Introduction
Gallbladder cancer (GBC) is considered the most
common bile tract cancer (1), and
patients with advanced disease have a 5-year survival rate of
<5% (2). Given that there are no
obvious symptoms and relevant biomarkers for GBC, patients are
commonly diagnosed at advanced stages (3,4).
Chemotherapy, radiotherapy and surgical resection remain the most
common treatment strategies for GBC (5). Despite advancements in treatment
approaches, the overall survival rate of patients with GBC remains
low (5). Thus, it is important to
determine the molecular mechanisms underlying GBC
tumorigenesis.
Circular RNAs (circRNAs) are derived from endogenous
non-coding RNAs with a closed loop structure (6). circRNAs have been reported to play
important roles in different types of cancer by binding to
microRNAs (miRNAs/miRs) (7).
Circular mitochondrial translation optimization 1 homologue
(circMTO1) inhibits progression of gastric cancer (8), colorectal cancer (9), bladder cancer (10) and glioblastoma (11). However, the role of circMTO1 in GBC
remains unknown. Recently, Wang et al (12) demonstrated that circMTO1 acts as a
biomarker in GBC. Shi et al (13) reported that overexpression of
circMTO1 suppresses cell apoptosis via flow cytometry. Thus, it is
important to determine the molecular mechanism underlying
circMTO1-induced GBC tumorigenesis.
miRNAs are small non-coding RNAs of 20–22
nucleotides in length, which regulate gene expression at the
post-transcriptional level (14).
Notably, miRNAs are involved in tumor progression (15,16).
Previous studies have reported that miR-219a-5p inhibits cancer
cell proliferation and migration (17,18). In
addition, miR-219a-5p can sensitize non-small cell lung cancer
cells to cisplatin by regulating fibroblast growth factor 9
expression (17). miR-219a-5p
promotes irradiation-induced apoptosis of non-small cell lung
cancer cells (19). Furthermore,
miR-219a-5p can also induce neuronal apoptosis (20). Based on these findings, it was
hypothesized that miR-219a-5p acts as a potential effector for
circMTO1 in GBC.
The present study aimed to determine the underlying
molecular mechanism during GBC progression. In addition, rationales
for diagnosing and treating GBC via the circMTO1-targeted approach
are discussed.
Materials and methods
Cell culture
Human GBC-SD cells were purchased from Kunming Cell
Bank, while NOZ cells were purchased from the Japanese Collection
of Research Bioresources Cell Bank. Cells were maintained in DMEM
supplemented with 10% fetal bovine serum (FBS) and 1%
penicillin/streptomycin (all purchased from Gibco; Thermo Fisher
Scientific, Inc.), at 37°C with 5% CO2, according to the
manufacturer's instructions. Human dermal lymphatic endothelial
cells (HDLECs; PromoCell GmbH) and maintained in endothelial cell
growth medium (PromoCell GmbH), at 37°C with 5% CO2.
Generating stable cell lines
To generate stable cell lines, 1 µg pLKO.1-short
hairpin (sh)RNAs (Invitrogen; Thermo Fisher Scientific, Inc.),
packaging vectors 0.5 µg pCMV–VSVG (Addgene, Inc.) and 0.5 µg
pCMV-PAX2 (Addgene, Inc.) were introduced into 293T cells to
generate 2nd lentiviruses. Lentiviruses were collected from the
medium after 36–48 h of transfection and maintained in fresh media.
We used lentiviruses (MOI=5) to infect into GBC-SD and NOZ cells
for 24 h. Puromycin (2 µg/ml) was used to obtain pooled resistant
cells after 24 h and 1 µg/ml of puromycin were used for
maintenance. After 1 week, subsequent experiments were performed.
The following sequences were used: sh-negative control (NC),
5′-AUCAGCCAAUCGGUCAACCUUC-3′; sh-circMTO1-1,
5′-GUGGGGUUGUUUUGGGUCAGA-3′; and sh-circMTO1-2,
5′-GUUGUUUUGGGUCAGAUGUCA-3′.
