Introduction
Doxorubicin (Dox) belongs to the anthracycline
family of antibiotics and is widely used for the treatment of
several types of cancer, including gastric cancer, breast cancer,
leukemia, sarcoma and lymphoma (1).
Dox efficiently induces apoptosis of tumor cells and prevents cell
proliferation by intercalating into DNA and stabilizing a ternary
complex with topoisomerase II to interfere with DNA replication
(1). The use of Dox is limited due
to the development of cardiotoxicity; the risk of heart disease is
3–5% in patients who receive a cumulative dose of 400
mg/m2 and up to 48% in patients receiving 700
mg/m2 (2). A number of
studies have demonstrated that Dox is a mitochondrial toxin
(1,2). Dox induces over-production of reactive
oxygen species (ROS) and ATP imbalance through electron transport
chain uncoupling, resulting in cardiac injury (1).
MicroRNAs (miRNAs/miRs) are a class of small
non-coding single-stranded RNAs (~22 nucleotides in length) that
bind to complementary sequences found in the 3′-untranslated region
of mRNAs and regulate post-transcriptional gene expression by
inducing mRNA degradation or inhibiting mRNA translation (3). miRNAs are ubiquitously involved in
developmental and pathological processes, such as cell
differentiation (4), organogenesis
(5), tissue injury and remodeling
(6), and tumorigenesis. Increasing
data have revealed that miRNAs serve an important role in cardiac
functions and several cardiovascular diseases, such as ischemic
heart diseases and arrhythmia (7,8). Several
biological and pathological functions of miRNAs in the regulation
of cardiomyocyte survival or apoptosis have been directly confirmed
in animal or cell models (9,10). In recent years, miRNA expression
profiling of heart tissues in mouse or rat models of Dox-induced
cardiotoxicity have been investigated by miRNA array or
quantitative PCR assay, and numerous changes in miRNA expression
were reported 2–8 weeks after Dox treatment (11,12). It
is well known that the transcription of miRNAs is sensitive to
cellular stress and triggers a number of downstream biological
processes (13). Changes in miRNA
expression profiling in cardiomyocytes resulting from Dox at the
early stage (12 or 24 h post Dox treatment) remain unknown. In the
present study, the rat cardiomyocyte H9c2 cell line was used to
investigate changes in miRNA expression at the early stage of Dox
treatment and the contribution of miRNAs in the initial phase of
Dox-induced cardiomyocytes dysfunction.
Materials and methods
Cell culture and treatment
Rat H9c2 cardiomyocytes were purchased from The Cell
Bank of Type Culture Collection of the Chinese Academy of Sciences.
Cells were maintained in DMEM containing 10% FBS (both Gibco;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin (Thermo Fisher Scientific, Inc.) at 37°C in a
humidified incubator containing 5% CO2. Dox (Zhejiang
Hisun Chemical Co., Ltd.) was obtained from Tongren Hospital
(Shanghai, China). Cells were seeded in a 15-cm culture dish at a
density of 1×106. Dox was dissolved in normal saline.
Cells were treated with a final concentration of 5 µg/ml Dox for 0,
12 or 24 h at 37°C in a humidified incubator containing 5%
CO2, and then samples were collected. The morphology of
H9c2 cells was observed at the aforementioned time points under a
light microscope (Nikon Eclipse Ti-S; Nikon Corporation;
magnification, ×10) (Fig. S1).
RNA isolation, RNA-sequencing library
construction and sequencing
Total RNA was extracted from cells using
TRIzol® according to the manufacturer's instructions
(cat. no. 15596026; Thermo Fisher Scientific, Inc.). A total of 3
µg total RNA per sample was used to generate the small RNA library.
Sequencing libraries were prepared using TruSeq® Small
RNA Sample Prep kit following the manufacturer's protocol (cat. no.
RS-200-0012; Illumina, Inc.). The library quality was examined
using Agilent Bioanalyzer 2100 (Agilent Technologies, Inc.).
NEBNext Multiplex Small RNA Library Prep Set for Illumina was used
for sequencing (cat. no. E7300S; New England BioLabs, Inc.). Deep
sequencing of the library was performed on Illumina NextSeq
platform using single-end 1×75 sequencing mode.
Quality control and alignment of
sequencing data
Raw reads were filtered to obtain high-quality clean
reads by removing sequencing adapters, short reads (<35 bp in
length) and low-quality reads using Cutadapt v1.9.1 (14) and Trimmomatic v0.35 (14). Subsequently, FastQC v0.11.5 (15) was used for ensuring high quality
reads. The resulting clean reads were mapped to rat genome
(assembly Rnor_6.0) using bowtie2 v2.2.9 with default parameters
(16). Using miRBase as the
reference database (http://www.mirbase.org), reads of each miRNA were
determined, and reads per million (RPM) was adopted as the
normalization method to quantify the abundance of each miRNA.
Analysis of differential
expression
Differential miRNA expression analyses were
conducted using DESeq2 v1.20.0 (17). The false discovery rate control
method was used to calculate the adjusted P-values in multiple
testing, in order to determine the significance of the differences.
Genes with an adjusted P<0.05, regression to the mean value of
10 at the 0-h time point and an expression level varying ≥1.5-fold
at 24 vs. 0 h or 12 vs. 0 h were used for subsequent analysis. The
cluster analysis of differentially expressed miRNAs was performed
using the Hierarchical Clustering Explorer v3.5 software
(http://www.cs.umd.edu/hcil/hce/hce3.html).
Functional enrichment and target genes
interaction network analysis
The target genes of rat miRNAs were downloaded from
mirTarBase v7.0 (10), the online
miRNA reference database that stores manually curated collections
of experimentally validated miRNA targets. Enrichment analysis
based on Gene Ontology (GO; http://geneontology.org/) annotation and Kyoto
Encyclopedia of Genes and Genomes (KEGG; http://www.genome.jp/kegg/pathway.html) pathways for
the target genes of differentially expressed miRNAs was implemented
using the ClusterProfiler package v3.11.0 (7). A pathway (or GO term) with an adjusted
P<0.05 was considered to be a significantly enriched pathway (or
GO term). Based on the lists of differentially expressed miRNAs,
information on the target gene interactions was obtained from the
Search Tool for the Retrieval of Interacting Genes/Proteins
(www.string-db.com). Target gene
interaction networks were mapped using Cytoscape v3.7 software
(https://cytoscape.org/index.html).
Reverse transcription-quantitative
(q)PCR analysis for miRNA expression
Total RNA was extracted from cells using
TRIzol® (cat. no. 15596026; Thermo Fisher Scientific,
Inc.). miRNA first-strand cDNA was synthesized from 1 µg total RNA
from cells using the PrimeScript™ II 1st Strand cDNA Synthesis kit
(Takara Bio, Inc.). The 20-µl volume reaction system contained 1 µg
total RNA, 5 nM reverse transcription primer (Stem-loop RT primer
from Shanghai GenePharma Co., Ltd.), 20 U reverse transcriptase, 20
U RNase inhibitor and 0.2 mM dNTPs. The mixture was incubated at
42°C for 15 min and at 85°C for 5 min. qPCR was performed with the
SYBR Green Hairpin-it MicroRNA Quantitation PCR kit (cat. no.
