Introduction
Colorectal cancer (CRC) is one of the most common
types of cancer, with 1.8 million cases and 862,000 deaths
worldwide, only during 2018 (1). In
total, ~2/3 of CRC cases are associated with colon cancer (CC) and
the rest with rectal cancer (RC). In addition, CRC is the third
most commonly occurring type of cancer in men and the second in
women in the world (2). The
incidence and mortality rates of CRC are increasing in younger
individuals for whom screening is limited and key symptoms may be
not obvious. The aforementioned finding is very important since for
the majority of patients with cancer, the 1- and 5-year survival
rate is much higher when cancer is diagnosed at the early stages of
the disease (3). Mexico is one of
the principal countries with the highest incidence of CRC in young
individuals, with a rate of 22.8% among patients <40 years old
(4,5).
The development of CRC is a multifactorial and
multistep process involving different genomic and epigenetic
modifications. The majority of CRC cases are sporadic and develop
slowly for over a decade through the adenoma-carcinoma sequence
(6). Although substantial progress
has been made in the treatment and understanding of the molecular
mechanisms involved in CRC in recent years, the overall survival
(OS) rate remains relatively low compared with other types of
cancer (7,8).
Emerging evidence has suggested that the Wnt pathway
is involved in the maintenance of the self-renewal capacity of stem
cells of the gastrointestinal tract (9). Paneth cells are a highly specialized
epithelial cell type located at the crypts, which serve a critical
role in controlling intestinal stem cell niche, and supporting
crypt base columnar cells and the regeneration of absorptive and
secretory cell types (10,11). SRY-related high-mobility group box 9
(SOX9) is a member of the SOX family of transcription factors, and
is known to act as a regulator in a variety of developmental
processes such as male sex determination, chondrogenesis,
neurogenesis and neural crest development (12). In the healthy colon, SOX9 is
expressed in stem/progenitor and Paneth cells by the
transcriptional effector of the Wnt pathway, namely the
β-catenin/transcription factor 4 (TCF4) complex (13). Mutations in important components of
the Wnt pathway, such as the tumor suppressor adenomatous polyposis
coli (APC) and the multifunctional β-catenin gene, have been
reported in the majority of CC cases (14). These mutations result in the
stabilization of β-catenin that constantly interacts with TCF4,
thus leading to constitutive transcription of TCF4-target genes,
such as SOX9, eventually promoting the development of CC through
transformation of the colonic epithelium (15). It has been reported that mutations in
the SOX9 gene, such as recurrent distal truncating and frameshift
or nonsense mutations, result in SOX9 overexpression in 11% of CC
cases. In addition, SOX9 mutations may coexist with KRAS mutations
and wild-type tumor protein 53 status (16).
Despite the importance of SOX9 in maintaining colon
homeostasis and stemness properties within the crypts (13), it has been ambiguously reported as
both an oncogene and tumor suppressor gene. As an oncogene,
overexpression of SOX9 in vitro and in vivo has been
associated with different pro-oncogenic properties in CRC, such as
promotion of proliferation, inhibition of senescence, and
collaboration with other oncogenes in cancer development (17). Interestingly, a study suggested that
SOX9 could serve as a marker of CRC progression and/or
differentiation, since it could negatively regulate the gene
expression of CDX2 and mucin 2, oligomeric mucus/gel-forming via
β-catenin (18). Moreover, a
previous study has suggested that the increased levels of SOX9 are
strongly associated with poor prognosis in CRC (19). SOX9 overexpression also promoted
S100P upregulation, which in turn enhanced the invasiveness and
metastasis of CC cells via activating the receptor for advanced
glycation end products/ERK signaling and promoting
epithelial-mesenchymal transition (EMT) (20). In this regard, SOX9 could regulate
hypoxia-induced ubiquitin-specific protease 47 upregulation to
promote EMT and metastasis of CRC cells (21). Additionally, another study
demonstrated that the increased secretion of TGF-β by tumor
microenvironment-associated macrophages upregulated SOX9 expression
and facilitated EMT (22).
Furthermore, it has been reported that high levels
of SOX9 enhance the invasion potential of CRC cells by providing
stemness properties (23), and are
associated with decreased survival rate and adverse prognosis, thus
supporting its use as a potential diagnostic and prognostic marker
in CRC in the Spanish population, via deregulating the BMI1
oncogene (24).
As a tumor suppressor, SOX9 is associated with the
human carcinoembryonic antigen (CEA) gene, which induces apoptosis
(25) and the CEA-related cell
adhesion molecule 1 (26) in the
human CC cell line HT29-16E. Interestingly, low expression of SOX9
was associated with a higher relapse risk in stage II CRC cases in
a Denmark population (27).
CRC is classified based on three important points
combined into an overall stage, namely local invasion depth (T
stage), lymph node involvement (N stage) and presence of distant
metastases (M stage) (7). However,
CC shows high tumor heterogeneity among patients, even at the same
TNM stage. Nowadays, prognostic markers, such as hypoalbuminemia
are used to predict poor surgical outcomes of colon cancer;
hypoalbuminemia is a poor prognosis factor for long-term survival
of colon cancer after curative surgery (28) and albumin level <3.5 g/dl predict
a poor survival chance for patients with colorectal carcinoma
(29). For this reason, the
pathological approach should be complemented with molecular
classification using diagnostic and prognostic biomarkers to
provide the appropriate therapy (30). Although populations growing in
different environments show some similarities in the instauration
of CC, they also present with heterogeneity in the biomarkers
associated with the onset of the disease. Therefore, the present
study aimed to evaluate the clinical value of SOX9 expression in
CC.
Materials and methods
Patients and tumor tissue samples
An observational, descriptive, and retrospective
study was conducted between January 2012 and June 2017. A total of
97 adult patients (48 male and 49 female; mean age ± SD, 56.2±14.45
years; age range, 18–90 years) were diagnosed with early-stage I
and II colon cancer at the National Cancer Institute of Mexico.
Tissue samples were obtained from all patients during surgical
treatment and were then paraffin-embedded. The study protocol was
approved by the Institutional Review Board of the National Cancer
Institute of Mexico (permit no. Rev/42/16) and due to its
retrospective nature, patient informed consent was not needed. The
clinicopathological features of patients were recorded and
evaluated. According to Vera et al (31), minimal follow-up should be between
3–6 months during the first 5 years, thus, patients who were lost
to follow-up for >6 months were excluded. The median follow-up
period for all patients was 66 months.
Immunohistochemistry (IHC)
Paraffin-embedded tissues were cut into 3-µm thick
slices and the SOX9 expression levels were determined by IHC.
Briefly, tissue slices were rehydrated and antigenically
reactivated in citrate buffer (0.01 M citric acid, 0.01 M sodium
citrate) for 10 min at 95°C. Tissues were then washed twice with 1X
PBS. Subsequently, IHC was carried out using the Mouse and Rabbit
Specific HRP Detection IHC kit (cat. no. ab-93686; Abcam) following
the manufacturer's instructions. Tissue endogenous peroxidase was
blocked following slice incubation with hydrogen peroxide for 10
min at room temperature followed by washing twice with 1X PBS.
Non-specific antigenic sites were blocked by a protein blocking
solution for 10 min at room temperature, and the tissue samples
were washed once followed by incubation with a polyclonal rabbit
SOX9 antibody (cat. no. ab185966; Abcam; 1:100) diluted in 0.01%
Triton X-100 and 0.1% BSA overnight at 4°C. The slides were then
washed three times with 1X PBS, incubated with enhancer solution
for 10 min at room temperature, washed again once and incubated
with the rabbit secondary antibody (cat. no. ab185966; Abcam;
1:800) for 30 min at room temperature. Following incubation with
the secondary antibody, the tissues were washed, and incubated for
5 min at room temperature with the DAB Black kit (cat. no. BR140 H,
L; Biocare Medical, LLC). The slides were counterstained with eosin
for 5 sec and mounted using the Aqua Mounter Solution (cat. no.
BSB0091; Bio SB). Digital images of the tissue sections were
captured under a light microscope (magnification, ×400 using the
color AxioCam MRc5 camera (Zeiss). SOX9 expression was evaluated at
the invasive front of the tumor by a pathologist (LLS) who was
blinded to the clinical data. The percentage score of positive
tumor nuclei was defined as: 0 (0-5%), 1 (>5-25%), 2
(>25-50%), 3 (>50-75%) and 4 (>75%). Signal intensity was
scored as follows: 0 (negative), 1 (low signal), 2 (moderate
signal) and 3 (high signal). The overall score for SOX9 expression
was determined by multiplying the score of positive nuclei with
that for signal intensity (32).