Cell transfection
miRNAs (50 µM; Shanghai GenePharma Co., Ltd.) were
transfected into GBC-SD and NOZ cells using
Lipofectamine® (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. Following
incubation for 48 h at 37°C, the cells were collected for
subsequent analysis. The following sequences were used: NC mimic,
5′-CAUUCAUCCAUCAAUCGGGCAGGCCUUUAAGCUAACAUGGAA-3′; miR-219a-5p
mimic, 5′-UGAUUGUCCAAACGCAAUUCUAAUUGCGUUUGGACAAUCAUU-3′; and NC
inhibitor, 5′-UUCAGGCAAUCCAAAUGCAGG-3′; miR-219a-5p inhibitor,
5′-AGAAUUGCGUUUGGACAAUCA-3′.
StarBase and TargetScan
prediction
The StarBase 2.0 online software (http://starbase.sysu.edu.cn/starbase2)
created by Sun Yat-sen University was used to predict the potential
interacting miRNAs. In addition, TargetScan v7.2 (http://www.targetscan.org) created by MIT was used to
predict protein-coding genes as potential targets.
MTT assay
GBC-SD and NOZ cells were seeded into 96-well plates
at a density of ~5,000 cells/well. MTT reagent was added to each
well and incubated for 4 h at 37°C with 5% CO2.
Following the MTT incubation, the purple formazan crystals were
dissolved using 150 µl DMSO for 10 min and viability was
subsequently analyzed at a wavelength of 490 nm.
Migration assay
GBC cells (1×105 cells) were suspended in
200 µl of DMEM media (Gibco; Thermo Fisher Scientific, Inc.). In
this assay, the upper chambers had 8-µm-pore membranes and
incubated for 4–6 h at 37°C. DMEM media (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 20% FBS was added to the lower
chambers and incubated for 24 h at 37°C. Migratory cells were
stained with 0.005% crystal violet dye for 2 h at room temperature
and observed under an invert light microscope (magnification,
×200).
Immunofluorescence
GBC cells were fixed with 4% paraformaldehyde for 10
min at room temperature. Triton was subsequently used to
permeabilize the cell membranes. Annexin V-FITC (eBioscience;
Thermo Fisher Scientific, Inc.) was used to probe the signal, and
cell nuclei were stained with DAPI at room temperature for 30 min
and observed under a fluorescence microscope (magnification,
×200).
Western blotting
GBC cells were lysed using RIPA buffer and total
proteins were extracted. The Rapid Gold BCA method (Thermo Fisher
Scientific, Inc.) was used to determine protein concentrations,
according to the manufacturer's protocol and ~50 µg total
protein/lane was separated via SDS-PAGE on a 10% gel. The separated
proteins were subsequently transferred onto PVDF membranes and
blocked with 5% non-fat milk at room temperature for 1 h. The
membranes were incubated with primary antibodies against Smad2
(cat. no. 5339), Smad4 (cat. no. 46535), epidermal growth factor
receptor (EGFR; cat. no. 2085), cleaved caspase-3 (cat. no.
25546-1-AP) and GAPDH (cat. no. 8884) overnight at 4°C (all 1:1,000
and purchased from Cell Signaling Technology, Inc.). Following the
primary incubation, membranes were incubated with anti-rabbit
HRP-conjugated IgG (cat. no. 7074) and anti-mouse HRP-conjugated
IgG (cat. no. 7076) secondary antibodies at room temperature for 1
h (both 1:2,000 and purchased from Cell Signaling Technology,
Inc.). Protein bands were visualized using ECL reagents (Thermo
Fisher Scientific, Inc.) and Bio-Rad gel imaging machine (Bio-Rad
Laboratories, Inc.).
The Cancer Genome Atlas (TCGA)
analysis
The gene expression data (RNAseq, NGS) of GBC were
downloaded from TCGA dataset (36 tumor tissues and nine normal
tissues, http://tcga.xenahubs.net). The
present study did not have any inclusion or exclusion criteria.
circMTO1 expression was analyzed using a two-tailed unpaired
Student's t-test. Clinical characteristics of patients with
gallbladder cancer were presented in Table I.
 | Table I.Clinical characteristics of patients
with gallbladder cancer (n=36). |
Table I.
