Introduction
Breast cancer is one of the most common malignancies
in women (1). Since 1990, the
incidence of breast cancer in China has been continually increasing
and nearly 1.2 million new cases of breast cancer are reported
annually (2,3). The incidence, development and
metastasis of breast cancer occur due to genetic and several
environmental factors, such as long-term or repeated X-ray exposure
and air pollution (3,4). However, the pathogenesis of this
disease remains unclear; thus, current studies are focusing on the
identification of effective treatment or preventative
strategies.
Recently, traditional Chinese medicine has been
demonstrated to possess notable potential in the prevention and
treatment of tumors. Lutein is a carotene extracted from plants
with high chlorophyll content that exhibits several biological
functions, such as induction of immune cell activity, prevention of
atherosclerosis and age-associated macular degeneration and
antitumor activity (5–9). Our previous study demonstrated that
lutein can inhibit breast cancer cell proliferation by inactivating
the NF-κB pathway and activating the Nuclear factor erythroid
2-related factor (Nrf2)/antioxidant response element (ARE)
signaling pathway (10). However,
the exact mechanism of its cancer preventative action remains
unknown.
Non-coding RNAs are a class of RNA molecules without
protein-coding function (11).
Recent studies have reported that non-coding RNAs, such as long
non-coding RNAs (lncRNAs) and microRNAs (miRNAs/miRs), play
important roles in the development and progression of tumors
(12–14). lncRNAs and miRNAs can influence the
development of tumors by regulating physiological processes, such
as cell proliferation and migration, apoptosis and autophagy
(15–19). To the best of our knowledge,
previous studies have not investigated whether lncRNAs and miRNAs
enhance or affect the anticancer activity of lutein on breast
cancer cells.
The present study aimed to investigate the molecular
mechanism of the anticancer effect of lutein by investigating its
association with specific lncRNAs and miRNAs. This can provide
novel targets for the development of pharmacological agents that
can be used for the treatment of breast cancer.
Materials and methods
Cell lines and reagents
The MCF-7 and T47D breast cancer cell lines were
purchased from the American Type Culture Collection. Cells were
maintained in RPMI-1640 medium (Sigma-Aldrich; Merck KGaA)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), at 37°C with 5% CO2.
MCF-7 and T47D cells were analyzed for mycoplasma to
ensure that they were not contaminated with mycoplasma using the
Mycoplasma Detection kit (Beijing Solarbio Science & Technology
Co., Ltd.), according to the manufacturer's instructions. Lutein
was purchased from the Agri-Food Canada Research Centre. Lutein was
dissolved in different concentrations of DMSO (Beijing Solarbio
Science & Technology Co., Ltd., 0.00, 6.25, 12.50, 25.00 and
50.00 µg/ml).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was isolated using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). A
PrimeScript™ RT reagent kit (cat. no. RR047A; Takara Biotechnology
Co., Ltd.) was used to reverse transcribe the extracted RNA from
MCF-7 and T47D cells into cDNA. RT was performed at 37°C for 60
min. qPCR was subsequently performed using SYBR Green (cat. no.
DRR041A; Takara Biotechnology Co., Ltd.). The following primer
sequences were used for qPCR: miR-590-3p forward,
5′-TAATTTTATGTATAAGCTAGT-3′ and reverse, 5′-GCAGGGTCCGAGGTATTC-3′;
Cancer Susceptibility 9 (CASC9) forward, 5′-CAGGTAATCTCAGCAGTCAT-3′
and reverse, 5′-ACATCCACAGGTCTCCAA-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; and GAPDH forward,
5′-GGAGTCCACTGGCGTCTT-3′ and reverse, 5′-ATCTTGAGGCTGTTGTCATAC-3′.
The following thermocycling conditions were used for qPCR: Initial
denaturation for 15 min at 95°C, followed by 40 cycles of
denaturation for 10 sec at 95°C, annealing for 30 sec at 60°C and
extension for 20 sec at 72°C. Relative expression levels were
calculated using the 2−ΔΔCq method (20) and normalized to the internal
reference genes GAPDH and U6.
Microarray analysis
MCF7 cells were divided into two groups as follows:
One group was treated with 50.00 µg/ml lutein, whereas the other
group was treated with solvent alone (control group). The cells
were incubated for 24 h and total RNA was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The RNA samples were purified using the RNasey Mini kit
(Qiagen, Inc.), amplified and labeled using the Quick Amp Labeling
kit, One-Color (Agilent Technologies, Inc.), according to the
manufacturer's protocol. An equal amount of labeled cRNA from each
sample was hybridized using the Agilent Gene Expression
Hybridization kit (Agilent Technologies, Inc.), followed by image
acquisition and data analysis using the Agilent Feature Extraction
software (version 11.0.1.1; Agilent Technologies, Inc.).
Cell transfection
The sequences of CASC9 small interfering RNA (siRNA)
(si-CASC9), scrambled siRNA [si-negative control (NC)], miR-590-3p
mimic and the NC mimic were purchased from Shanghai GenePharma Co.,
Ltd. The sequences were as follows: si-CASC91#,
5′-AUGAACAUCCACAAACACCAA-3′; si-CASC92#,
5′-UAAUAUUUCUUGAUAGUGCCA-3′; si-CASC9 NC,
5′-GAAUCCUACUUUCACAGCCAU-3′; miRNA-590-3p mimic,
5′-TAATTTTATGTATAAGCTAGT-3′; mimic NC, 5′-CTAGTCACTATATAGGAGCTG-3′.