SOX9 expression was considered as ‘low’ when the overall score was
0–4 and as ‘high’ when the overall score was 4–12.
Cell culture
The human CC HCT116 cell line and the human normal
colon cell line CCD-18co were purchased from the American Type
Culture Collection. HCT116 cells were cultured in DMEM/F12 (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin.
CCD-18co cells were maintained in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS, 1%
penicillin/streptomycin and 1% non-essential amino acids. Both cell
lines were incubated at 37°C in a humidified atmosphere containing
5% CO2.
SOX9 silencing using small interfering
RNAs (siRNAs)
A total of 3×105 HCT116 cells were seeded into
6-well plates and when confluence reached 70%, cells were
transfected with 30 nM Silencer Select siRNA (cat. no. s532658;
targeted exon 3, siRNA location 1243; Thermo Fisher Scientific,
Inc.) specific to SOX9 (HCT116-siSOX9) and a negative control
(HCT116-Scr, cat. no. 4404021), using the Lipofectamine®
RNAiMAX reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Transfection with a
scrambled sequence served as control in functional assays. The
efficiency of SOX9 silencing was verified 24 h post
transfection.
RNA purification and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from transfected cells using
the TRIzol® reagent (Thermo Fisher Scientific, Inc) and
transcribed into cDNA using the High-Capacity cDNA Reverse
Transcription kit (Thermo Fisher Scientific, Inc.), following the
manufacturer's protocol: 25°C for 10 min, 37°C for 2 h and 85°C for
5 min. qPCR was carried out using the TaqMan Gene Expression Assay
for SOX9 (cat. no. Hs00165814_m1; Applied Biosystems; Thermo Fisher
Scientific, Inc.; amplicon length 102, exon boundary 2–3) and
TaqMan Universal PCR Master mix (Thermo Fisher Scientific, Inc.).
The PCR reactions were incubated at 50°C for 2 min, 95°C 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. PCR
analysis was carried out in triplicate and gene expression was
analyzed using the 2−ΔΔCq method (33). ACTB (cat. no. Hs01060665_g1, amplicon
length 63, exon boundary 2–3) was used as the endogenous
control.
Immunofluorescence analysis
Immunofluorescence was performed 48 h
post-transfection as previously described (34). Briefly, cells were washed with
ice-cold PBS and fixed in 4% p-formaldehyde for 10 min. Then, the
samples were washed three times with PBS and permeabilized with
0.1% Triton X-100 for 10 min. Following washing and blocking with
22.52 mg/ml glycine and 1% BFS, samples were hybridized with rabbit
anti-human SOX9 antibody (dilution, 1:200; cat. no. ab185966;
Abcam) for 1 h at room temperature, washed again and incubated with
donkey anti-rabbit IgG Alexa 647 secondary antibody (dilution,
1:1,000; cat. no. ab150075; Abcam) for 1 h at room temperature.
Finally, the samples were washed and mounted using Fluoroshield
with DAPI (Sigma-Aldrich; Merck KGaA). Images were captured at a
magnification of ×40 under the Leica LSCM SP8 (Leica Microsystems,
Inc.) microscope.
Sphere-formation assay
HCT116 and HCT116-siSOX9 cells were seeded at a