Clinical characteristics of patients
with gallbladder cancer (n=36).
| Characteristic | Patients, n |
|---|
| Age, years |
|
|
≤60 | 17 |
|
>60 | 19 |
| Sex |
|
|
Male | 20 |
|
Female | 16 |
|
Cholecystolithiasis |
|
|
Absent | 28 |
|
Present | 8 |
| Diabetes |
|
|
Absent | 12 |
|
Present | 24 |
| Jaundice |
|
|
Absent | 21 |
|
Present | 15 |
| Pathological
type |
|
|
Adenocarcinoma | 36 |
|
Adenosquamous carcinoma | 0 |
|
Papillocarcinoma | 0 |
| Degree of
differentiation |
|
|
Poor | 22 |
|
Moderate-well | 14 |
| T stage |
|
| T1 | 10 |
| T2 | 9 |
| T4 | 17 |
Patient sorting criterion
The patients were sorted into high and low
expression groups according to the median circMTO1 RNA expression
(cut-off value=2.8) in patients with GBC tumors. A total of 18
patients were classified into the high circMTO1 expression group
and 18 into the low circMTO1 expression group.
Tube formation assay
GFR Matrigel was mixed with PBS (1:5) and coated in
24-well plates at 37°C for 4–5 h. HDLECs were subsequently seeded
into the plates (12 wells) at a density of ~5,000 cells/well, and
the tubes were observed under an inverted light microscope
(magnification, ×200).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from GBC cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA (1 µg) was reverse transcribed into cDNA using the
PrimeScript RT reagent kit (Takara Bio, Inc.) at 37°C for 15 min
and 85°C for 1 min. qPCR was subsequently performed to measure
relative gene expression levels, using the 2−ΔΔCq method
(21). SYBR Green dye (Roche Applied
Science) was used for qPCR. The following thermocycling conditions
were used: Initial denaturation at 95°C for 5 min (step 1), 95°C
for 30 sec (step 2), 60°C for 30 sec (step 3), 72°C for 30 sec
(step 4), 40 cycles (step 2–4). β-actin was used as the reference
gene for circMTO1, Smad2, Smad4 and EGFR. U6 was used as the
reference gene for miR-219a-5p. The following primer sequences were
used for qPCR: circMTO1 forward, 5′-GCCTGAACACACTGGGAAAT-3′ and
reverse, 5′-CACAGATGCGAGAACACAGG-3′; Smad2 forward,
5′-CTTTGTGCAGAGCCCCAATT-3′ and reverse, 5′-CTTGTTACCGTCTGCCTTCG-3′;
Smad4 forward, 5′-TCCAGCCTCCCATTTCCAAT-3′ and reverse,
5′-ACCTTGCTCTCTCAATGGCT-3′; EGFR forward,
5′-AGGTGAAAACAGCTGCAAGG-3′ and reverse, 5′-AGGTGATGTTCATGGCCTGA-3′;
miR-219a-5p forward, 5′-CTCCTGATTGTCCAAAC-3′ and reverse,
5′-CGCTCGAGGTTTGGGG-3′; β-actin forward, 5′-TGGCATCCACGAAAC TAC
CT-3′ and reverse, 5′-TCTCCTTCTGCATCCTGTCG-3′; and U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8.0 software (GraphPad Software, Inc.). Unpaired Student's
t-test was used to compare differences between two groups, while
one-way ANOVA followed by Tukey's post hoc test was used to compare
differences between multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
circMTO1 knockdown impairs GBC
tumorigenesis
To determine the role of circMTO1 in GBC
progression, circMTO1 expression was suppressed using shRNAs
(sh-circMTO1-1 and sh-circMTO1-2). RT-qPCR analysis was performed
to detect circMTO1 expression. The results demonstrated that
circMTO1 expression significantly decreased (~70-80%) in circMTO1
knockdown cells compared with sh-NC cells (Fig. 1A) (GBC-SD, sh-circMTO1-1 vs. sh-NC,
P=0.0002; sh-circMTO1-2 vs. sh-NC, P=0.0003; NOZ, sh-circMTO1-1 vs.
sh-NC, P=0.0005; sh-circMTO1-2 vs. sh-NC, P=0.0006).