E01006; Shanghai GenePharma Co., Ltd.) according to the
manufacturer's instructions. cDNA (2 µl) was used as the template
for the qPCR reaction system. PCR was performed using the
StepOnePlus PCR system (Thermo Fisher Scientific, Inc.), and the
PCR parameters were as follows: 95°C for 3 min, followed by 40
cycles at 95°C for 12 sec and 62°C for 40 sec. Rox was used as the
reference dye in PCR. The expression levels of rno-miR-29b-3p,
rno-miR-145-5p and rno-miR-378a-3p were analyzed using the 7500
real-time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The primers were included in the aforementioned SYBR Green
Hairpin-it MicroRNA Quantitation PCR kit. Primers used were as
follows: Rno-mir-29b-3p forward, 5′-ACAGCAATTAGCACCATTTGAA-3′ and
reverse, 5′-TATGCTTCTTCTCGTCTCTGTGTC-3′; rno-mir-145-5p forward,
5′-CAGTCTTGTCCAGTTTTCCCAG-3′ and reverse,
5′-TATGCTTGTTCTCGTCTCTGTGTC-3′; rno-mir-378a-3p forward,
5′-ATGGTGGACTGGACTTGGAGT-3′, and reverse, 5′-GTGCAGGGTCCGAGGT-3′;
rat U6 forward, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′. Relative miRNA expression was
normalized to U6 expression and calculated using the relative
2−ΔΔCq method (18).
Statistical analysis
Statistical analysis was performed using SPSS v21
(IBM Corp.) and data were analyzed using one-way ANOVA followed by
Tukey's HSD test. All data were presented as the mean ± SD. All
experiments were repeated at least three times. P<0.05 was
considered to indicate a statistically significant difference.
Results
Dynamic changes in miRNA expression
profiling in cardiomyocytes treated with Dox
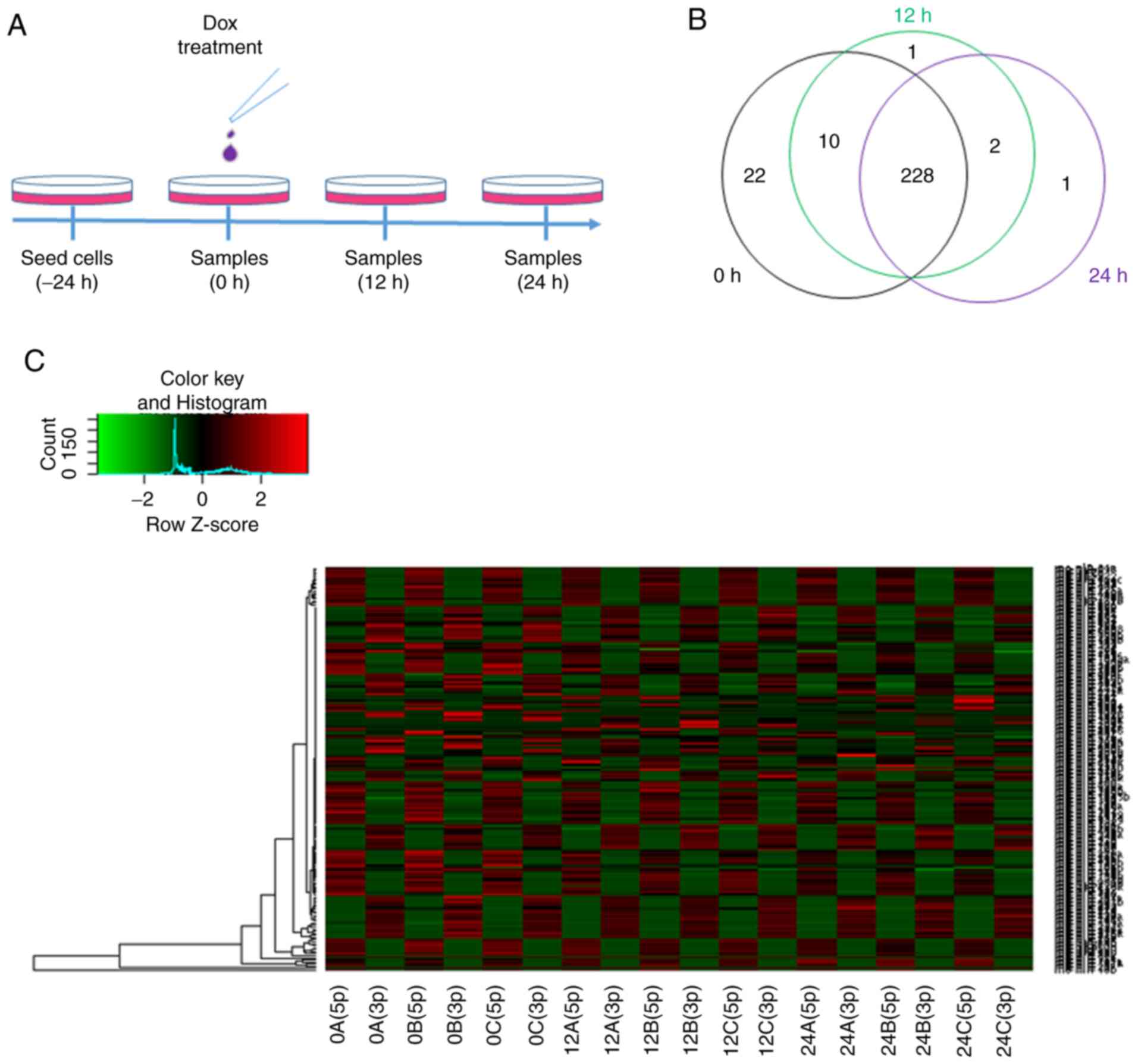
In order to reveal the dynamic changes in miRNAs at
the early stage of Dox treatment, deep miRNA sequencing was
performed to determine miRNA expression at the 0-, 12- and 24-h
time points (Fig. 1A). Read counts
of each miRNA were normalized to RPM, and the expression levels of
386 rat miRNAs were identified to be altered after 12 or 24 h of
Dox treatment compared with the 0-h time point (Tables SI and SII). In the present study, 228 miRNAs were
expressed continuously at the 0-, 12- and 24-h time points by
intersecting miRNAs (Fig. 1B).
Mature miRNAs can be generated from the 5p- or 3p-arm of miRNA
precursors, and the majority of miRNA isoform accumulation is
tissue-dependent (19). A total of
198 pairs of miRNAs were analyzed, and 5p- or 3p-biased miRNA
expression was presented at different time points (Fig. 1C). The results revealed that Dox
decreased the expression levels of most miRNAs; however, the
efficiency of regulation was different between the 5p and 3p
isoforms in each pair of miRNAs. The mechanism of 5p and 3p
selectivity and accumulated expression remains unknown.