density of 4×104 cells/well into Ultra-Low Attachment 6-well plates
(Costar; Corning Inc.). Following incubation for 72 h,
sphere-formation capacity was evaluated under an inverted light
microscope at a magnification of ×10.
Statistical analysis
For clinical and experimental data, categorical
variables were compared by χ2 test and Fisher's exact test was used
for relapsed patients, while continuous variables were analyzed
using t-test or ANOVA followed by Bonferroni's post hoc with the
GraphPad Prism 8.0 software (GraphPad Software, Inc.). For survival
analysis, the OS was defined as the interval between surgery and
the last visit or death. In addition, relapse-free survival (RFS)
was defined as the interval between surgery and the local or
distant relapse. The estimate curves for 3-year OS and RFS were
obtained by the Kaplan-Meier method and the differences between
groups were determined by log-rank test. Statistical analyses were
performed using the STATA v.12.0 software (Stata Corp.). P<0.05
was considered to indicate a statistically significant
difference.
Results
SOX9 expression is associated with
clinical stage, tumor size and tumor location in CC
SOX9 has been identified as an oncogene in several
types of cancer, and is therefore considered to be a possible
biomarker of aggressive disease (35). Herein, the clinical value of SOX9
expression was evaluated in CC samples. The clinical and
pathological features from the 97 patients recruited in this study
are summarized in Table I. The data
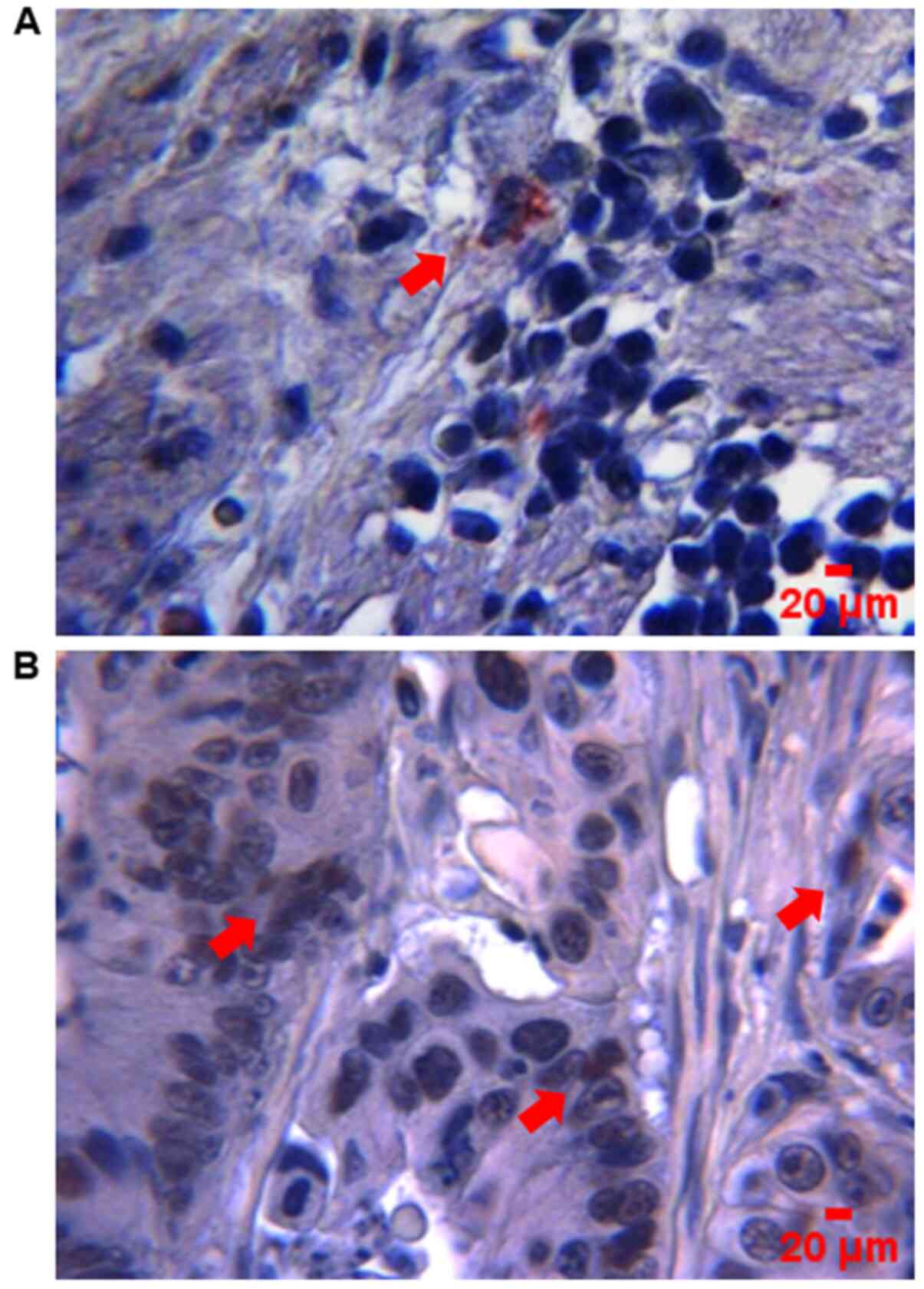
showed that SOX9 immunoreactivity was predominantly located in the
nuclei of tumor cells (Fig. 1). High
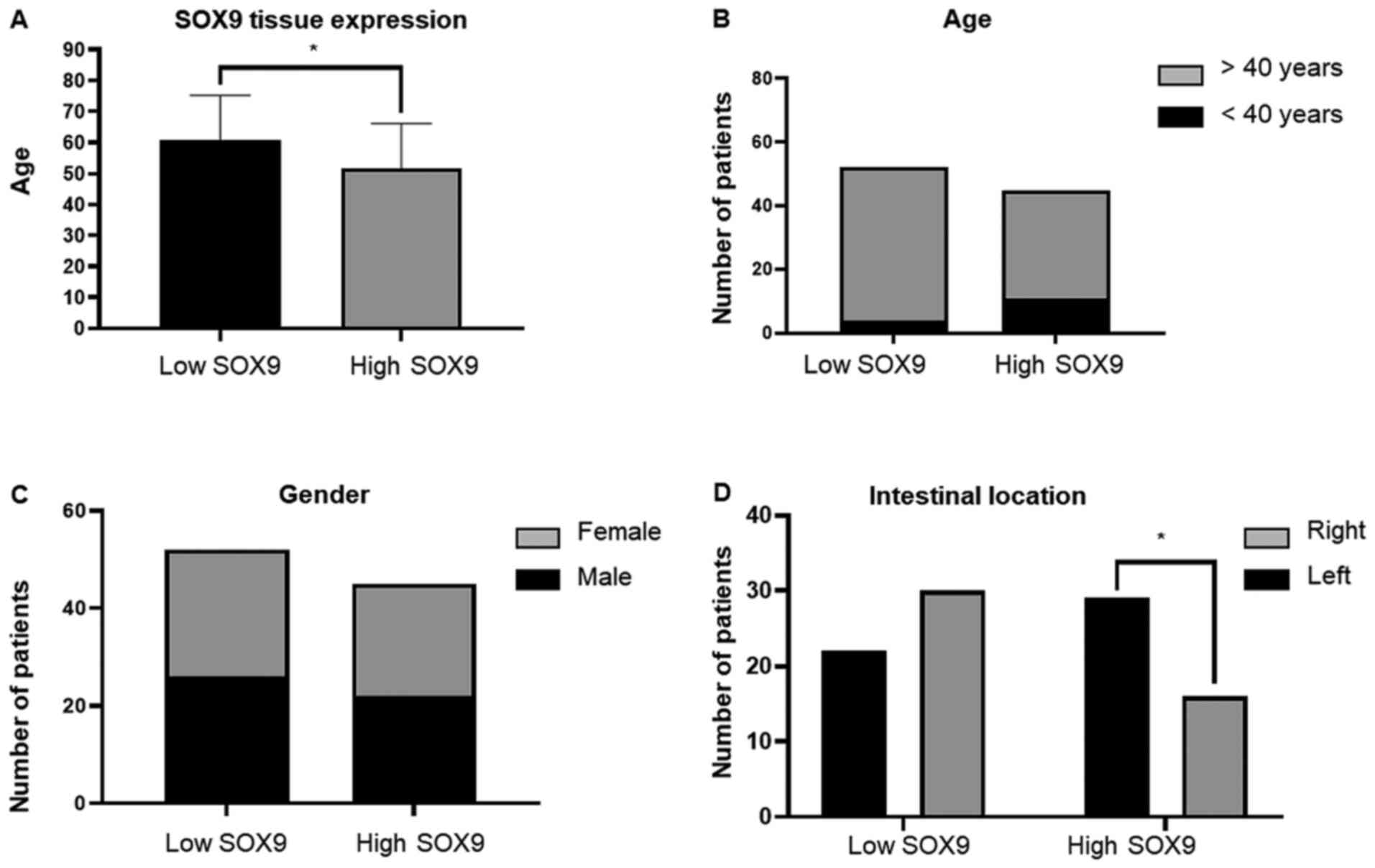
SOX9 expression was observed in 47% of tumors, while low one in 53%
(Fig. 2A). Patients <40 years
exhibited higher SOX9 expression compared with those >40 years
(P<0.046), suggesting that SOX9 could be important for the early
onset of CC (Fig. 2B). This age
cut-off was selected given the epidemiological data showed by Ahnen
et al (5), where more than
11% of colon cancer and 18% of rectal cancer have a young-onset,
occurring in individuals younger than 50 years and more
importantly, by the data showed by Rojas-Puente et al
(4), where CRC in Mexican young
people less than 40 years old, shows a disease characterized by
biological aggressiveness, with unique features such as
localization, adverse histologic factors, not associated with prior
or concomitant adenomas, advanced stage at diagnosis, and worse
prognosis. In addition, SOX9 expression was not associated with sex
(Fig. 2C); however, it was
positively associated with intestinal tumor location since its high
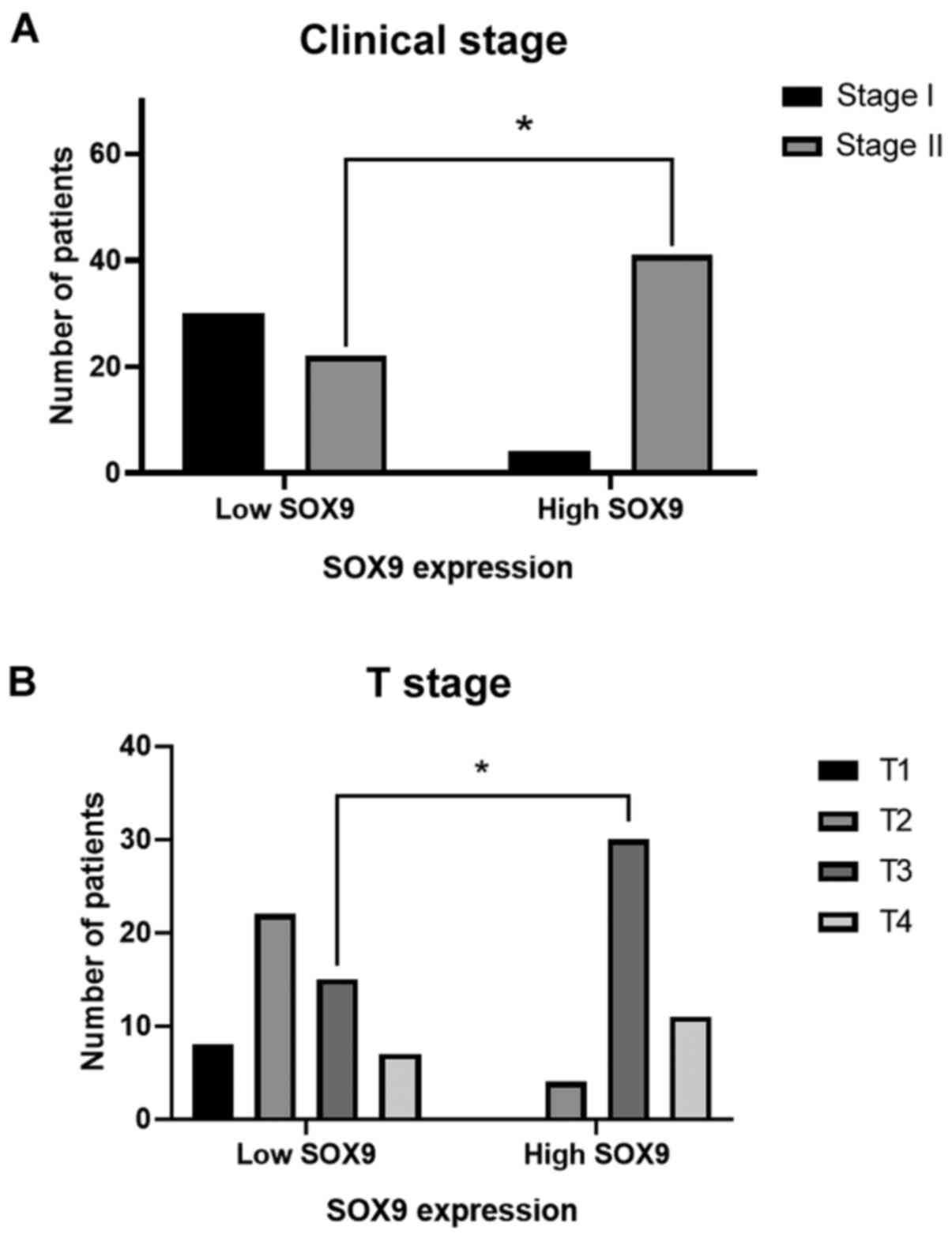
expression was predominant in the right side (P<0.048; Fig. 2D). Interestingly, χ2 test showed,
that in the high SOX9 expression group, 91% of patients
corresponded to the disease stage II (P<0.001) and 67% to the
advanced T3 stage (P<0.001). This finding indicated that SOX9
could be involved in tumor progression, size and extension
(Fig. 3).
 | Table I.Overall patient characteristics. |
Table I.
