Introduction
Lung cancer is the most common type of cancer with a
survival time of <5 years and is one of the main causes of
cancer-associated death worldwide (1,2).
Although chemotherapy and radiotherapy have clearly improved the
prognosis, these treatments cannot be applied for a long time owing
to adverse effects on the human body. Therefore, new antitumor
drugs are needed for treating lung cancer. Human cathelicidin
(hCAP)-18/LL-37 has attracted much attention due to its role in
regulating the release of proinflammatory factors in the lung
innate and adaptive immune system (3).
The LL-37 peptide, named for its 37 amino acids, has
antitumor and antimicrobial effects. A previous study showed that
the antimicrobial peptide LL-37 can exerting antitumor effects both
in vitro and in vivo (4). Thus, LL-37 is a promising antitumor
drug candidate. LL-37 can be divided into functional components
that have different roles, which possess antibacterial, antitumor,
antiparasitic and antiviral properties. The antimicrobial peptide
17BIPHE2, the shortest of the LL-37-derived peptides, was designed
and synthesized by Professor Guangshun Wang of the Nebraska Medical
Center (5). The amino acid sequence
of this peptide is GBKRLVQRLKDBLRNLV (6). Previously we have shown that the
17BIPHE2 can effectively inhibit the proliferation and biofilm
formation of methicillin-resistant staphylococcus aureus by
disrupting the integrity of microbial membranes (7). However, whether the novel synthetic
drug can inhibit tumors is unknown, and the mechanism by which
17BIPHE2 exerts its anticancer effect is not completely
understood.
ERK is a serine/threonine protein kinase that is
widely present in eukaryotic cells and belongs to the MAPK family
of proteins. The ERK signaling pathway affects cell proliferation,
invasion and migration (8). A study
has shown that phosphorylated (p-)ERK can mediate apoptosis in
pancreatic cancer cells by regulating the ratio of BAX/BCL-2
(9). Studies have also showed that
ERK controls colorectal tumor cell apoptosis by regulating the
expression of BAX and BCL-2 (8,10).
In our previous studies, we reported that 17BIPHE2
peptide can inhibit the formation of biofilms of Staphylococcus
aureus and Pseudomonas aeruginosa, and 17BIPHE2 peptide is superior
to LL-37 due to reduced cell hemolysis and quality guarantee in
artificial synthesis. Non-small cell lung cancer (NSCLC) accounts
for >85% of lung cancer cases (11), and studies on the effect of 17BIPHE2
on lung cancer cells have not yet been performed to the best of our
knowledge. A series of relevant experiments were conducted to
investigate the effect of the 17BIPHE2 peptide against the human
lung adenocarcinoma cell line, A549, and identified novel
mechanisms by which 17BIPHE2 induced apoptosis in these cells. The
effects of 17BIPHE2 on tumor biology, including proliferation,
chemotaxis, invasiveness and changes in the expression levels of
related genes and proteins were also investigated. The results may
assist in the development of a novel promising anticancer drug that
can be used as an alternative therapy for lung cancer with lower
toxicity and resistance and higher safety and efficiency.
Materials and methods
Cell culture
The human lung adenocarcinoma cancer cell line A549
was kindly supplied by the Affiliated Hospital of the Ningxia
Medical University (The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences). The lung cancer cell line NCI-H1975
and the normal human lung epithelial cell line BEAS-2B were
purchased from the BeNa Culture Collection. A549 cells were
cultured in in Roswell Park Memorial Institute (RPMI)-1640 medium
(Hyclone; Cyvita) and NCI-H1975 cells were cultured in Dulbecco's
modified Eagle's medium (Hyclone; Cyvita). BEAS-2B cells was
cultured in F12K medium (Gibco; Thermo Fisher Scientific, Inc.).
All cells were supplemented with 10% FBS, 100 U/ml penicillin and
streptomycin (all Gibco; Thermo Fisher Scientific, Inc.). The A549
cells were cultured at 37°C in a cell culture incubator with 5%
CO2. Phosphate-buffered saline (PBS) and L-glutamine
(Biological Industries) were used to wash cells. The cells were
cultured in 25-cm plates and passaged once every 1–2 days after a
1:3 split. All cell lines were used for experiments within 50
passages after thawing.
Synthesis and characterization of
17BIPHE2
The LL-37 peptide contains 37 amino acids, with two
leucine residues at the N-terminus (6). Previously, Professor Guangshun Wang
identified the minimum sequence responsible for the antimicrobial
effect of LL-37 from peptide activity assays to be the 17–29th or
18–29th amino acids of LL-37 (amino acid sequence:
GBKRLVQRLKDBLRNLV. RI-10 is a random control peptide with an amino
acid sequence of RIVQRIKDFL derived from LL-37 (5). Solid-phase chemistry was used to
synthesize the peptides with ≥95% purity.
Cytotoxicity of 17BIPHE2
Exponentially proliferating A549, NCI-H1975 and
BEAS-2B cells were harvested from 96-well plates, and
5×103 cells/well were seeded in 96-well plates and
cultured overnight at 37°C. The cells were treated with 0, 5, 10,
15, 20, 25, 30, 35 and 40 µmol/l 17BIPHE2 and RI-10 for 24 h
(treatment group) respectively. 17BIPHE2 (1:10 solved in
ddH2O) was not added to the control group, and no cells
were added in the blank group. In total, four duplicate wells were
used per group. After culturing for 24 h, the medium in the 96-well
plate was removed; then, 50 µl MTT (5 g/l; Nanjing KeyGen Biotech
Co., Ltd.) solution was added, and the cells were incubated in a
cell culture incubator at 37°C. After 4 h, 150 µl dimethyl
sulfoxide (DMSO) was added to each well and the plate was left to
stand at room temperature. After reaction for 10 min, the
absorbance at 490 nm was measured using a multiscan microplate
reader (Thermo Fisher Scientific, Inc.). Percentage cell viability
was calculated as follows: Cell viability %=[(A treatment-blank)/(A
control-A blank)] ×100%, where A is the absorbance. The experiment
was conducted in triplicate.
Cell cycle detection using flow
cytometry
A549 cells in the exponential growth phase were
collected after trypsinization, resuspended in RPMI-1640 culture
medium and seeded in a six-well plate to obtain 1×108
cells/well. The cells were cultured in 37°C incubator for 24 h and
treated with 0, 25 and 35 µmol/l 17BIPHE2 for 24 h at 37°C. The
cells in the non-treated group were used as the control. The cells
were digested with 0.25% trypsin without EDTA after 17BIPHE2
treatment. The A549 cells were washed twice with PBS for 5 min each
and then fixed with pre-cooled 70% ethanol (Hyclone; Cyvita) for 2
h at 4°C. The supernatant was removed via centrifugation at 800 × g
and 4°C for 5 min, and the pellet was washed twice with PBS,
followed by the addition of 100 µl RNaseA and incubation in a water
bath at 37°C for 30 min. Subsequently, 400 µl propidium iodide (PI)
was added, and the cell suspension was incubated at 4°C for 30 min
in the dark, followed by treatment with the Cell Cycle Detection
kit (Nanjing KeyGen Biotech Co., Ltd.) according to the
manufacturer's instructions. Finally, the cells were detected using
flow cytometry (Accuri C6; BD Biosciences) and FlowJo X version
10.0.7 (BD Biosciences) was used to analyze the data.
Detection of cellular
ultrastructure
The A549 cells were subcultured in 25-cm plates at a
density of 5×106 cells/well and treated with 0, 25 and
35 µmol/l 17BIPHE2 for 24 h. Then, the medium was discarded and the
cells were rinsed thrice with PBS and detached using trypsin with
0.25% EDTA (Beijing Solarbio Science & Technology) in PBS,
Afterward, the cells were, collected, and fixed for 2 h at 4°C with
3% glutaraldehyde solution. The cells were rinsed thrice with PBS
for 15 min each, after which they were incubated at 4°C for 1 h
with 1% osmium tetroxide, followed by two 15-min PBS washes. Next,
the cells were incubated in 30, 50 and 70% ethanol at 4°C for 10
min each; in 80 and 90% ethanol at room temperature for 10 min each
and in anhydrous ethanol (twice) at room temperature for 15 min.
The cells were then observed using a transmission electron
microscope (H-7650; Hitachi High-Technologies Corporation).
Detection of cell morphology
The A549 cells were collected at the exponential
growth stage and seeded into 24-well plates at a density of
5×104 cells/well. After culturing for at 37°C for 24 h,
the A549 cells were treated with 0, 25 and 35 µmol/l 17BIPHE2 for
24 h at 37°C in a 5% CO2 incubator. The medium was
discarded and the cells were rinsed thrice with PBS, followed by
the addition of 1 ml freshly prepared 4% paraformaldehyde into each
well and incubated for 1 h at room temperature. The cells were
washed thrice with PBS, blocked in 3% H2O2 at
room temperature for 10 min, washed again thrice with PBS, then
incubated with 0.1% Triton-X 100-sodium citrate for 2 min for
permeabilization. Next, the cells were washed thrice with PBS for 5
min each, followed by the addition of 50 µl working TUNEL solution
to each well and incubation for 1 h at 37°C. Afterward, the cells
were washed thrice with PBS and incubated with 50 µl POD working
solution for 30 min at 37°C. The cells were again washed thrice
with PBS for 5 min each, and incubated with the
3,3′-diaminobenzidine (DAB) substrates at room temperature for 10
min. Morphological changes of A549 cells were observed under a
microscope (Nikon Corporation) and 10 fields were observed under
the microscope and the mean TUNEL positive ratio was counted.
Transwell assay to detect the effect
of 17BIPHE2 on the migration and invasion of A549 cells
For the cell migration assay, the A549 cells were
cultured to the exponential growth stage and treated with 0, 25 and
35 µmol/l 17BIPHE2 for 24 h. The cells were digested with 0.25%
trypsin without EDTA and washed twice with fresh PBS for 5 min
each, after which they were resuspended in FBS-free RPMI-1640
medium. In the control group, 100 µl of cells (5×103
cells/well) were seeded in the upper chamber of the Transwell
apparatus chamber in which the concentration of 17BIPHE2 was 0
µmol/l. In the treatment group, the A549 cells were treated with 25
and 35 µmol/l 17BIPHE2 for 24 h, 100 µl of these treated cells
(5×103 cells/well) were added to the upper chamber.
Then, 600 µl RPMI-1640 medium supplemented with 10% FBS was added
to the lower chamber. After culturing for 24 h at 37°C, the culture
medium in the upper chamber was discarded, and the cells were fixed
at room temperature for 30 min with 4% paraformaldehyde, and then
stained with 0.5% purple crystal violet for 10 min at room
temperature. Next, the cells were removed from the bottom of the
upper chamber with a cotton swab and the number of cells that
penetrated the membrane was counted using a light microscope. Each
experiment was performed in duplicate and repeated thrice.
The procedure of the cell invasion assay was
identical to that described above, except for the addition of 1
mg/ml Matrigel matrix (precoating for 30 min at 37°C; Corning,
Inc.) and culturing for 24 h at 37°C. In brief, the six wells were
coated with colloidal Matrigel and each well was filled with
3×104 A549 cells/ml treated with 0, 25 and 35 µmol/l
17BIPHE2, then incubated at 37°C for 24 h before the number of
invading cells was counted using a light microscope.
Detection of 24 h cell apoptosis rate
using flow cytometry
Apoptosis assays were performed using the Annexin V
apoptosis detection kit (Nanjing Fengfeng Biomedical Technology
Co., Ltd.) according to the manufacturer's instructions. A549 cells
in the exponential growth phase were resuspended in RPMI-1640
culture medium to a density of 1×108 cells/l. The cells
were seeded and cultured in a six-well plate for 24 h and then
treated with 0, 25 and 35 µmol/l 17BIPHE2 for 24 h. The culture
solution was discarded, and the cells were washed twice with PBS
and detached using trypsin and 0.25% EDTA (Beijing Solarbio Science
& Technology) in PBS for collection. After the 2-min reaction,
excess trypsin was discarded and the digestion was continued for 3
min in trypsin. Afterwards, culture medium containing 10% FBS was
added. Finally, the cells were collected in a 1.5 ml EP tube,
followed by the addition of 400 µl Annexin V binding solution and
adjustment of the suspension concentration to ~1×106
cells/ml. Then, 5 µl Annexin V-FITC working solution was added, and
the cells were incubated in the dark for 15 min at 4°C.