MCF-7 and T47D cells were cultured until they reached 70–80%
confluence and subsequently each well was transfected with si-CASC9
(45 nM), si-NC (45 nM), miR-590-3p mimic (40 nM) and NC mimic (40
nM) using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions, at
37°C for 48 h. Subsequent experiments were performed 48 h
post-transfection.
Bioinformatics analysis
The DIANA TOOLS database (http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2/index)
was used to determine the molecular mechanism by which CASC9
regulates miR-590-3p.
Dual-luciferase reporter assay
The CASC9 fragments containing the predicted
miR-590-3p binding sites were separately amplified via RT-qPCR
analysis and cloned into a pmirGLO dual luciferase miRNA target
expression vector (Invitrogen; Thermo Fisher Scientific, Inc.) to
create a wild-type CASC9 reporter vector. Subsequently, the
putative mutant miR-590-3p binding sites were constructed in CASC9
by altering the sequences and replacing them to form a
CASC9-mutated-type. The miRNAs (miR-590-3p mimics or miR-NC) and
recombinant plasmids were co-transfected into 293T cells (American
Type Culture Collection), using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.). Following incubation
for 48 h at 37°C, luciferase activities were detected using a dual
luciferase reporter gene assay system (Promega Corporation).
Firefly luciferase activity was normalized to Renilla
luciferase activity.
MTT assay
Following transfection, MCF-7 and T47D cells were
seeded into 96-well microplates at a density of 5×104
cells/ml (100 µl/well) and treated with different concentrations of
lutein (0.00, 6.25, 12.50, 25.00 and 50.00 µg/ml), at 37°C for 24,
48 and 72 h. Subsequently, cells were incubated with 20 µl MTT
solution (KGA312; Nanjing KeyGen Biotech Co., Ltd.) for 4–6 h at
37°C. The MTT solvent included in the assay kit was used to
dissolve the purple formazan. The absorbance was measured at a
wavelength of 570 nm, using a microplate reader (SpectraMax M5;
Molecular Devices, LLC).
Statistical analysis
Statistical analysis was performed using SPSS 22.0
software (IBM Corp.). All experiments were performed in triplicate
and data are presented as the mean ± standard deviation. Unpaired
Student's t-test was used to compare differences between two
groups, while one-way ANOVA followed by Tukey's post hoc test were
used to compare differences between multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Lutein downregulates CASC9 expression