Overall patient characteristics.
|
Characteristics | Low SOX9 expression
n=52, n (%) | High SOX9
expression n=45, n (%) | P-value |
|---|
| Mean of age ± SD,
years | 60.7±14.5 | 51.7±14.4 | 0.003 |
| Age, years |
|
|
|
|
≤40 | 4 (8) | 11 (24) | 0.046 |
|
>40 | 48 (92) | 34 (76) |
|
| Sex |
|
| 0.913 |
|
Male | 26 (50) | 22 (49) |
|
|
Female | 26 (40) | 23 (51) |
|
| BMI |
|
| 0.34 |
|
Malnutrition | 1 (2) | 3 (6) |
|
|
Normal | 26 (50) | 27 (60) |
|
|
Overweight | 20 (38) | 13 (29) |
|
|
Obesity | 5(10) | 2 (5) |
|
| Albumin level |
|
| 0.065 |
|
<3.5 | 16 (30) | 23 (51) |
|
|
>3.5 | 36 (70) | 22 (49) |
|
| Clinical stage |
|
| <0.001 |
| I | 30 (58) | 4 (9) |
|
| II | 22 (42) | 41 (91) |
|
| T stage |
|
| <0.001 |
| T1 | 8 (15) | 0 (0) |
|
| T2 | 22 (42) | 4 (9) |
|
| T3 | 15 (29) | 30 (67) |
|
| T4 | 7 (14) | 11 (24) |
|
| Histological
type |
|
| 0.74 |
|
Mucinous | 8 (15) | 9 (20) |
|
| Not
mucinous | 44 (85) | 36 (80) |
|
| Histological
grade |
|
| 0.16 |
| 1 | 21(40) | 12 (27) |
|
| 2 | 20 (39) | 26 (58) |
|
| 3 | 11 (21) | 7 (15) |
|
| Tumor side |
|
| 0.048 |
|
Right | 22 (42) | 29 (64) |
|
|
Left | 30 (58) | 16 (36) |
|
| Intestinal
perforation |
|
| 0.81 |
|
Yes | 4 (8) | 2 (5) |
|
| No | 48 (92) | 43 (95) |
|
| Intestinal
occlusion |
|
| 0.98 |
|
Yes | 8 (15) | 7 (16) |
|
| No | 44 (85) | 38 (84) |
|
| Lymphovascular
invasion |
|
| 0.87 |
|
Yes | 11 (21) | 8 (18) |
|
| No | 41 (79) | 37 (82) |
|
| Perineural
invasion |
|
| 0.14 |
|
Yes | 1 (2) | 5 (11) |
|
| No | 51 (98) | 40 (89) |
|
SOX9 expression is associated with
local site recurrence
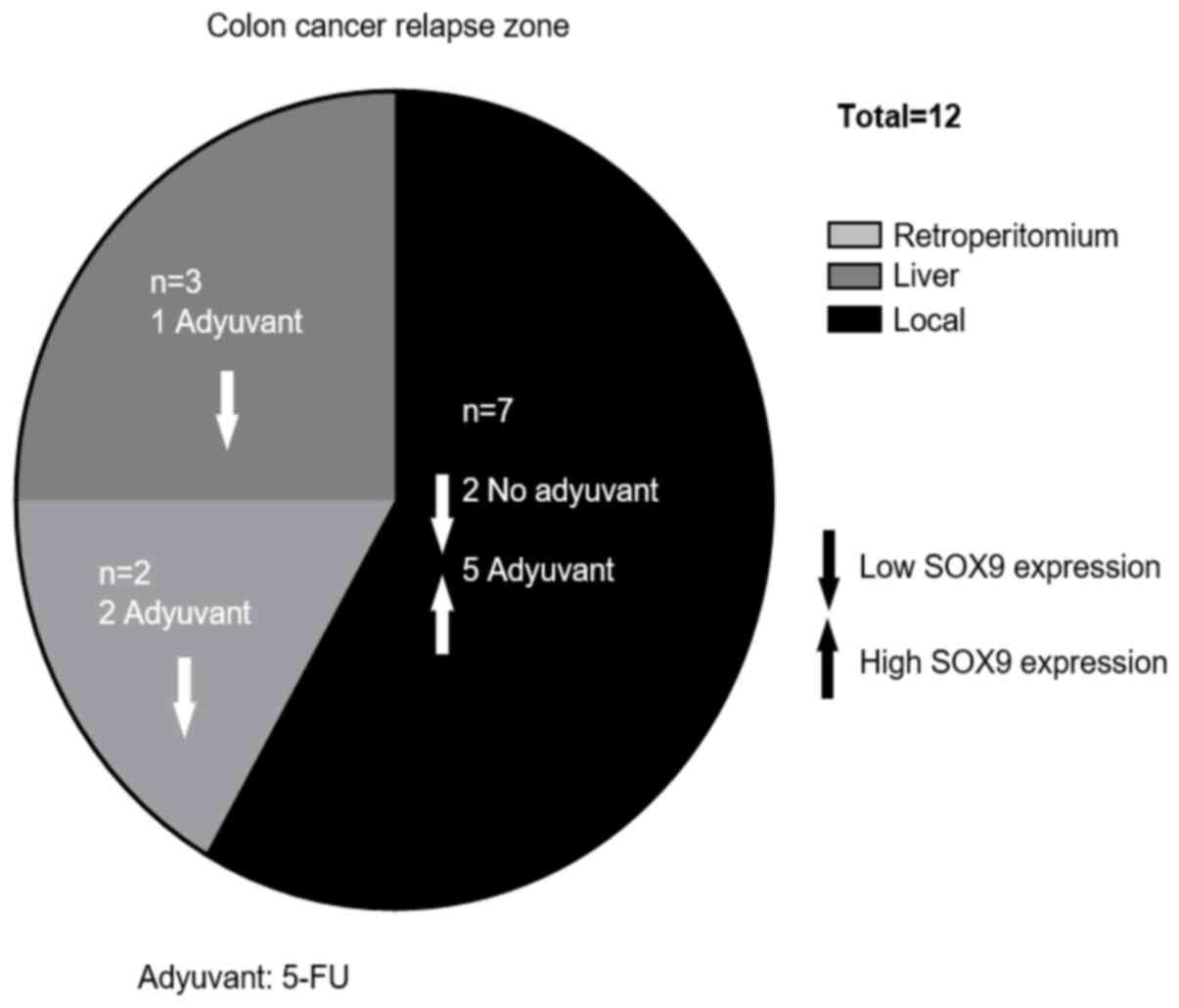
Relapse occurred in 12.4% of patients during
follow-up, while 3% of them died. The median RFS was 17 months
(range, 4–39 months). The characteristics of the relapsed patients
are illustrated in Table II.
