Introduction
Breast cancer is one of the most commonly diagnosed
cancer among women and the fifth cause of cancer-associated death
worldwide (1). Although systemic
therapies, including surgery, chemo/radiotherapy and targeted
therapy, lead to marked improvements in clinical outcome, the
survival rates of patients with breast cancer remain unsatisfactory
in developing countries, considering that breast cancer in
developing countries accounts for 50% of all breast cancer cases
and 62% of cancer-associated deaths (2). Docetaxel (DOC), an effective
chemotherapeutic drug, is indispensable for standard treatment in
breast cancer, according to the latest National Comprehensive
Cancer Network guidelines (3).
However, the presence of chemoresistance is considered as one of
the barriers that leads to poor efficacy (4). Therefore, it is imperative to gain a
deeper understanding of the key molecular mechanisms associated
with chemoresistance in breast cancer.
Long non-coding RNAs (lncRNAs) are a class of
non-coding RNAs >200 nucleotides in length (5). Evidence has indicated that lncRNAs have
a critical role in tumorigenesis, serving either as oncogenes or
tumor suppressors in different types of cancer, such as gastric,
liver and colon cancer (6,7). In addition, lncRNAs are also associated
with chemoresistance of tumors (8).
For example, the lncRNA Colorectal Neoplasia Differentially
Expressed has been shown to facilitate colorectal cancer
chemoresistance through the Wnt/β-catenin signaling pathway
(9). Metastasis-associated lung
adenocarcinoma transcript 1 modulates autophagy-associated
chemoresistance in gastric cancer (10), whereas the lncRNA Urothelial Cancer
Associated 1 promotes cisplatin resistance in oral squamous cell
carcinoma by inhibiting miR-184 (11).
There are numerous different forms of lncRNAs that
are involved in the processes of biological regulation, including
cell proliferation, apoptosis and cell cycle regulation, among
which antisense (AS) lncRNA is usually reverse-transcribed from the
corresponding gene (7). TMPO-AS1,
located on chromosome 12, was previously identified in human lung
cancer (12). Emerging evidence has
indicated that TMPO-AS1 could act as an indicator of the prognosis
of patients with lung cancer through regulation of the cell cycle
and cell adhesion (13). Another
study demonstrated that TMPO-AS1 served an oncogenic role and may
be a potentially novel prognostic biomarker as well as a
therapeutic target in prostate cancer (14). However, the function and mechanism of
TMPO-AS1 in DOC resistance and invasion in breast cancer cells has
yet to be fully elucidated. In addition, tripartite
motif-containing protein 37 (TRIM37) has been shown to be involved
in the development of breast cancer (15), but the role of TRIM37 in breast
cancer chemotherapy is unclear.
The purpose of the present study was to investigate
the association between TMPO-AS1 and breast cancer drug resistance,
and to identify the key targets and molecular axes of TMPO-AS1
affecting breast chemotherapy resistance. The current findings may
provide novel insights into the mechanism in which breast cancer
acquires chemoresistance, facilitating the development of novel
therapeutical drugs for breast cancer.
Materials and methods
Cell lines and cell culture
Breast cancer cell lines (MDA-MB-231 and MCF7 cell
lines) were acquired from The Cell Bank of Type Culture Collection
of The Chinese Academy of Sciences. Cells were treated with
increasing concentrations of DOC (20-1,000 nM) for 48 h. The
surviving DOC-resistant sublines, MDA-231/DOC and MCF-7/DOC, were
established successfully. The cells cultured in DMEM medium
(Nanjing KeyGen Biotech. Co., Ltd.) were supplemented with 10%
fetal bovine serum (FBS) (Gibco; Thermo Fisher Scientific, Inc.) at
37°C in a humidified atmosphere of 5% CO2.
Cell transfection
pcDNA3.1-TMPO-AS1, pcDNA3.1-TRIM37 and empty vector
plasmids (2 µg used for transfection) were acquired from
GeneCopoeia, Inc. In total, three siRNAs specifically targeting
TMPO-AS1 (10 nM; si-TMPO-AS1 #1, 5′-CCGCCAAACGCCCGCCTTT-3′;
si-TMPO-AS1 #2, 5′-AGGTAGAAACGCAGTTTAA-3′; and si-TMPO-AS1 #3,
5′-GAGCCGAACUACGAACCAATT-3′) or TRIM37 (10 nM; si-TRIM37,
5′-GGAGAAGATTCAGAATGAA-3′) and scrambled siRNA negative control (10
nM; si-NC, 5′-UUCUCCGAACGUGUCACGU-3′), miR-1179 mimic (10 nM;
5′-AAGCAUUCUUUCAUUGGUUGG-3′), scrambled miRNA control (10 nM;
miR-NC, 5′-AGGTAGAAACGCAGTTTAA-3′), miR-1179 inhibitor (10 nM;
miR-inhibitor, 5′-CAGUACUUUUGUGUAGUACAA-3′) and scrambled inhibitor
control (10 nM; inhibitor-NC, 5′-CAGUACUUUUGUGUAGUACAA-3′) were
acquired from Shanghai GenePharma Co., Ltd. All transfection
processes were performed with Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C for 48 h
according to the manufacturer's instructions. Cells were subjected
to subsequent experimentation 48 h following transfection.
Reverse transcription-quantitative PCR
analysis (RT-qPCR)
Total RNA from the breast cancer cells was extracted
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). First-strand cDNA was synthesized with Reverse
Transcriptase XL (Takara Bio, Inc.) in the presence of random
primers and oligo(dT) with the following temperature protocol: 30
min at 16°C, 30 min at 42°C and 5 min at 85°C. RT-qPCR was
conducted on an ABI 7300 real-time PCR machine (Applied Biosystems;
Thermo Fisher Scientific, Inc.) with the SYBR Green reaction mix
(Applied Biosystems; Thermo Fisher Scientific, Inc.). β-actin was
used as the internal reference of lncRNA and mRNA qPCR, while U6 as
the internal reference of miRNA. Primers for TMPO-AS1 were as
follows: Forward, 5′-AGCCCACACACTACAGGCA-3′ and reverse,
5′-GCACAAAAGCAGTACGACCT-3′. Primers for miR-1179 as follows:
Forward, 5′-AAGCATTCTTTCATTGGTTGGA-3′ and reverse
CTCTACAGCTATATTGCCAGCCAC. Primers for U6 were: Forward,
5′-TGCGGGTGCTCGCTTCGGCAGC-3′ and reverse 5′-GTGCAGGGTCCGAGGT-3′.