in breast cancer cells
Our previous study demonstrated that lutein exhibits
antitumor effects in breast cancer cells (10). In the present study, to identify
whether specific lncRNAs are involved in the antiproliferative
effects of lutein, their expression levels were detected in MCF7
cells with or without lutein treatment (50.00 µg/ml). A total of
1,083 lncRNAs exhibited significant changes in their expression
levels following treatment of cells with lutein. In the present
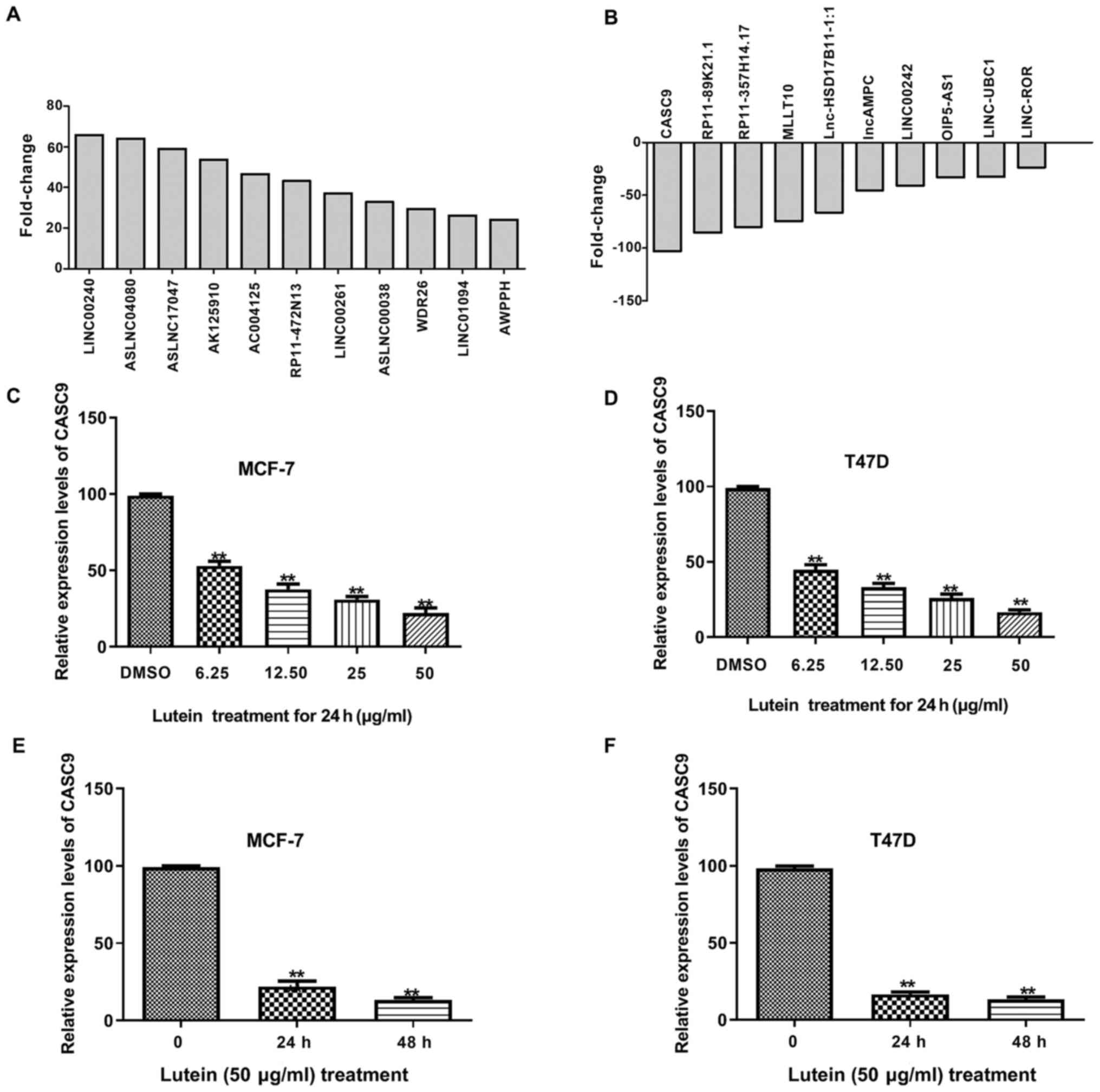
study, 11 lncRNAs (fold change ≥20; P≤0.05; Fig. 1A) were identified with significantly
upregulated expression levels and 10 demonstrated the most
significantly downregulated expression levels (fold change ≤20;
P≤0.05; Fig. 1B). Among these,
LINC00240 was the most highly upregulated lncRNA (fold
change=65.65; P=0.024), whereas CASC9 exhibited the highest
downregulation compared with the expression levels of the remaining
lncRNAs (fold change=103.119800; P=0.033) (Fig. 1A and B). These effects were noted in
MCF7 cells treated with lutein. Thus, RT-qPCR analysis was
performed to detect the expression levels of CASC9 following
treatment of MCF-7 and T47D cells with different concentrations of
lutein for 24 h. The results demonstrated that CASC9 expression
exhibited a negative association with increasing concentrations of
lutein (Fig. 1C and D). In
addition, CASC9 expression was the lowest when the cells were
treated with 50.00 µg/ml lutein. Thus, the present study further
investigated whether CASC9 expression levels decreased in a
time-dependent manner following treatment of cells with 50.00 µg/ml
lutein. The results demonstrated that CASC9 expression decreased in
MCF-7 and T47D cells treated with 50.00 µg/ml lutein in a
time-dependent manner (Fig. 1E and
F). Taken together, these results suggest that lutein
downregulates CASC9 expression in breast cancer cells.
CASC9 knockdown promotes the
suppressive role of lutein on the proliferation of breast cancer
cells
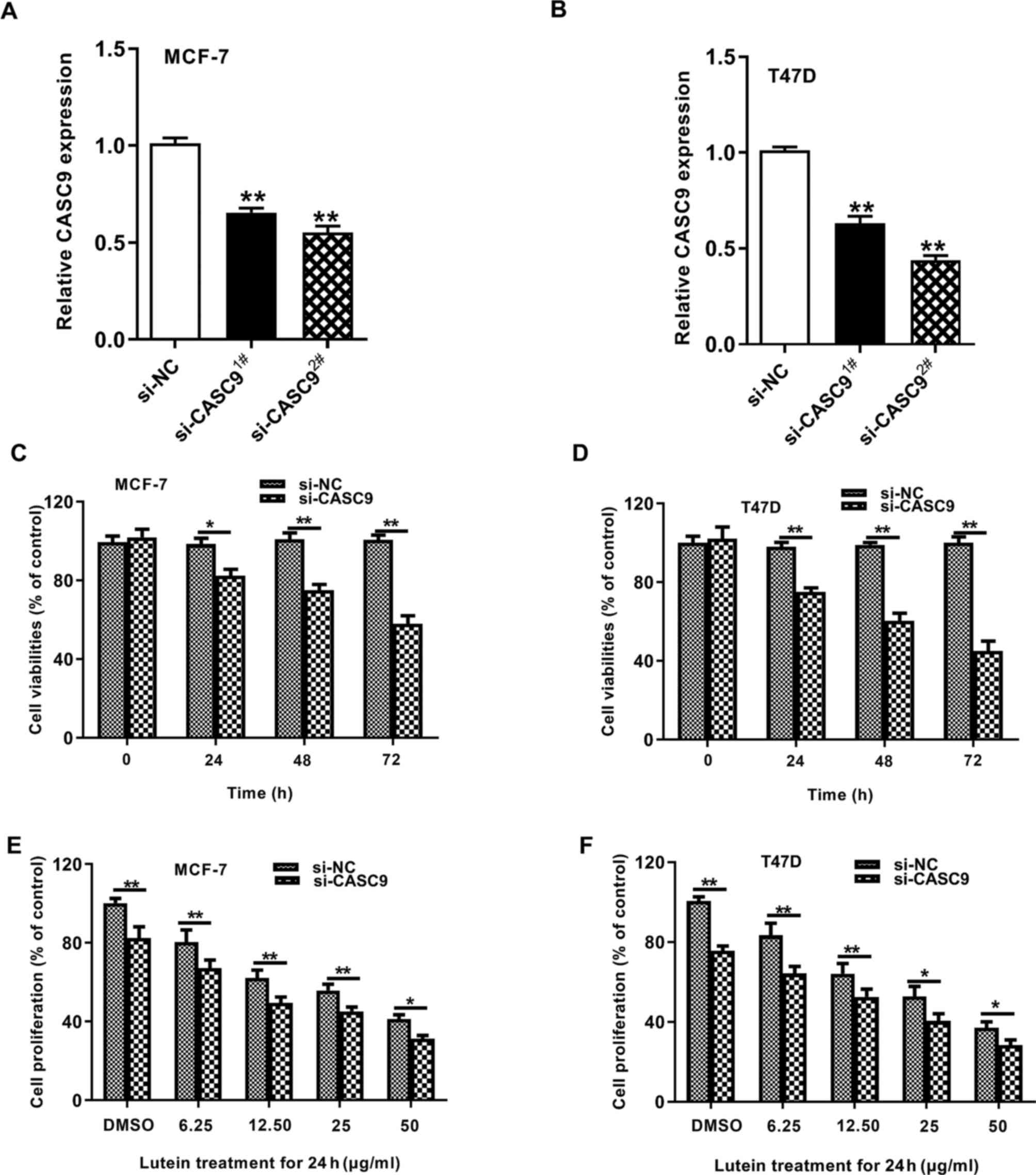
To assess the potential functional roles of CASC9 on
the antiproliferative effects of lutein, CASC9 expression was
knocked down using the siRNA1 and siRNA2 sequences. RT-qPCR
analysis demonstrated that transfection with both siRNAs into the
cells was successful (Fig. 2A and
B). The knockdown efficiency of siRNA2 was higher, and thus
this sequence was selected for subsequent experimentation.
