUNE1/DDP cells. The results of
RT-qPCR indicated that knockdown of HOXA11-AS increased miR-98
expression, whereas the transfection with HOXA11-AS plasmid
decreased miR-98 expression (Fig. 3H and
I). Furthermore, miR-98 expression was markedly decreased in
NPC tissues and cell lines, particularly in DDP-resistant NPC
tissues and cell lines (Fig. 3J-N).
In summary, the results demonstrated that HOXA11-AS acted as a
miR-98 sponge in NPC cells.
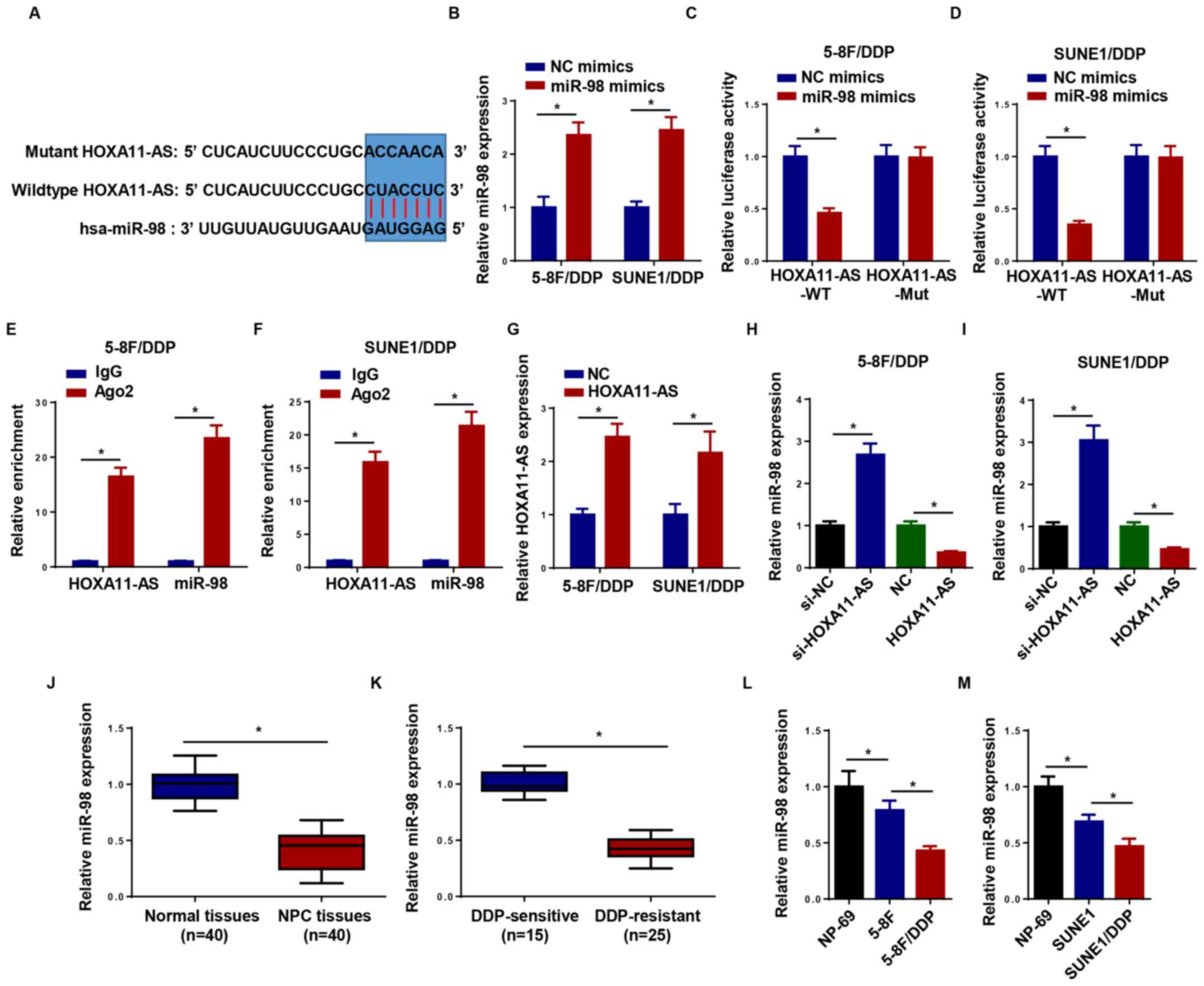 | Figure 3.HOXA11-AS serves as a molecular
sponge for miR-98. (A) Putative binding regions of HOXA11-AS and
miR-98 predicted using biological software. (B) RT-qPCR was used to
detect the expression levels of miR-98 in 5–8F/DDP and SUNE1/DDP
cells transfected with NC mimics or miR-98 mimics. Luciferase
activity of WT or Mut HOXA11-AS in (C) 5–8F/DDP and (D) SUNE1/DDP
cells following transfection with NC mimics or miR-98 mimics as
determined by a luciferase reporter assay. The interaction between
HOXA11-AS and miR-98 was further confirmed using an RNA
immunoprecipitation assay in (E) 5–8F/DDP and (F) SUNE1/DDP cells.