Primers for β-actin were: Forward, 5′-CCTCTCCCAAGTCCACACAG-3′ and
reverse, 5′-GGGCACGAAGGCTCATCATT-3′. Primers for TRIM37 were:
Forward, 5′-TGGACTTACTCGCAAATG-3′ and reverse:
5′-ATCTGGTGGTGACAAATC-3′. The following thermocycling conditions
were used for PCR: Pre-denaturation at 93°C for 15 sec, then
denaturation at 90°C for 25 sec, annealing at 58°C for 25 sec and
extension at 72°C for 30 sec for 35 cycles. The 2−ΔΔCq
method was used to analyze gene expression levels (16).
DOC-resistance analysis
In vitro cancer cell proliferation was
detected using a Cell Counting Kit-8 (CCK-8) assay. Briefly, the
proliferation of non-DOC-treated cancer cells was regarded as the
100% standard. MDA-231/DOC and MCF-7/DOC cells were processed with
various concentrations of DOC in order to determine the
half-maximal inhibitory concentration (the IC50 value).
Subsequently, following incubation at room temperature for 48 h,
cell viability was detected with CCK-8 (Dojindo Molecular
Technologies, Inc.), following the manufacturer's instructions. The
absorbance of the breast cancer cells was measured at 450 nm using
a microplate reader (Bio-Rad Laboratories, Inc.).
Western blot analysis
The cancer cells were first lysed in RIPA buffer
incubated with cocktail protease inhibitors (both from Beyotime
Institute of Biotechnology) in order to extract the total protein.
Subsequently, proteins (30 µg/lane) were separated via 10% SDS-PAGE
and then blotted onto a PVDF membrane (EMD Millipore).
Subsequently, the PVDF membrane was probed with primary antibodies
(anti-TRIM37, dilution 1:1,000; Abcam) was incubated at 4°C
overnight. The membrane was then incubated with the HRP-conjugated
goat anti-rabbit IgG secondary antibody (1:1,000; cat. no.
ab205718; Abcam) at room temperature for 2 h. Finally, blots were
detected and visualized with an ECL detection kit (Santa Cruz
Biotechnology, Inc.).
Transwell assay
The transfected MDA-231/DOC and MCF-7/DOC cells were
rinsed with PBS and incubated at 37°C for 1 h with serum-free DMEM.
Subsequently, the cell suspension (5×104 cells) was
transferred to the upper chamber of a Boyden apparatus (BD
Biosciences), with the lower chamber filled with Complete™ medium
containing 20% FBS (Gibco; Thermo Fisher Scientific, Inc.). After
the non-migrating cells had been cleared from the upper chamber,
the lower chamber of migrating cells were incubated with 4%
methanol for 20 min at room temperature, followed by staining with
1% crystal violet for 20 min at room temperature. The stained cells
were then observed and counted using an inverted light microscope
(magnification, ×200; Olympus Corporation).
Apoptosis analysis
MDA-231/DOC and MCF-7/DOC cells were incubated in
six-well plates (2×104 cells/well) for 24 h, followed by
treatment with 50 ng/l DOC at room temperature for 24 h. The
harvested cells were subsequently washed thrice with pre-cooled
PBS. The cells were treated with 3 ml annexin V for 10 min at room
temperature in the dark, followed by an incubation with 2 ml
propidium iodide (PI) at room temperature for 5 min. Subsequently,
the extent of apoptosis with annexin V and PI was determined in the
dark through analysis using an Attune NxT flow cytometer (Thermo
Fisher Scientific, Inc.). The apoptosis results were analyzed using
FlowJo version 10.4 (FlowJo LLC).
Target prediction and luciferase
reporter assay
Starbase version 3.0 (http://starbase.sysu.edu.cn/) and TargetScan
(www.targetscan.org; Human 7.2) databases
were used to predict the potential binding sites. The pMIR-REPORT
Luciferase vector (Promega Corporation) of TMPO-AS1-wild-type (Wt)
or TMPO-AS1-mutant (Mut) reporters were co-transfected with NC
mimics or miR 1179 mimics into 293T cells (The Cell Bank of Type
Culture Collection of The Chinese Academy of Sciences) with
Lipofectamine 2000 as aforementioned. Similarly, pMIR-TRIM37-Wt or
pMIR-TRIM37-Mut reporters were co-transfected with NC mimics or
miR-1179 mimics into 293T cells with Lipofectamine 2000. After 36
h, the relative luciferase activity was detected using a Dual
Luciferase Reporter assay system (Promega Corporation) and
normalized to Renilla luciferase activity.
Statistical analysis
The data are presented as the mean ± SD with three
independent experiments. All data were analyzed using GraphPad
Prism software, version 7.0 (GraphPad Software, Inc.). Unpaired
Student's t-test or one-way ANOVA analyses followed by Tukey's post
hoc test were performed to distinguish differences between two and
three or more groups, respectively. P<0.05 was considered to
indicate a statistically significant difference.