Subsequently, 10 µl PI working solution was added and the cells
were incubated for 5 min in the dark at 4°C. The samples were
excited at 488 nm and detected using flow cytometry and FlowJo X
10.0.7 was used to analyze the data.
Microplate-based Ca2+
measurements
The A549 cells were detached at the exponential
growth stage using trypsin with 0.25% EDTA in PBS and resuspended
(1×108 cells/well) in FBS-free RPMI-1640 medium. The
cells were then seeded in six-well plates and incubated at 37°C for
24 h. The next day, the A549 cells were treated with 0, 25 and 35
µmol/l 17BIPHE2 for 10, 20 and 30 min, respectively, and digested
with 0.25% trypsin without EDTA. The A549 cells in the treatment
and control groups were collected and cell suspensions of
~1×106 cells/ml were prepared after washing the cells
thrice with PBS for 5 min each. Then, the calcium fluorescent probe
Fluo-4AM working buffer was added (Beyotime Institute of
Biotechnology) and the suspensions were incubated for 30 min,
followed by washing thrice with PBS for 5 min each. Finally, the
cells were resuspended in cell culture medium without FBS and
analyzed using flow cytometry as previously mentioned.
Analysis of the mitochondrial membrane
potential (ψm)
A549 cells in the logarithmic growth phase were
treated with 0.25% trypsin with EDTA and harvested, followed by
resuspension in RPMI-1640 medium supplemented with 10% FBS to a
concentration of 1×108 cells/l. The cells were plated in
six-well plates with 5×103 cells per well and incubated
for 24 h at 37°C. The next day, the A549 cells were treated with 0,
25 and 35 µmol/l 17BIPHE2 for 24 h at 37°C, followed by treatment
with 0.25% trypsin without EDTA. The collected A549 cells were
washed twice for 5 min each with 0.1 M PBS, and incubated with the
JC-1 working buffer (Beijing Solarbio Science & Technology) for
20 min at 37°C. Then, cell culture medium without FBS was added to
the six-well plates, and emission fluorescence was detected at 525
nm using flow cytometry as previously mentioned.
Determination of intracellular
reactive oxygen species (ROS) levels
A549 cells in the exponential growth stage were
detached using trypsin with 0.25% EDTA in PBS, plated in six-well
plates with 5×103 cells per well, and treated with 0, 25
and 35 µmol/l 17BIPHE2 for 30 min at 37°C. Intracellular ROS
production was detected using the ROS assay kit (Beyotime Institute
of Biotechnology) according to the manufacturer's instruction. The
A549 cells were collected, washed thrice with PBS for 5 min each,
and incubated with DCFH-DA working solution, which was diluted
1:1,000-fold with serum-free medium to a final concentration of 10
µmol/l. The cells were collected and suspended in diluted DCFH-DA
at the concentration of 1–20 million cells/ml, and incubated at
37°C for 20 min. The samples were mixed by inversion every 3–5 min.
The cells were washed thrice with serum-free cell culture medium to
remove the DCFH-DA at did not enter the cells. Then, the cells were
gently mixed and placed in a cell culture incubator. After 30 min
at 37°C, the cells were washed thrice with PBS for 5 min each,
followed by the addition of the cell culture medium without FBS and
the detection at 488 nm using flow cytometry as previously
mentioned.
Protein extraction and western
blotting
The A549 cells were seeded in T-75 flask plates and
cultured for 24 h until the cells reached a confluence of 80–90%.
Next, 0, 25 and 35 µmol/l 17BIPHE2 was added and the cells were
incubated for 24 h. The medium then was aspirated, and the cells
were harvested and washed twice with pre-cold PBS buffer with
shaking several times to remove the culture solution. Then, the
cells were centrifuged at 800 × g at 37°C for 30 sec and lysed via
violent oscillations for 30 sec in pre-cooled lysis buffer
containing 1× protease and phosphatase inhibitors. The samples
placed on ice and removed every 4 min, which was repeated five
times. Next, the cells were centrifuged at 16,000 × g at 4°C for 30
min and the supernatant was collected. Protein concentration was
determined using a bicinchoninic acid protein assay kit (Nanjing
KeyGen Biotech Co., Ltd.). The proteins were denatured at 95°C for
10 min and 30–50 µg of each protein was loaded per lane and
separated on a 10–15% sodium dodecyl sulfate-polyacrylamide gel.
Next, the proteins were electro-transferred to a polyvinylidene
fluoride membrane. The membrane was blocked with 5% skimmed milk or
bovine serum albumin (5% BSA for the phosphorylated protein;
Beijing Solarbio Science & Technology Co., Ltd.) at room
temperature for 1 h, followed by overnight incubation at 4°C with
the following antibodies: Anti-ERK (1:5,000; Abcam), anti-p-ERK
(1:5,000; Abcam), anti-BCL-2 (1:1,000; Abcam) and anti-BAX
(1:1,000; Abcam). The next day, the blots were washed thrice with
Tris-buffered 5% saline-Tween-20 (TBST) for 5 min each, followed by
incubation with the secondary antibody HRP-conjugated Affinipure
Goat Anti-rabbit IgG (H+L) (1:10,000 in TBST and 5% skim milk;
Abcam) at room temperature for 1 h. The blots were then again
washed thrice with TBST for 5 min each. The enhanced
chemiluminescence substrate (Thermo Fisher Scientific, Inc.) was
used to detect the protein bands and images were captured using the
Chemidoc XRS system (Bio-Rad Laboratories, Inc.). GAPDH and β-actin
(both 1:1,000; Abcam) were used as internal controls as the
molecular weight of ERK is close to that of GAPDH, while the
molecular weight of β-actin is larger compared with of ERK.
Relative protein levels were determined using the ImageJ 1.8.0
software (National Institutes of Health).
RNA interference
A549 cells (5×105 cells/l) in the
logarithmic growth phase were seeded in six-well plates and
cultured with 2 ml RPMI1640 with 10% FBS per well for 24 h until
confluence of 60–70% was reached at the time of transfection. The
sequences were as follows: siRNA-ERK, 5′-GAGCAAAUGAAAGAUGUAUTT-3′
and siRNA-Control, 5′-UUCUCCGAACGUGUCACGUTT-3′. In total, 80 pmol
siRNA (siRNA-ERK or siRNA Control, Shanghai GenePharma Co., Ltd.)
or 3.75 µl Lipofectamine® 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.) reagent was diluted in 125 µl opti-MEM
(Gibco; Thermo Fisher Scientific, Inc.) and incubated at room
temperature for 5 min. The diluted siRNA was added to each tube of
diluted Lipofectamine 3000 reagent (1:1 ratio) and incubated for 15
min at room temperature to form the siRNA-lipid complex. Then, the
siRNA-lipid complex was added to the A549 cells and incubated for
24 h. The transfection efficiency was visualized using western blot
analysis. si-RNA-transfected A549 cells were treated with 17BIPHE2
(25 or 35 µmol/l) and apoptosis was detected using flow cytometry.
Western blotting was used to detect the expression of ERK, p-ERK,
BAX and BCL-2 in A549 cells as aforementioned.
In vivo xenotransplanted tumor
model
In total, SPF grade BALB/C nude mice (n=24, average
body weight of 18–22 g) were obtained from Beijing Medical
University Animal Research Center and raised in the Experimental
Animal Center of the Ningxia Medical University. The study was
approved by the Animal Experimentation Committee of The Ningixia
Medical University (Yin Chuan, China). Animals were clinically
healthy and kept under hygienic conditions in plastic cages in an
environment maintained at 24±2°C, 55±10% humidity and 12/12 h cycle
of light and darkness for acclimatization. The animals had free
access to standard pellet diet and water ad libitum. A549
cells (5×106 cells/mouse) were injected subcutaneously
near the axilla into the right flank of BALB/c nude mice (n=5 per
experimental group) to form xenograft tumors and tumor formation
and volume were monitored on alternate days. After 21 days, tumors
with diameters of ~5 mm were observed in the back, indicating that
the nude mouse model of lung adenocarcinoma was successfully
established. Nude mice were divided into four groups: Negative
control (PBS injection), low-dose 17BIPHE2 treatment (10 mg/kg),
high-dose 17BIPHE2 treatment (20 mg/kg) and positive control
(paclitaxel injection, 10 mg/kg). The body weight and tumor growth
of the nude mice were observed for two week after the peptide
treatment. Tumor length and width were measured every other day
from the day of A549 cell injection, and the tumor volume was
calculated as follows: Tumor volume (V)=(length ×
width2/2). At 4 weeks post-cell injection, the mice were
sacrificed and the tumors were removed and measured. The mice were
placed in a 10-L box, and CO2 was slowly filled at a
flow rate of 20% to make the indoor air and CO2 reach
equilibrium. The mice were observed continuously until they did not
move (rigor mortis).
Detection of the morphology of
transplanted tumor cells using the TUNEL assay
Tissue samples from the negative control (PBS),
low-dose treatment (25 µmol/l 17BIPHE2) and high-dose treatment
group (35 µmol/l 17BIPHE2) were sectioned using a freezing
microtome, fixed with 4% paraformaldehyde and washed thrice with
PBS for 30 min each. Then, the samples were blocked with sealing
buffer (3% H2O2 in methanol) at room
temperature for 10 min, and washed thrice with PBS for 5 min each,
followed by treatment with permeabilizing buffer (Triton X-100 in
0.1% sodium citrate) at room temperature for 2 min. After washing
thrice with PBS, the TUNEL working buffer was added, and the
samples were placed in a cell culture box for 1 h. After washing
thrice with PBS for 5 min each, the samples were incubated with POD
working buffer for 30 min, followed by three washes with PBS and
incubation with DAB at room temperature for 10 min. After two PBS
washes, the samples were covered with neutral balsam and images
were captured using a light microscope for five fields of each
group.
Immunohistochemistry (IHC) for
Ki67
IHC was performed according to the manufacturer's
instructions of KI67 Cell Proliferation kit (Sangon Biotech Co.,
Ltd.) to detect Ki67 expression in the transplanted tumor. Briefly,
fresh tumor tissue samples from the negative control (PBS),
low-dose treatment (25 µmol/l 17BIPHE2) and high-dose treatment
groups (35 µmol/l 17BIPHE2) were fixed by soaking in 10% formalin
and were embedded in paraffin overnight at 65°C. Then, the samples
were sectioned at 4-µm thickness after heating for 10 min at 37°C,
deparaffinized in xylene, and rehydrated in graded ethanol [100
(twice), 90, 80, 70, 50 and 0%, each for 5 min). The tissue
sections were immersed into a dye vat filled with the antigen
repair solution and incubated under high pressure for 5 min in a
pressure cooker to remove the endogenous peroxidase. After removing
the water from the tissue sections, the samples were incubated with
3% H2O2 in a wet box for 15 min at room
temperature and washed thrice with PBS for 5 min each to remove the
excess H2O2. The slides were then incubated
with goat serum (Gibco; Thermo Fisher Scientific, Inc.) for 20 min
at room temperature. Subsequently, the slides were incubated
overnight with mouse anti-Ki67 (cat. no. PB9026; 1:200; Wuhan
Boster Biological Technology, Ltd.) antibody at 4°C. The next day,
the samples were removed and equilibrated at room temperature for
30 min. The primary antibody was washed off with PBS. Next, the
slides were incubated with reagent A at 37°C for 20 min, washed
thrice with PBS for 5 min each, and incubated with reagent B at
37°C for 30 min. The slides were washed thrice with PBS for 5 min
each and then incubated with DAB (1:20) for 2–10 min for color
development, and the reaction was stopped using pure water. Then
the nuclei were re-stained for 4 min with hematoxylin at room
temperature, rinsed thoroughly with pure water and dipped in tap
water until the color returned to blue. All slides were dehydrated
with graded ethanol [50, 70, 80, 90, 100 (twice) and 50% each for 2
min] and covered with neutral balsam for analysis under light
microscope with five fields of each group.