Interestingly, in the case of relapsed patients that received
adjuvant therapy (66.7%), all with local relapse (62.5%) showed
tumors with high SOX9 expression levels; while, in those without
chemotherapy (33.3%), all with local (50%) and liver (50%) relapse
showed low SOX9 expression levels. However, due to the low number
of cases no statistically significant association was obtained by
Fisher's exact test (Table II;
Fig. 4). Additionally, SOX9
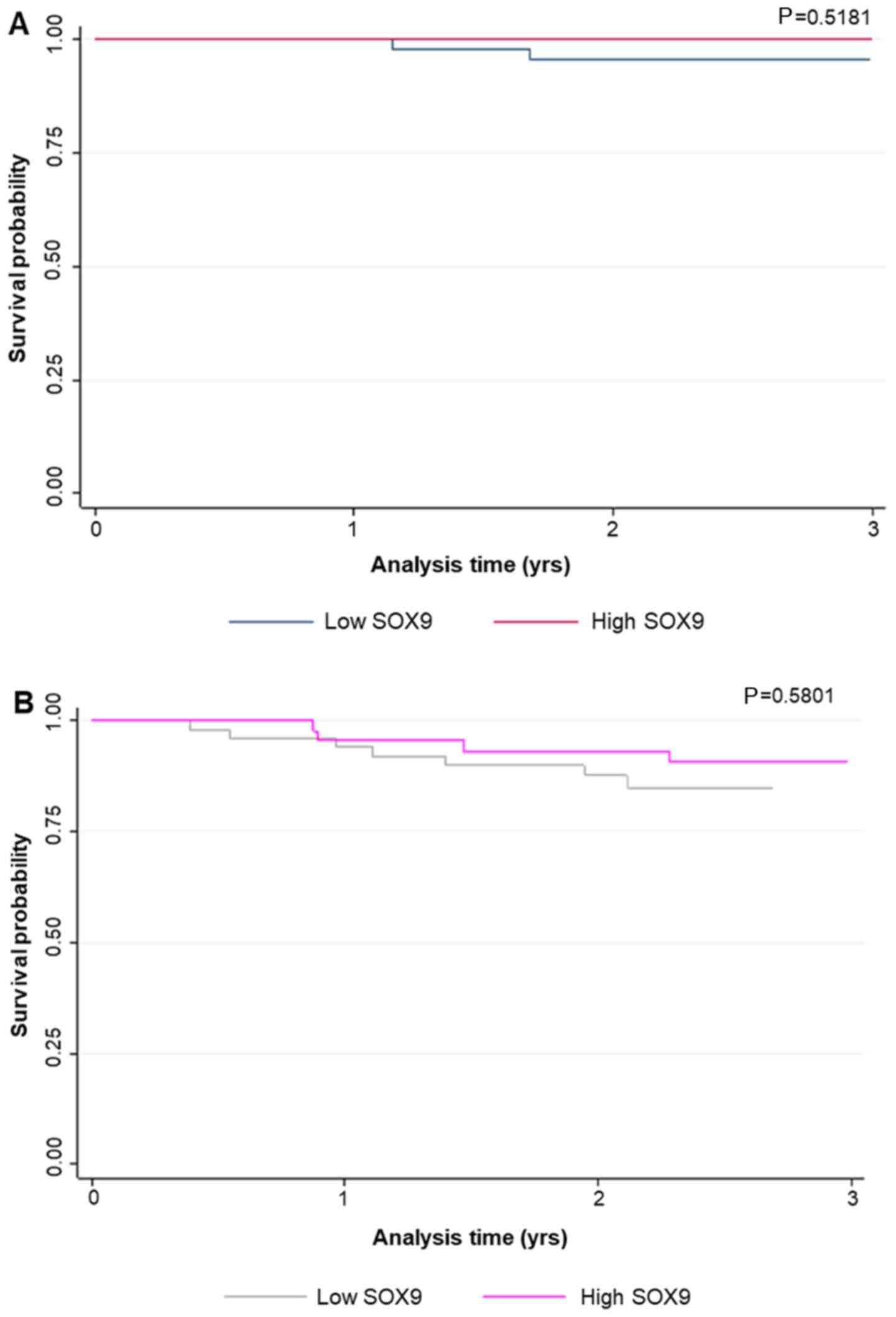
expression levels were not associated with survival probability,
neither in global patients (P<0.66) nor in relapsed ones
(P<0.21; Fig. 5).
| Table II.Characteristics of relapsed
patients. |
Table II.
Characteristics of relapsed
patients.
| Adjuvant | Relapse site | Age, years | Sex | Stage | Tumor location | RFS, months |
|---|
| Yes
(fluoropyrimidine) | Local | 66 | M | IIA | L | 29↑ |
| (N=8) | Local | 59 | F | IIA | L | 19↑ |
|
| Local | 59 | F | IIA | R | 8↑ |
|
| Local | 46 | M | IIB | R | 10↑ |
|
|
Retroperitoneum | 72 | F | IIB | L | 29↓ |
|
|
Retroperitoneum | 74 | M | IIA | L | 40↓ |
|
| Liver | 57 | F | IIB | R | 22↓ |
|
| Local | 64 | F | IIA | R | 11↑ |
| No (N=4) | Liver | 86 | F | I | L | 9↓ |
|
| Liver | 51 | F | IIB | R | 13↓ |
|
| Local | 45 | M | I | R | 16↓ |
|
| Local | 54 | F | I | L | 6↓ |
SOX9 silencing inhibits colonosphere
formation
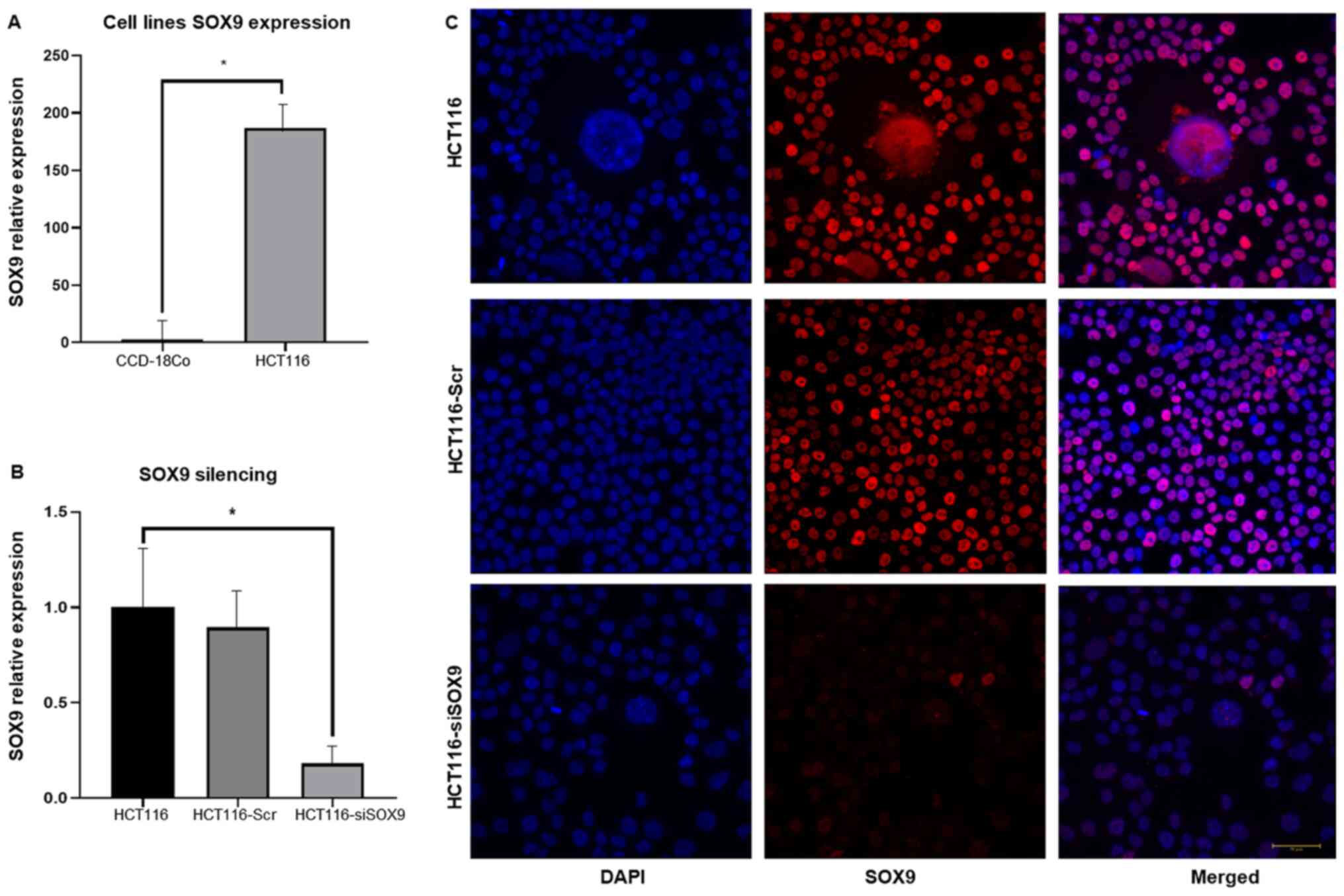
To investigate SOX9 effects in CC, its expression
was knocked down using specific siRNA targeting SOX9. The results
demonstrated that the HCT116 cell line overexpressed SOX9 (~180
fold) when compared with the non-tumorigenic colon cell line
CCD-18Co (P<0.001; Fig. 6A).
Following transfection of HCT116 cells with SOX9-siRNA, SOX9
expression was decreased by ~80% (P<0.002; Fig. 6B). Silencing efficiency was also
confirmed at the protein level by immunofluorescence assays. The
results showed that SOX9 was predominantly located in the cell
nuclei and its expression was significantly downregulated in cells
transfected with SOX9-siRNA (Fig.