Results
TMPO-AS1 levels are markedly elevated
in DOC-resistant breast cancer cells
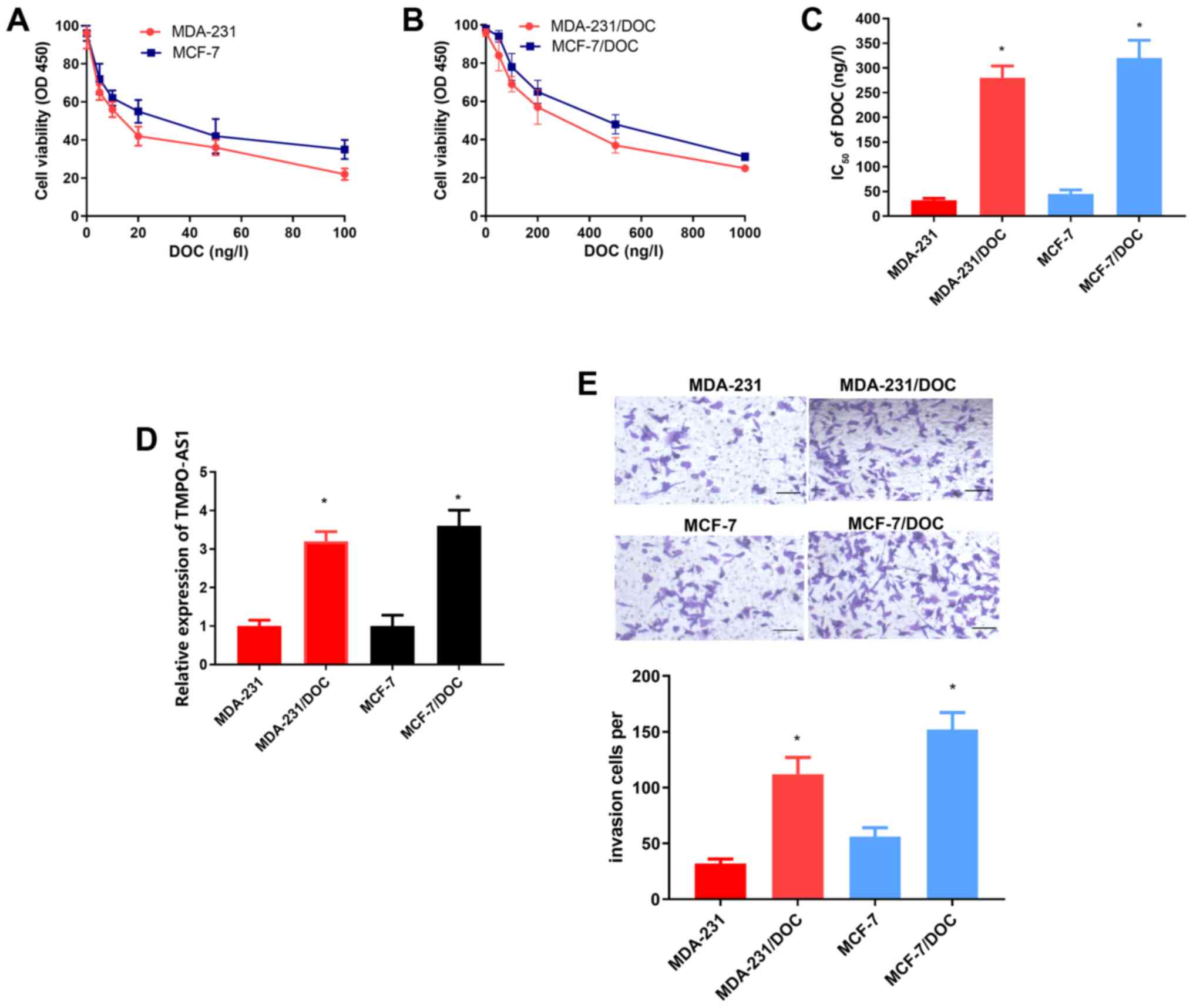
To confirm whether TMPO-AS1 was associated with DOC
resistance in breast cancer, DOC-resistant MDA-231/DOC and
MCF-7/DOC breast cancer cell lines were constructed. The
IC50 values of DOC in MDA-231/DOC and MCF-7/DOC cells
were significantly increased compared with that of their parental
cells (both P<0.05; Fig. 1C),
demonstrating the successful establishment of breast cancer cells
resistant to DOC. Furthermore, the level of TMPO-AS1 was
significantly increased in MDA-231/DOC and MCF-7/DOC cells compared
with their parental cells (both P<0.05; Fig. 1D). In invasion assays, MDA-231/DOC
and MCF-7/DOC exhibited an enhanced invasion capability compared
with that of their parental MDA-231 and MCF-7 cells (both
P<0.05; Fig. 1E). Taken together,
these results suggested that TMPO-AS1 was overexpressed in
DOC-resistant breast cancer cells, and this may provide one of the
reasons underlying the resistance of breast cancer cells to
DOC.
TMPO-AS1 knockdown enhances
sensitivity to DOC and suppresses migration in DOC-resistant breast
cancer cells
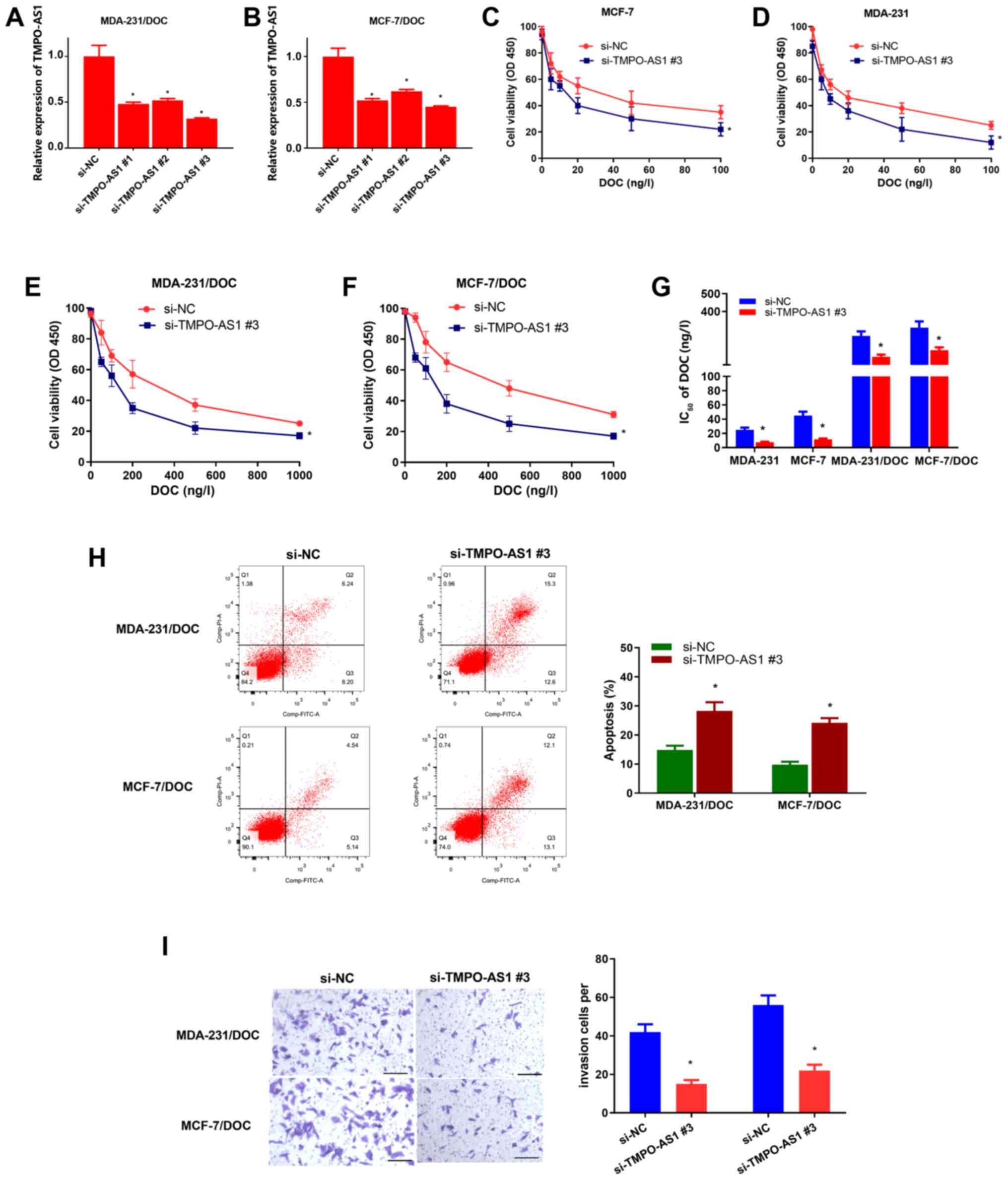
To further confirm the functional role of TMPO-AS1
in breast cancer cells, MDA-231/DOC and MCF-7/DOC cells were
transfected with TMPO-AS1 siRNAs (si-TMPO-AS1#1, si-TMPO-AS1#2 or
si-TMPO-AS1#3) or si-NC. As shown in Fig. 2A and B, transfection with TMPO-AS1
siRNAs (especially si-TMPO-AS1#3) led to a significant reduction in
TMPO-AS1 expression in MDA-231/DOC and MCF-7/DOC cells (P<0.05).