(G) Expression levels of HOXA11-AS in 5–8F/DDP and SUNE1/DDP cells
transfected with NC or HOXA11-AS were detected by RT-qPCR. miR-98
expression was detected in (H) 5–8F/DDP and (I) SUNE1/DDP cells
transfected with si-HOXA11-AS or HOXA11-AS overexpression plasmid.
(J) miR-98 expression was measured in NPC tissues (n=40) or normal
tissues (n=40). (K) miR-98 expression in DDP-sensitive or
DDP-resistant NPC tissues was measured by RT-qPCR. (L) miR-98
expression in NP-69, 5–8F and 5–8F/DDP cells was measured by
RT-qPCR. (M) miR-98 expression in NP-69, SUNE1 and SUNE1/DDP cells
was measured by RT-qPCR. Data are presented as the mean ± SD.
*P<0.05. Ago2, argonaute RISC catalytic component 2; DDP,
cisplatin; HOXA11-AS, homeobox A11-antisense RNA; miR-98,
microRNA-98; Mut, mutant; NC, negative control; NPC, nasopharyngeal
carcinoma; RT-qPCR, reverse transcription-quantitative PCR; si,
small interfering RNA; WT, wild-type. |
miR-98 inhibition abolishes the
effects of si-HOXA11-AS on DDP resistance in NPC cells
The levels of miR-98 were markedly decreased in
5–8F/DDP and SUNE1/DDP cells transfected with miR-98 inhibitor
(Fig. 4A). To further explore
whether HOXA11-AS regulated the DDP resistance of NPC via miR-98,
5–8F/DDP and SUNE1/DDP cells were transfected with si-NC,
si-HOXA11-AS and si-HOXA11-AS + miR-98 inhibitor. RT-qPCR analysis
indicated that HOXA11-AS interference upregulated the expression
levels of miR-98, whereas the treatment of the cells with the
miR-98 inhibitor partially counteracted the induced expression of
miR-98 (Fig. 4B). Furthermore,
knockdown of HOXA11-AS increased the DDP sensitivity of 5–8F/DDP
and SUNE1/DDP cells, which was reversed following the transfection
of the cells with the miR-98 inhibitor (Fig. 4C). Furthermore, the miR-98 inhibitor
neutralized the promoting effects of si-HOXA11-AS on the induction
of cell apoptosis (Fig. 4D). The
depletion of HOXA11-AS increased the protein expression levels of
Bax and inhibited Bcl-2 expression, which was reversed following
the transfection of cells with the miR-98 inhibitor (Fig. 4E). The data indicated that HOXA11-AS
silencing promoted the DDP sensitivity of NPC cells by sponging
miR-98.
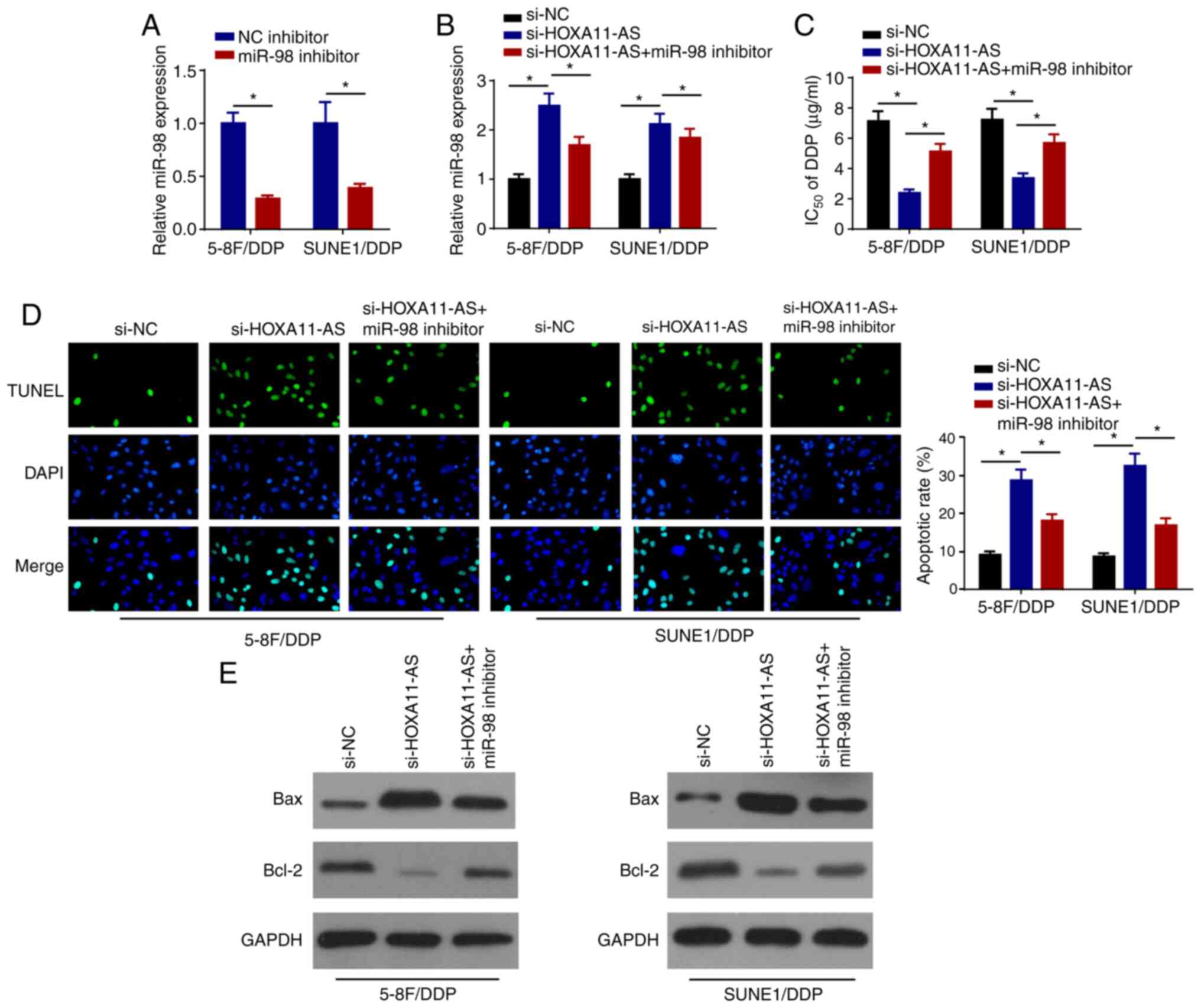 | Figure 4.miR-98 inhibition abolishes the
effects of si-HOXA11-AS on DDP resistance of nasopharyngeal
carcinoma cells. (A) RT-qPCR was performed to examine the
expression levels of miR-98 in 5–8F/DDP and SUNE1/DDP cells
transfected with NC inhibitor or miR-98 inhibitor. (B) RT-qPCR
analysis was performed to determine miR-98 expression in 5–8F/DDP
and SUNE1/DDP cells transfected with si-NC, si-HOXA11-AS or