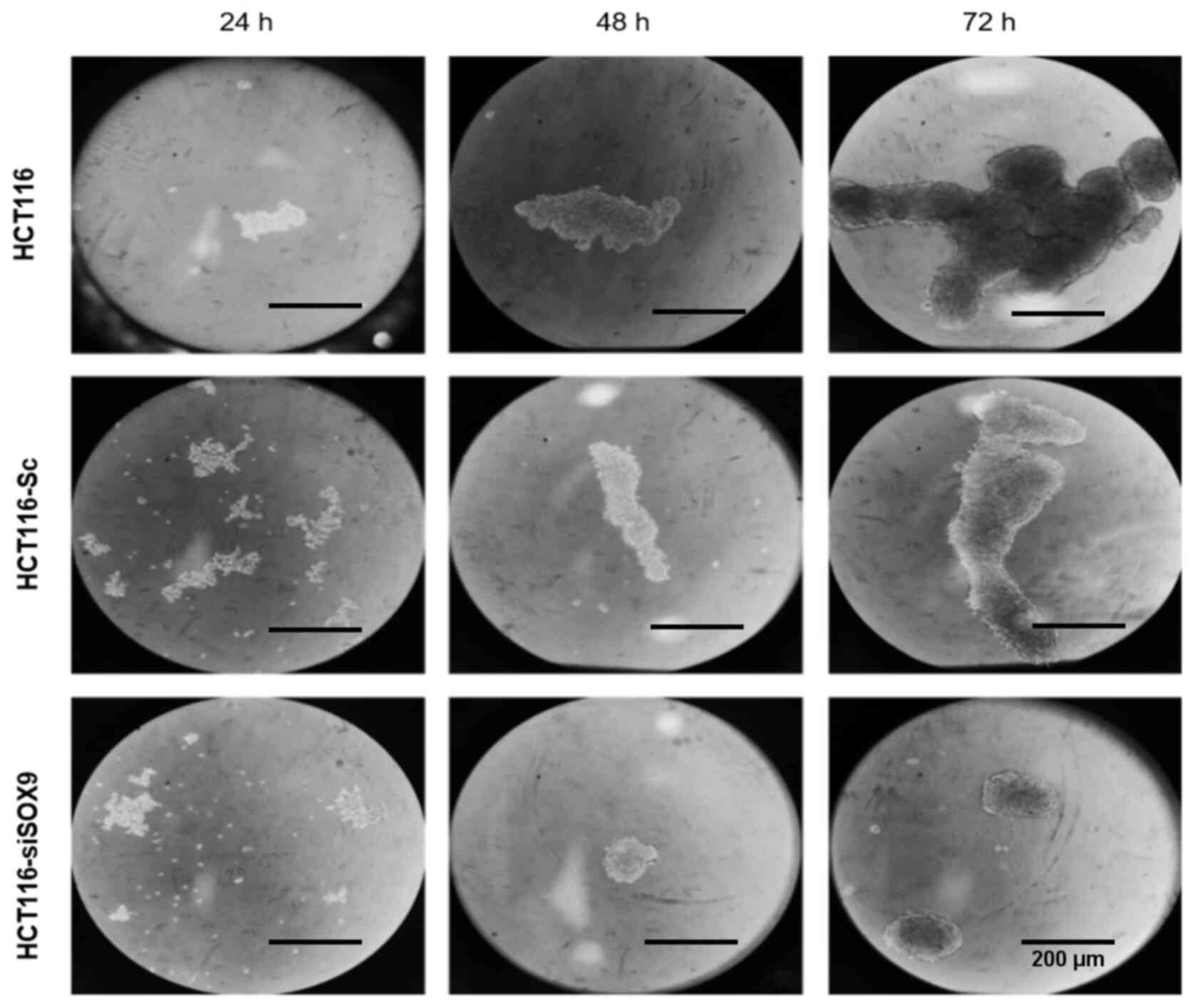
6C). In addition, HCT116 cells were able to generate solid and
well-defined colonospheres following incubation for 72 h in
ultra-low attachment plates. On the other hand, SOX9-deficient
HCT116 cells showed labile cellular aggregates and a significantly
reduced number of colonospheres (Fig.
7).
Discussion
The present study was the first to evaluate the
expression of SOX9 in Mexican patients diagnosed with early-stage
colon cancer. A limitation of the study was that samples were only
obtained from one hospital. Nevertheless, this national hospital
attends patients from all states of the center of the country.
Herein, 20% of all patients were <40 years old.
The significance of the current study is based on
the fact that in recent years CC has been diagnosed in younger
individuals, increasing by 2% each year (36). In this regard, it has been reported
that early-onset CRC can be caused by APC mutations in combination
with environmental factors that may have an impact on epigenetic
mechanisms modulating SOX9 expression in the intestinal epithelium
(37). Nonetheless, in other studies
on diverse types of cancer, including CC, it has been reported that
age is not associated with SOX9 expression (19,27,38,39).
Herein, in vitro experiments revealed that
the increased SOX9 expression could play an important role in
tumorigenesis and clinically associate mainly with advanced
T-stages in clinical-stage II patients, but not with relapse or
survival. These findings were inconsistent with those reported by
Maiken et al (27), where
high and low levels of SOX9 in primary stage II colon tumors could
predict low and high risk of relapse, respectively, suggesting that
SOX9 could be used as a biomarker for predicting risk of relapse.
Besides, Lü et al (19)
demonstrated that SOX9 immunoreactivity was the same among
different CRC stages. This finding could be attributed to the
heterogeneity of CRC tumors among patients. Nevertheless, given the
differences found in SOX9 expression in different studies,
Prévostel and Blache (40) proposed
a molecular model suggesting that in order to ensure a sustained
proliferation of intestine epithelial cells, a critical level of
active SOX9 should be maintained. In addition, according to this
model, CRC cells can implement several strategies in order the
levels of active SOX9 to be sufficient for cell proliferation,
namely heterozygous inactivating mutations, regulation of SOX9
expression, and regulation of SOX9 activity, mediated by SOX9
binding proteins such as transcription factors (40). Nonetheless, more experimental work
needs to be addressed to better understand the molecular mechanisms
of SOX9 in CC.
Emerging evidence has suggested that higher SOX9
expression is associated with advanced pathological grade and
different clinical stages in other types of cancer such as renal
cell carcinoma and bladder cancer (38). These findings could explain the local
recurrence mediated by SOX9 capability on inducing stemness state
and chemoresistance. However, longer follow-up and a higher sample
size are needed to further understand the effects of SOX9 in
CC.
Intriguingly, the current study demonstrated that
the expression levels of SOX9 were not associated with survival
probability, neither in global (P<0.66) nor in relapsed patients
(P<0.21). In this regard, there is evidence supporting that
cancer therapy is not equally effective against all types of
cancer, and cells that are not eliminated may contribute to
residual disease and may be the key drivers of cancer relapse
(41). Cancer stem cells (CSCs)
could be responsible of this behavior since they are usually
resistant to chemotherapy, eventually promoting recurrence
(42,43). Furthermore, the expression levels of
SOX9 are associated with stemness phenotype, since SOX9 levels are
sufficient to acquire and maintain colorectal-CSCs traits (23). In the present study, overexpressed
SOX9 was predominant in the right side. These right-sided tumors
(RSTs) tend to be advanced, bigger, and are often poorly
differentiated. In addition, RSTs are associated with mutations in
the DNA mismatch repair pathway, and do not respond well to
conventional therapies, whereas they benefit from immunotherapy
given their antigenic load (44,45).
Furthermore, it has been reported that SOX9
expression is associated with chemoresistance in gastric (46,47),
pancreatic (48) and CRC, while its
high expression in several solid tumors is associated with poor OS,
RFS and disease-free survival (17).
These results could suggest that SOX9 was not associated with
survival probability, at least in a small cohort of patients as
described here. However, it has been reported that higher levels of
SOX9 are associated with reduced survival probability in
hepatocellular carcinoma (49).
Additionally, the expression of SOX9 has been previously associated
with poor prognosis and RFS in different types of cancer (50). In terms of molecular
characterization, frequent mutations in AT-rich interaction domain
1A, APC membrane recruitment protein 1 and SOX9 genes have been
identified in CC and RC (51),
supporting the diversity of SOX9 behavior in CC among different
populations. These results indicated that SOX9 expression was not
relevant to survival probability, suggesting that a larger number
of patients should be analyzed to obtain a significant
correlation.
Herein, SOX9 silencing attenuated the colonosphere
formation ability of HCT116 cells, a hallmark of cancer stem cells.
According to Bahmad et al, these studies primarily relied on
two main criteria in their assessment: average volume of the
generated spheres (size) and the average number of sphere-forming
units (SFU) (52). Although our
study did not evaluate SFU, size of colonosphere represents the
proliferation of cells in the tumor mass, the capability of
adhesion, and the anoikis resistance (AR) (53,54). A
study demonstrated that SOX9 silencing resulted in AR loss in
HCT116 cells. Therefore, the AR tumor cells could detach from their
primary tumor and metastasize (52,55).