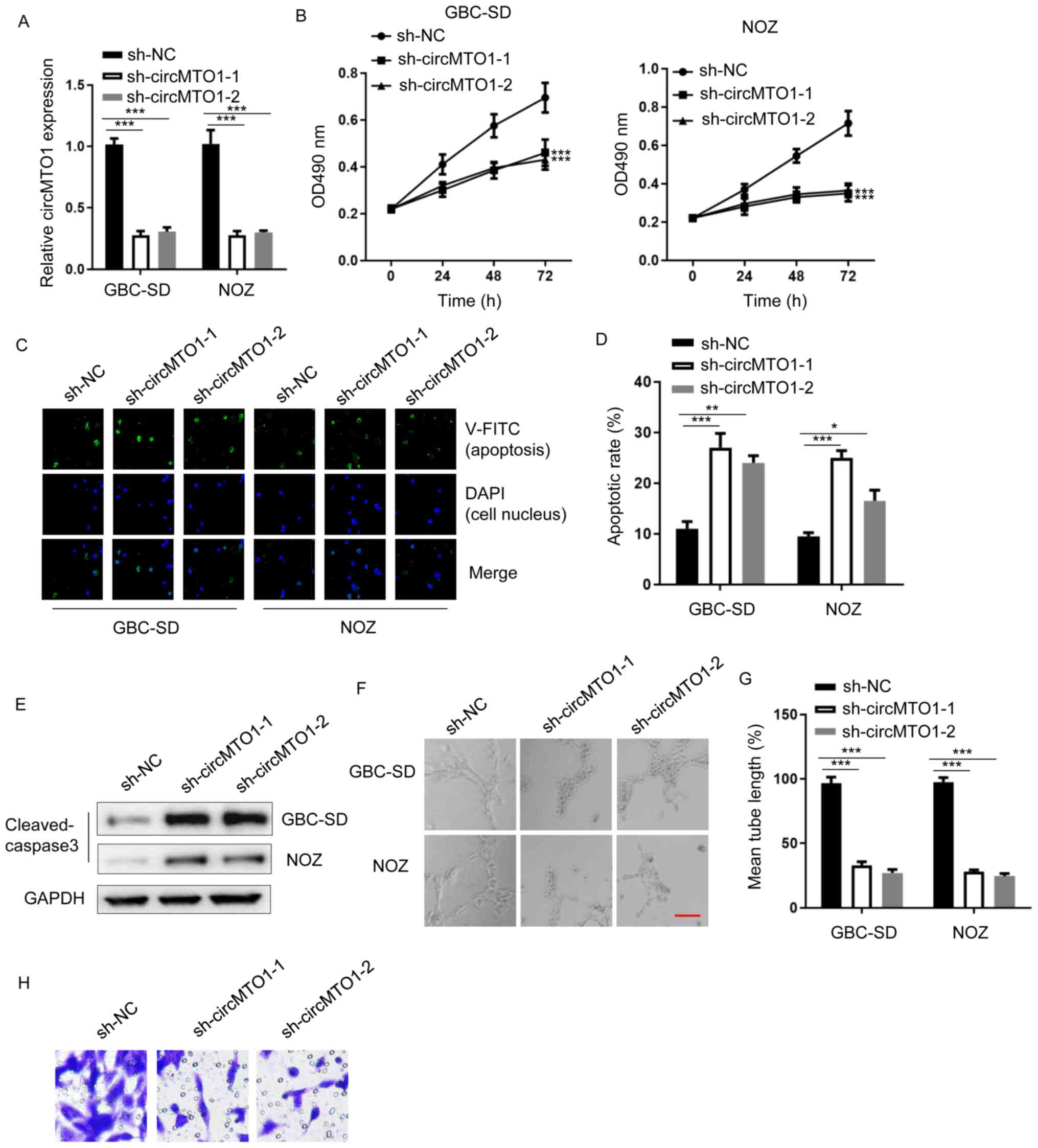 | Figure 1.circMTO1 knockdown impairs GBC
tumorigenesis. (A) Reverse transcription-quantitative PCR analysis
was performed to detect circMTO1 expression in GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-2. (B) The MTT
assay was performed to assess the viability of GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-2. (C and D)
Annexin V staining was performed to assess the apoptosis of GBC-SD
and NOZ cells transfected with sh-NC, sh-circMTO1-1 or
sh-circMTO1-2. (E) Western blot analysis was performed to assess
cleaved caspase-3 protein expression in GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-2. GAPDH was
used as the internal control. (F and G) The lymphatic vessel
formation assay was performed to assess tube formation of human
dermal lymphatic endothelial, GBC-SD and NOZ cells transfected with
sh-NC, sh-circMTO1-1 or sh-circMTO1-2 (scale bar, 2 µm). (H) The
Transwell assay was performed to assess the migratory ability of
GBC-SD cells transfected with sh-NC, sh-circMTO1-1 or
sh-circMTO1-2. *P<0.05; **P<0.01; ***P<0.001. circMTO1,
circular mitochondrial translation optimization 1 homologue; GBC,
gallbladder cancer; sh, short hairpin; NC, negative control; OD,
optical density. |
circMTO1 has been reported to promote tumorigenesis
of cervical cancer (22). The MTT
assay was performed to assess the viability of GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 and sh-circMTO1-2. The
results demonstrated that cells transfected with sh-circMTO1-1 or
sh-circMTO1-2 exhibited evidently lower viability compared with
sh-NC cells (Fig. 1B) (GBC-SD,
sh-circMTO1-1 vs. sh-NC, P=0.0003; sh-circMTO1-2 vs. sh-NC,
P=0.0004; NOZ, sh-circMTO1-1 vs. sh-NC, P=0.0002; sh-circMTO1-2 vs.
sh-NC, P=0.0007). Annexin V staining was performed to assess
apoptosis of GBC-SD and NOZ cells transfected with sh-NC,