Statistical analysis
All values were expressed as mean ± standard
deviation calculated from three independent experiments. The data
were analyzed with one-way ANOVA followed by Tukey's multiple
comparison post hoc tests (SPSS 17.0; SPSS, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of the antibacterial peptide
17BIPHE2 on the proliferation of lung cancer cells
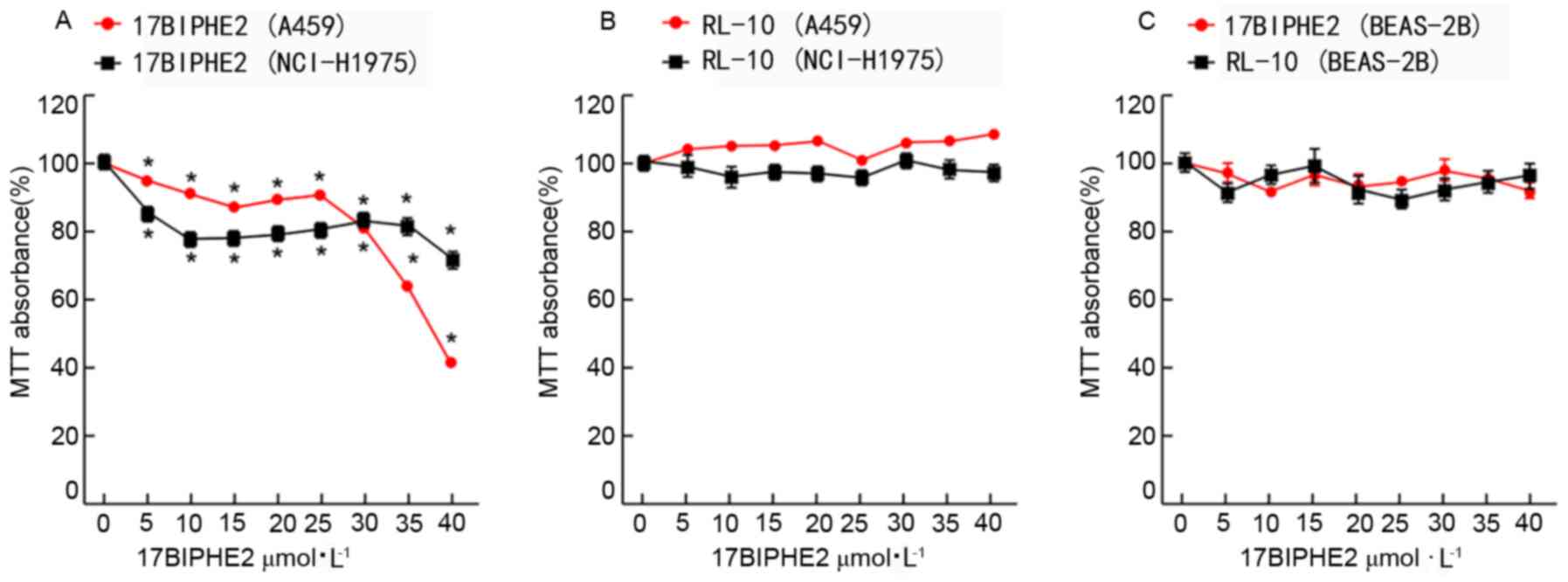
To confirm the cytotoxic effect of 17BIPHE2 in a
concentration (0, 5, 10, 15, 20, 25, 30, 35 and 40 µmol/l) -and
time (24 and 48 h)-dependent manner, the viability of treated A549
and NCI-H1975 cells and that of the normal human lung epithelial
cell line BEAS-2B was evaluated using the MTT assay. After
treatment with various concentrations of 17BIPHE2 (0-40 µmol/l) for
24 h, the viability of the A549 cells decreased from 99.1±1.3 to
40.3±0.7%, with a determined IC50 value of 34.33 µmol
(Fig. 1A). The viability of the
NCI-H1975 cells treated with 0–40 µmol/l 17BIPHE2 from 99.2±2.0% (0
µmol/l) to 71.6±2.9%, respectively, and the IC50 was
71.42 µmol/l. 17BIPHE2 treatment showed an inhibitory effect on the
viability of A549 and NCI-H1975 cells from 5 µmol/l, while
treatment with RI-10 at the same concentration range (0-40 µmol/l)
for 24 h did not show any significant inhibitory effect (Fig. 1B). The proliferation of BEAS-2B cells
was inhibited neither by 17BIPHE2 nor by RI-10 (Fig. 1C). Overall, the results showed that
17BIPHE2 inhibited the proliferation of A549 and NCI-H1975, and the
IC50 of NCI-H1975 was higher compared with that of
A549.
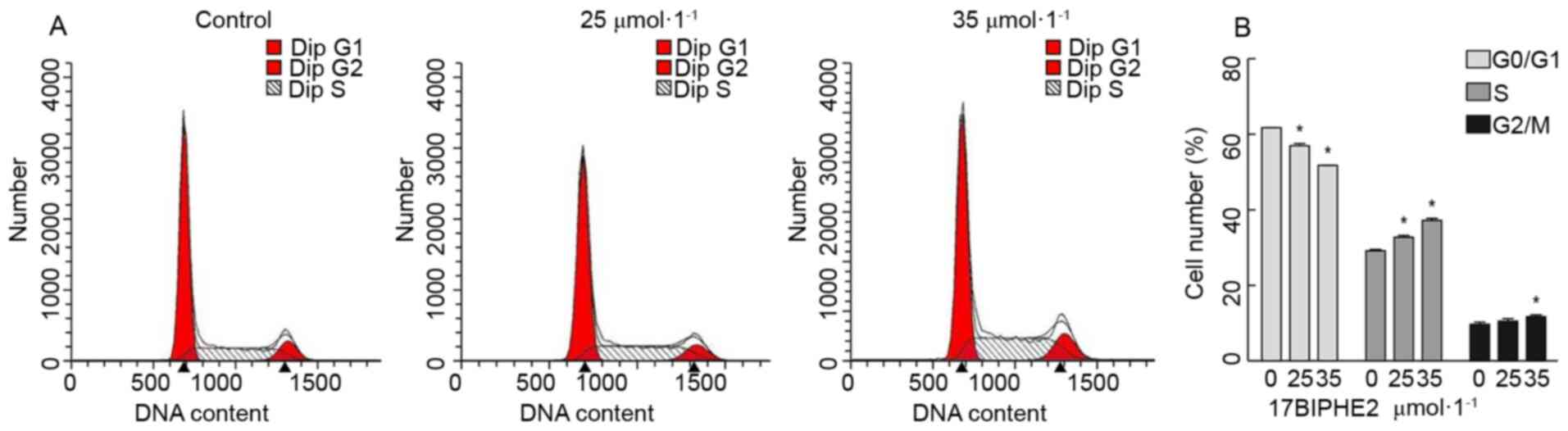
17BIPHE2 induced cell-cycle arrest in
the G1/S phase
Next, it was investigated whether the proapoptotic
effect of 17BIPHE2 is indeed mediated by the inhibition of cell
proliferation. To this end, the effect of 17BIPHE2 on the cell
cycle in A549 cells was analyzed using flow cytometry. As shown in
Fig. 2, significant S phase arrest
was observed in A549 cells was detected after treatment with 25 and
35 µmol/l 17BIPHE2 for 24 h. This effect was dose- and
time-dependent. In contrast to this, the number of cells in the
G0/G1 phase decreased with increasing
17BIPHE2 concentrations. The number of cells in the G2/M
phase increased in response to 35 µmol/l 17BIPHE2 compared with 0
µmol/l group. These results indicated the mechanism of 17BIPHE2
action involves the inhibition of the proliferation of A549 cells
and cell cycle arrest at the S phase.
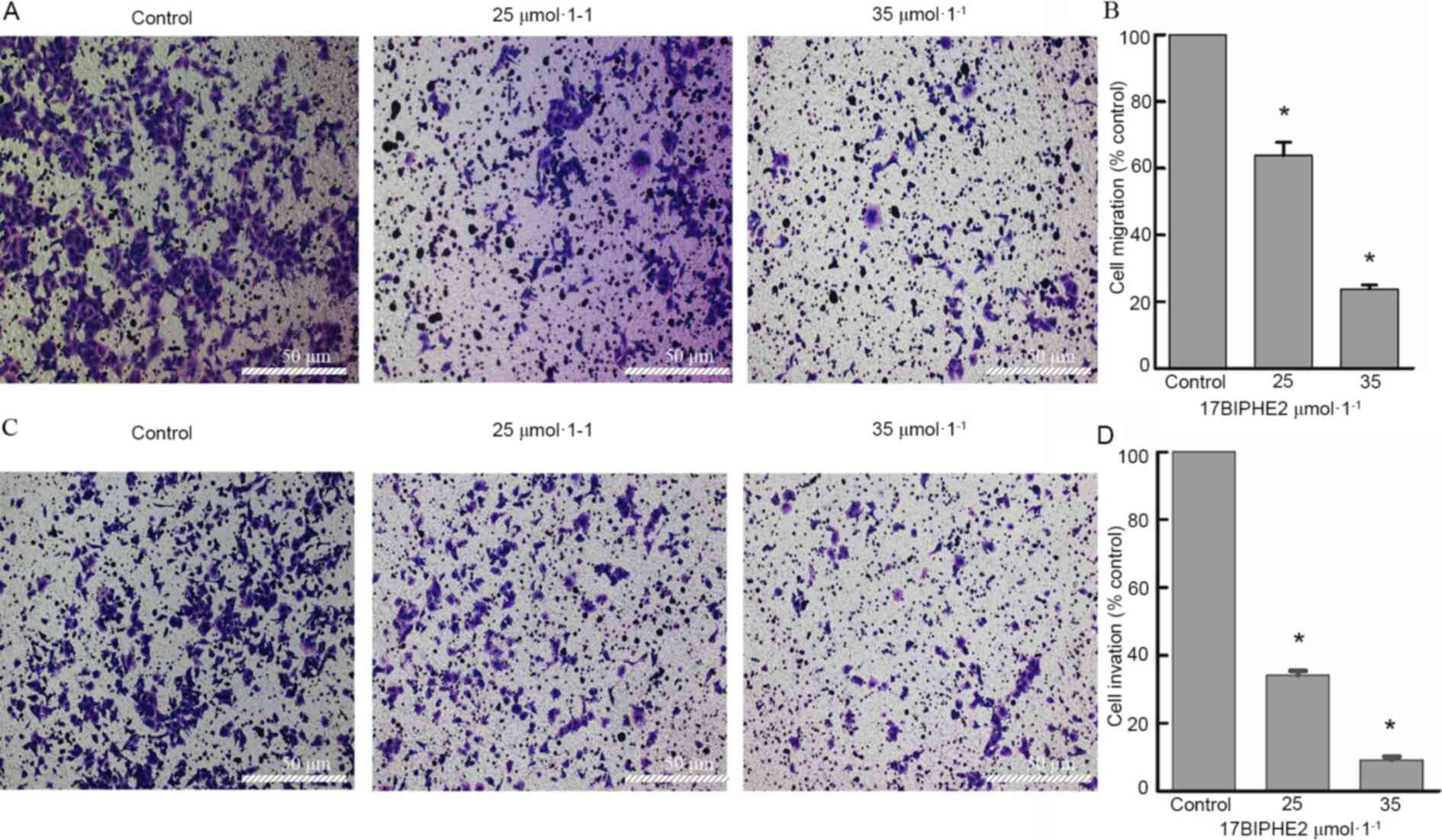
17BIPHE2 inhibits the migration and
invasion of lung cancer cells in vitro
To further investigate the effect of 17BIPHE2 in
vitro, cell migration and invasion assays were performed with
A549 cells treated with 25 and 35 µmol/l 17BIPHE2 for 24 h. The
results showed that compared with 0 µmol/l control group, the
migration rates of the A549 cells treated with 25 and 35 µmol/l
17BIPHE2 were 65.6±6.3 and 23.7±1.3%, respectively. The
corresponding invasion rates at the above indicated 17BIPHE2
concentrations were 34.2±1.2 and 9.2±0.9%, respectively. Thus, both
migration and invasion of the lung cancer cells showed a downward
trend after 17BIPHE2 treatment (Fig.