Subsequently, the effect of CASC9 knockdown on cell proliferation
was assessed. The results demonstrated that CASC9 knockdown
inhibited cell proliferation (Fig. 2C
and D). The cells were treated with different concentrations of
lutein and concomitantly transfected with the siRNA sequences to
assess the effect of CASC9 on lutein-mediated tumor suppression.
The results demonstrated that transfection with si-CASC9 sequences
significantly increased the tumor-inhibitory effects of lutein
(Fig. 2E and F). Collectively,
these results suggest that CASC9 knockdown can potentiate the
suppressive role of lutein on breast cancer.
Overexpression of miR-590-3p enhances
the tumor suppressive effect of lutein on breast cancer cells
Recent studies have reported that the
lncRNA/miRNA/mRNA regulatory axis plays important roles in tumor
development (21,22). Thus, the present study aimed to
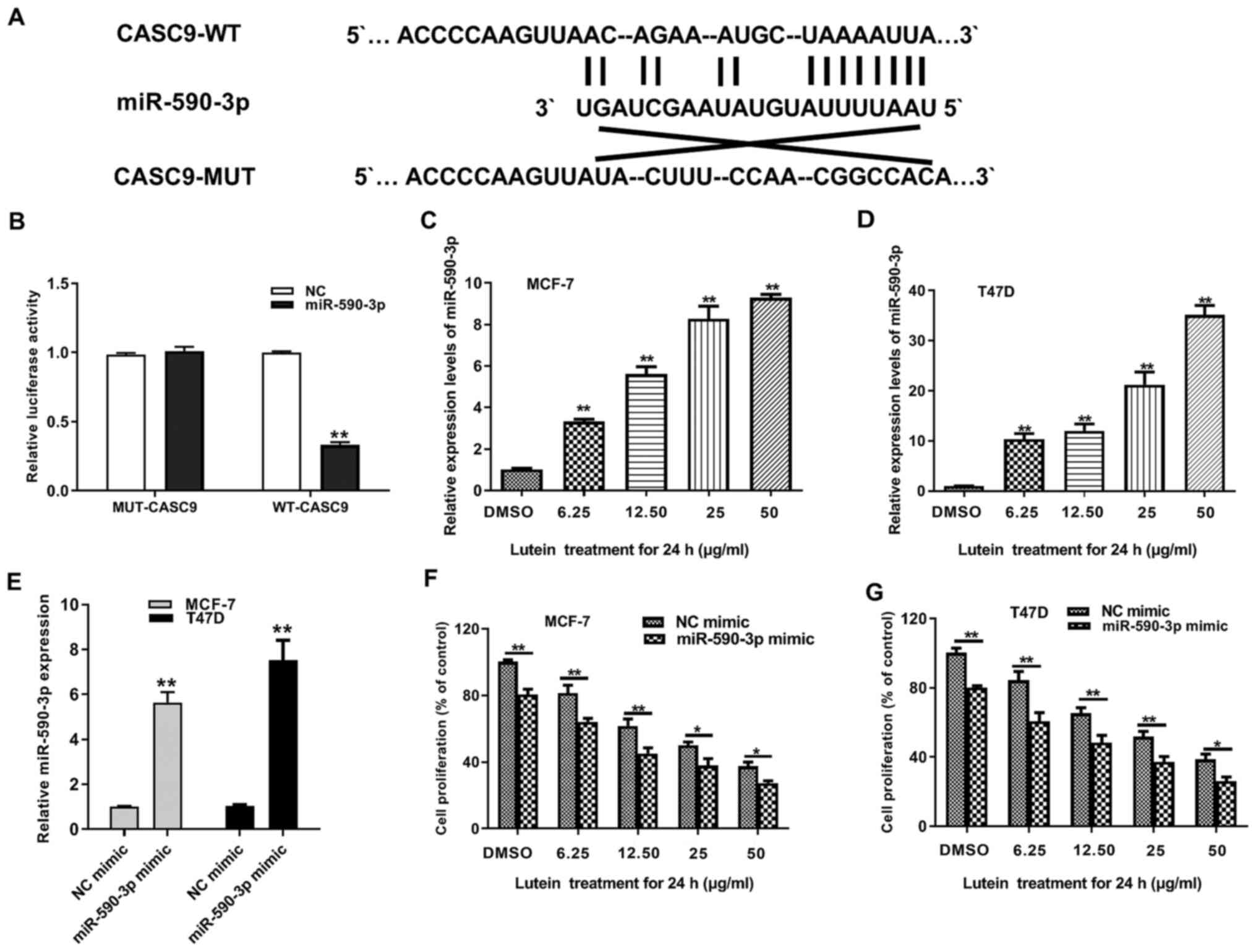
identify the potential targets of CASC9. Bioinformatic analysis
revealed that miR-590-3p is a target gene of CASC9 (Fig. 3A). The dual-luciferase reporter
assay was performed to determine whether CASC9 possesses putative
binding sites of miR-590-3p. Following co-transfection with
miR-590-3p and si-CASC9, the luciferase activity of the wild-type
CASC9 reporter was attenuated (Fig.