sh-circMTO1-1 or sh-circMTO1-2. The results indicated that circMTO1
knockdown increased the apoptotic rate of cells (Fig. 1C and D) (GBC-SD, sh-circMTO1-1 vs.
sh-NC, P=0.0005; sh-circMTO1-2 vs. sh-NC, P=0.0063; NOZ,
sh-circMTO1-1 vs. sh-NC, P=0.0008; sh-circMTO1-2 vs. sh-NC,
P=0.036). To confirm, western blot analysis was performed to detect
cleaved caspase-3 protein expression in GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-2. The results
demonstrated that cleaved caspase-3 protein expression was
upregulated in circMTO1 knockdown cells (Fig. 1E). In addition, the effect of
circMTO1 on tube formation of HDLECs was investigated. The results
indicated that circMTO1 knockdown attenuated tube formation of
HDLECs (Fig. 1F and G) (GBC-SD,
sh-circMTO1-1 vs. sh-NC, P=0.0004; sh-circMTO1-2 vs. sh-NC,
P=0.0003; NOZ, sh-circMTO1-1 vs. sh-NC, P=0.0007; sh-circMTO1-2 vs.
sh-NC, P=0.0005). The results of the Transwell assay demonstrated
that circMTO1 knockdown decreased the number of migratory GBC-SD
cells (Fig. 1H). Taken together,
these results suggest that circMTO1 acts as an oncogene in GBC
progression.
miR-219a-5p inhibitor partially
restores sh-circMTO1-attenuated GBC tumorigenesis
The StarBase database was used to determine the
potential molecular mechanism underlying circMTO1-induced
tumorigenesis of GBC. Analysis revealed that miR-219a-5p is an
interacting molecule for circMTO1 in GBC (Fig. 2A). RT-qPCR analysis demonstrated that
miR-219a-5p expression was substantially regulated in GBC-SD or NOZ
cells transfected with NC mimic, miR-219a-5p mimic, NC inhibitor or
miR-219a-5p inhibitor (Fig. 2B)
(GBC-SD, miR-219a-5p mimic vs. NC mimic, P=0.0004; miR-219a-5p
inhibitor vs. NC inhibitor, P=0.0005; NOZ, miR-219a-5p mimic vs. NC
mimic, P=0.0003; miR-219a-5p inhibitor vs. NC inhibitor,
P=0.0003).