si-HOXA11-AS + miR-98 inhibitor. (C) IC50 of DDP was
evaluated in 5–8F/DDP and SUNE1/DDP cells transfected with si-NC,
si-HOXA11-AS or si-HOXA11-AS + miR-98 inhibitor using a Cell
Counting Kit-8 assay. (D) Cell apoptosis was detected by TUNEL
staining (magnification, ×200) in 5–8F/DDP and SUNE1/DDP cells
transfected with si-NC, si-HOXA11-AS or si-HOXA11-AS + miR-98
inhibitor. (E) Protein expression levels of Bax and Bcl-2 in
5–8F/DDP and SUNE1/DDP cells transfected with si-NC, si-HOXA11-AS
or si-HOXA11-AS + miR-98 inhibitor were detected by western blot
analysis. Data are presented as the mean ± SD. *P<0.05. DDP,
cisplatin; HOXA11-AS, homeobox A11-antisense RNA; miR-98,
microRNA-98; NC, negative control; RT-qPCR, reverse
transcription-quantitative PCR; si, small interfering RNA. |
HOXA11-AS confers DDP resistance by
upregulating PBX3 expression in NPC cells via miR-98 sponging
TargetScan version 7.2 was used to predict the
candidate targets of miR-98. As shown in Fig. 5A, PBX3 contained a miR-98 binding
sequence in the 3′-untranslated region. Furthermore, it was
revealed that miR-98 mimics markedly decreased the relative
luciferase activity of PBX3-WT, while the activity of PBX3-Mut
remained unchanged, suggesting that PBX3 was a direct target of
miR-98 (Fig. 5B and C). The effects
of miR-98 and HOXA11-AS on the regulation of PBX3 expression were
investigated in 5–8F/DDP and SUNE1/DDP cells following transfection
with NC mimics, miR-98 mimics, si-NC, si-HOXA11-AS and si-HOXA11-AS
+ miR-98 inhibitor. RT-qPCR analysis revealed that PBX3 expression
was downregulated in DDP-resistant NPC cells transfected with
miR-98 mimics and si-HOXA11-AS, whereas the inhibitory effects of
HOXA11-AS-knockdown on PBX3 expression were abrogated by miR-98
inhibition (Fig. 5D). Upregulated
levels of PBX3 were directly positively associated with lymph node
metastasis, distant metastasis and clinical stage (Table I). In addition, PBX3 levels were
upregulated in NPC tissues and DDP-resistant NPC tissues (Fig. 5E and F). To further explore whether
HOXA11-AS was associated with NPC DDP resistance by regulating PBX3
expression, 5–8F/DDP and SUNE1/DDP cells were transfected with
si-NC, si-HOXA11-AS or si-HOXA11-AS + PBX3. RT-qPCR revealed that
PBX3 overexpression plasmid transfection increased PBX3 expression
in 5–8F/DDP and SUNE1/DDP cells compared with the cells transfected
with NC (pcDNA3.1) (Fig. 5G), and
the overexpression of PBX3 abolished the effects of HOXA11-AS
silencing on the DDP sensitivity of 5–8F/DDP and SUNE1/DDP cells
(Fig. 5H). Furthermore, the
overexpression of PBX3 reversed the inductive effects of
si-HOXA11-AS on cell apoptosis (Fig.