3B). Notably, this was not observed in the mutant CASC9
reporter or in the transfected NC-miR 293T cells, suggesting that
miR-590-3p can bind to CASC9. RT-qPCR analysis was performed to
detect miR-590-3p expression following treatment of MCF-7 and T47D
cells with different concentrations of lutein for 24 h. The results
demonstrated that miR-590-3p expression was positively associated
with increasing concentrations of lutein treatment (Fig. 3C and D). miR-590-3p mimics or NC
mimics were transfected into MCF-7 and T47D cells to assess the
potential functional roles of miR-590-3p on lutein-mediated tumor
suppression. RT-qPCR analysis demonstrated that transfection was
successful (Fig. 3E). Subsequently,
the cells were treated with different concentrations of lutein and
concomitantly transfected with miR-590-3p mimics to assess the
effect of miR-590-3p on lutein-mediated tumor suppression. The
results demonstrated that following 24 h treatment of the cells
with lutein, miR-590-3p mimic caused a significant increase in the
tumor-inhibitory effect of this compound (Fig. 3F and G). Taken together, these
results suggest that overexpression of miR-590-3p can enhance the
suppressive role of lutein on breast cancer.
Inhibition of proliferation by lutein
requires the lncRNACASC9/miR-590-3p axis
Taken together, the results of the present study
suggest that CASC9 and miR-590-3p are both involved in the tumor
suppressive role of lutein on breast cancer. Thus, subsequent
experiments were performed to investigate the interaction between
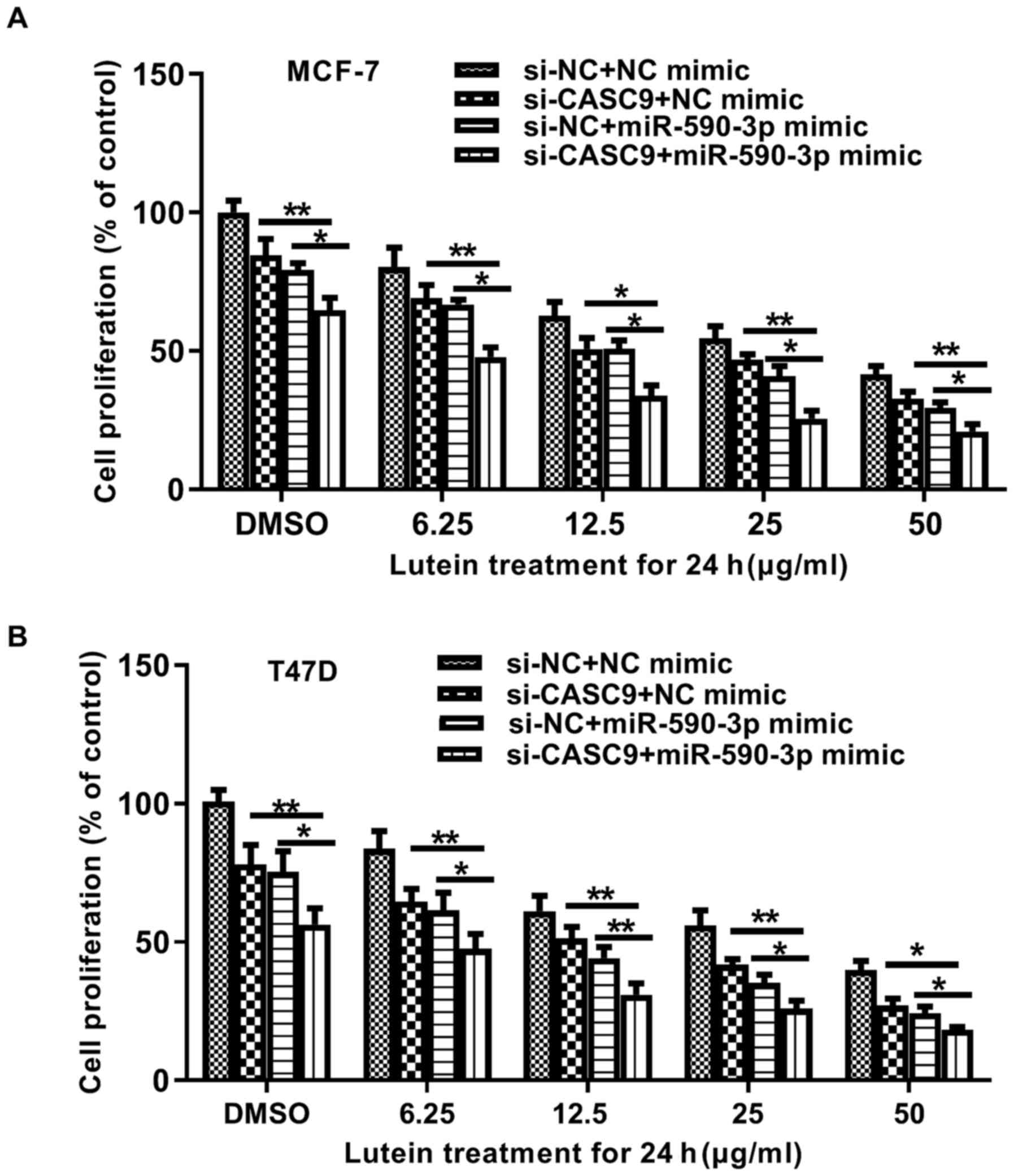
the CASC9/miR-590-3p axis and lutein. MCF-7 and T47D cells were
treated with different concentrations of lutein and transfected
concomitantly with miR-590-3p mimics and si-CASC9. Transfection of
the cells with miR-590-3p mimics or si-CASC9 alone led to a
significant increase in the tumor-inhibitory effects of lutein
(Fig. 4A and B). Notably, the
simultaneous transfection of miR-590-3p mimics and si-CASC9
enhanced the antiproliferative activity of lutein on breast cancer
cells, suggesting that the CASC9/miR-590-3p axis participates in
the antitumor action of this compound.