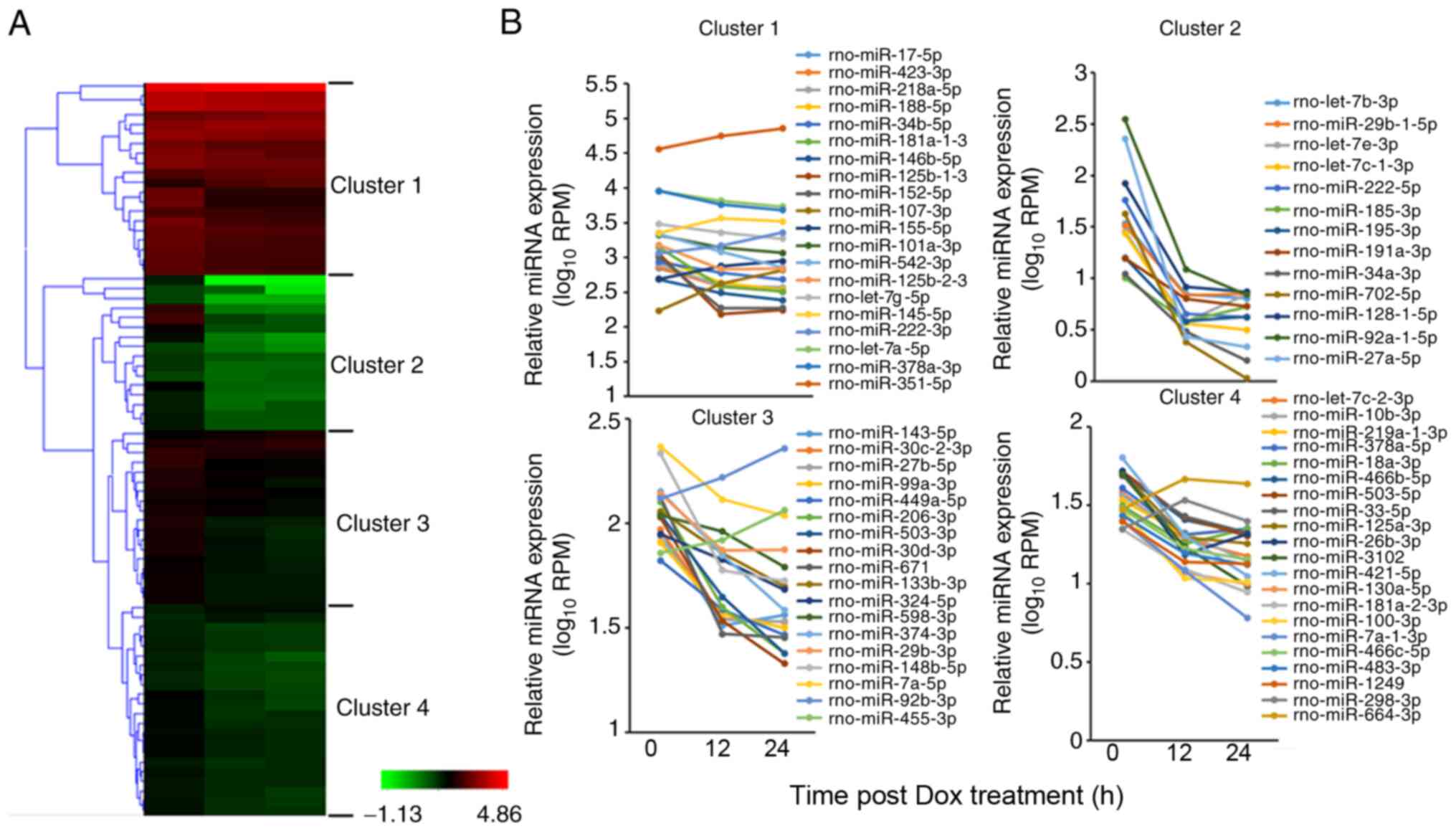
In order to narrow down the target miRNAs, 76 miRNAs
whose expression levels changed by ≥1.5-fold compared with the 0-h
time point were selected, including 67 downregulated miRNAs and 9
upregulated miRNAs (Table SIII),
and were further classified into 4 clusters using the Hierarchical
Clustering Explorer v3.5 software (Fig.
2A). Changes in miRNA expression patterns for 4 typical
clusters are shown in Fig. 2B.
miRNAs in cluster 1 had high expression levels, with average RPM
values >100 at the 0-h time point in H9c2 cells. Notably,
miR-351-5p exhibited the highest RPM value in cluster 1. The
expression levels of miRNAs of cluster 2 rapidly decreased at both
12 and 24 h after Dox treatment. The expression levels of miRNAs of
cluster 3 and cluster 4 tended to decrease following Dox treatment,
although the range of change was smaller compared with that in
cluster 2. The expression levels of most miRNAs in the 4 clusters
were downregulated at 12 and 24 h after Dox administration. Among
the selected miRNAs, some miRNAs are known to function in
Dox-induced heart diseases, such as members of the miR-30 family or
let-7 family, miR-133b, miR-143, miR-298, miR-29 and miR-34a
(6–13). However, the functions of most of the
selected miRNAs in Dox-induced cardiotoxicity or heart injury
remain unclear.
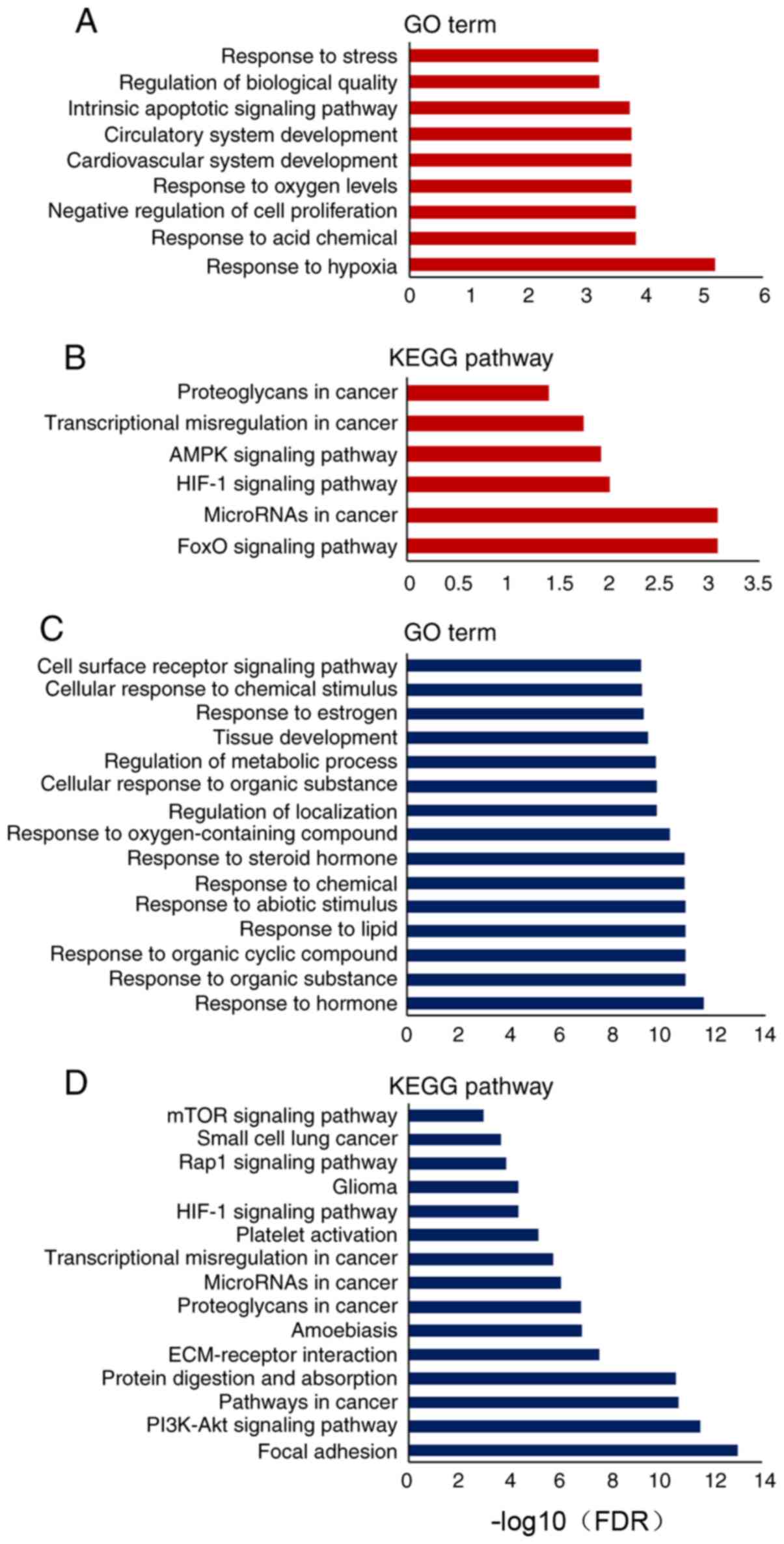
Contribution of Dox-mediated miRNA
expression to different biological processes and pathways
miRNAs directly bind to target mRNAs to induce mRNA
degradation or inhibit mRNA translation in cells. Based on the
mirTarBase database of differentially expressed miRNAs, information
on the downstream targets of upregulated or downregulated miRNAs
was collected. A total of 87 genes targeted by the downregulated
miRNAs, and 20 genes targeted by the upregulated miRNAs were
analyzed according to GO terms and KEGG pathways (Tables SIV–SVII). The upregulated miRNAs contributed
to the biological processes of ‘response to hypoxia’, ‘negative
regulation of cell proliferation’ and ‘cardiovascular system
development’ (Fig. 3A). KEGG pathway
analysis revealed that the upregulated miRNAs were involved in the
regulation of cell survival and apoptotic pathways, such as the
‘hypoxia-inducible factor 1 (HIF-1) signaling pathway’, ‘AMPK
signaling pathway’ and ‘FoxO signaling pathway’ (Fig. 3B). Notably, the analysis of
downregulated miRNAs revealed that several processes of cell
response to chemicals were disturbed, such as ‘response to organic
substance’, ‘response to lipid’ and ‘response to steroid hormone’
(Fig. 3C). Additionally, these
downregulated miRNAs were involved in ‘extracellular matrix
(ECM)-receptor interaction’, ‘focal adhesion’ and ‘PI3K-Akt
signaling pathway’ (Fig. 3D).