5I). The interference of HOXA11-AS increased the protein
expression levels of Bax and inhibited Bcl-2 expression, which was
partially rescued by the overexpression of PBX3 (Fig. 5J). In summary, these findings
indicated that HOXA11-AS promoted DDP resistance in NPC cells by
regulating the miR-98/PBX3 axis.
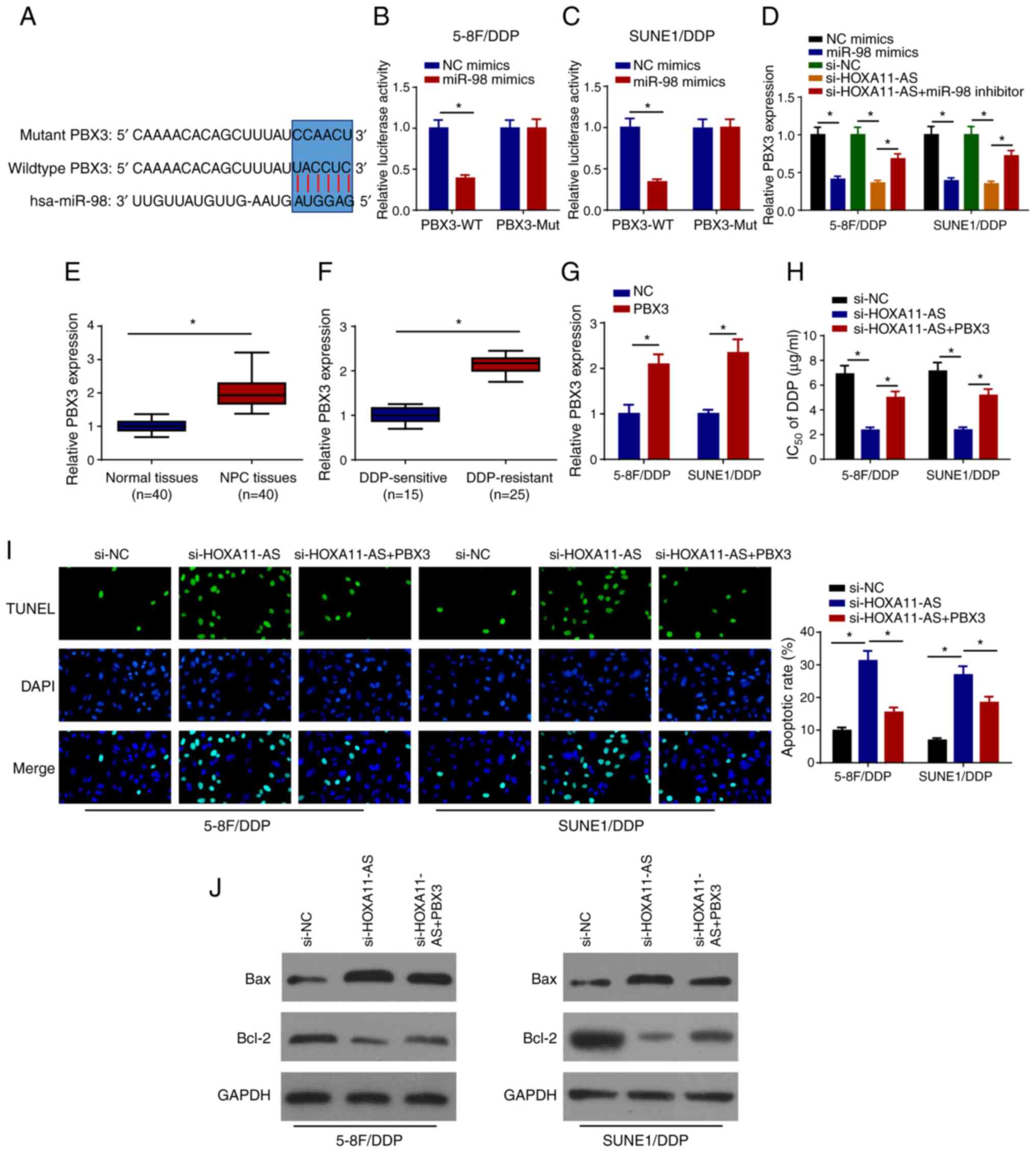 | Figure 5.HOXA11-AS confers DDP resistance via
upregulation of PBX3 expression in NPC cells by sponging miR-98.