3). Overall, the results revealed that 17BIPHE2 attenuated cell
migration and invasion in a dose-dependent manner.
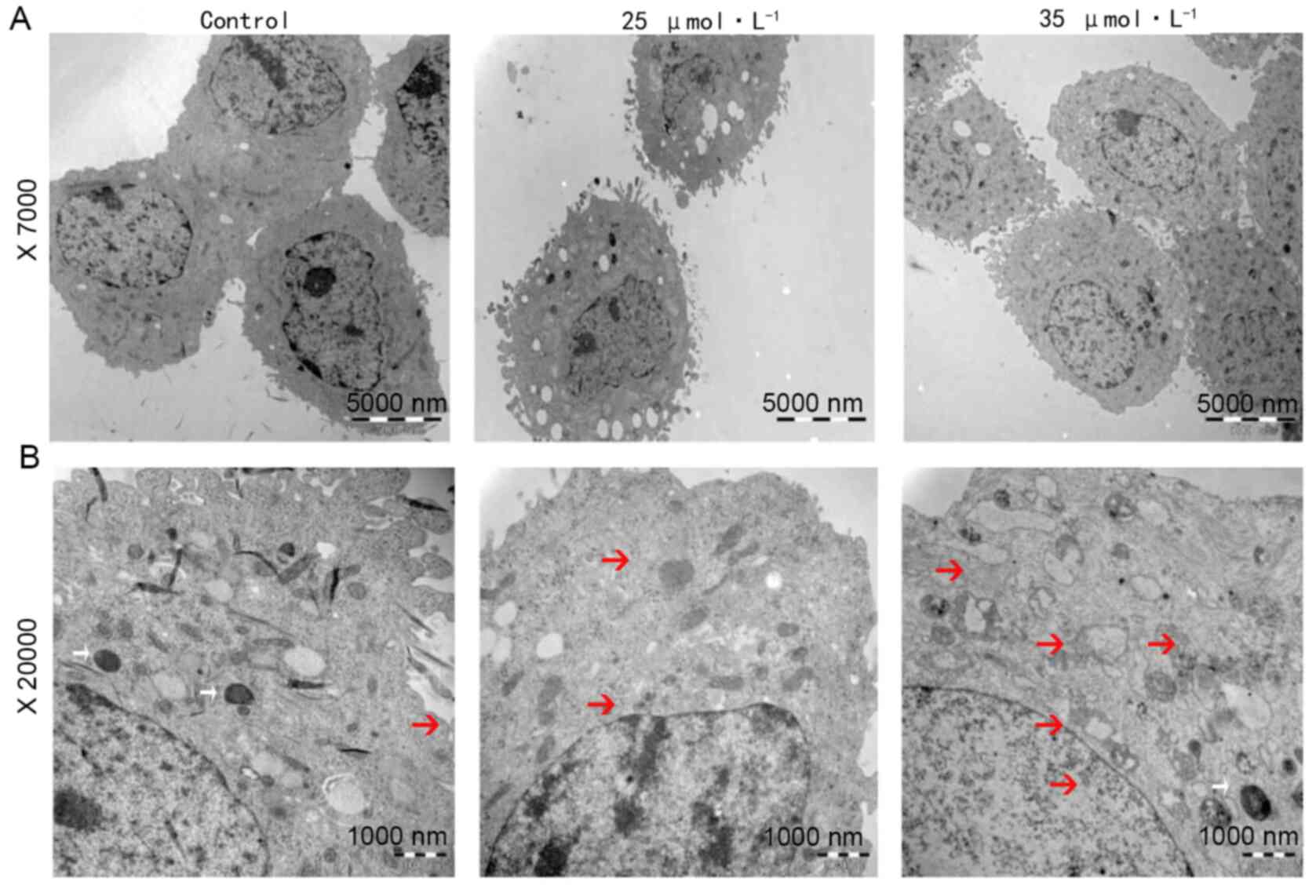
17BIPHE2-treated A549 cells show
typical morphological characteristics of apoptotic cells
To investigate the morphological changes in
apoptotic cells, A549 cells were treated with 25 and 35 µmol/l
17BIPHE2 for 24 h. Cell morphology was analyzed using transmission
electron microscopy. Cells in the control group had visible
organelles in the cytoplasm with complete structure, large number
of endoplasmic reticula, ribosome, mitochondria, uniform
distribution of chromatin in the nucleus and a clear nucleoli
structure. In contrast to this, treatment with 25 µmol/l 17BIPHE2
increased the number of intracytoplasmic vacuoles and induced
nuclear wrinkling, apoptotic body formation and cytoskeleton
disintegration. The peptide also damaged the mitochondrial crest.
Furthermore, cells treated with 35 µM 17BIPHE2 showed destruction
of the cytoplasmic organelle structure, mitochondrial ridge
rupture, endoplasmic reticulum swelling and vacuolation (Fig. 4).
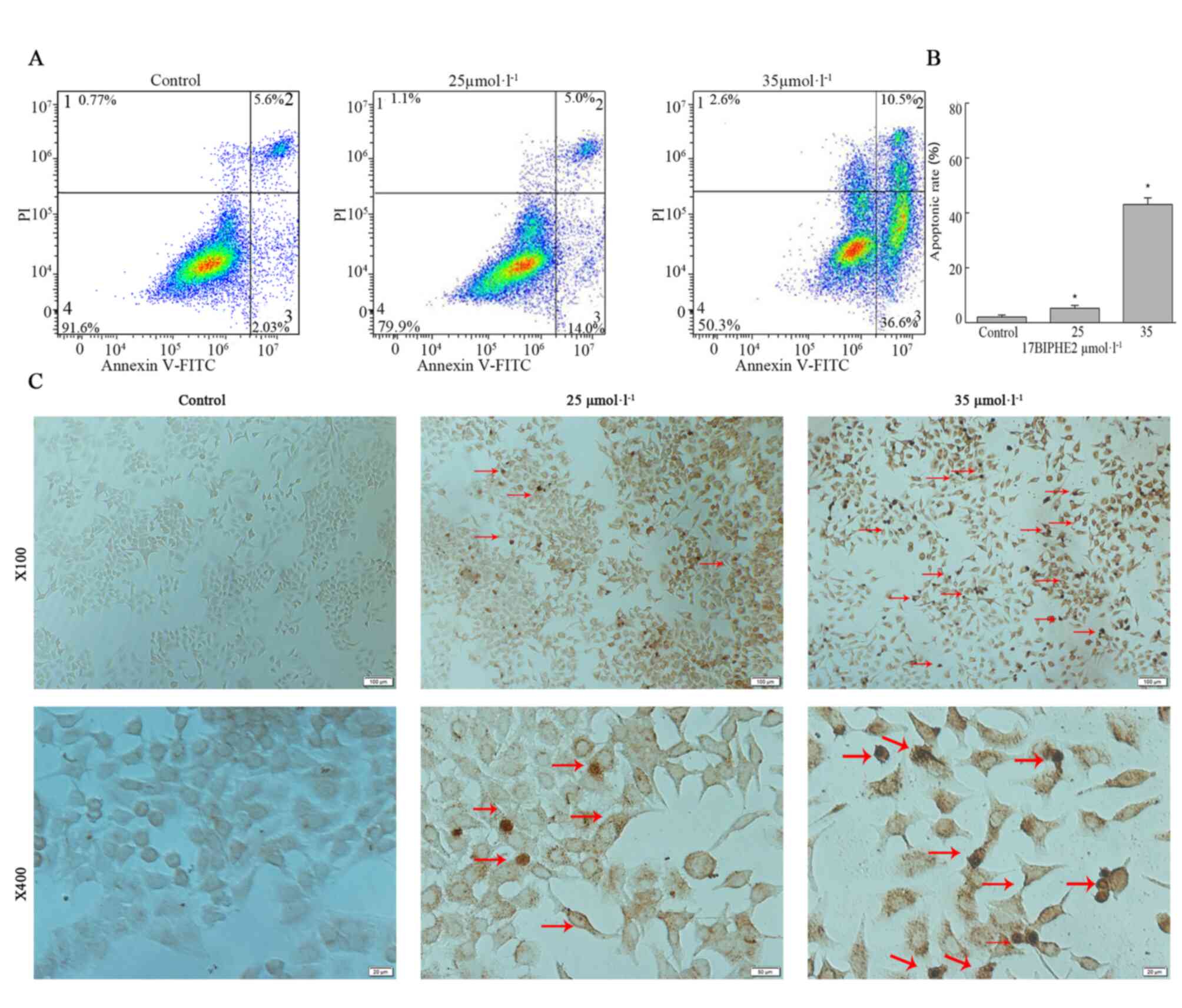
17BIPHE2 induces apoptosis in A549
cells
The morphological changes in A549 cells were evident
after 17BIPHE2 treatment, especially after treatment with high
concentrations of the peptide. Cell shrinkage was induced after
treatment with 25 µmol/l 17BIPHE2 for 24 h (Fig. 4), which may have contributed to
apoptosis. It was confirmed that 17BIPHE2 promoted apoptosis using
Annexin V/PI staining coupled with flow cytometry. As shown in
Fig. 5A, the early apoptotic
fraction increased when cells were treated with 25 µmol/l 17BIPHE2,
while treatment with 35 µmol/l 17BIPHE2 increased both the early
and late apoptotic fractions, compared with the control cells.
These observations indicated that the proportion of apoptotic cells
increased in a concentration-dependent manner after 17BIPHE2
treatment (Fig. 5A). Furthermore,
the apoptosis rate of the A549 cells increased significantly with
17BIPHE2 concentration (P<0.05; Fig.
5B).
To further demonstrate the ability of 17BIPHE2 to
induce apoptosis in lung cancer cells, a TUNEL assay was performed
using A549 cells treated with 25 and 35 µmol/l 17BIPHE2 for 24 h.
In the control group, the nuclei were round and uniformly brown and
lightly stained, and the chromatin distribution was uniform. In
contrast to this, after treatment 17BIPHE2, the cells showed
typical apoptotic morphological characteristics such as cell
shrinkage, dense and hyperchromatic chromatin and loss of integrity
(Fig. 5B). Furthermore, the
apoptosis rate of A549 cells increased significantly with 17BIPHE2
concentration (P<0.05; Fig.
5B).
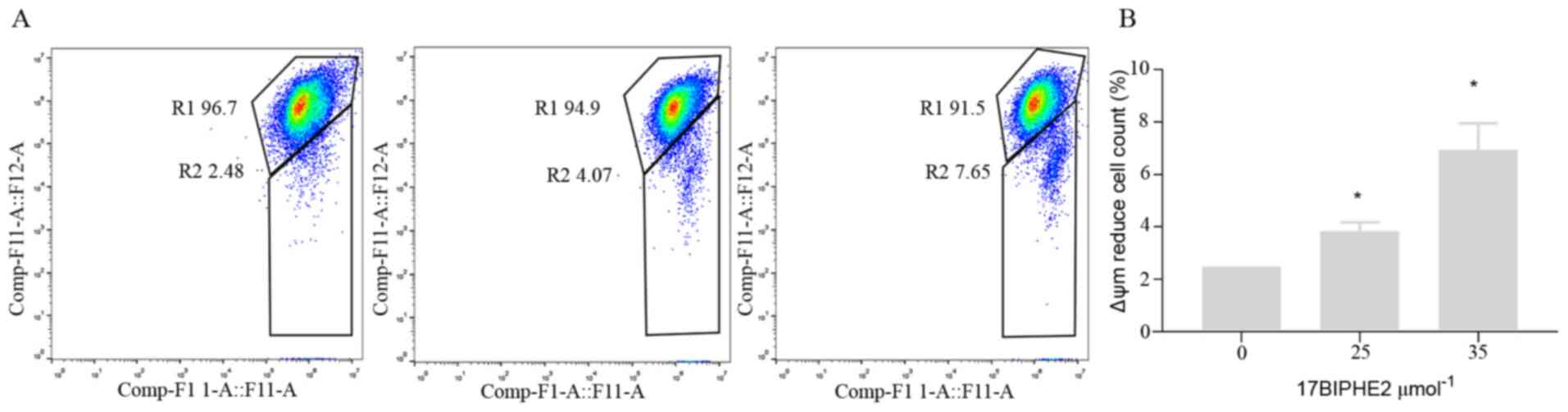
17BIPHE2 induces apoptosis in lung
cancer cells via the mitochondria-mediated apoptotic pathway
Apoptosis is mediated by two major pathways, namely
the death-receptor pathway and the mitochondrial pathway (12). Therefore, the loss of ψm, which is
another hallmark of apoptosis (13)
was further determined. Accordingly, A549 cells were treated with
25 and 35 µmol/l 17BIPHE2 for 24 h, followed by measurement of ψm.