Therefore, si-TMPO-AS1 #3 was selected for the subsequent
experiments. The CCK-8 assay revealed that TMPO-AS1-knockdown led
to a marked decrease in the proliferative activity of MDA-231,
MCF-7, MDA-231/DOC and MCF-7/DOC cells (Fig. 2C-F). TMPO-AS1 knockdown also caused a
marked decrease in the IC50 value of MDA-231, MCF-7,
MDA-231/DOC and MCF-7/DOC cells (Fig.
2G), demonstrating that TMPO-AS1-knockdown increased the
sensitivity of MDA-231/DOC and MCF-7/DOC cells to DOC. To confirm
the mechanism by which TMPO-AS1-knockdown reversed DOC resistance,
flow cytometric analysis was performed following transfection in
MDA-231/DOC and MCF-7/DOC cells treated with DOC. The results
indicated that TMPO-AS1-knockdown significantly increased
DOC-induced apoptosis in MDA-231/DOC and MCF-7/DOC cells (both
P<0.05; Fig. 2H). In invasion
assays, TMPO-AS1-knockdown attenuated the rate of invasion of
MDA-231/DOC and MCF-7/DOC cells (both P<0.05; Fig. 2I). Collectively, these results
demonstrated that knockdown of TMPO-AS1 diminished DOC resistance
and invasion in MDA-231/DOC and MCF-7/DOC cells.
TMPO-AS1 knockdown sensitizes
DOC-resistant OC cells to DOC by sponging miR-1179
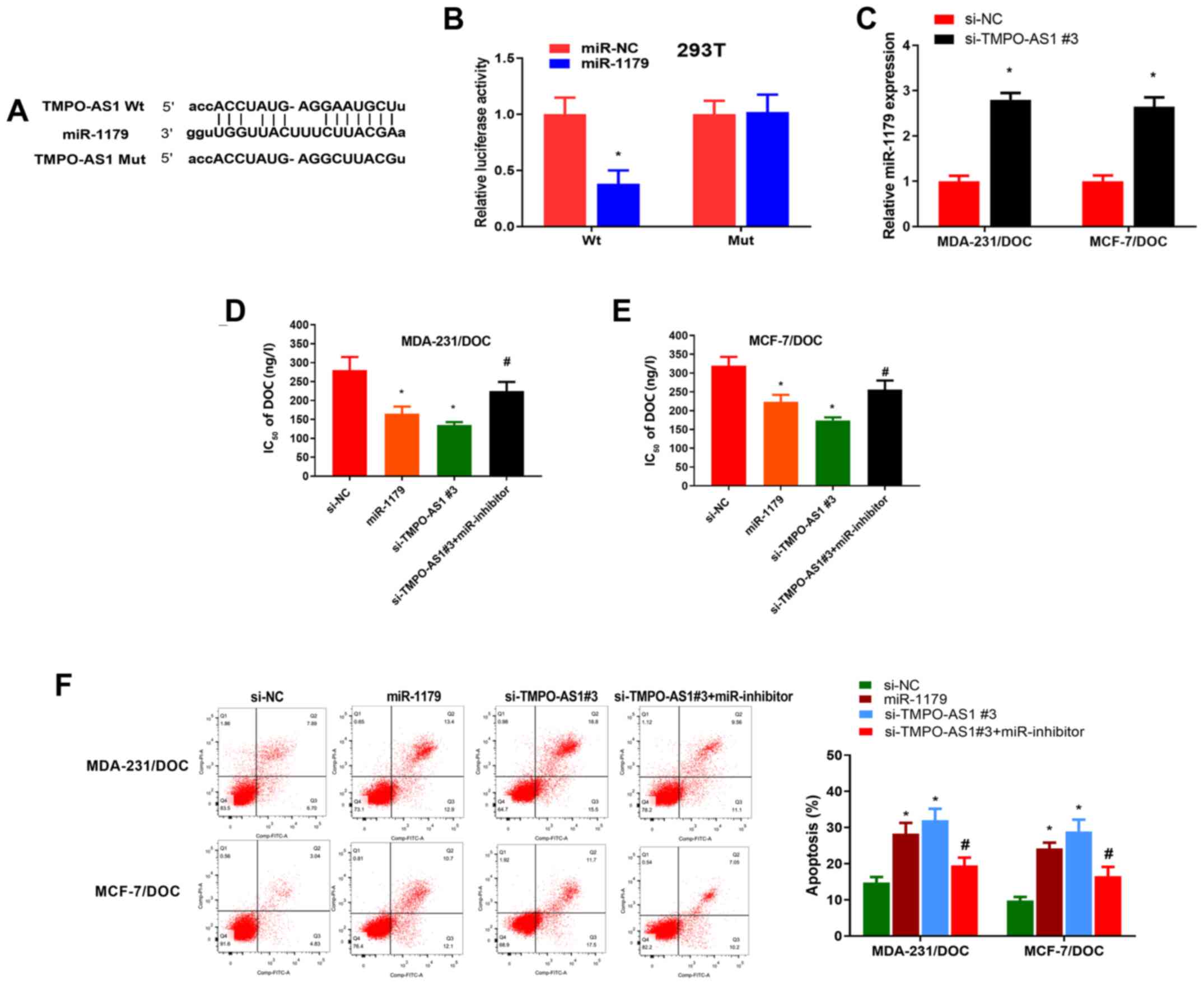
Searches of the Starbase and TargetScan databases
revealed that miR-1179, miR-198, −205-5p and −425-5p may interact
with TMPO-AS1; however, our pre-experimental results showed that
knockdown of TMPO-AS1 only affected miR-1179 (data not shown), so
miR-1179 was therefore selected for subsequent studies. According
to the bioinformatics analysis, miR-1179 was predicted to share
binding sites with TMPO-AS1 (Fig.