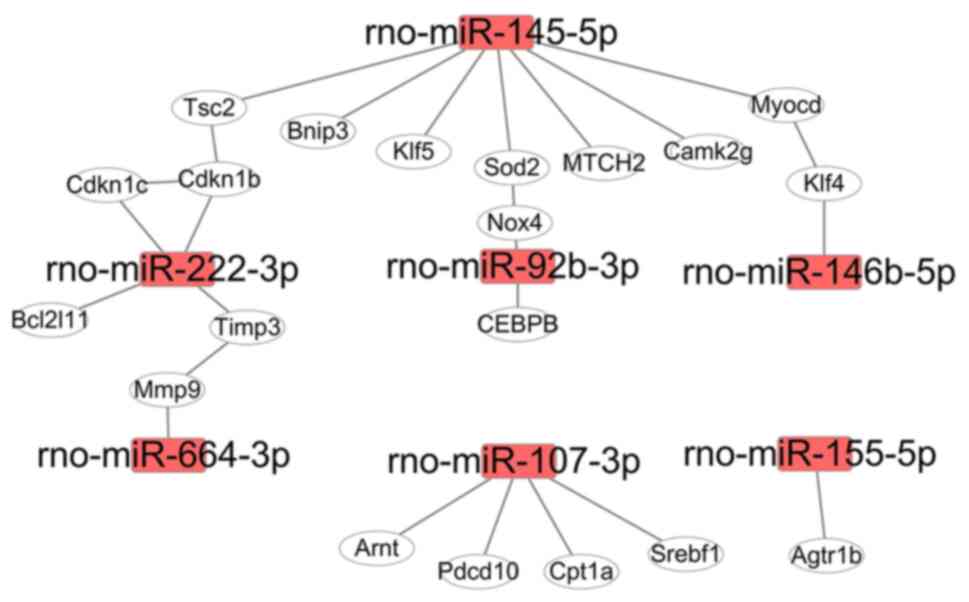
Regulation of gene networks by miRNAs
in H9c2 cells
Proteins, as products of gene translation, execute
biological functions in cells. Most proteins need be associated
with their partners to form a protein complex, which then has the
capability to be involved in biological functions (20). Differentially expressed miRNAs
induced by Dox in the present study regulated a number of target
genes, and the proteins translated from these genes were used for
mapping protein-protein interaction networks. For the upregulated
miRNAs (miR-455-3p, −92b-3p, −222-3p, −155-5p, −298-3p, −351-5p,
664-3p, −145p-5p and −107-3p), the interaction network consisted of
20 genes (Fig. 4), which were
involved in the regulation of cell fate and stress response
pathways (Table SIV). For example,
rno-miR-145-5p targeted 7 genes in rats (Fig. 4), and these genes were involved in
the regulation of autophagy, lipid metabolism and mitochondrial
homeostasis (Table SIV). The
function of rno-miR-222-3p in heart disease or cardiomyocytes has
been previously reported (13);
however, its expression profiling during Dox treatment was newly
discovered (Fig. 4). rno-miR-92b-3p
expression was significantly increased at 12 and 24 h after Dox
treatment in H9c2 cells, and it targeted NADPH oxidase 4 (Nox4),
whose expression levels and mRNA alternative splicing induce the
development of heart failure by increasing ROS production (3).
In the downregulated miRNA group, the range of
fold-change was 1.76–384.67 at 12 or 24 h vs. the 0-h time point
(Table SII), and the network
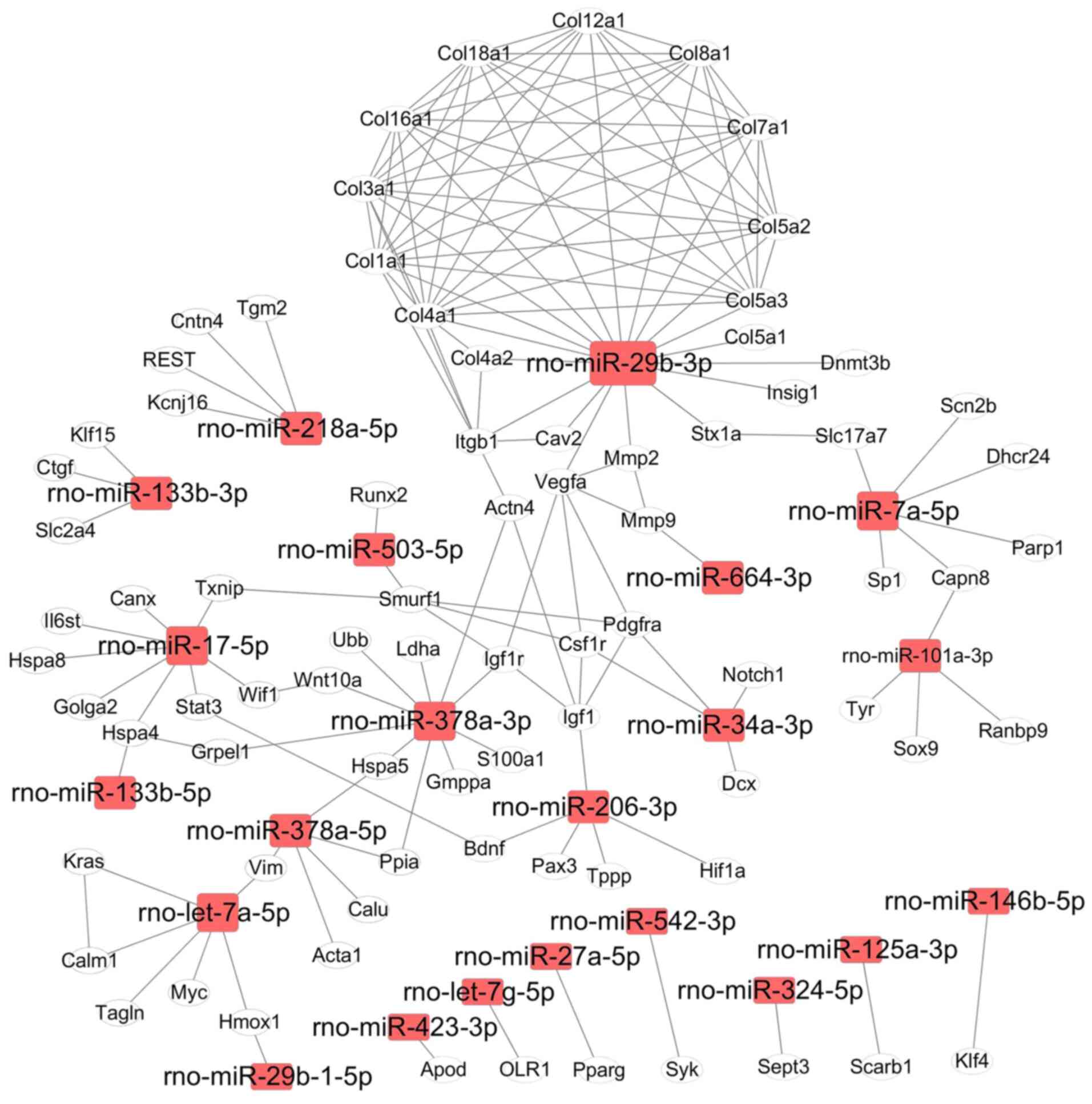
comprised a total of 178 interactions between genes (Fig. 5). According to the number of targets
genes and biological functions, four miRNAs (rno-miR-29b-3p,
rno-miR-378a-3p, rno-let-7a-5p and rno-miR-17-5p) were localized on
critical network nodes and regulated over 40 genes (Fig. 5). It is well know that heat shock
proteins (Hsp) are important for regulating cell survival and
protein homeostasis in cells. Notably, rno-miR-378a-5p targeted
Hspa5, and rno-miR-17-5p targeted Hspa4 and Hspa8 (Fig. 5). In the current map, rno-let-7a-5p
regulated the expression of 6 genes, whose functions are associated
with maintaining cell phenotype (such as Myc, Kras and Tagln) and
functions (such as Hmox1, Calm1 and Vim) (21). rno-miR-29b-3p expression was
significantly downregulated by Dox in H9c2 cells, and it interfered
with the expression levels of 10 genes in the collagen protein
family, which are important proteins in the ECM. It is well know
that the ECM serves a critical role in signal transduction and
material exchange between the environment and cells. Three miRNAs,
namely rno-miR-145-5p (upregulated), rno-miR-29b-3p and
rno-miR-378a-3p (both downregulated), served as important nodes in
the gene networks, and their expression levels were therefore
further analyzed using qPCR. The expression levels of these three
miRNAs were consistent with the results from deep sequencing
analysis, with the expression levels of rno-miR-145-5p being
significantly upregulated and those of rno-miR-29b-3p and
rno-miR-378a-3p being significantly downregulated following Dox
treatment for 12 and 24 h (Fig.
S2).
Discussion
Cardiotoxicity is the main side effect of Dox
treatment that restricts its clinical application in cancer therapy
(1). Specific biomarkers at the
early stage of cardiotoxicity resulting from Dox treatment are very
important to predict and decrease the risk of heart failure in the
clinic. Previous studies have revealed that circulating miRNAs
(8,9), as potential biomarkers of heart
failure, were very sensitive in patients with heart failure
(13). In the present study, 76
miRNAs were identified to be dysregulated by Dox treatment in H9c2
myoblast cells. In the upregulated miRNA group, the expression