The results showed that ψm decreased significantly as the
concentration of 17BIPHE2 increased (P<0.05; Fig. 6). Taken together, these results
demonstrated that 17BIPHE2 can induces apoptosis in A549 cells via
mitochondrial dysfunction in a dose-dependent manner.
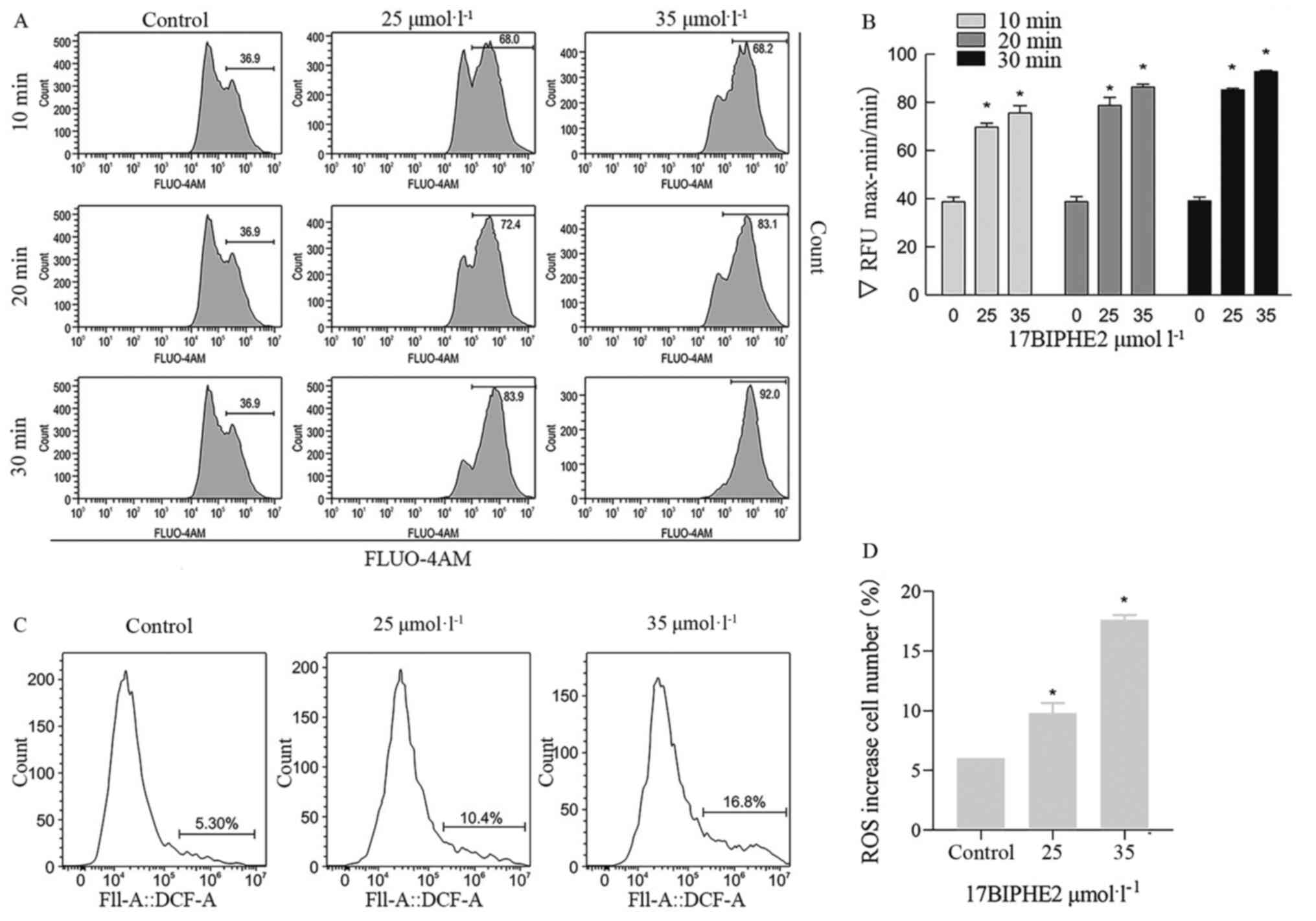
Intracellular Ca2+ and ROS
levels are elevated in A549 cells after 17BIPHE2 treatment
To further examine whether 17BIPHE2-induced
apoptosis is ROS-mediated and/or associated with the intracellular
calcium ion concentration, the intracellular calcium and ROS levels
in A549 cells after treatment with 25 and 35 µmol/l 17BIPHE2 for
10, 20 and 30 min was determined. The results showed that the
Ca2+ level in A549 cells increased significantly
(P<0.05) with the concentration of 17BIPHE2. This indicated that
17BIPHE2 increases intracellular Ca2+ activity
significantly in a time- and dose-dependent manner (Fig. 7A and B). In addition, 17BIPHE2
induced apoptosis in lung cancer cells also in a ROS-dependent
manner (Fig. 7C and D), because flow
cytometry analysis showed that the intracellular ROS levels in A549
cells were significantly elevated in a 17BIPHE2
concentration-dependent manner.
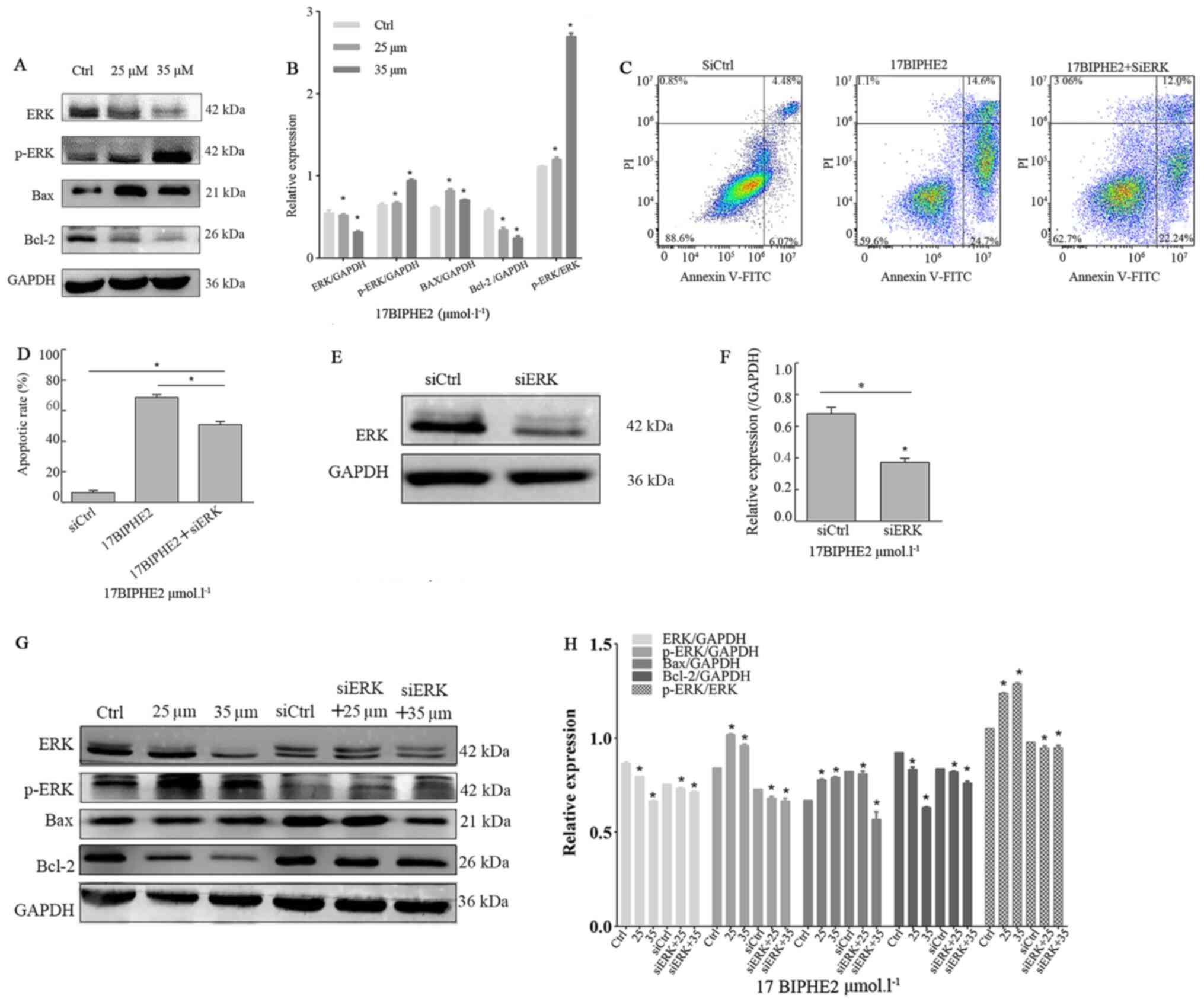
17BIPHE2 elevates p-ERK and BAX levels
while downregulating the expression of ERK and BCL-2
Next, the effect of a 24-h treatment with 25 and 35
µmol/l 17BIPHE2 on the endogenous expression of ERK, p-ERK, BAX and
BCL-2 was investigated in A549 cells using western blot analysis
(Fig. 8A and C). The protein levels
of p-ERK and BAX were upregulated in 17BIPHE2-treated cells
compared with those in the control group. In contrast to this, the
expression of ERK and BCL-2 was significantly downregulated in the
peptide-treated A549 cells compared with that in the untreated
cells. Importantly, the p-ERK/ERK ratio increased significantly
after treatment with the peptide. To further evaluate the effect of
17BIPHE2 on the expression of ERK, p-ERK, BAX and BCL-2 in A549
cells, ERK expression was silenced using siRNA targeting ERK and it
was observed that knockdown significantly affected the protein
levels of lung cancer cells compared with that of cells transfected
with the negative control siRNA (P<0.05; Fig. 8G and H). Flow cytometry analysis
showed that the apoptotic ratio in A549 cancer cells significantly
increased when they were transfected with siRNA targeting ERK after
treatment with 0 and 35 µmol/l 17BIPHE2 for 24 h (Fig. 8B). However, the apoptotic rate in
A549 cells in the control group was significantly lower compared
with that in the si-ERK-treated group (P<0.05; Fig. 8D). These data showed that
17BIPHE2-induced apoptosis of A549 cells was regulated by ERK.
Next, the role of ERK in 17BIPHE2-induced apoptosis was assessed in
A549 cells using western blot analysis. The results showed, that
after si-ERK transfection, cells treated with 17BIPHE2 showed
significantly lower expression levels of ERK and p-ERK compared
with the NC-transfected cells (Fig. 8E
and F). Furthermore, BAX was significantly upregulated in the
si-ERK group treated with 25 µmol/l 17BIPHE2 compared with the
control group; however, it was upregulated when the cells were
treated with 35 µmol/l 17BIPHE2 (P<0.05). The ratio of BAX/BCL-2
in the si-ERK group was upregulated, indicating that p-ERK
regulates the levels of BAX and BCL-2 in A549 cells, which in turn
regulates 17BIPHE2-induced apoptosis in A549 cells (Fig. 8G and H).