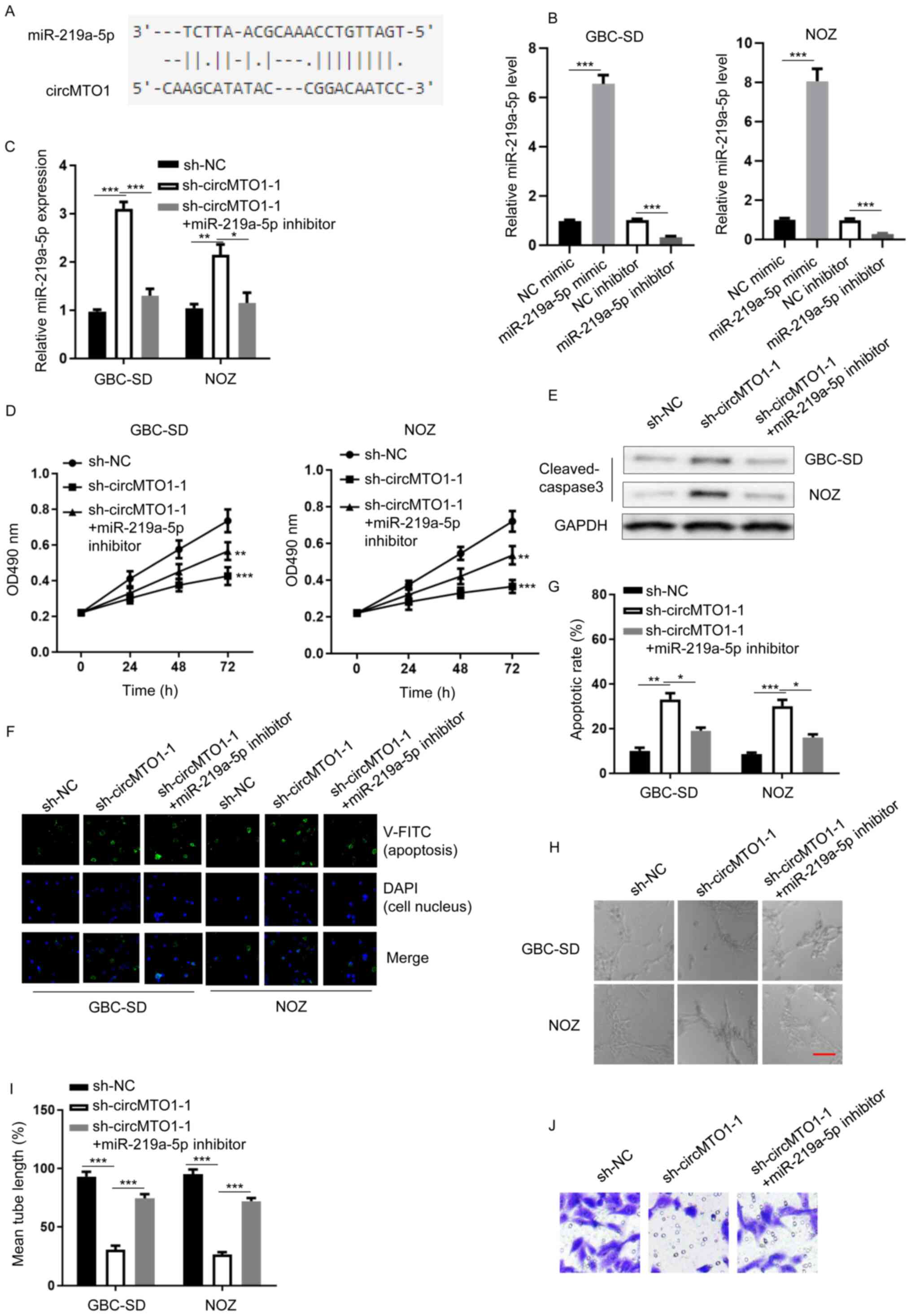 | Figure 2.miR-219a-5p inhibitor partially
restores sh-circMTO1-attenuated GBC tumorigenesis. (A)
Bioinformatics analysis revealed that miR-219-5p is a potential
target of circMTO1. (B) RT-qPCR analysis was performed to detect
miR-219a-5p expression in GBC-SD and NOZ cells transfected with NC
mimic, miR-219a-5p mimic, NC inhibitor or miR-219a-5p inhibitor.