levels of 9 miRNAs (miR-455-3p, −92b-3p, −222-3p, −155-5p, −298-3p,
−351-5p, 664-3p, −145p-5p and −107-3p) were significantly
upregulated at 12 or 24 h after Dox treatment. Altered expression
levels of miR-145-5p, miR-155-5p and miR-222-3p have been
previously observed in animal models of Dox administration
(22–24). In particular, miR-145-5p has been
previously reported as a cardioprotective molecule in myocardial
ischemic injury (25) and as a
biomarker in LaminA/C-related dilated cardiomyopathy (26). miR-351-5p exhibited the highest RPM
value in this group, and has been shown to inhibit Pten expression
in H9c2 cells (27). It is well
known that PTEN serves an important role in the development of
heart failure by regulating multiple signaling pathways (28,29).
Additionally, miR-351-5p expression is upregulated by ROS and
functions as a proinflammatory and proapoptotic factor by targeting
the MAPK signaling pathway in both ICE-6 and H9c2 cells (30,31).
Furthermore, several studies have identified that MAPK/NF-κB, a
classical proinflammatory and proapoptotic signaling pathway, is
involved in cardiotoxicity induced by Dox (32,33).
In the downregulated miRNA group, two out of the ten
most altered miRNAs, miR-133b (34)
and let-7e-3p (23), have been
reported in previous studies, revealing a minor increase in
circulating miR-133b expression and a dominant contribution from
the skeletal muscle toxicity induced by Dox (34,35). In
the present study, miR-133b-5p expression in H9c2 cells was
inhibited by Dox and markedly decreased. The main side effect of
Dox are both acute or chronic heart failure, whose progression
begins from myocardial injury that develops into left ventricular
dysfunction (36). These
pathological changes are very important for the progression to
heart failure. miR-133 expression in the heart functions as a
suppressor of cardiac fibrosis and cardiac remodeling (10).
miR-29b-3p expression in the present study was
markedly downregulated after Dox administration, in accordance with
previous studies (13,22,23).
miR-29b-3p targeted 12 collagen genes, whose protein levels have
been detected and used as prognostic biomarkers in patients with
chronic heart failure (37,38). Additionally, inhibition of miR-29b-3p
expression directly results in apoptosis of myoblast cells in
vitro (39). Recent studies have
indicated that miR-29b-3p serves a central role in the processes of
cardiac fibrosis formation and adverse ventricular remodeling
(40,41). Both cell proliferation and apoptosis
are involved in the process of tissue remodeling. In the present
study, miR-702-5p and miR-128-1-5p expression was inhibited by Dox
in H9c2 cells. In previous studies, miR-702 has been shown to
inhibit apoptosis and improve survival of different types of cells,
such as NIH3T3, 293T and embryonic stem cells (42,43).
miR-128 serves a critical role in promoting cardiomyocyte
proliferation and heart regeneration by targeting Suz12, a
component of polycomb repressive complex 2, and downregulating
Suz12 protein expression in cardiomyocytes (44).
The members of the let-7 miRNA family are involved
in the regulation of cardiomyocyte differentiation and metabolism,
as well as myocardial infarction (45). In the present study, the expression
levels of seven members of let-7 (rno-let-7a-1-3p, -a-5p, -b-3p,
-c-1-3p, -c-2-3p, -e-3p and -g-5p) were inhibited by Dox in H9c2
cells, and, to the best of our knowledge, the expression profiles
of six let-7 members in Dox induced cardiomyocytes have not been
previously reported, except for let-7g (9). In previous studies, both let-7a and −7e
downregulated β1-adrenoceptors expression and modulated their
downstream signaling pathways in cardiomyocytes of rat heart
failure models (46,47). Inhibition of let-7a and −7e
expression significantly results in deterioration of cardiac
fibrosis and heart failure (46).