17BIPHE2 decreases the tumorigenicity
of lung cancer cells in vivo
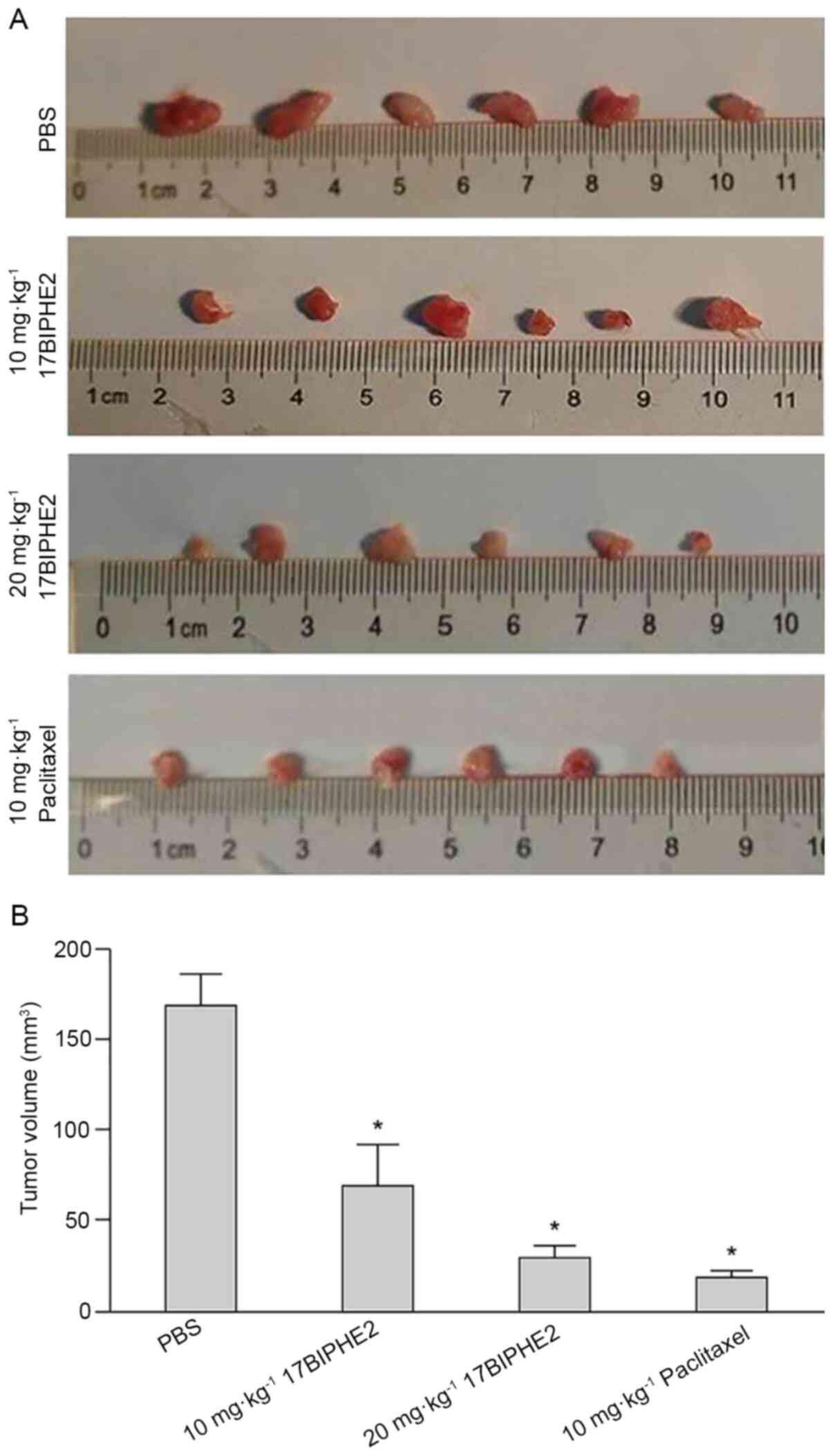
The effect of 17BIPHE2 in vivo was validated
in the BALB/c mouse model of lung tumor. Mouse xenograft models
were established by subcutaneously injecting A549 cells
(5×106 cells/mouse) into the right flank of BALB/c nude
mice, and tumor formation and volume were monitored on alternate
days. The tumor grew rapidly and locally after the injection, and
after 21 days, the tumor size was ~5 mm in diameter, indicating
that the mouse xenograft model was successfully established. Tumor
sizes of the tumors in four groups was measured after two weeks of
treatment with 17BIPHE2. The body weights differed between the
negative control group (22.06±1.46 g), low-dose treatment
(21.20±1.41 g), high-dose treatment (21.42±0.94 g) and positive
control groups (20.93±1.03 g). The mice were sacrificed and the
tumors were dissected and images were captured. The tumor sizes in
the low-dose and high-dose treatment groups were significantly
lower compared with those in the negative control group (P<0.05;
Fig. 9A and B). Thus, 17BIPHE2
significantly reduced tumor volume in xenograft mice and thereby
decreased the tumorigenicity of lung cancer cells in
vivo.
17BIPHE2 suppresses lung tumor growth
and inhibits proliferation in vivo
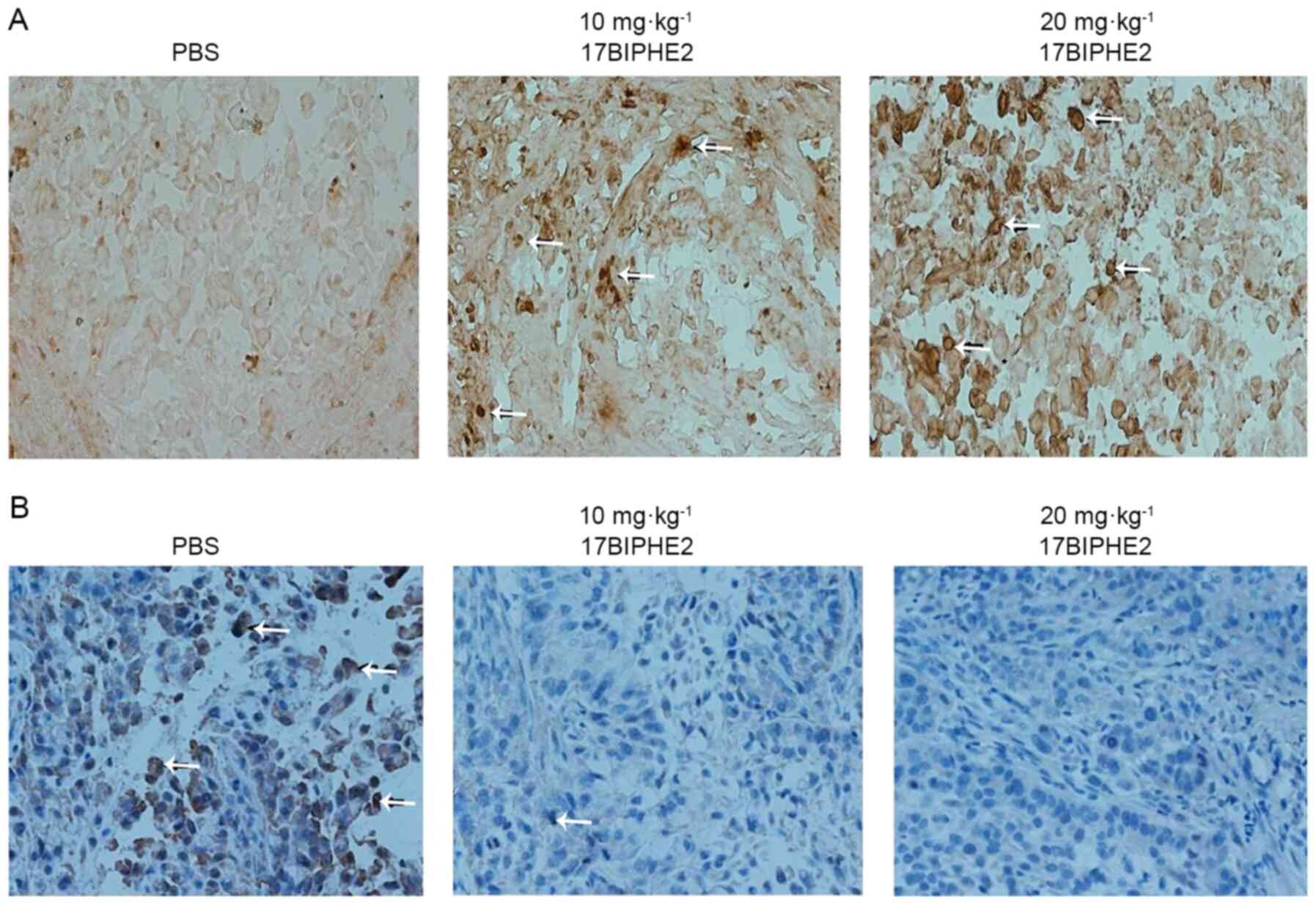
To further determine the effects of 17BIPHE2 on
apoptosis and proliferation in A549 cells in vivo, TUNEL
staining of tumor samples from nude mice treated with 17BIPHE2 was
performed. TUNEL staining of 17BIPHE2 treated tumor sections showed
that the cell nuclei stained brown and yellow (Fig. 10A). Most of the transplanted tumor
cells in the PBS-treated group were cancer cells, and hardly any
apoptotic cells were observed. Compared with the PBS group, the
groups treated with 10 and 20 mg/kg 17BIPHE2 had an increased
number of apoptotic cells in the transplanted tumor (Fig. 10B). It was then examined whether
this finding was associated with the proliferation of A549 cells.
For this reason, tumor samples were sectioned and analyzed using
immunohistochemistry for Ki67 expression, which is highly expressed
in proliferating cells (13).
Compared with the cells in the control (PBS) group,
17BIPHE2-treated cells showed suppressed expression of Ki67, shown
by the tan, diffuse and granular staining of Ki67 (Fig. 10B). Taken together, these data
demonstrated that 17BIPHE2 attenuated tumor growth via induction of
apoptosis in tumor cells and inhibition of proliferation ability of
the transplanted tumors.
Discussion
17BIPHE2 is a derivative of LL-37 and has lower
hemolytic activity and cytotoxicity compared with LL-37 (5). 17BIPHE2 have hardly been addressed in
research to the best of our knowledge, especially its effects on
lung cancer. Research shows that LL-37 plays an important role in
antibacterial (14) and antitumor
responses (15). The results showed
that 17BIPHE2 promoted apoptosis in lung cancer A549 cells. In
addition, 17BIPHE2 inhibited tumor growth in vivo and cell
proliferation in vitro. Therefore, the present study sought
to investigate whether 17BIPHE2 has similar functions to LL-37 and
aimed to explore the effects of 17BIPHE2 on lung cancer.
LL-37 is a C-terminal cleavage product of hCAP-18,
which is mainly expressed within neutrophils (3,4,9,16–19). A
study reported that LL-37 has multiple biological functions,
including antibacterial, antiviral, antifungal and antiparasitic
effects, at low concentrations (20). In addition, LL-37 inhibits the
formation of bacterial biofilms (21). The peptide does not only directly
eliminate harmful pathogenic microorganisms, but it also regulates
immune responses, which requires the interaction between multiple
molecules and cells involved in the innate and adaptive immunity
(22). However, research shows that
LL-37 may participate in the pathogenesis of malignant tumors by
regulating apoptosis in colorectal cancer (23), bladder cancer (24) and leukemia (8). LL-37 exerts its antitumor effects by
inducing apoptosis in cancer cells (25). However, LL-37 has been shown to
promote the growth of and has variable expression in ovarian cancer
(26), malignant melanoma (27) and lung cancer (28). The role of LL-37 is also
tissue-specific (29). Studies have
indicated that LL-37 plays a prominent and complex role in
antitumor response (26).
Overexpression of LL-37 promotes the development and progression of
ovarian, lung and breast cancer, and it suppresses tumorigenesis in
colon and gastric cancer (30).
Under normal conditions, LL-37 plays a role in the tumor
surveillance system. However, since LL-37 is involved in
proliferation, migration and angiogenesis, uncontrolled expression
of LL-37 could also lead to aberrant control of the aforementioned
processes and promote cancer development (31).
LL-37 kills tumor cells by regulating the
cytotoxicity of natural killer cells (32–34).