(A) Binding sites between PBX3 and miR-98 were predicted.
Luciferase activities of WT or Mut PBX3 in (B) 5–8F/DDP and (C)
SUNE1/DDP cells following transfection of NC mimics or miR-98
mimics were detected using a luciferase reporter assay. (D) RT-qPCR
was used to detect PBX3 expression in 5–8F/DDP and SUNE1/DDP cells
transfected with NC mimics, miR-98 mimics, si-NC, si-HOXA11-AS,
si-HOXA11-AS + miR-98 inhibitor. (E) RT-qPCR was used to detect the
expression levels of PBX3 in NPC and normal tissues. (F) PBX3
expression in DDP-sensitive or DDP-resistant NPC tissues was
detected using RT-qPCR. (G) Expression levels of PBX3 were detected
by RT-qPCR in 5–8F/DDP and SUNE1/DDP cells transfected with NC or
PBX3. (H) IC50 of DDP was measured in 5–8F/DDP and
SUNE1/DDP cells transfected with si-NC, si-HOXA11-AS or
si-HOXA11-AS + PBX3 using a Cell Counting Kit-8 assay. (I) Cell
apoptosis was measured by TUNEL staining (magnification, ×200) in
5–8F/DDP and SUNE1/DDP cells transfected with si-NC, si-HOXA11-AS
or si-HOXA11-AS + PBX3. (J) Protein expression levels of Bax and
Bcl-2 in 5–8F/DDP and SUNE1/DDP cells transfected with si-NC,
si-HOXA11-AS or si-HOXA11-AS + PBX3 were assessed by western blot
analysis. Data are presented as the mean ± SD. *P<0.05. DDP,
cisplatin; HOXA11-AS, homeobox A11-antisense RNA; miR-98,
microRNA-98; Mut, mutant; NC, negative control; NPC, nasopharyngeal
carcinoma; PBX3, pre-B-cell leukemia homeobox 3; RT-qPCR, reverse
transcription-quantitative PCR; si, small interfering RNA; WT,
wild-type. |
 | Table I.Association between
clinicopathological characteristics and PBX3 expression in
nasopharyngeal carcinoma. |
Table I.
Association between
clinicopathological characteristics and PBX3 expression in
nasopharyngeal carcinoma.
| Variable | No. of
patients | Low PBX3
expression, n | High PBX3
expression, n | P-value |
|---|
| Age, years |
|
|
| 0.934 |
|
<55 | 19 | 10 | 9 |
|
|
≥55 | 21 | 11 | 10 |
|
| Sex |
|
|
| 0.973 |
|
Male | 22 | 12 | 10 |
|
|
Female | 18 | 9 | 9 |
|
| Lymph node
metastasis |
|
|
| 0.001 |
| No | 23 | 16 | 7 |
|
|
Yes | 17 | 5 | 12 |
|
| Distant
metastasis |
|
|
| 0.012 |
| No | 22 | 14 | 8 |
|
|
Yes | 18 | 7 | 11 |
|
| Clinical stage |
|
|
| 0.001 |
|
I/II | 21 | 15 | 6 |
|
|
III/IV | 19 | 6 | 13 |
|
Discussion
Chemoresistance has been considered to be a barrier
to the effective treatment of NPC (25,26).
Therefore, it is necessary to clarify the underlying mechanism of
chemoresistance in NPC. In the present study, knockdown of
HOXA11-AS enhanced the DDP sensitivity of NPC cells by sponging
miR-98 and suppressing PBX3. Preliminary evidence suggested that
the HOXA11-AS/miR-98/PBX3 axis was involved in the regulation of
DDP resistance in NPC.
The expression levels of HOXA11-AS have been
investigated in multiple studies (27–29).
According to previous studies, HOXA11-AS has been identified as an
oncogene in several types of cancer, including gastric cancer
(30), glioma (31) and hepatocellular carcinoma (32). In addition, HOXA11-AS promotes the
DDP resistance of lung cancer via regulation of the miR-454/STAT3
axis (33). Similarly, HOXA11-AS
contributes to the DDP resistance and proliferation of oral
squamous cell carcinoma by inhibiting miR-214 expression (34). In the present study, HOXA11-AS
expression was increased in DDP-resistant NPC tissues and cells.