3A). A dual luciferase reporter assay demonstrated that forced
expression of miR-1179 markedly mitigated the luciferase activity
of TMPO-AS1-Wt reporter, but not the TMPO-AS1-Mut reporter in 293T
cells (Fig. 3B and Fig. S1A). To further confirm the effect of
TMPO-AS1 on miR-1179 expression, MDA-231/DOC and MCF-7/DOC cells
were transfected with si-TMPO-AS1#3 or si-NC, and the results
obtained demonstrated that TMPO-AS1-knockdown increased the
miR-1179 expression levels in MDA-231/DOC and MCF-7/DOC cells (both
P<0.05; Fig. 3C). All the
aforementioned data demonstrated that TMPO-AS1 directly sponged
miR-1179. Subsequently, function and rescue experiments were
conducted to investigate whether the actions of TMPO-AS1 were
mediated through sponging miR-1179. The results obtained suggested
that forced expression of miR-1179 or silencing TMPO-AS1 increased
DOC-sensitivity in MDA-231/DOC and MCF-7/DOC cells; however,
miR-1179 inhibition reversed the chemosensitization effect of
TMPO-AS1-knockdown (Fig. 3D and E
and Fig. S1B). In addition, miR-117
expression or TMPO-AS1-silencing increased DOC-induced apoptosis in
MDA-231/DOC and MCF-7/DOC cells; nevertheless, miR-1179 inhibition
reversed the co-operative effect of TMPO-AS1-knockdown on
DOC-induced apoptosis (Fig. 3F).
Taken together, these data demonstrated that TMPO-AS1-knockdown led
to decreased DOC resistance in DOC-resistant breast cancer cells
through promoting miR-1179.
TRIM37 levels increased by the
TMPO-AS1/miR-1179 axis contribute to DOC resistance in
DOC-resistant breast cancer cells
The TRIM family of proteins may have an important
role in tumor regulation (15),
although the role of the TRIM protein family in breast cancer drug
resistance has yet to be fully established. Using the Starbase and
TargetScan databases, it was shown that only TRIM37 of the TRIM
family of proteins was predicted to be a potential target of
miR-1179 (Fig. 4A). To confirm the
interaction, a dual luciferase reporter assay was conducted, which
revealed that forced expression of miR-1179 led to a marked
attenuation of the luciferase activity of the TRIM37-Wt reporter,
which was rescued by overexpression of TMPO-AS1 (Fig. 4B). Nevertheless, the luciferase
activity of TRIM37-Mut reporter exhibited no significant changes
(Fig. 4B). Subsequently, TRIM37 mRNA
and protein levels were further detected in MDA-231/DOC and
MCF-7/DOC cells. Silencing TMPO-AS1 or forced expression of
miR-1179 led to a clear decrease in the levels of TRIM37 mRNA and
protein expression; however, miR-1179 inhibition mitigated the
inhibition of TRIM37 mRNA induced by TMPO-AS1-knockdown (Fig. 4C). Western blot analysis revealed
that TMPO-AS1 silencing or miR-1179-overexpression clearly
decreased the TRIM37 protein levels (Fig. 4D). These results suggested that
TMPO-AS1 promoted TRIM37 expression by sponging miR-1179.
Subsequently, the effect of TRIM37 on DOC resistance was
investigated in MDA-231/DOC and MCF-7/DOC cells. The
IC50 assay showed that inhibition of TRIM37 markedly
promoted DOC-sensitivity in MDA-231/DOC and MCF-7/DOC cells, and
TRIM37-overexpression reversed the TMPO-AS1 knockdown-induced
chemosensitization effect (Fig. 4E and
F and Fig. S1C and D).
Additionally, TRIM37-knockdown led to an increased rate of
DOC-induced apoptosis of MDA-231/DOC and MCF-7/DOC cells, and
TRIM37-overexpression caused a diminution of the co-operative
effect of inhibition of TMPO-AS1 on DOC-induced apoptosis (Fig. 4G). Considered together, the
aforementioned results demonstrated that inhibition of TMPO-AS1
sensitized DOC-resistant OC cells to DOC via inhibition of TRIM37,
and this was mediated via inhibiting miR-1179 expression.