However, data on the let-7 miRNA family in regulating heart failure
are not consistent across different studies and their biological
functions in cardiomyocytes need to be further investigated
(48,49). miR-378 improves cardiomyocyte
survival (50) and protects
cardiomyocytes against myocardial fibrosis in vivo (51). In the present study, Dox repressed
miR-378-3p and −5p expression in H9c2 cells at the 12- and 24-h
time points. In vivo and in vitro studies have
confirmed that miR-378 directly targets caspase-3 (50) and four key components of the MAPK
signaling pathway (Mapk1, Igfr1, Gfrbp 2 and Ksr1) (52).
H9c2 rat cardiac cells, as immortalized
cardiomyocytes, are widely used in establishing cell models of
cardiomyocyte injury induced by different compounds in the medium
with or without the addition of serum (32,53).
Previous studies have indicated that serum withdrawal can
significantly induce changes in miRNA expression in HUVECs and A549
cells (54,55). In the present study, cells were
treated with Dox in complete medium with serum, since serum
deprivation could have induced disturbances of miRNA expression
associated with apoptosis. There is a limitation in the present
study, as the effects of cell proliferation and cell cycle on miRNA
expression in H9c2 cells could not be avoided. To further
investigate the impacts of serum on miRNA expression, miRNA
expression profiles should be analyzed after treating H9c2 cells
with Dox in medium with or without serum in future studies.
In conclusion, 76 miRNAs were identified to have
altered expression levels directly induced by Dox in a
cardiomyocyte cell line. Notably, to the best of our knowledge, the
present study described for the first time the altered expression
levels of miRNAs in cardiomyocytes treated with Dox, including
let-7 family, miR-664-3p, −455-3p, −298-3p, −702-5p, −128-1-5p,
−671, −421-5p, −378-3/5p and −351-3p. Pathway analysis revealed
that these miRNAs were involved in the dysregulation of multiple
signaling pathways, including the FoxO, HIF-1, AMPK, focal
adhesion, PI3K-Akt, ECM-receptor and mTOR signaling pathways. The
expression levels of these miRNAs served an important role in the
regulation of biological processes that support cell metabolism and
cell stability. Accordingly, previous studies have reported that
the MAPK and PI3K-Akt signaling pathways serve an important role in
Dox-induced cardiotoxicity (53,56).
However, the present study presents some limitations, since the
direct cardiotoxicity of Dox was only investigated in vitro.
Dox and products of Dox metabolism may serve a role in these
biological processes in vivo. The control group always
serves an important role in experiments, and the 0-h time point
group (untreated group) was chosen as a control in the present
study. However, H9c2 cells treated with a vehicle for 0, 12 and 24
h could also be used as experiment controls. Additionally, using
different doses of Dox on H9c2 cells may exclude effects of vehicle
on miRNA expression. These strategies may be useful for future
research in Dox-induced cardiotoxicity in vitro. In future
studies, lower doses of Dox and longer treatment times in H9c2
cells may more efficiently mimic the progression of chronic
cardiotoxicity induced by Dox. In the future, animal models of
spontaneous tumors or primary tumors should be used to study Dox
cardiotoxicity and analyze miRNA expression profiles in
cardiomyocytes. Analysis of tissues from patients with Dox-induced
heart failure may provide further insight into miRNA expression
profiles, but samples cannot be collected at the early stage of
Dox-induced cardiotoxicity. The use of induced pluripotent stem
cell-derived human heart organoids may replace some in vivo
experiments and partly overcome the limitations of cell models in
the study of Dox cardiotoxicity. The present study provided new
insights for understanding the dysregulation of miRNA expression in
H9c2 cardiomyocytes induced by Dox at an early stage. Some of these
miRNAs may have potential as biomarkers or therapeutic targets.
Overall, the present study focused on investigating
alterations of miRNA expression in rat cardiomyocyte within 24 h
(12- and 24-h time points) of Dox treatment. The expression levels
of 76 miRNAs were significantly altered in cardiomyocytes treated
with Dox, and had >1.5-fold-changes compared with the 0-h time
point group. Only 9 of these miRNAs have been reported in previous
studies of Dox-induced cardiotoxicity (22–24,34,35).
These differentially expressed miRNAs were ubiquitously involved in
numerous biological processes and functional pathways involved in
the regulation of apoptosis, ECM remodeling, cell differentiation
and cell metabolism. Based on the present data and biological
information database, the interactions with the target genes of
these miRNAs were analyzed and visual gene interaction networks
were mapped. These interaction networks may help to further
understand the roles of miRNA dysregulation in the development of
Dox-induced cardiotoxicity at the early stage, as well as provide
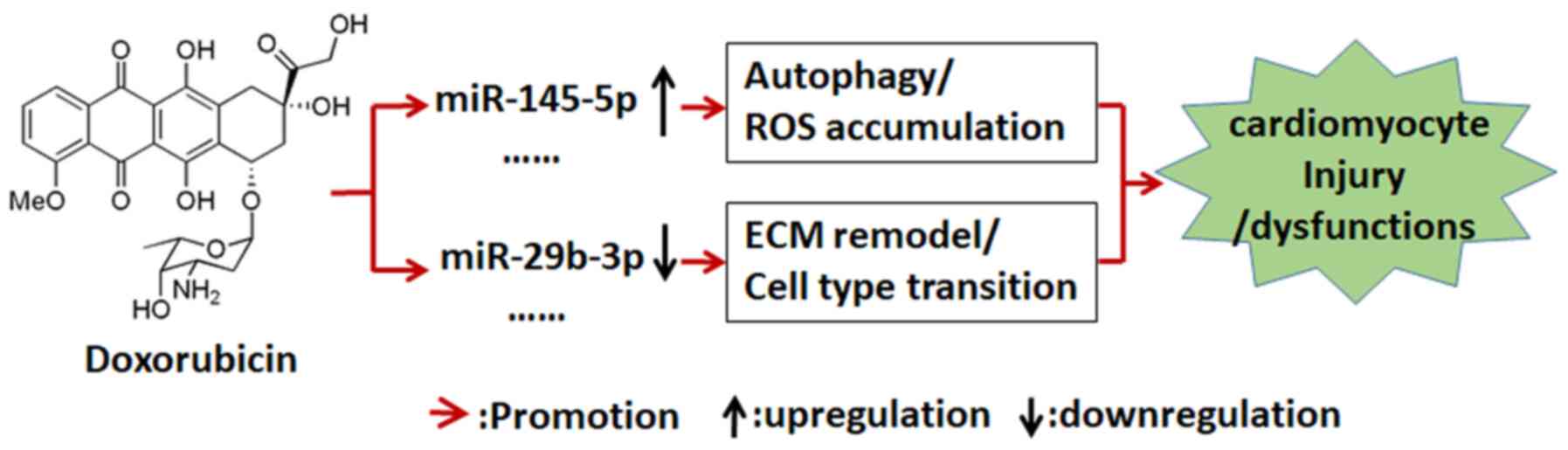
further insight in the involved molecular mechanisms (Fig. 6).
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
The authors would like to thank Dr Yali Kang and Dr
Lingyi Li (Shanghai Jiao Tong University School of Biomedicine
Engineering, Shanghai, China) for helping to analyze the microRNA
sequencing data and data normalization, and Dr Xiaohong Ma (Huadong
Hospital Affiliated to Fudan University, Shanghai, China) for his
advice on this project and for reviewing the manuscript.
Funding
The present study was supported by the Shanghai
Municipal Commission of Health and Family planning (grant no.
201540302), the Science and Technology Commission of Shanghai
Changning (grant no. CNKW2016Y03) and Shanghai Jiao Tong University
Medicine-Engineering Joint Fund (grant no. YG2015MS64).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the National Genomics Data Center
repository (https://bigd.big.ac.cn/; accession
no. CRA003714).