During tumor onset, normal cells acquire mutations that modify the
gene expression of specific genes, thus promoting a series of
molecular changes, which in turn stimulate clonal expansion and the
development of invasive properties of aberrant cells (56). It has been shown that the fusion of
apoptotic blebs, retention of full-length p70S6K, pH, and
lipid-dependent fusion between blebbishields to form spheres is a
key aspect for survival of CSCs (57). Also, important downstream genes of
the Wnt/β-catenin signaling, such as SOX9, are involved in several
crucial processes of cancer development, such as the long-term
self-renewal and tumor formation, during the early steps (58). Emerging evidence has supported the
inhibition of colonosphere formation in response to conditional
expression of wild type SOX9, which also attenuates the activity of
the Wnt/β-catenin pathway (59).
These results suggested that SOX9 could have a pivotal role in the
generation of CSCs via modulating other molecules in the form of
spheroids, as previously described in other types of cancer
(60,61). Nevertheless, the effect of SOX9
overexpression in SOX9-negative CRC cancer cells needs to be
examined as well as the expression of p70S6k as a marker of cell
transformation in colonospheres to better support this
hypothesis.
In conclusion, in the present study the expression
levels of SOX9 were determined in Mexican patients diagnosed with
early-stage CC and showed that SOX9 expression was associated with
younger CC cases with advanced T stages of clinical stage II, but
not with relapse or disease-free survival. However, further studies
with a larger sample size are needed to assess the response of
cells to the restoration of SOX9 expression to further understand
the underlying molecular mechanisms involved in CRC.
Acknowledgements
Not applicable.
Funding
The authors acknowledge Consejo Nacional de Ciencia
y Tecnología CONACYT (grant no. 290311) for funding.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ELV, MAF, MB and MPN conducted laboratory tests.
TGC, EFF, HAS, IVS and ERG diagnosed and treated the patients. MAM,
RRP and RCO collected the data and analyzed the results. LLS
analyzed SOX9 expression in patients' samples. MAM, CLC and RRP
conceived and designed the study. CLC and MAM confirmed the
authenticity of the data. CLC, RRP, MB, MAM and ERG wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study protocol was approved by the Institutional
Review Board of the National Cancer Institute of Mexico (permit no.
Rev/42/16) and due to its retrospective nature, patient informed
consent was not needed.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rawla P, Sunkara T and Barsouk A:
Epidemiology of colorectal cancer: Incidence, mortality, survival,
and risk factors. Prz Gastroenterol. 14:89–103. 2019.PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hawkes N: Cancer survival data emphasise
importance of early diagnosis. BMJ. 364:l4082019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rojas-Puente L, de la Garza-Salazar JG,
Calderillo-Ruiz G, Lino-Silva LS, Millan SV, Noveron NR,
Meneses-Garcia A, de la Vega HA and Ruiz Garcia EB: Increased
incidence of colorectal cancer in young people (less than 40 years
old) over the last ten years. J Cancerology. 1:16–22. 2014.
|
|
5
|
Ahnen DJ, Wade SW, Jones WF, Sifri R,
Mendoza Silveiras J, Greenamyer J, Guiffre S, Axilbund J, Spiegel A
and You YN: The increasing incidence of young-onset colorectal
cancer: A call to action. Mayo Clin Proc. 89:216–224. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mishra DK, Veena U, Kaliki S, Kethiri AR,
Sangwan VS, Ali MH, Naik MN and Singh V: Differential expression of
stem cell markers in ocular surface squamous neoplasia. PLoS One.
11:e01618002016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li W, Zhang G, Wang HL and Wang L:
Analysis of expression of cyclin E, p27kip1 and Ki67 protein in
colorectal cancer tissues and its value for diagnosis, treatment
and prognosis of disease. Eur Rev Med Pharmacol Sci. 20:4874–4879.
2016.PubMed/NCBI
|
|
9
|
Krausova M and Korinek V: Wnt signaling in
adult intestinal stem cells and cancer. Cell Signal. 26:570–579.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gassler N: Paneth cells in intestinal
physiology and pathophysiology. World J Gastrointest Pathophysiol.
8:150–160. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sailaja BS, He XC and Li L: The regulatory
niche of intestinal stem cells. J Physiol. 594:4827–4836. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jo A, Denduluri S, Zhang B, Wang Z, Yin L,
Yan Z, Kang R, Shi LL, Mok J, Lee MJ, et al: The versatile
functions of Sox9 in development, stem cells, and human diseases.
Genes Dis. 1:149–161. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bastide P, Darido C, Pannequin J, Kist R,
Robine S, Marty-Double C, Bibeau F, Scherer G, Joubert D, Hollande
F, et al: Sox9 regulates cell proliferation and is required for
Paneth cell differentiation in the intestinal epithelium. J Cell
Biol. 178:635–648. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Morin PJ, Sparks AB, Korinek V, Barker N,
Clevers H, Vogelstein B and Kinzler KW: Activation of
beta-catenin-Tcf signaling in colon cancer by mutations in
beta-catenin or APC. Science. 275:1787–1790. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Korinek V, Barker N, Morin PJ, van Wichen
D, de Weger R, Kinzler KW, Vogelstein B and Clevers H: Constitutive
transcriptional activation by a beta-catenin-Tcf complex in APC-/-
colon carcinoma. Science. 275:1784–1787. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Javier BM, Yaeger R, Wang L, Sanchez-Vega
F, Zehir A, Middha S, Sadowska J, Vakiani E, Shia J, Klimstra D, et
al: Recurrent, truncating SOX9 mutations are associated with SOX9
overexpression, KRAS mutation, and TP53 wild type status in
colorectal carcinoma. Oncotarget. 7:50875–50882. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Aguilar-Medina M, Avendaño-Félix M,
Lizárraga-Verdugo E, Bermúdez M, Romero-Quintana JG, Ramos-Payan R,
Ruíz-García E and López-Camarillo C: SOX9 stem-cell factor:
Clinical and functional relevance in cancer. J Oncol.
2019:67540402019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Blache P, van de Wetering M, Duluc I,
Domon C, Berta P, Freund JN, Clevers H and Jay P: SOX9 is an
intestine crypt transcription factor, is regulated by the Wnt
pathway, and represses the CDX2 and MUC2 genes. J Cell Biol.
166:37–47. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lü B, Fang Y, Xu J, Wang L, Xu F, Xu E,
Huang Q and Lai M: Analysis of SOX9 expression in colorectal
cancer. Am J Clin Pathol. 130:897–904. 2008. View Article : Google Scholar
|
|
20
|
Shen Z, Deng H, Fang Y, Zhu X, Ye GT, Yan
L, Liu H and Li G: Identification of the interplay between SOX9 and
S100P in the metastasis and invasion of colon carcinoma.
Oncotarget. 6:20672–20684. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Choi BJ, Park SA, Lee SY, Cha YN and Surh
YJ: Hypoxia induces epithelial-mesenchymal transition in colorectal
cancer cells through ubiquitin-specific protease 47-mediated
stabilization of Snail: A potential role of Sox9. Sci Rep.
7:159182017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang S, Che D, Yang F, Chi C, Meng H,
Shen J, Qi L, Liu F, Lv L, Li Y, et al: Tumor-associated
macrophages promote tumor metastasis via the TGF-β/SOX9 axis in
non-small cell lung cancer. Oncotarget. 8:99801–99815. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Carrasco-Garcia E, Lopez L, Aldaz P,
Arevalo S, Aldaregia J, Egaña L, Bujanda L, Cheung M, Sampron N,
Garcia I, et al: SOX9-regulated cell plasticity in colorectal
metastasis is attenuated by rapamycin. Sci Rep. 6:323502016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Matheu A, Collado M, Wise C, Manterola L,
Cekaite L, Tye AJ, Canamero M, Bujanda L, Schedl A, Cheah KS, et
al: Oncogenicity of the developmental transcription factor Sox9.
Cancer Res. 72:1301–1315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jay P, Berta P and Blache P: Expression of
the carcinoembryonic antigen gene is inhibited by SOX9 in human
colon carcinoma cells. Cancer Res. 65:2193–2198. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zalzali H, Naudin C, Bastide P,
Quittau-Prévostel C, Yaghi C, Poulat F, Jay P and Blache P:
CEACAM1, a SOX9 direct transcriptional target identified in the
colon epithelium. Oncogene. 27:7131–7138. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Marcker Espersen ML, Linnemann D,
Christensen IJ, Alamili M, Troelsen JT and Høgdall E: SOX9
expression predicts relapse of stage II colon cancer patients. Hum
Pathol. 52:38–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lai CC, You JF, Yeh CY, Chen JS, Tang R,
Wang JY and Chin CC: Low preoperative serum albumin in colon
cancer: A risk factor for poor outcome. Int J Colorectal Dis.