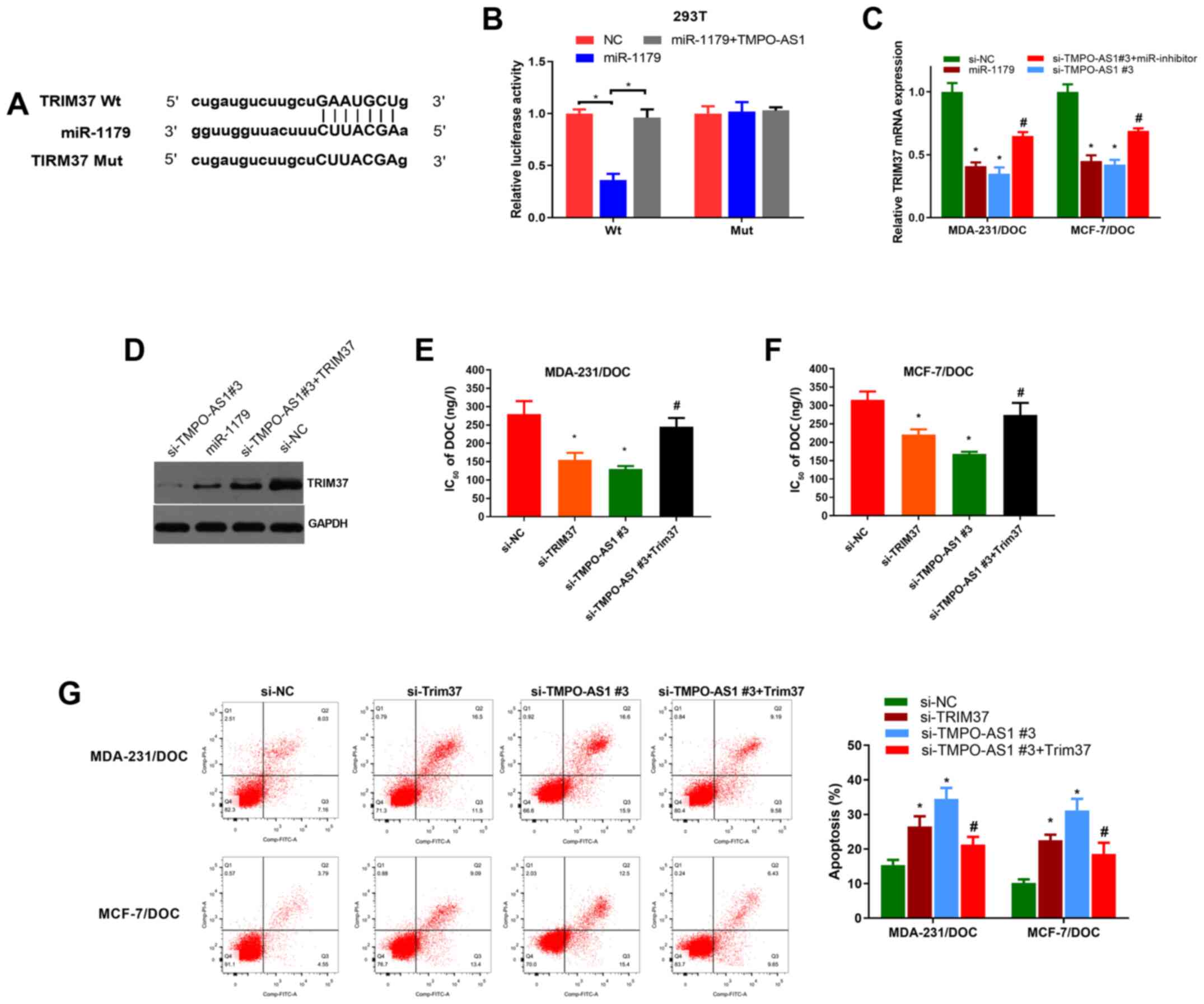 | Figure 4.TRIM37 positively modulated by the
TMPO-AS1/miR-1179 axis promotes DOC resistance in DOC-resistant
breast cells. (A) Predicted binding sites between miR-1179 and
TRIM37. (B) Luciferase reporter assay was conducted in 293T cells
transfected with TRIM37-Wt or TRIM37-Mut reporter vector and
miR-1179, miR-NC or pcDNA-TMPO-AS1. (C) TRIM37 mRNA was detected by
reverse-transcription-quantitative PCR analysis in transfected
MDA-231/DOC and MCF-7/DOC cells. (D) TRIM37 protein was detected
using western blot analysis in transfected MDA-231/DOC and
MCF-7/DOC cells. (E and F) Cell Counting Kit-8 assay was conducted
to compare IC50 values of DOC in MDA-231/DOC and
MCF-7/DOC cells after transfection. (G) Apoptotic rate was measured
via flow cytometry in MDA-231/DOC and MCF-7/DOC cells following
transfection. *P<0.05 vs. si-NC, #P<0.05 vs.
si-TMPO-AS1 #3. DOC, docetaxel; TRIM37, tripartite motif-containing
protein 37; Wt, wild-type; Mut, mutated; si, small interfering; NC,
negative control; AS1, antisense 1; miR, microRNA. |
Discussion
The poor prognosis of patients with breast cancer is
predominantly due to DOC resistance and chemotherapy failure
(1). Evidence has suggested that the
mechanisms underlying this chemoresistance are modulated by several
factors and are complex (17). In
present study, the results obtained revealed that the levels of
TMPO-AS1 were raised in DOC-resistant breast cells, and
TMPO-AS1-knockdown sensitized DOC-resistant breast cancer cells to
DOC, thereby attenuating the invasion capability of breast cancer
cells through modulating the miR-1179/TRIM37 axis.
lncRNAs have been widely identified as molecules
crucial for the chemoresistance and invasion of different cancer
types, including breast cancer (18). For example, BMP/OP-responsive gene
confers chemoresistance in triple-negative breast cancer (19). Arf-GAP with GTPase, ANK repeat and PH
domain-containing protein 2-AS1 was shown to facilitate
chemoresistance of breast cancer through regulation of myeloid
differentiation primary response 88 (20). Furthermore, Ferritin Heavy Chain 1
Pseudogene 3 promotes paclitaxel resistance in breast cancer
through sponging the miR-206/ABCB1 axis (21). Additionally, Cancer Susceptibility 9
was shown to facilitate doxorubicin-resistant breast cancer via
binding to enhancer of zeste homolog 2 (22). A recent study indicated that
TMPO-AS1, as an oncogenic lncRNA, may function as a potential
diagnostic and prognostic biomarker, and as a therapeutic target in
prostate cancer (13). However,
prior to that study, the functional and molecular role of TMPO-AS1
in the regulation of DOC resistance in different types of breast
cancer was poorly understood. In the present study, it was observed
that TMPO-AS1 was markedly upregulated in DOC-resistant breast
cancer cells. TMPO-AS1 silencing sensitized DOC-resistant breast
cancer cells to DOC and led to an improvement in DOC-induced
apoptosis; moreover, TMPO-AS1-knockdown decreased the number of
invasive cells. Therefore, collectively these results demonstrated
that TMPO-AS1 could both enhance resistance to DOC and enhance
invasion in breast cancer cells. To the best of the authors'
knowledge, the present study is the first to have shown the
promotional role of TMPO-AS1 in DOC resistance and invasion in
breast cancer, thereby revealing a potentially novel therapeutic
target for DOC resistance in breast cancer. In terms of mechanism
of lncRNA dysfunction in chemoresistance, there is evidence
indicating that transcription factors (TFs) take a key role. For
instance, lncRNA AGAP2-AS1 is induced by transcription factor SP1
in breast cancer cells to promote chemoresistance (20). In lung adenocarcinoma cells, TMPO-AS1
could be activated by E2F transcription factor 1 (23). Therefore, the authors speculated that
the abnormal expression of TMPO-AS1 caused by transcription factors
plays an important role in chemotherapy resistance of breast
cancer. However, the specific mechanism of the increased expression
of TMPO-AS1 needs to be confirmed in the future.