Authors' contributions
LJ and WJ designed the experiments. YC and YX
performed the tissue culture and established the cell models. ZD
and YW prepared the microRNA library. YC, YZ and YJ analyzed the
data of the miRNA expression profiles and the biological
information. LJ and WJ wrote the manuscript. YC, YX, LJ and WJ
confirm the authenticity of all the raw data and the processed data
in the present study. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mitry MA and Edwards JG: Doxorubicin
induced heart failure: Phenotype and molecular mechanisms. Int J
Cardiol Heart Vasc. 10:17–24. 2016.PubMed/NCBI
|
|
2
|
Koleini N and Kardami E: Autophagy and
mitophagy in the context of doxorubicin-induced cardiotoxicity.
Oncotarget. 8:46663–46680. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhu HJ, Han ZY, He SF, Jin SY, Xu SJ, Fang
XD and Zhang Y: Specific MicroRNAs comparisons in hypoxia and
morphine preconditioning against hypoxia-reoxgenation injury with
and without heart failure. Life Sci. 170:82–92. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li N, Wang WB, Bao H, Shi Q, Jiang ZL, Qi
YX and Han Y: MicroRNA-129-1-3p regulates cyclic stretch-induced
endothelial progenitor cell differentiation by targeting Runx2. J
Cell Biochem. 120:5256–5267. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu L, Yuan Y, He X, Xia X and Mo X:
MicroRNA-1 upregulation promotes myocardiocyte proliferation and
suppresses apoptosis during heart development. Mol Med Rep.
15:2837–2842. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ren N and Wang M: microRNA-212-induced
protection of the heart against myocardial infarction occurs via
the interplay between AQP9 and PI3K/Akt signaling pathway. Exp Cell
Res. 370:531–541. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo L, Zheng X, Wang E, Jia X, Wang G and
Wen J: Irigenin treatment alleviates doxorubicin (DOX)-induced
cardiotoxicity by suppressing apoptosis, inflammation and oxidative
stress via the increase of miR-425. Biomed Pharmacother.
125:1097842020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ji X, Ding W, Xu T, Zheng X, Zhang J, Liu
M, Liu G and Wang J: MicroRNA-31-5p attenuates doxorubicin-induced
cardiotoxicity via quaking and circular RNA Pan3. J Mol Cell
Cardiol. 140:56–67. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fu J, Peng C, Wang W, Jin H, Tang Q and
Wei X: Let-7 g is involved in doxorubicin induced myocardial
injury. Environ Toxicol Pharmacol. 33:312–317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li N, Zhou H and Tang Q: miR-133: A
suppressor of cardiac remodeling? Front Pharmacol. 9:9032018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu JN, Fu YH, Hu ZQ, Li WY, Tang CM, Fei
HW, Yang H, Lin QX, Gou DM, Wu SL and Shan ZX: Activation of
miR-34a-5p/Sirt1/p66shc pathway contributes to doxorubicin-induced
cardiotoxicity. Sci Rep. 7:118792017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao L, Qi Y, Xu L, Tao X, Han X, Yin L
and Peng J: MicroRNA-140-5p aggravates doxorubicin-induced
cardiotoxicity by promoting myocardial oxidative stress via
targeting Nrf2 and Sirt2. Redox Biol. 15:284–296. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ruggeri C, Gioffré S, Achilli F, Colombo
GI and D'Alessandra Y: Role of microRNAs in doxorubicin-induced
cardiotoxicity: An overview of preclinical models and cancer
patients. Heart Fail Rev. 23:109–122. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bolger AM, Lohse M and Usadel B:
Trimmomatic: A flexible trimmer for Illumina sequence data.
Bioinformatics. 30:2114–2120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Andrews S: FastQC: A quality control tool
for high throughput sequence data. 2010.http://www.bioinformatics.babraham.ac.uk/projects/fastqc/April
26–2010
|
|
16
|
Langmead B and Salzberg SL: Fast
gapped-read alignment with Bowtie 2. Nat Methods. 9:357–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Love MI, Huber W and Anders S: Moderated
estimation of fold change and dispersion for RNA-seq data with
DESeq2. Genome Biol. 15:5502014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Buermans HP, Ariyurek Y, van Ommen G, den
Dunnen JT't and Hoen PA: New methods for next generation sequencing
based microRNA expression profiling. BMC Genomics. 11:7162010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Legrain P and Rain JC: Twenty years of
protein interaction studies for biological function deciphering. J
Proteomics. 107:93–97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee H, Han S, Kwon CS and Lee D:
Biogenesis and regulation of the let-7 miRNAs and their functional
implications. Protein Cell. 7:100–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Desai VG, C Kwekel J, Vijay V, Moland CL,
Herman EH, Lee T, Han T, Lewis SM, Davis KJ, Muskhelishvili L, et
al: Early biomarkers of doxorubicin-induced heart injury in a mouse
model. Toxicol Appl Pharmacol. 281:221–229. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vacchi-Suzzi C, Bauer Y, Berridge BR,
Bongiovanni S, Gerrish K, Hamadeh HK, Letzkus M, Lyon J, Moggs J,
Paules RS, et al: Perturbation of microRNAs in rat heart during
chronic doxorubicin treatment. PLoS One. 7:e403952012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roca-Alonso L, Castellano L, Mills A,
Dabrowska AF, Sikkel MB, Pellegrino L, Jacob J, Frampton AE, Krell
J, Coombes RC, et al: Myocardial MiR-30 downregulation triggered by
doxorubicin drives alterations in β-adrenergic signaling and
enhances apoptosis. Cell Death Dis. 6:e17542015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yuan M, Zhang L, You F, Zhou J, Ma Y, Yang
F and Tao L: MiR-145-5p regulates hypoxia-induced inflammatory
response and apoptosis in cardiomyocytes by targeting CD40. Mol
Cell Biochem. 431:123–131. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Toro R, Blasco-Turrión S, Morales-Ponce
FJ, Gonzalez P, Martínez-Camblor P, López-Granados A, Brugada R,
Campuzano O, Pérez-Serra A, Rosa Longobardo F, et al: Plasma
microRNAs as biomarkers for Lamin A/C-related dilated
cardiomyopathy. J Mol Med (Berl). 96:845–856. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
da Silva W, dos Santos RA and Moraes KC:
Mir-351-5p contributes to the establishment of a pro-inflammatory
environment in the H9c2 cell line by repressing PTEN expression.
Mol Cell Biochem. 411:363–371. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Billia F, Hauck L, Konecny F, Rao V, Shen
J and Mak TW: PTEN-inducible kinase 1 (PINK1)/Park6 is
indispensable for normal heart function. Proc Natl Acad Sci USA.