Discussion
Breast cancer is the most common malignancy in
Chinese women and exhibits the second highest mortality rate among
all female-associated cancer types (23). Over the past decade, the mortality
of breast cancer has declined due to improvements in diagnosis,
surgery and chemotherapy (24).
However, due to heterogeneity and high degree of breast cancer
metastasis, the mortality rate of patients with metastatic breast
cancer remains considerably high (25).
A previous study demonstrated that lutein inhibits
breast cancer cell proliferation by inactivating the NF-κB pathway
via inactivation of the Nrf2/ARE signaling pathway (10). However, to the best of our
knowledge, the ability of lutein to inhibit the proliferation of
breast cancer cells by regulating other associated signaling
transduction pathways has not yet been investigated. The present
study assessed the role of the lncRNA/miRNA axis in lutein-mediated
inhibition of breast cancer cell proliferation. The results
demonstrated that lutein inhibited breast cancer progression by
regulating the CASC9/miR-590-3p axis. To the best of our knowledge,
the present study is the first to demonstrate the involvement of
specific lncRNAs and miRNAs in the antiproliferative action of
lutein on breast cancer cells.
lncRNAs are a class of non-coding RNAs with a length
>200 nucleotides (26). lncRNAs
can regulate the expression levels of specific genes by histone
modification, chromatin remodeling, transcriptional regulation and
mRNA splicing, as well as the regulation of protein activity and
intracellular localization (27).
Recently, lncRNAs, have been demonstrated to regulate certain
cellular functions and participate in the development of specific
diseases, such as Alzheimer's disease, cardiovascular disease, male
infertility, epilepsy and tumor progression (28–32).
It has been reported that several important lncRNA molecules are
involved in the regulation of proliferation and apoptosis of breast
cancer cells, such as H19 imprinted maternally expressed
transcript, urothelial cancer associated 1 and growth arrest
specific 5 (33–35). In addition, neuroblastoma associated
transcript 1 and metastasis associated lung adenocarcinoma
transcript 1 have been demonstrated to be involved in the invasion
and migration of breast cancer cells (36,37).
Furthermore, mitosis associated long intergenic non-coding RNA 1 is
closely associated with drug resistance of breast cancer cells, and
can be used as a sensitizing target for paclitaxel in the treatment
of breast cancer (38). In the
present study, the lncRNA microarray profiles of breast cancer
cells were analyzed to identify candidate lncRNAs associated with
the antiproliferative activity of lutein. Among these, CASC9 was
identified as the most significantly downregulated lncRNA. RT-qPCR
analysis confirmed that lutein inhibited CASC9 expression in MCF-7
and T47D cells, in dose- and time-dependent manners, respectively.
Furthermore, CASC9 knockdown inhibited cell proliferation and
promoted the tumor inhibitory role of lutein on breast cancer.
miRNAs are short RNAs with a length of 20–25
nucleotides (39). They can inhibit
or degrade their target mRNAs by binding to the 3′non-coding region
of their target gene mRNA (18,19).
It has been reported that miRNAs are involved in a series of
important processes, such as tumor proliferation, invasion and
migration (40,41). Increasing evidence suggest that
lncRNAs and miRNAs can interact to cross-regulate the biological
processes of tumor cells (42,43).
In the present study, bioinformatics analysis revealed that
miR-590-3p is a potential target of CASC9. Furthermore, the
dual-luciferase reporter assay demonstrated the direct interaction
between CASC9 and miR-590-3p. RT-qPCR analysis demonstrated that
lutein increased miR-590-3p expression in MCF-7 and T47D cells, in
a dose-dependent manner. Notably, overexpression of miR-590-3p
inhibited cell proliferation and enhanced the antiproliferative
effect of lutein. Furthermore, simultaneous transfection with
miR-590-3p mimics and si-CASC9 potentiated the effect of lutein on
inhibiting the proliferation of breast cancer cells compared with
the effects noted by miR-590-3p or si-CASC9 alone, which suggests
that the CASC9/miR-590-3p axis participates in the
antiproliferative effect of lutein.