lncRNAs have been shown to serve as regulators of
miRNAs that function via modulating cell proliferation and
participating in chemoresistance. The mechanism through which they
accomplish these roles is usually via adsorbing and interacting
with miRNAs, and they are thereby said to act as ‘molecular
sponges’ (24). For example,
lncRNAPVT1 sponging miR-152 enhances the chemoresistance of
osteosarcoma to gemcitabine through the c-MET/PI3K/AKT pathway
(25). Additionally, Small Nucleolar
RNA Host Gene 16 restrained hepatocellular carcinoma (HCC) cell
proliferation and chemoresistance by sponging miR-93 (26). A recent study showed that TMPO-AS1
may promote the proliferation and invasion of cervical cancer cells
by regulating the miR-143-3p/zinc finger E-box-binding homeobox 1
axis (27); however, the role of
TMPO-AS1 in drug resistance of breast cancer has yet to be fully
elucidated. In the present study, TMPO-AS1 was first described to
directly inhibit miR-1179 expression. Furthermore, overexpression
of miR-1179 mitigated DOC resistance in DOC-resistant breast cancer
cells. Consistently, miR-1179 inhibition reversed the inhibitory
effect on DOC resistance induced by TMPO-AS1-knockdown. These
findings showed that silencing TMPO-AS1 increased DOC-sensitivity
via direct inhibition of miR-1179. A recent study demonstrated that
miR-1179 acts as a tumor suppressor that inhibits cell invasion via
the Notch signaling pathway in breast cancer cells (28). Another study demonstrated that
miR-1179 inhibits the proliferation, migration and invasion of
human pancreatic cancer cells by targeting E2F5 (29). In the present study, using the
Starbase and TargetScan databases, it was shown that only TRIM37
was predicted to be a potential target of miR-1179. Moreover, the
results indicated that miR-1179 could attenuate DOC resistance in
DOC-resistant breast cancer cells by targeting TRIM37.
TRIM37, a recently identified E3 ubiquitin ligase
and a member of the TRIM protein family, is dysfunctional in a
number of cancer types, and an increase in the level of TRIM37
expression is notably associated with cancer progression. For
instance, TRIM37 facilitates cell migration and metastasis in HCC
through activation of the Wnt/β-catenin signaling pathway (30). TRIM37 enhances the
epithelial-mesenchymal transition in colorectal cancer (31). Furthermore, TRIM37 overexpressed in a
subset of breast cancer types was shown to promote tumorigenesis by
the silencing of tumor suppressors (32). However, to date, the role of TRIM37
in chemoresistance of breast cancer and invasion has not been
reported. In the present study, knockdown of TRIM37 via silencing
of TMPO-AS1 significantly promoted DOC sensitivity and apoptosis,
with decreased invasion of breast cancer cells. Importantly,
TRIM37-overexpression rescued the inhibition of invasion and
sensitization effects attributable to TMPO-AS1-knockdown, which
demonstrated that TRIM37 is a key regulator in DOC resistance and
invasion in breast cancer. To the best of our knowledge, the
present study is the first to suggest the involvement of a
TMPO-AS1/miR-1179/TRIM37 molecular axis in DOC resistance and
invasion in breast cancer. However, there are some limitations to
the current study, such as the lack of further confirmation using
animal experiments and the lack of an argonatue-2-RIP experiment to
further confirm the interaction between TMPO-AS1, miR-1179 and
TRIM37 mRNA. The present results may be helpful to further confirm
the role of TMPO-AS1 in chemotherapy resistance of breast
cancer.
In conclusion, the present study revealed that
TMPO-AS1 facilitated DOC resistance and invasion in breast cancer
cells via an miR-1179 sponging-induced increase in the level of
TRIM37, revealing a novel understanding of the mechanisms
underlying chemoresistance in breast cancer, and also providing a
potential therapeutic target for breast cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the scientific
research project of Wuhan Municipal Health Commission (grant no.
WX17Q01).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XN, BL and JR designed the present study. XN, JZ, FH
and YY performed all the experiments, analyzed the data and
prepared the figures. XN drafted the initial manuscript. BL and JR
reviewed and revised the manuscript. XN and JR confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Merino Bonilla JA, Torres Tabanera M and
Ros Mendoza LH: Breast cancer in the 21st century: From early
detection to new therapies. Radiologia. 59:368–379. 2017.(In
English, Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fan L, Strasser Weippl K, Li JJ, St Louis
J, Finkelstein DM, Yu KD, Chen WQ, Shao ZM and Goss PE: Breast
cancer in China. Lancet Oncol. 15:e279–e289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Goetz MP, Gradishar WJ, Anderson BO,
Abraham J, Aft R, Allison KH, Blair SL, Burstein HJ, Dang C, Elias
AD, et al: NCCN guidelines insights: Breast cancer, version 3.2018.
J Natl Compr Canc Netw. 17:118–126. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang M, Li Y, Shen X, Ruan Y, Lu Y, Jin X,
Song P, Guo Y, Zhang X, Qu H, et al: CLDN6 promotes chemoresistance
through GSTP1 in human breast cancer. J Exp Clin Cancer Res.
36:1572017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jarroux J, Morillon A and Pinskaya M:
History, discovery, and classification of lncRNAs. Adv Exp Med
Biol. 1008:1–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peng WX, Koirala P and Mo YY:
LncRNA-mediated regulation of cell signaling in cancer. Oncogene.