Furthermore, knockdown of HOXA11-AS promoted the sensitivity and
apoptosis of DDP-resistant NPC cells. These data revealed that
HOXA11-AS conferred DDP resistance in NPC cells.
Accumulating evidence has demonstrated that lncRNAs
act as competing endogenous RNAs by sponging miRNAs to participate
in tumor progression and drug resistance (35). In the present study, HOXA11-AS served
as a molecular sponge for miR-98. miR-98 has been demonstrated to
be a cancer-associated miRNA, with abnormal expression in multiple
cancer types (36–38). For instance, miR-98 suppresses the
migration of breast cancer by targeting E2F transcription factor 5
(39). Upregulation of miR-98
expression suppresses the development of non-small cell lung cancer
by targeting MAPK6 (40). In
addition, miR-98 sensitizes DDP-resistant lung adenocarcinoma cells
by regulating high mobility group AT-hook 2 (41). miR-98 overexpression impedes
proliferation and enhances the sensitivity of leukemia cells to
Adriamycin (42). The present study
demonstrated that miR-98 was a direct target of HOXA11-AS.
Overexpression of HOXA11-AS suppressed miR-98 expression, whereas
knockdown enhanced miR-98 expression. Furthermore,
HOXA11-AS-knockdown enhanced the DDP sensitivity of NPC cells by
sponging miR-98.
PBX3 is a member of the PBX family, which is
implicated in the development of various cancer types (43). For example, the overexpression of
PBX3 accelerates the development of gastric cancer (44). Knockdown of PBX3 inhibits the
cervical cancer cell epithelial-mesenchymal transition process
(45). To the best of our knowledge,
the present study provided the first preliminary evidence regarding
the interaction between PBX3 (target gene) and miR-98. PBX3
expression was markedly increased in NPC tissues, notably in
DDP-resistant NPC tissues. miR-98 overexpression and
HOXA11-AS-knockdown markedly decreased the expression levels of
PBX3, whereas miR-98 inhibition partly abolished the repressive
effects of HOXA11-AS-knockdown on PBX3 expression. In addition,
overexpression of PBX3 weakened the increased DDP sensitivity of
5–8F/DDP and SUNE1/DDP cells caused by HOXA11-AS silencing.
In conclusion, the present study demonstrated that
HOXA11-AS conferred DDP resistance of NPC via the miR-98/PBX3 axis.
These findings suggested that HOXA11-AS may be a valuable target
for overcoming DDP resistance in patients with NPC. In future
studies, the mechanism by which the HOXA11-AS/miR-98/PBX3 axis
increases DDP resistance of NPC should be explored.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HNL, YZ and QW designed the present study. JH and SY
performed the experiments. HBL and HNL analyzed the data and
prepared the figures. HNL and YZ drafted the initial manuscript. QW
reviewed and revised the manuscript. The authenticity of all the
raw data must have been assessed by HNL and QW. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Shanghai Ninth People's Hospital (Shanghai, China).
All participants provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang W, Zhang Y and Xi S: Upregulation of
lncRNA HAGLROS enhances the development of nasopharyngeal carcinoma
via modulating miR-100/ATG14 axis-mediated PI3K/AKT/mTOR signals.
Artif Cells Nanomed Biotechnol. 47:3043–3052. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fan C, Tang Y, Wang J, Wang Y, Xiong F,
Zhang S, Li X, Xiang B, Wu X, Guo C, et al: Long non-coding RNA
LOC284454 promotes migration and invasion of nasopharyngeal
carcinoma via modulating the Rho/Rac signaling pathway.