26:473–481. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Boonpipattanapong T and Chewatanakornkul
S: Preoperative carcinoembryonic antigen and albumin in predicting
survival in patients with colon and rectal carcinomas. J Clin
Gastroenterol. 40:592–595. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dimitriou N, Arandjelović O, Harrison DJ
and Caie PD: A principled machine learning framework improves
accuracy of stage II colorectal cancer prognosis. NPJ Digit Med.
1:522018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vera R, Aparicio J, Carballo F, Esteva M,
González-Flores E, Santianes J, Santolaya F and Fernández-Cebrián
JM: Recommendations for follow-up of colorectal cancer survivors.
Clin Transl Oncol. 21:1302–1311. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
van Diest PJ, van Dam P, Henzen-Logmans
SC, Berns E, van der Burg ME, Green J and Vergote I; European
Organization for Research and Treatment of Cancer-Gynaecological
Cancer Cooperative Group, : A scoring system for
immunohistochemical staining: Consensus report of the task force
for basic research of the EORTC-GCCG. J Clin Pathol. 50:801–804.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lizárraga-Verdugo E, Ruiz-García E,
López-Camarillo C, Bermúdez M, Avendaño-Félix M, Ramos-Payán R,
Romero-Quintana G, Ayala-Ham A, Villegas-Mercado C, Pérez-Plasencia
C, et al: Cell curvival is regulated via SOX9/BCL2L1 axis in
HCT-116 colorectal cancer cell line. J Oncol. 2020:57015272020.
View Article : Google Scholar
|
|
35
|
Wang L, He S, Yuan J, Mao X, Cao Y, Zong
J, Tu Y and Zhang Y: Oncogenic role of SOX9 expression in human
malignant glioma. Med Oncol. 29:3484–3490. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mauri G, Sartore-Bianchi A, Russo AG,
Marsoni S, Bardelli A and Siena S: Early-onset colorectal cancer in
young individuals. Mol Oncol. 13:109–131. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xicola RM, Manojlovic Z, Augustus GJ,
Kupfer SS, Emmadi R, Alagiozian-Angelova V, Triche T Jr, Salhia B,
Carpten J, Llor X, et al: Lack of APC somatic mutation is
associated with early-onset colorectal cancer in African Americans.
Carcinogenesis. 39:1331–1341. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wan YP, Xi M, He HC, Wan S, Hua W, Zen ZC,
Liu YL, Zhou YL, Mo RJ, Zhuo YJ, et al: Expression and clinical
significance of SOX9 in renal cell carcinoma, bladder cancer and
penile cancer. Oncol Res Treat. 40:15–20. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mesquita P, Freire AF, Lopes N, Gomes R,
Azevedo D, Barros R, Pereira B, Cavadas B, Pópulo H, Boaventura P,
et al: Expression and clinical relevance of SOX9 in gastric cancer.
Dis Markers. 2019:82670212019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Prévostel C and Blache P: The
dose-dependent effect of SOX9 and its incidence in colorectal
cancer. Eur J Cancer. 86:150–157. 2017. View Article : Google Scholar
|
|
41
|
Lytle NK, Barber AG and Reya T: Stem cell
fate in cancer growth, progression and therapy resistance. Nat Rev
Cancer. 18:669–680. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Beck B and Blanpain C: Unravelling cancer
stem cell potential. Nat Rev Cancer. 13:727–738. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kuşoğlu A and Biray Avcı Ç: Cancer stem
cells: A brief review of the current status. Gene. 681:80–85. 2019.
View Article : Google Scholar
|
|
44
|
Boland CR, Thibodeau SN, Hamilton SR,
Sidransky D, Eshleman JR, Burt RW, Meltzer SJ, Rodriguez-Bigas MA,
Fodde R, Ranzani GN, et al: A National Cancer Institute Workshop on
Microsatellite Instability for cancer detection and familial
predisposition: Development of international criteria for the
determination of microsatellite instability in colorectal cancer.
Cancer Res. 58:5248–5257. 1998.PubMed/NCBI
|
|
45
|
Boland CR and Goel A: Microsatellite
instability in colorectal cancer. Gastroenterology.
138:2073–2087.e3. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Santos JC, Carrasco-Garcia E, Garcia-Puga
M, Aldaz P, Montes M, Fernandez-Reyes M, de Oliveira CC, Lawrie CH,
Araúzo-Bravo MJ, Ribeiro ML, et al: SOX9 Elevation Acts with
Canonical WNT Signaling to Drive Gastric Cancer Progression. Cancer
Res. 76:6735–6746. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang J, Xue X, Hong H, Qin M, Zhou J, Sun
Q, Liang H and Gao L: Upregulation of microRNA-524-5p enhances the
cisplatin sensitivity of gastric cancer cells by modulating
proliferation and metastasis via targeting SOX9. Oncotarget.
8:574–582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Higashihara T, Yoshitomi H, Nakata Y,
Kagawa S, Takano S, Shimizu H, Kato A, Furukawa K, Ohtsuka M and
Miyazaki M: Sex determining region Y box 9 induces chemoresistance
in pancreatic cancer cells by induction of putative cancer stem
cell characteristics and its high expression predicts poor
prognosis. Pancreas. 46:1296–1304. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Aguilar-Medina M, Avendaño-Félix M,
Lizárraga-Verdugo E, Bermúdez M, Romero-Quintana JG, Ramos-Payan R,
Ruíz-García E and López-Camarillo C: SOX9 stem-cell factor:
Clinical and functional relevance in cancer. J Oncol.
2019:67540402019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu C, Liu L, Chen X, Cheng J, Zhang H,
Shen J, Shan J, Xu Y, Yang Z, Lai M, et al: Sox9 regulates
self-renewal and tumorigenicity by promoting symmetrical cell
division of cancer stem cells in hepatocellular carcinoma.
Hepatology. 64:117–129. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Network TCGA; Cancer Genome Atlas Network,
: Comprehensive molecular characterization of human colon and
rectal cancer. Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bahmad HF, Cheaito K, Chalhoub RM, Hadadeh
O, Monzer A, Ballout F, El-Hajj A, Mukherji D, Liu YN, Daoud G, et
al: Sphere-formation assay: Three-dimensional in vitro culturing of
prostate cancer stem/progenitor sphere-forming cells. Front Oncol.
8:3472018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shaheen S, Ahmed M, Lorenzi F and Nateri
AS: Spheroid-Formation (Colonosphere) Assay for in vitro assessment
and expansion of stem cells in colon cancer. Stem Cell Rev Rep.
12:492–499. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Terasaki M, Matsumoto N, Hashimoto R, Endo
T, Maeda H, Hamada J, Osada K, Miyashita K and Mutoh M: Fucoxanthin
administration delays occurrence of tumors in xenograft mice by
colonospheres, with an anti-tumor predictor of glycine. J Clin
Biochem Nutr. 64:52–58. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rennebeck G, Martelli M and Kyprianou N:
Anoikis and survival connections in the tumor microenvironment: Is
there a role in prostate cancer metastasis? Cancer Res.
65:11230–11235. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Stratton MR: Exploring the genomes of
cancer cells: Progress and promise. Science. 331:1553–1558. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jinesh GG, Choi W, Shah JB, Lee EK, Willis
DL and Kamat AM: Blebbishields, the emergency program for cancer
stem cells: Sphere formation and tumorigenesis after apoptosis.
Cell Death Differ. 20:382–395. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Larsimont J-C, Youssef KK, Sánchez-Danés
A, Sukumaran V, Defrance M, Delatte B, Liagre M, Baatsen P, Marine
JC, Lippens S, et al: Sox9 controls self-renewal of oncogene
targeted cells and links tumor initiation and invasion. Cell Stem
Cell. 17:60–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Prévostel C, Rammah-Bouazza C, Trauchessec
H, Canterel-Thouennon L, Busson M, Ychou M and Blache P: SOX9 is an
atypical intestinal tumor suppressor controlling the oncogenic
Wnt/β-catenin signaling. Oncotarget. 7:82228–82243. 2016.
View Article : Google Scholar
|
|
60
|
Ma XL, Hu B, Tang WG, Xie S-H, Ren N, Guo
L and Lu RQ: CD73 sustained cancer-stem-cell traits by promoting
SOX9 expression and stability in hepatocellular carcinoma. J
Hematol Oncol. 13:112020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Richtig G, Aigelsreiter A, Schwarzenbacher
D, Ress AL, Adiprasito JB, Stiegelbauer V, Hoefler G, Schauer S,
Kiesslich T, Kornprat P, et al: SOX9 is a proliferation and stem
cell factor in hepatocellular carcinoma and possess widespread
prognostic significance in different cancer types. PLoS One.
12:e01878142017. View Article : Google Scholar : PubMed/NCBI
|