36:5661–5667. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xiao J, Lv Y, Jin F, Liu Y, Ma Y, Xiong Y,
Liu L, Zhang S, Sun Y, Tipoe GL, et al: LncRNA HANR promotes
tumorigenesis and increase of chemoresistance in hepatocellular
carcinoma. Cell Physiol Biochem. 43:1926–1938. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
YiRen H, YingCong Y, Sunwu Y, Keqin L,
Xiaochun T, Senrui C, Ende C, XiZhou L and Yanfan C: Long noncoding
RNA MALAT1 regulates autophagy associated chemoresistance via
miR-23b-3p sequestration in gastric cancer. Mol Cancer. 16:1742017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fang Z, Zhao J, Xie W, Sun Q, Wang H and
Qiao B: LncRNA UCA1 promotes proliferation and cisplatin resistance
of oral squamous cell carcinoma by sunppressing miR-184 expression.
Cancer Med. 6:2897–2908. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li DS, Ainiwaer JL, Sheyhiding I, Zhang Z
and Zhang LW: Identification of key long non-coding RNAs as
competing endogenous RNAs for miRNA-mRNA in lung adenocarcinoma.
Eur Rev Med Pharmacol Sci. 20:2285–2295. 2016.PubMed/NCBI
|
|
13
|
Peng F, Wang R, Zhang Y, Zhao Z, Zhou W,
Chang Z, Liang H, Zhao W, Qi L, Guo Z and Gu Y: Differential
expression analysis at the individual level reveals a lncRNA
prognostic signature for lung adenocarcinoma. Mol Cancer.
16:982017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang W, Su X, Yan W, Kong Z, Wang D,
Huang Y, Zhai Q, Zhang X, Wu H, Li Y, et al: Overexpression of
AR-regulated lncRNA TMPO-AS1 correlates with tumor progression and
poor prognosis in prostate cancer. Prostate. 78:1248–1261. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bhatnagar S and Green MR: TRIMming down
tumor suppressors in breast cancer. Cell Cycle. 14:1345–1346. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu Y, Yang Y and Liu Y, Hao Y, Zhang Y, Hu
Y, Jiang L, Gong Y, Wu K and Liu Y: Upregulation of PAG1/Cbp
contributes to adipose-derived mesenchymal stem cells promoted
tumor progression and chemoresistance in breast cancer. Biochem
Biophys Res Commun. 494:719–727. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guttman M and Rinn JL: Modular regulatory
principles of large non-coding RNAs. Nature. 482:339–346. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gooding AJ, Zhang B, Gunawardane L, Beard
A, Valadkhan S and Schiemann WP: The lncRNA BORG facilitates the
survival and chemoresistance of triple-negative breast cancers.
Oncogene. 38:2020–2041. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dong H, Wang W, Mo S, Chen R, Zou K, Han
J, Zhang F and Hu J: SP1-induced lncRNA AGAP2-AS1 expression
promotes chemoresistance of breast cancer by epigenetic regulation
of MyD88. J Exp Clin Cancer Res. 37:2022018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang R, Zhang T, Yang Z, Jiang C and Seng
J: Long non-coding RNA FTH1P3 activates paclitaxel resistance in
breast cancer through miR-206/ABCB1. J Cell Mol Med. 22:4068–4075.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang B, Li Y, Qu X, Zhu H, Tan Y, Fan Q,
Jiang Y, Liao M and Wu X: Long noncoding RNA cancer susceptibility
candidate 9 promotes doxorubicin-resistant breast cancer by binding
to enhancer of zeste homolog 2. Int J Mol Med. 42:2801–2810.
2018.PubMed/NCBI
|
|
23
|
Wei L, Liu Y, Zhang H, Ma Y, Lu Z, Gu Z
and Ding C: TMPO-AS1, a novel E2F1-regulated lncRNA, contributes to
the proliferation of lung adenocarcinoma cells via modulating
miR-326/SOX12 axis. Cancer Manag Res. 12:12403–12414. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ding B, Lou W, Xu L and Fan W: Non-coding
RNA in drug resistance of hepatocellular carcinoma. Biosci Rep.
38:BSR201809152018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun ZY, Jian YK, Zhu HY and Li B:
lncRNAPVT1 targets miR-152 to enhance chemoresistance of
osteosarcoma to gemcitabine through activating c-MET/PI3K/AKT
pathway. Pathol Res Pract. 215:555–563. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu F, Zha G, Wu Y, Cai W and Ao J:
Overexpressing lncRNA SNHG16 inhibited HCC proliferation and
chemoresistance by functionally sponging hsa-miR-93. Onco Targets
Ther. 11:8855–8863. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gang X, Yuan M and Zhang J: Long
non-coding RNA TMPO-AS1 promotes cervical cancer cell
proliferation, migration, and invasion by regulating
miR-143-3p/ZEB1 axis. Cancer Manag Res. 12:1587–1599. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li WJ, Xie XX, Bai J, Wang C, Zhao L and
Jiang DQ: Increased expression of miR-1179 inhibits breast cancer
cell metastasis by modulating Notch signaling pathway and
correlates with favorable prognosis. Eur Rev Med Pharmacol Sci.
22:8374–8382. 2018.PubMed/NCBI
|
|
29
|
Lin C, Hu Z, Yuan G, Su H, Zeng Y, Guo Z,
Zhong F, Jiang K and He S: MicroRNA-1179 inhibits the
proliferation, migration and invasion of human pancreatic cancer
cells by targeting E2F5. Chem Biol Interact. 291:65–71. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jiang J, Yu C, Chen M, Tian S and Sun C:
Over-expression of TRIM37 promotes cell migration and metastasis in
hepatocellular carcinoma by activating Wnt/β-catenin signaling.
Biochem Biophys Res Commun. 464:1120–1127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu CE and Gan J: TRIM37 promotes
epithelial-mesenchymal transition in colorectal cancer. Mol Med
Rep. 15:1057–1062. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bhatnagar S, Gazin C, Chamberlain L, Ou J,
Zhu X, Tushir JS, Virbasius CM, Lin L, Zhu LJ, Wajapeyee N and
Green MR: TRIM37 is a new histone H2A ubiquitin ligase and breast
cancer oncoprotein. Nature. 516:116–120. 2014. View Article : Google Scholar : PubMed/NCBI
|