108:9572–9577. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gao Y, Chu M, Hong J, Shang J and Xu D:
Hypoxia induces cardiac fibroblast proliferation and phenotypic
switch: A role for caveolae and caveolin-1/PTEN mediated pathway. J
Thorac Dis. 6:1458–1468. 2014.PubMed/NCBI
|
|
30
|
Zheng L, Han X, Hu Y, Zhao X, Yin L, Xu L,
Qi Y, Xu Y, Han X, Liu K and Peng J: Dioscin ameliorates intestinal
ischemia/reperfusion injury via adjusting
miR-351-5p/MAPK13-mediated inflammation and apoptosis. Pharmacol
Res. 139:431–439. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu Y, Mao Z, Xu L, Yin L, Tao X, Tang Z,
Qi Y, Sun P and Peng J: Protective effect of dioscin against
intestinal ischemia/reperfusion injury via adjusting
miR-351-5p-mediated oxidative stress. Pharmacol Res. 137:56–63.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Venkatakrishnan CD, Tewari AK, Moldovan L,
Cardounel AJ, Zweier JL, Kuppusamy P and Ilangovan G: Heat shock
protects cardiac cells from doxorubicin-induced toxicity by
activating p38 MAPK and phosphorylation of small heat shock protein
27. Am J Physiol Heart Circ Physiol. 291:H2680–H2691. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guo R, Wu K, Chen J, Mo L, Hua X, Zheng D,
Chen P, Chen G, Xu W and Feng J: Exogenous hydrogen sulfide
protects against doxorubicin-induced inflammation and cytotoxicity
by inhibiting p38MAPK/NFκB pathway in H9c2 cardiac cells. Cell
Physiol Biochem. 32:1668–1680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nishimura Y, Kondo C, Morikawa Y, Tonomura
Y, Torii M, Yamate J and Uehara T: Plasma miR-208 as a useful
biomarker for drug-induced cardiotoxicity in rats. J Appl Toxicol.
35:173–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rigaud VO, Ferreira LR, Ayub-Ferreira SM,
Ávila MS, Brandão SM, Cruz FD, Santos MH, Cruz CB, Alves MS, Issa
VS, et al: Circulating miR-1 as a potential biomarker of
doxorubicin-induced cardiotoxicity in breast cancer patients.
Oncotarget. 8:6994–7002. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cappetta D, Rossi F, Piegari E, Quaini F,
Berrino L, Urbanek K and De Angelis A: Doxorubicin targets multiple
players: A new view of an old problem. Pharmacol Res. 127:4–14.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dupuy AM, Kuster N, Curinier C, Huet F,
Plawecki M, Solecki K, Roubille F and Cristol JP: Exploring
collagen remodeling and regulation as prognosis biomarkers in
stable heart failure. Clin Chim Acta. 490:167–171. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nagao K, Inada T, Tamura A, Kajitani K,
Shimamura K, Yukawa H, Aida K, Sowa N, Nishiga M, Horie T, et al:
Circulating markers of collagen types I, III, and IV in patients
with dilated cardiomyopathy: Relationships with myocardial collagen
expression. ESC Heart Fail. 5:1044–1051. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou S, Lei D, Bu F, Han H, Zhao S and
Wang Y: MicroRNA-29b-3p targets SPARC gene to protect cardiocytes
against autophagy and apoptosis in hypoxic-induced H9c2 cells. J
Cardiovasc Transl Res. 12:358–365. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Han Z, Zhang T, He Y, Li G, Li G and Jin
X: Inhibition of prostaglandin E2 protects abdominal aortic
aneurysm from expansion through regulating miR-29b-mediated
fibrotic ECM expression. Exp Ther Med. 16:155–160. 2018.PubMed/NCBI
|
|
41
|
Drummond CA, Fan X, Haller ST, Kennedy DJ,
Liu J and Tian J: Na/K-ATPase signaling mediates miR-29b-3p
regulation and cardiac fibrosis formation in mice with chronic
kidney disease. PLoS One. 13:e01976882018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang WG, Chen L, Dong Q, He J, Zhao HD,
Li FL and Li H: Mmu-miR-702 functions as an anti-apoptotic mirtron
by mediating ATF6 inhibition in mice. Gene. 531:235–242. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim BM and Choi MY: Non-canonical
microRNAs miR-320 and miR-702 promote proliferation in
Dgcr8-deficient embryonic stem cells. Biochem Biophys Res Commun.
426:183–189. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huang W, Feng Y, Liang J, Yu H, Wang C,
Wang B, Wang M, Jiang L, Meng W, Cai W, et al: Loss of microRNA-128
promotes cardiomyocyte proliferation and heart regeneration. Nat
Commun. 9:7002018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kuppusamy KT, Jones DC, Sperber H, Madan
A, Fischer KA, Rodriguez ML, Pabon L, Zhu WZ, Tulloch NL, Yang X,
et al: Let-7 family of microRNA is required for maturation and
adult-like metabolism in stem cell-derived cardiomyocytes. Proc
Natl Acad Sci USA. 112:E2785–E2794. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Du Y, Zhang M, Zhao W, Shu Y, Gao M,
Zhuang Y, Yang T, Mu W, Li T, Li X, et al: Let-7a regulates
expression of β1-adrenoceptors and forms a negative feedback
circuit with the β1-adrenoceptor signaling pathway in chronic
ischemic heart failure. Oncotarget. 8:8752–8764. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li X, Wang B, Cui H, Du Y, Song Y, Yang L,
Zhang Q, Sun F, Luo D, Xu C, et al: let-7e replacement yields
potent anti-arrhythmic efficacy via targeting beta 1-adrenergic
receptor in rat heart. J Cell Mol Med. 18:1334–1343. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bao MH, Feng X, Zhang YW, Lou XY, Cheng Y
and Zhou HH: Let-7 in cardiovascular diseases, heart development
and cardiovascular differentiation from stem cells. Int J Mol Sci.
14:23086–23102. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tolonen AM, Magga J, Szabó Z, Viitala P,
Gao E, Moilanen AM, Ohukainen P, Vainio L, Koch WJ, Kerkelä R, et
al: Inhibition of Let-7 microRNA attenuates myocardial remodeling
and improves cardiac function postinfarction in mice. Pharmacol Res
Perspect. 2:e000562014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fang J, Song XW, Tian J, Chen HY, Li DF,
Wang JF, Ren AJ, Yuan WJ and Lin L: Overexpression of microRNA-378
attenuates ischemia-induced apoptosis by inhibiting caspase-3
expression in cardiac myocytes. Apoptosis. 17:410–423. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yuan J, Liu H, Gao W, Zhang L, Ye Y, Yuan
L, Ding Z, Wu J, Kang L, Zhang X, et al: MicroRNA-378 suppresses
myocardial fibrosis through a paracrine mechanism at the early
stage of cardiac hypertrophy following mechanical stress.
Theranostics. 8:2565–2582. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ganesan J, Ramanujam D, Sassi Y, Ahles A,
Jentzsch C, Werfel S, Leierseder S, Loyer X, Giacca M, Zentilin L,
et al: MiR-378 controls cardiac hypertrophy by combined repression
of mitogen-activated protein kinase pathway factors. Circulation.
127:2097–2106. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kobashigawa LC, Xu YC, Padbury JF, Tseng
YT and Yano N: Metformin protects cardiomyocyte from doxorubicin
induced cytotoxicity through an AMP-activated protein kinase
dependent signaling pathway: An in vitro study. PLoS One.
9:e1048882014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu X, Wei J, Ma Z and He Y: Rapamycin-
and starvation-induced autophagy are associated with miRNA
dysregulation in A549 cells. Acta Biochim Biophys Sin (Shanghai).
51:393–401. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kim JH, Lee DK, Kim J, Choi S, Park W, Ha
KS, Kim TH, Choe J, Won MH, Kwon YG and Kim YM: A miRNA-101-3p/Bim
axis as a determinant of serum deprivation-induced endothelial cell
apoptosis. Cell Death Dis. 8:e28082017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wu J, Sun C, Wang R, Li J, Zhou M, Yan M,
Xue X and Wang C: Cardioprotective effect of paeonol against
epirubicin-induced heart injury via regulating miR-1 and PI3K/AKT
pathway. Chem Biol Interact. 286:17–25. 2018. View Article : Google Scholar : PubMed/NCBI
|