The present study is not without limitations. For
example, additional targets of miR-590-3p that may interact with
CASC9, such as SIX homeobox 1, were not assessed. These molecular
targets may crosstalk to regulate the antiproliferative effects of
lutein on breast cancer cells. In addition, the present study only
performed in vitro experiments. Thus, prospective studies
with in vivo experiments are required to validate the
results presented here.
In conclusion, the present study investigated the
changes in the expression levels of specific lncRNAs in breast
cancer cells treated with lutein and verified that CASC9 was the
most significantly downregulated lncRNA. A novel mechanism of
antiproliferative action of lutein was presented, which involves
the activation of the CASC9/miR-590-3p axis. These data may provide
novel targets for the diagnosis and treatment of breast cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the key Science
and Technology project of Henan Province (grant no.
192102310093).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YZ and XY designed the present study. JC, YZ, SZ and
WJ performed the literature review and analyzed the data. YZ and SZ
confirmed the authenticity of all the raw data. All authors have
read and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Benson JR, Jatoi I, Keisch M, Esteva FJ,
Makris A and Jordan VC: Early breast cancer. Lancet. 373:1463–1479.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fan L, Strasser-Weippl K, Li JJ, St Louis
J, Finkelstein DM, Yu KD, Chen WQ, Shao ZM and Goss PE: Breast
cancer in China. Lancet Oncol. 15:e279–e289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Susini T, Olivieri S, Molino C,
Castiglione F, Tavella K and Viligiardi R: Ovarian cancer initially
presenting as intramammary metastases and mimicking a primary
breast carcinoma: A case report and literature review. J Womens
Health (Larchmt). 19:169–174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim HW, Chew BP, Wong TS, Park JS, Weng
BB, Byrne KM, Hayek MG and Reinhart GA: Dietary lutein stimulates
immune response in the canine. Vet Immunol Immunopathol.
74:315–327. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zou Z, Xu X, Huang Y, Xiao X, Ma L, Sun T,
Dong P, Wang X and Lin X: High serum level of lutein may be
protective against early atherosclerosis: The Beijing
atherosclerosis study. Atherosclerosis. 219:789–793. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carpentier S, Knaus M and Suh M:
Associations between lutein, zeaxanthin, and age-related macular
degeneration: An overview. Crit Rev Food Sci. 49:313–326. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Reynoso-Camacho R, González-Jasso E,
Ferriz-Martínez R, Villalón-Corona B, Loarca-Piña GF, Salgado LM
and Ramos-Gomez M: Dietary supplementation of lutein reduces colon
carcinogenesis in DMH-treated rats by modulating K-ras, PκB, and
β-catenin proteins. Nutr Cancer. 63:39–45. 2011.PubMed/NCBI
|
|
9
|
Bharti AC and Aggarwal BB: Chemopreventive
agents induce suppression of nuclear factor-kappaB leading to
chemosensitization. Ann N Y Acad Sci. 973:392–395. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chang J, Zhang Y, Li Y, Lu K, Shen Y, Guo
Y, Qi Q, Wang M and Zhang S: NrF2/ARE and NF-κB pathway regulation
may be the mechanism for lutein inhibition of human breast cancer
cell. Future Oncol. 14:719–726. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
García-Padilla C, Aránega A and Franco D:
The role of long non-coding RNAs in cardiac development and
disease. AIMS Genet. 5:124–140. 2018. View Article : Google Scholar
|
|
12
|
Yu Y, Wang L, Li Z, Zheng Y, Shi Z and
Wang G: Long noncoding RNA CRNDE functions as a diagnostic and
prognostic biomarker in osteosarcoma, as well as promotes its
progression via inhibition of miR-335-3p. J Biochem Mol Toxicol.
35:e227342021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mei J, Hao L, Wang H, Xu R, Liu Y, Zhu Y
and Liu C: Systematic characterization of non-coding RNAs in
triple-negative breast cancer. Cell Prolif. 53:e128012020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Grixti JM and Ayers D: Long noncoding RNAs
and their link to cancer. Noncoding RNA Res. 5:77–82. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ratti M, Lampis A, Ghidini M, Salati M,
Mirchev MB, Valeri N and Hahne JC: MicroRNAs (miRNAs) and long
non-coding RNAs (lncRNAs) as new tools for cancer therapy: First
steps from bench to bedside. Target Oncol. 15:261–278. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guan H, Shang G, Cui Y, Liu J, Sun X, Cao
W, Wang Y and Li Y: Long noncoding RNA APTR contributes to
osteosarcoma progression through repression of miR-132-3p and
upregulation of yes-associated protein 1. J Cell Physiol.
234:8998–9007. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu G, Xue M, Zhao Y, Han Y, Li C, Zhang S,
Zhang J and Xu J: Long noncoding RNA ZEB1-AS1 acts as a Sponge of
miR-141-3p to inhibit cell proliferation in colorectal cancer. Int
J Med Sci. 17:1589–1597. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guan H, Liu J, Lv P, Zhou L, Zhang J and
Cao W: MicroRNA-590 inhibits migration, invasion and
epithelial-to-mesenchymal transition of esophageal squamous cell
carcinoma by targeting low-density lipoprotein receptor-related
protein 6. Oncol Rep. 44:1385–1392. 2020.PubMed/NCBI
|
|
19
|
Sun P, Feng Y, Guo H, Li R, Yu P, Zhou X,
Pan Z, Liang Y, Yu B, Zheng Y, et al: MiR-34a inhibits cell
proliferation and induces apoptosis in human nasopharyngeal
carcinoma by targeting lncRNA MCM3AP-AS1. Cancer Manag Res.