Previously, numerous studies have reported that LL-37 inhibits the
proliferation of various tumor cell lines such as those from
colorectal cancer (25), leukemia
(34) and bladder cancer (35). In the present study, it was
demonstrated that the antimicrobial peptide 17BIPHE2 affected the
proliferation, migration and invasion of human lung adenocarcinoma
A549 cells. 17BIPHE2 exerted cytotoxic effects on A549 cells but
not in normal pulmonary epithelial BEAS-2B and NCI-H1795 cells. The
MTT assay showed an evident dose- and time-dependent
antiproliferative effect for 17BIPHE2 in A549 cells. Meanwhile,
17BIPHE2 did not inhibit the proliferation of BEAS-2B cells,
indicating that the peptide was selective lung adenocarcinoma
cells, and hence has potential as an adjuvant drug for the
treatment of lung cancer.
NSCLC (of which A549 cell is the most typical cell
line) accounts for the largest proportion of lung cancer cases
(36). The poor prognosis in lung
cancer is associated with tumor metastasis (37). Cancer metastasis includes
uncontrolled cell proliferation, angiogenesis, adhesion, migration
and invasion (34). Metastasis is
one of causes of treatment failure in some malignant tumors
(38). A number of studies have
confirmed that antimicrobial peptides can inhibit tumor cell
proliferation (39–41). Liu et al (42) showed that the antimicrobial peptide
Cecropin XJ effectively inhibits the migration and invasion of
human gastric cancer AGS cells. Tian et al (43) reported that the migratory ability and
invasiveness of breast cancer cells were inhibited by the
antimicrobial peptide RP39. The present study showed that the cell
migration and invasion of A549 cells were inhibited by 17BIPHE2 in
a dose-dependent manner when the cells were treated with 25 and 35
µmol/l 17BIPHE2 for 24 h.
Inhibition of cell proliferation is closely
associated with cell cycle regulation as an anticancer mechanism,
and apoptosis usually follows cell cycle arrest (44). The cell cycle represents a survival
mechanism for tumor and normal cells because it allows cells to
repair damaged DNA (45). In the
present study, the cell cycle of A549 cells was arrested at the S
phase by 17BIPHE2 treatment, which would have inhibited DNA
synthesis (46,47) and damage repair and thereby prevented
cells from entering the division phase. These results demonstrating
DNA damage in S phase are consistent with those of Guangrui et
al (24). A previous study
showed that bladder cancer EJ and T24 cells were arrested at the S
phase by the antimicrobial peptide LL-37 (48). The mechanism of action includes
either the direct binding of antibacterial peptides to the DNA or
the inhibition of DNA polymerase activity. However, most studies
have reported that tumor cell cycle arrest induced by antimicrobial
peptides occurs at the G1/S phase (49). Although 17BIPHE2 blocked the tumor
cell cycle of A549 cells at the S phase in the present study, the
peptide showed a strong inhibitory effect on the proliferation of
tumor cells. Overall, it was demonstrated that 17BIPHE2 can promote
apoptosis and inhibit cell proliferation by blocking the cell cycle
of A549 cells.
Morphological changes accompany apoptosis and the
main characteristics of apoptotic cells include nuclear shrinkage,
DNA fragmentation, chromatin condensation, mitochondrial crest
rupture, loss of cell adhesion, cell fragmentation into apoptotic
bodies, and vacuolation and destruction of the endoplasmic
reticulum (50). In the current
study, 17BIPHE2-treated A549 cells exhibited morphological
characteristics of apoptosis. TUNEL staining and transmission
electron microscopy showed that 17BIPHE2 treatment of A549 cells
led to nuclear shrinkage, chromatin condensation and darkening,
mitochondrial ridge rupture, vacuolization of the endoplasmic
reticulum and the destruction of its normal structure (51). Compared with the control group,
treatment with 25 µmol/l 17BIPHE2 significantly increased the
percentage of early apoptotic cells, while treatment with 35 µmol/l
17BIPHE2 increased the percentage of cells in both the early and
late apoptotic stages. These results demonstrated that 17BIPHE2
induced apoptosis by inhibiting the proliferation of lung cancer
cells.
The results of the present animal experiment showed
that 17BIPHE2 promoted apoptosis of tumor cells also in
vivo. TUNEL staining of tumor sections from xenograft mice with
transplanted tumors showed that the number of apoptotic cells in
the 17BIPHE2 treatment group was higher compared with that in the
negative control group. The results of immunohistochemical staining
showed that Ki67 expression was significantly lower in the 17BIPHE2
treatment group compared with in the negative control group,
indicating that the proliferative activity of the transplanted
tumor in the treatment group was reduced.
In summary, the antimicrobial peptide 17BIPHE2
exhibited anti-lung cancer effects in vitro and in
vivo by blocking the cell cycle, inhibiting migration and
invasion and promoting apoptosis of A549 cells. Extensive research
has revealed the pivotal role of mitochondria in apoptosis.
Mitochondria are the executors of apoptosis (52). A previous study revealed that the
change in the mitochondrial Δψm is the earliest change observed in
cells after the after initiation of apoptosis (53). Mitochondria store calcium and thus
regulate the intracellular levels of calcium. When the level of
accumulated Ca2+ in the mitochondria exceeds the
threshold of the highly conductive permeability transition pore,
the pore opens, leading to a reduction in the mitochondrial
membrane potential, enhanced permeability and apoptosis (54). ROS are the products of cellular
metabolism, which are continuously produced and cleared in cells
and play important roles in the occurrence of apoptosis (55). ROS accumulation and the increase in
Ca2+ concentration coincide. Oxidative pressure
increases the Ca2+ levels in the cytoplasm and
eventually leads to calcium overload in the mitochondria. This
further increases intracellular ROS levels, which in turn
stimulates the release of intracellular calcium reserves and
results in the intracellular accumulation of Ca2+
(56). In the present study, ψm
decreased significantly when A549 cells were treated with 17BIPHE2.
However, intracellular Ca2+ and ROS levels increased
significantly and were associated with increasing concentrations of
17BIPHE2.
ERK is a serine/threonine protein kinase, which is
ubiquitous in eukaryotic cells and is a member of the MAPK family
of proteins (57). Kuriyama et
al (58) observed that ROS and
Ca2+ activate the ERK signaling pathway and participate
in the regulation of cell proliferation, differentiation, and
apoptosis. Chuang et al (53)
also reported that ROS activates ERK to mediate apoptosis in skin
cells. Cabrera Zapata et al (59) revealed that neuronal cell apoptosis
is regulated by ERK, which is activated by Ca2+. BCL-2
has an important function in the apoptosis process (60). Liu et al (61) found that the downregulation of BCL-2
expression can promote apoptosis in Vero cells. Under normal
circumstances, BAX mainly exists in the cytoplasm. When cells are
stimulated by apoptotic signals, BAX is transferred from the
cytoplasm to the mitochondria, where it increases mitochondrial
permeability and mediates apoptosis (62). Wang et al (63) observed that p-ERK can regulate the
ratio of BAX/BCL-2 to regulate apoptosis of pancreatic tumor cells.
In addition, Wang et al (64)
revealed that increased expression of p-ERK and BCL-2 and decreased
expression of BAX reduces the apoptotic rate in mouse nerve cells,
thereby protecting them from apoptotic cell death. In the present
study, the expression of p-ERK and BAX was higher in A549 cells
treated with 17BIPHE2 compared with in untreated control cells.
BCL-2 was downregulated, and the BAX/BCL-2 ratio was increased
after 17BIPHE2 treatment. After si-ERK transfection, the expression
of ERK, p-ERK and BAX decreased, whereas that of BCL-2 increased
compared with the corresponding expression rates in the control
group. The BAX/BCL-2 ratio also decreased significantly with
treatment with the peptide. These results indicated that p-ERK
regulates BAX and BCL-2 in A549 cells, which in turn regulates
apoptosis mediated by the antimicrobial peptide 17BIPHE2.
The antibacterial peptide 17BIPHE2 inhibited the
proliferation of lung adenocarcinoma A549 cells. Furthermore, it
blocked the cell cycle at the G1/S phase, decreased the
mitochondrial membrane potential and elevated the ROS and
Ca2+ levels, thereby promoting apoptosis and inhibiting
the proliferation of A549 cells. 17BIPHE2 treatment also activated
ERK, which regulated the expression of the apoptosis-related
proteins BAX and BCL-2 to mediate apoptosis of A549 cells further.
However, the mechanism of action underpinning the anticancer action
of 17BIPHE2 is still unclear, therefore further studies are
required to lay a foundation for future clinical application.
Acknowledgements
The authors would like to thank Professor Guangshun
Wang (Department of Pathology and Microbiology, College of
Medicine, University of Nebraska Medical Center, Omaha, NE, USA)
who designed and synthesized 17BIPHE2.
Funding
This research was supported by The National Natural
Science Foundation of China (grant nos. 81560573 and 81760661).
Availability of data and materials
All data are available from the corresponding author
upon reasonable request.
Authors' contributions
TY and JL confirm the authenticity of all raw data.
JL conceived and designed the present study, acquired, analyzed and
interpreted the data. TY drafted the manuscript and analyze the
relevant data. QJ prepared some relevant experimental data. YW, QZ
and SZ contributed to the data analysis of this paper. XW helped
analyze the data and critically revised the manuscript for
important intellectual content. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal
Experimentation Committee of The Ning Xia of Medical University
(Yin Chuan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Singh RD, Shandilya R, Bhargava A, Kumar
R, Tiwari R, Chaudhury K, Srivastava RK, Goryacheva IY and Mishra
PK: Quantum dot based nano-biosensors for detection of circulating
cell free miRNAs in lung carcinogenesis: From Biology to Clinical
translation. Front Genet. 9:6162018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fahy RJ and Wewers MD: Pulmonary defense
and the human cathelicidin hCAP-18/LL-37. Immunol Res. 31:75–89.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gennaro R and Zanetti M: Structural
features and biological activities of the cathelicidin-derived
antimicrobial peptides. Biopolymers. 55:31–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang X, Mishra B, Lushnikova T, Narayana
JL and Wang G: Amino acid composition determines peptide activity
spectrum and hot-spot-based design of merecidin. Adv Biosyst.
2:17002592018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang G, Hanke ML, Mishra B, Lushnikova T,
Heim CE, Chittezham Thomas V, Bayles KW and Kielian T:
Transformation of human cathelicidin LL-37 into selective, stable,
and potent antimicrobial compounds. ACS Chem Biol. 9:1997–2002.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zasloff M: Antimicrobial peptides of
multicellular organisms. Nature. 415:389–395. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mader JS, Mookherjee N, Hancock RE and
Bleackley RC: The human host defense peptide LL-37 induces
apoptosis in a calpain- and apoptosis-inducing factor-dependent
manner involving Bax activity. Mol Cancer Res. 7:689–702. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu Z, Zhang F, Bai C, Yao C, Zhong H, Zou
C and Chen X: Sophoridine induces apoptosis and S phase arrest via
ROS-dependent JNK and ERK activation in human pancreatic cancer
cells. J Exp Clin Cancer Res. 36:1242017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M,
Tafuri A, et al: Roles of the Raf/MEK/ERK pathway in cell growth,
malignant transformation and drug resistance. Biochim Biophys Acta.
1773:1263–1284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Le Chevalier T: Adjuvant chemotherapy for
resectable non-small-cell lung cancer: Where is it going? Ann
Oncol. 21 (Suppl 7):vii196–vii198. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu L, Zhang T, Liu D, Guan G, Huang J,
Proksch P, Chen X and Lin W: Notoamide-type alkaloid induced
apoptosis and autophagy via a P38/JNK signaling pathway in
hepatocellular carcinoma cells. RSC Adv. 9:19855–19868. 2019.
View Article : Google Scholar
|
|
13
|
Tian SW, Ren Y, Pei JZ, Ren BC and He Y:
Pigment epithelium-derived factor protects retinal ganglion cells
from hypoxia-induced apoptosis by preventing mitochondrial
dysfunction. Int J Ophthalmol. 10:1046–1054. 2017.PubMed/NCBI
|
|
14
|
Kang J, Dietz MJ and Li B: Antimicrobial
peptide LL-37 is bactericidal against Staphylococcus aureus
biofilms. PLoS One. 14:e02166762019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ko JK and Zhang Z: LL-37 inhibits
pancreatic cancer development through inhibition of autophagy and
reprogramming of the tumor microenvironment. Pergamon. 110:S5–S6.