Carcinogenesis. 40:380–391. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kang M, Zhou P, Li G, Yan H, Feng G, Liu
M, Zhu J and Wang R: Validation of the 8th edition of the UICC/AJCC
staging system for nasopharyngeal carcinoma treated with
intensity-modulated radiotherapy. Oncotarget. 8:70586–70594. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wei F, Wu Y, Tang L, He Y, Shi L, Xiong F,
Gong Z, Guo C, Li X, Liao Q, et al: BPIFB1 (LPLUNC1) inhibits
migration and invasion of nasopharyngeal carcinoma by interacting
with VTN and VIM. Br J Cancer. 118:233–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kamran SC, Riaz N and Lee N:
Nasopharyngeal carcinoma. Surg Oncol Clin North Am. 24:547–561.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim ED and Sung S: Long noncoding RNA:
Unveiling hidden layer of gene regulatory networks. Trends Plant
Sci. 17:16–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kong X, Duan Y, Sang Y, Li Y, Zhang H,
Liang Y, Liu Y, Zhang N and Yang Q: LncRNA-CDC6 promotes breast
cancer progression and function as ceRNA to target CDC6 by sponging
microRNA-215. J Cell Physiol. 234:9105–9117. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang J, Su Z, Lu S, Fu W, Liu Z, Jiang X
and Tai S: LncRNA HOXA-AS2 and its molecular mechanisms in human
cancer. Clin Chim Acta. 485:229–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chan JJ and Tay Y: Noncoding RNA:RNA
regulatory networks in cancer. Int J Mol Sci. 19:13102018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Z, Feng L, Liu P and Duan W: ANRIL
promotes chemoresistance via disturbing expression of ABCC1 by
regulating the expression of Let-7a in colorectal cancer. Biosci
Rep. 38:BSR201806202018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cao C, Zhou S and Hu J: Long noncoding RNA
MAGI2-AS3/miR-218-5p/GDPD5/SEC61A1 axis drives cellular
proliferation and migration and confers cisplatin resistance in
nasopharyngeal carcinoma. Int Forum Allergy Rhinol. 10:1012–1023.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cui Z, Pu T, Zhang Y, Wang J and Zhao Y:
Long non-coding RNA LINC00346 contributes to cisplatin resistance
in nasopharyngeal carcinoma by repressing miR-342-5p. Open Biol.
10:1902862020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Huang J, Yu S and Lou Z: Long
non-coding RNA DLEU1 Up-regulates BIRC6 expression by competitively
sponging miR-381-3p to promote cisplatin resistance in
nasopharyngeal carcinoma. Onco Targets Ther. 13:2037–2045. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Q, Zhang W and Hao S: LncRNA CCAT1
modulates the sensitivity of paclitaxel in nasopharynx cancers
cells via miR-181a/CPEB2 axis. Cell Cycle. 16:795–801. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lenkala D, LaCroix B, Gamazon ER, Geeleher
P, Im HK and Huang RS: The impact of microRNA expression on
cellular proliferation. Hum Genet. 133:931–938. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tufekci KU, Meuwissen RL and Genc S: The
role of microRNAs in biological processes. Methods Mol Biol.
1107:15–31. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mihanfar A, Fattahi A and Nejabati HR:
MicroRNA-mediated drug resistance in ovarian cancer. J Cell
Physiol. 234:3180–3191. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
van Jaarsveld MT, Helleman J, Berns EM and
Wiemer EA: MicroRNAs in ovarian cancer biology and therapy
resistance. Int J Biochem Cell Biol. 42:1282–1290. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao Y, Wang P and Wu Q: miR-1278
sensitizes nasopharyngeal carcinoma cells to cisplatin and
suppresses autophagy via targeting ATG2B. Mol Cell Probes.
53:1015972020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuan TZ, Zhang HH, Lin XL, Yu JX, Yang QX,
Liang Y, Deng J, Huang LJ and Zhang XP: microRNA-125b reverses the
multidrug resistance of nasopharyngeal carcinoma cells via
targeting of Bcl-2. Mol Med Rep. 15:2223–2228. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carey LA, Metzger R, Dees EC, Collichio F,
Sartor CI, Ollila DW, Klauber-DeMore N, Halle J, Sawyer L, Moore DT
and Graham ML: American Joint Committee on Cancer
tumor-node-metastasis stage after neoadjuvant chemotherapy and
breast cancer outcome. J Natl Cancer Inst. 97:1137–1142. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang W, Lou W, Ding B, Yang B, Lu H, Kong
Q and Fan W: A novel mRNA-miRNA-lncRNA competing endogenous RNA
triple sub-network associated with prognosis of pancreatic cancer.
Aging. 11:2610–2627. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo H, Xu C, Le W, Ge B and Wang T: lncRNA
CASC11 promotes cancer cell proliferation in bladder cancer through
miRNA-150. J Cell Biochem. 120:13487–13493. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang J, Xie T, Zhong X, Jiang HL, Li R,
Wang BY, Huang XT, Cen BH and Yuan YW: Melatonin reverses
nasopharyngeal carcinoma cisplatin chemoresistance by inhibiting
the Wnt/beta-catenin signaling pathway. Aging (Albany NY).
12:5423–5438. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang R, Li SW, Liu L, Yang J, Huang G and
Sang Y: TRIM11 facilitates chemoresistance in nasopharyngeal
carcinoma by activating the beta-catenin/ABCC9 axis via
p62-selective autophagic degradation of Daple. Oncogenesis.