12:4799–4806. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma W, Xue N, Zhang J, Wang D, Yao X, Lin L
and Xu Q: circUBAP2 regulates osteosarcoma progression via the
miR-204-3p/HMGA2 axis. Int J Oncol. 58:298–311. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ma W, Zhao X, Xue N, Gao Y and Xu Q: The
LINC01410/miR-122-5p/NDRG3 axis is involved in the proliferation
and migration of osteosarcoma cells. IUBMB Life. 73:705–717. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guestini F, McNamara KM, Ishida T and
Sasano H: Triple negative breast cancer chemosensitivity and
chemoresistance: Current advances in biomarkers indentification.
Expert Opin Ther Targets. 20:705–720. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Echeverria GV, Powell E, Seth S, Ge Z,
Carugo A, Bristow C, Peoples M, Robinson F, Qiu H, Shao J, et al:
High-resolution clonal mapping of multi-organ metastasis in triple
negative breast cancer. Nat Commun. 9:5079–5095. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiong H, Shen J, Chen Z, Yang J, Xie B,
Jia Y, Jayasinghe U, Wang J, Zhao W, Xie S, et al: H19/let 7/Lin28
ceRNA network mediates autophagy inhibiting epithelial mesenchymal
transition in breast cancer. Int J Oncol. 56:794–806.
2020.PubMed/NCBI
|
|
27
|
Scacalossi KR, van Solingen C and Moore
KJ: Long non-coding RNAs regulating macrophage functions in
homeostasis and disease. Vascul Pharmacol. 114:122–130. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu KS, Li TP, Ton H, Mao XD and Chen YJ:
Advances of long noncoding RNAs-mediated regulation in
reproduction. Chin Med J (Engl). 131:226–234. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cortini F, Roma F and Villa C: Emerging
roles of long non-coding RNAs in the pathogenesis of Alzheimer's
disease. Ageing Res Rev. 50:19–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schulte C, Barwari T, Joshi A, Zeller T
and Mayr M: Noncoding RNAs versus protein biomarkers in
cardiovascular disease. Trends Mol Med. 26:583–596. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Su Y, Zhou LL, Zhang YQ and Ni LY: Long
noncoding RNA HOTTIP is associated with male infertility and
promotes testicular embryonal carcinoma cell proliferation. Mol
Genet Genomic Med. 7:e8702019. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu Q, Zhao MW and Yang P: LncRNA UCA1
suppresses the inflammation via modulating miR-203-mediated
regulation of MEF2C/NF-κB signaling pathway in epilepsy. Neurochem
Res. 45:783–795. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guan H, Mei Y, Mi Y, Li C, Sun X, Zhao X,
Liu J, Cao W, Li Y and Wang Y: Downregulation of lncRNA ANRIL
suppresses growth and metastasis in human osteosarcoma cells. Onco
Targets Ther. 11:4893–4899. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang J, Sun J and Yang F: The role of long
non-coding RNA H19 in breast cancer. Oncol Lett. 19:7–16.
2020.PubMed/NCBI
|
|
35
|
Liu C, Jiang F, Zhang X and Xu X: Long
non-coding RNA UCA1 modulates paclitaxel resistance in breast
cancer via miR-613/CDK12 Axis. Cancer Manag Res. 12:2777–2788.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li Y, Guo XB and Wei YH: LncRNA GAS5
affects epithelial-mesenchymal transition and invasion of breast
cancer cells by regulating miR-216b. Eur Rev Med Pharmacol Sci.
24:4873–4881. 2020.PubMed/NCBI
|
|
37
|
Hu P, Chu J, Wu Y, Sun L, Lv X, Zhu Y, Li
J, Guo Q, Gong C, Liu B and Su S: NBAT1 suppresses breast cancer
metastasis by regulating DKK1 via PRC2. Oncotarget. 6:32410–23425.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang Y, Zhou Y, Yang Z, Chen B, Huang W,
Liu Y and Zhang Y: MiR-204/ZEB2 axis functions as key mediator for
MALAT1-induced epithelial-mesenchymal transition in breast cancer.
Tumour Biol. 39:10104283176909982017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bida O, Gidoni M, Ideses D, Efroni S and
Ginsberg D: A novel mitosis-associated lncRNA, MA-linc1, is
required for cell cycle progression and sensitizes cancer cells to
Paclitaxel. Oncotarget. 6:27880–27890. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wei F, Yang S and Wang S: MicroRNAs: A
critical regulator under mechanical force. Histol Histopathol.
33:335–342. 2018.PubMed/NCBI
|
|
41
|
Xie T, Wu D, Li S, Li X, Wang L, Lu Y,
Song Q, Sun X and Wang X: microRNA-582 potentiates liver and lung
metastasis of gastric carcinoma cells through the FOXO3-mediated
PI3K/Akt/Snail pathway. Cancer Manag Res. 12:5201–5212. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen ZF, Wang J, Yu Y and Wei W:
MicroRNA-936 promotes proliferation and invasion of gastric cancer
cells by down-regulating FGF2 expression and activating P13K/Akt
signaling pathway. Eur Rev Med Pharmacol Sci. 24:6707–6715.
2020.PubMed/NCBI
|
|
43
|
Jiang Q, Xing W, Cheng J and Yu Y:
Knockdown of lncRNA XIST suppresses cell tumorigenicity in human
non-small cell lung cancer by regulating miR-142-5p/PAX6 axis. Onco
Targets Ther. 13:4919–4929. 2020. View Article : Google Scholar : PubMed/NCBI
|