2019.
|
|
16
|
Agerberth B, Charo J, Werr J, Olsson B,
Idali F, Lindbom L, Kiessling R, Jörnvall H, Wigzell H and
Gudmundsson GH: The human antimicrobial and chemotactic peptides
LL-37 and alpha-defensins are expressed by specific lymphocyte and
monocyte populations. Blood. 96:3086–3093. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dale BA and Fredericks LP: Antimicrobial
peptides in the oral environment: Expression and function in health
and disease. Curr Issues Mol Biol. 7:119–133. 2005.PubMed/NCBI
|
|
18
|
Rico-Mata R, De Leon-Rodriguez LM and
Avila EE: Effect of antimicrobial peptides derived from human
cathelicidin LL-37 on Entamoeba histolytica trophozoites. Exp
Parasitol. 133:300–306. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rekha RS, Rao Muvva SS, Wan M, Raqib R,
Bergman P, Brighenti S, Gudmundsson GH and Agerberth B:
Phenylbutyrate induces LL-37-dependent autophagy and intracellular
killing of Mycobacterium tuberculosis in human macrophages.
Autophagy. 11:1688–1699. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bucki R, Leszczyńska K, Namiot A and
Sokołowski W: Cathelicidin LL-37: A multitask antimicrobial
peptide. Arch Immunol Ther Exp (Warsz). 58:15–25. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dosler S and Karaaslan E: Inhibition and
destruction of Pseudomonas aeruginosa biofilms by antibiotics and
antimicrobial peptides. Peptides. 62:32–37. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Amatngalim GD, Nijnik A, Hiemstra PS and
Hancock RE: Cathelicidin Peptide LL-37 Modulates TREM-1 expression
and inflammatory responses to microbial compounds. Inflammation.
34:412–425. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shaykhiev R, Beisswenger C, Kändler K,
Senske J, Püchner A, Damm T, Behr J and Bals R: Human endogenous
antibiotic LL-37 stimulates airway epithelial cell proliferation
and wound closure. Am J Physiol Lung Cell Mol Physiol.
289:L842–L848. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pan G: Inhibitory effect of antimicrobial
peptide LL-37 on bladder tumor cells (unpublished PhD thesis).
Kunming Medical University; 2015
|
|
25
|
Ren SX, Shen J, Cheng AS, Lu L, Chan RL,
Li ZJ, Wang XJ, Wong CC, Zhang L, Ng SS, et al: FK-16 Derived from
the Anticancer Peptide LL-37 induces caspase-independent apoptosis
and autophagic cell death in colon cancer cells. PLoS One.
8:e636412013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Coffelt SB, Waterman RS, Florez L, Höner
zu Bentrup K, Zwezdaryk KJ, Tomchuck SL, LaMarca HL, Danka ES,
Morris CA and Scandurro AB: Ovarian cancers overexpress the
antimicrobial protein hCAP-18 and its derivative LL-37 increases
ovarian cancer cell proliferation and invasion. Int J Cancer.
122:1030–1039. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim JE, Kim HJ, Choi JM, Lee KH, Kim TY,
Cho BK, Jung JY, Chung KY, Cho D and Park HJ: The antimicrobial
peptide human cationic antimicrobial protein-18/cathelicidin LL-37
as a putative growth factor for malignant melanoma. Br J Dermatol.
163:959–967. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Von Haussen J, Koczulla R, Shaykhiev R,
Herr C, Pinkenburg O, Reimer D, Wiewrodt R, Biesterfeld S, Aigner
A, Czubayko F and Bals R: The host defence peptide LL-37/hCAP-18 is
a growth factor for lung cancer cells. Lung Cancer. 59:12–23. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chieosilapatham P, Ikeda S, Ogawa H and
Niyonsaba F: Tissue-specific regulation of innate immune responses
by human cathelicidin LL-37. Curr Pharm Des. 24:1079–1091. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ou Hongyu, Zhu Haihong, Zhu Wenjun, et al:
Antimicrobial peptide LL-37 induces apoptosis in AGS cells of
gastric cancer by activating p53 signaling pathway. J Anhui Med
Uni. 56:571–576. 2021.
|
|
31
|
Vandamme D, Landuyt B, Luyten W and
Schoofs L: A comprehensive summary of LL-37, the factotum human
cathelicidin peptide. Cell Immunol. 280:22–35. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang G: Structures of Human host defense
cathelicidin LL-37 and its smallest antimicrobial peptide KR-12 in
lipid micelles. J Biol Chem. 283:32637–32643. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Doss M, White MR, Tecle T and Hartshorn
KL: Human defensins and LL-37 in mucosal immunity. J Leukoc Biol.
87:79–92. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
An LL, Ma XT, Yang YH, Lin YM, Song YH and
Wu KF: Marked reduction of LL-37/hCAP-18, an antimicrobial peptide,
in patients with acute myeloid leukemia. Int J Hematol. 81:45–47.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Choi SY, Kim SJ, Chi BH, Kwon JK and Chang
IH: Modulating the internalization of bacille calmette-guérin by
cathelicidin in bladder cancer cells. Urology. 85:964.e7–964.e12.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ang MK and Mok TSK: Twenty-five years of
Respirology: Advances in lung cancer. Respirology. 25:26–31. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu N, Jia D, Chen W, Wang H, Liu F, Ge H,
Zhu X, Song Y, Zhang X, Zhang D, et al: FoxM1 is associated with
poor prognosis of non-small cell lung cancer patients through
promoting tumor metastasis. PLoS One. 8:e594122013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xuan Y, Zhao S, Xiao X, Xiang L and Zheng
HC: Inhibition of chaperone-mediated autophagy reduces tumor growth
and metastasis and promotes drug sensitivity in colorectal cancer.
Mol Med Rep. 23:3602021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu WK, Sung JJ, To KF, Yu L, Li HT, Li ZJ,
Chu KM, Yu J and Cho CH: The host defense peptide LL-37 activates
the tumor-suppressing bone morphogenetic protein signaling via
inhibition of proteasome in gastric cancer cells. J Cell Physiol.
223:178–186. 2010.PubMed/NCBI
|
|
40
|
Murray NP, Aedo S, Fuentealba C, Salazar
A, Reyes E, Lopez MA and Minzer S: Circulating prostate cells and
bone marrow micro-metastasis and not treatment modality determine
the risk and time to biochemical failure in low risk prostate
cancer. Arch Esp Urol. 72:1000–1009. 2019.(In Spanish). PubMed/NCBI
|
|
41
|
Sancar A, Lindsey-Boltz LA, Unsal-Kaçmaz K
and Linn S: Molecular mechanisms of mammalian DNA repair and the
DNA damage checkpoints. Annu Rev Biochem. 73:39–85. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu X, Li J, Huang L, Wang Y, Yang M, Tang
M and Qiu T: Preparation and evaluation of MPEG-PCL polymeric
nanoparticles against gastric cancer. J Wuhan Univ Technol-Mat Sci
Edit. 35:1162–1168. 2021. View Article : Google Scholar
|
|
43
|
Tian W, Li B, Zhang X, Dang W, Wang X,
Tang H, Wang L, Cao H and Chen T: Suppression of tumor invasion and
migration in breast cancer cells following delivery of siRNA
against Stat3 with the antimicrobial peptide PR39. Oncol Rep.
28:1362–1368. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhu M, Miao S, Zhou W, Elnesr SS, Dong X
and Zou X: MAPK, AKT/FoxO3a and mTOR pathways are involved in
cadmium regulating the cell cycle, proliferation and apoptosis of
chicken follicular granulosa cells. Ecotoxicol Environ Saf.
214:1120912021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bartek J, Lukas C and Lukas J: Checking on
DNA damage in S phase. Nat Rev Mol Cell Biol. 5:792–804. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Al-Ejeh F, Kumar R, Wiegmans A, Lakhani
SR, Brown MP and Khanna KK: Harnessing the complexity of DNA-damage
response pathways to improve cancer treatment outcomes. Oncogene.
29:6085–6098. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Campos A and Clemente-Blanco A: Cell cycle
and DNA repair regulation in the damage response: Protein
phosphatases take over the reins. Int J Mol Sci. 21:4462020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zeng Q: Expression of antimicrobial
peptide LL-37 in urothelial carcinoma of the bladder. Kunming
Medical University; 2015
|
|
49
|
Su L, Xu G, Shen J, Tuo Y, Zhang X, Jia S,
Chen Z and Su X: Anticancer bioactive peptide suppresses human
gastric cancer growth through modulation of apoptosis and the cell
cycle. Oncol Rep. 23:3–9. 2010.PubMed/NCBI
|
|
50
|
Fink SL and Cookson BT: Apoptosis,
pyroptosis, and necrosis: Mechanistic description of dead and dying
eukaryotic cells. Infect Immun. 73:1907–1916. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li Jun: Antimicrobial peptide 17BIPHE2
inhibits lung adenocarcinoma A549 cells and its mechanism. Ningxia
Medical University; 2018
|
|
52
|
Song J, Ham J, Hong T, Song G and Lim W:
Fraxetin suppresses cell proliferation and induces apoptosis
through mitochondria dysfunction in human hepatocellular carcinoma
cell lines Huh7 and Hep3B. Pharmaceutics. 13:1122021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chuang KC, Chen FW, Tsai MH and Shieh JJ:
EGR-1 plays a protective role in AMPK inhibitor compound C-induced
apoptosis through ROS-induced ERK activation in skin cancer cells.
Oncol Lett. 21:3042021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Schwartz GK and Shah MA: Targeting the
cell cycle: A new approach to cancer therapy. J Clin Oncol.
23:9408–9421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dong Y, Yang Y, Wei Y, Gao Y, Jiang W,
Wang G and Wang D: Facile synthetic nano-curcumin encapsulated
Bio-fabricated nanoparticles induces ROS-mediated apoptosis and
migration blocking of human lung cancer cells. Process
Biochemistry. 95:91–98. 2020. View Article : Google Scholar
|
|
56
|
Hong Y, Sun Y, Rong X, Li D, Lu Y and Ji
Y: Exosomes from adipose-derived stem cells attenuate UVB-induced
apoptosis, ROS, and the Ca2+ level in HLEC cells. Exp
Cell Res. 396:1123212020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang CL, Liu C, Niu LL, Wang LR, Hou LH
and Cao XH: Surfactin-induced apoptosis through ROS-ERS-Ca2+-ERK
pathways in HepG2 cells. Cell Biochem Biophys. 67:1433–1439. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kuriyama I, Miyazaki A, Tsuda Y, Yoshida H
and Mizushina Y: Inhibitory effect of novel somatostatin peptide
analogues on human cancer cell growth based on the selective
inhibition of DNA polymerase β. Bioorg Med Chem. 21:403–411. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Cabrera Zapata LE, Bollo M and Cambiasso
MJ: Estradiol-mediated axogenesis of hypothalamic neurons requires
ERK1/2 and ryanodine receptors-dependent intracellular
Ca2+ rise in male rats. Front Cell Neurosci. 13:1222019.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Adams JM and Cory S: The Bcl-2 protein
family: Arbiters of cell survival. Science. 281:1322–1326. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Liu Y, et al: HSV-2 miR-H4-5p negatively
regulates CDKL2 gene expression, blocking actinomycin D
(ActD)-induced apoptosis in vero cells. Chin J Biochem Mol Bio.
17:9728–9735. 2015.PubMed/NCBI
|
|
62
|
Desagher S and Martinou JC: Mitochondria
as the central control point of apoptosis. Trends in Cell Biol.
10:369–377. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wang M, Lu X, Dong X, Hao F, Liu Z, Ni G
and Chen D: pERK1/2 silencing sensitizes pancreatic cancer BXPC-3
cell to gemcitabine-induced apoptosis via regulating Bax and Bcl-2
expression. World J Surg Oncol. 13:662015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang Z, Xu Z, Niu Z, Liang B and Niu J:
Epieriocalyxin A induces cell apoptosis through JNK and ERK1/2
signaling pathways in colon cancer cells. Cell Biochem Biophys.
73:559–564. 2015. View Article : Google Scholar : PubMed/NCBI
|