9:452020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xue JY, Huang C, Wang W, Li HB, Sun M and
Xie M: HOXA11-AS: A novel regulator in human cancer proliferation
and metastasis. Onco Targets Ther. 11:4387–4393. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu J, Zhang Y, You Q, Fu H, Zhao X, Lu K,
Yan R and Yang D: LncRNA PTCSC3 alleviates the postoperative
distant recurrence of gastric cancer by suppression of lncRNA
HOXA11-AS. Cancer Manag Res. 12:2623–2629. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Niu X, Yang B, Liu F and Fang Q: LncRNA
HOXA11-AS promotes OSCC progression by sponging miR-98-5p to
upregulate YBX2 expression. Biomed Pharmacother. 121:1096232020.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sun M, Nie F, Wang Y, Zhang Z, Hou J, He
D, Xie M, Xu L, De W, Wang Z and Wang J: LncRNA HOXA11-AS promotes
proliferation and invasion of gastric cancer by scaffolding the
chromatin modification factors PRC2, LSD1, and DNMT1. Cancer Res.
76:6299–6310. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu CH, Xiao LM, Liu Y, Chen LK, Zheng SY,
Zeng EM and Li DH: The lncRNA HOXA11-AS promotes glioma cell growth
and metastasis by targeting miR-130a-5p/HMGB2. Eur Rev Med
Pharmacol Sci. 23:241–252. 2019.PubMed/NCBI
|
|
32
|
Zhan M, He K, Xiao J, Liu F, Wang H, Xia
Z, Duan X, Huang R, Li Y, He X, et al: LncRNA HOXA11-AS promotes
hepatocellular carcinoma progression by repressing miR-214-3p. J
Cell Mol Med. 22:3758–3767. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao X, Li X, Zhou L, Ni J, Yan W, Ma R,
Wu J, Feng J and Chen P: LncRNA HOXA11-AS drives cisplatin
resistance of human LUAD cells via modulating miR-454-3p/Stat3.
Cancer Sci. 109:3068–3079. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang X, Li H and Shi J: LncRNA HOXA11-AS
promotes proliferation and cisplatin resistance of oral squamous
cell carcinoma by suppression of miR-214-3p expression. Biomed Res
Int. 2019:86451532019.PubMed/NCBI
|
|
35
|
Xie C, Chen B, Wu B, Guo J and Cao Y:
LncRNA TUG1 promotes cell proliferation and suppresses apoptosis in
osteosarcoma by regulating miR-212-3p/FOXA1 axis. Biomed
Pharmacother. 97:1645–1653. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luo H, Yang L, Liu C, Wang X, Dong Q, Liu
L and Wei Q: TMPO-AS1/miR-98-5p/EBF1 feedback loop contributes to
the progression of bladder cancer. Int J Biochem Cell Biol.
122:1057022020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yahya SM and Yahya SM: The effect of
miR-98 and miR-214 on apoptotic and angiogenic pathways in
hepatocellular carcinoma HepG2 cells. Indian J Clin Biochem.
35:353–358. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Qiu K, Xie Q, Jiang S and Lin T: miR-98-5p
promotes apoptosis and inhibits migration and cell growth in
papillary thyroid carcinoma through Bax/Caspase-3 by HMGA2. J Clin
Lab Anal. 34:e230442020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cai C, Huo Q, Wang X, Chen B and Yang Q:
SNHG16 contributes to breast cancer cell migration by competitively
binding miR-98 with E2F5. Biochem Biophys Res Commun. 485:272–278.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu F, Mo Q, Wan X, Dan J and Hu H:
NEAT1/hsa-mir-98-5p/MAPK6 axis is involved in non-small-cell lung
cancer development. J Cell Biochem. 120:2836–2846. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xiang Q, Tang H, Yu J, Yin J, Yang X and
Lei X: MicroRNA-98 sensitizes cisplatin-resistant human lung
adenocarcinoma cells by up-regulation of HMGA2. Pharmazie.
68:274–281. 2013.PubMed/NCBI
|
|
42
|
Huang Y, Hong X, Hu J and Lu Q: Targeted
regulation of MiR-98 on E2F1 increases chemosensitivity of leukemia
cells K562/A02. Onco Targets Ther. 10:3233–3239. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Guo H, Chu Y, Wang L, Chen X, Chen Y,
Cheng H, Zhang L, Zhou Y, Yang FC, Cheng T, et al: PBX3 is
essential for leukemia stem cell maintenance in MLL-rearranged
leukemia. Int J Cancer. 141:324–335. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang S, Li C, Wang W and Xing C: PBX3
promotes gastric cancer invasion and metastasis by inducing
epithelial-mesenchymal transition. Oncol Lett. 12:3485–3491. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li H, Wang J, Xu F, Wang L, Sun G, Wang J
and Yang Y: By downregulating PBX3, miR-526b suppresses the
epithelial-mesenchymal transition process in cervical cancer cells.
Future Oncol. 15:1577–1591. 2019. View Article : Google Scholar : PubMed/NCBI
|