(C) RT-qPCR analysis was performed to detect miR-219a-5p expression
in GBC-SD and NOZ cells transfected with sh-NC, sh-circMTO1-1 or
sh-circMTO1-1 + miR-219a-5p inhibitor. (D) The MTT assay was
performed to assess the viability of GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-1 +
miR-219a-5p inhibitor. (E) Western blot analysis was performed to
detect cleaved caspase-3 protein expression in GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-1 +
miR-219a-5p inhibitor. GAPDH was used as the internal control. (F
and G) Annexin V staining was performed to assess the apoptosis of
GBC-SD and NOZ cells transfected with sh-NC, sh-circMTO1-1 or
sh-circMTO1-1 + miR-219a-5p inhibitor. (H and I) The lymphatic
vessel formation assay was performed to assess tube formation of
human dermal lymphatic endothelial, GBC-SD and NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-1 +
miR-219a-5p inhibitor (scale bar, 2 µm). (J) The Transwell assay
was performed to assess the migratory ability of NOZ cells
transfected with sh-NC, sh-circMTO1-1 or sh-circMTO1-1 +
miR-219a-5p inhibitor. *P<0.05; **P<0.01; ***P<0.001. miR,
microRNA; sh, short hairpin; circMTO1, circular mitochondrial
translation optimization 1 homologue; GBC, gallbladder cancer;
RT-qPCR, reverse transcription-quantitative PCR; NC, negative
control. |
The function of miR-219a-5p in circMTO1-regulated
GBC phenotypes was investigated. RT-qPCR analysis demonstrated that
miR-219a-5p expression was upregulated following circMTO1 knockdown
(Fig. 2C) (GBC-SD, sh-circMTO1-1 vs.
sh-NC, P=0.0002; sh-circMTO1-1 + miR-219a-5p inhibitor vs.
sh-circMTO1-1, P=0.0006; NOZ, sh-circMTO1-1 vs. sh-NC, P=0.0067;
sh-circMTO1-1 + miR-219a-5p inhibitor vs. sh-circMTO1-1, P=0.028).
The results of the MTT assay demonstrated that transfection with
miR-219a-5p inhibitor rescued the viability of GBC-SD or NOZ cells
attenuated by circMTO1 knockdown (Fig.
2D) (GBC-SD, sh-circMTO1-1 vs. sh-NC, P=0.0005; sh-circMTO1-1 +
miR-219a-5p inhibitor vs. sh-circMTO1-1, P=0.0036; NOZ,
sh-circMTO1-1 vs. sh-NC, P=0.0007; sh-circMTO1-1 + miR-219a-5p
inhibitor vs. sh-circMTO1-1, P=0.0083). Western blot analysis
revealed that cleaved caspase-3 protein expression was upregulated
in circMTO1 knockdown cells, the effects of which were reversed
following transfection with miR-219a-5p inhibitor (Fig. 2E). In addition, Annexin V staining
demonstrated that transfection with miR-219a-5p inhibitor decreased
the apoptotic rate of GBC-SD or NOZ cells enhanced by circMTO1
knockdown (Fig. 2F and G) (GBC-SD,
sh-circMTO1-1 vs. sh-NC, P=0.0072; sh-circMTO1-1 + miR-219a-5p
inhibitor vs. sh-circMTO1-1, P=0.032; NOZ, sh-circMTO1-1 vs. sh-NC,
P=0.0007; sh-circMTO1-1 + miR-219a-5p inhibitor vs. sh-circMTO1-1,
P=0.014). Notably, transfection with miR-219a-5p inhibitor enhanced
sh-circMTO1-modulated tube formation of HDLECs (Fig. 2H and I) (GBC-SD, sh-circMTO1-1 vs.
sh-NC, P=0.0002; sh-circMTO1-1 + miR-219a-5p inhibitor vs.
sh-circMTO1-1, P=0.0003; NOZ, sh-circMTO1-1 vs. sh-NC, P=0.0006;
sh-circMTO1-1 + miR-219a-5p inhibitor vs. sh-circMTO1-1, P=0.0003).
The results of the Transwell assay indicated that circMTO1
knockdown decreased migration of NOZ cells, which was reversed
following transfection with miR-219a-5p inhibitor (Fig. 2J). Collectively, these results
suggest that miR-219a-5p serves as a downstream effector for
circMTO1 in GBC cells.
miR-219a-5p regulates the TGF-β/Smad
pathway and EGFR expression
To further investigate the detailed downstream
pathway responsible for miR-219a-5p-associated GBC progression,
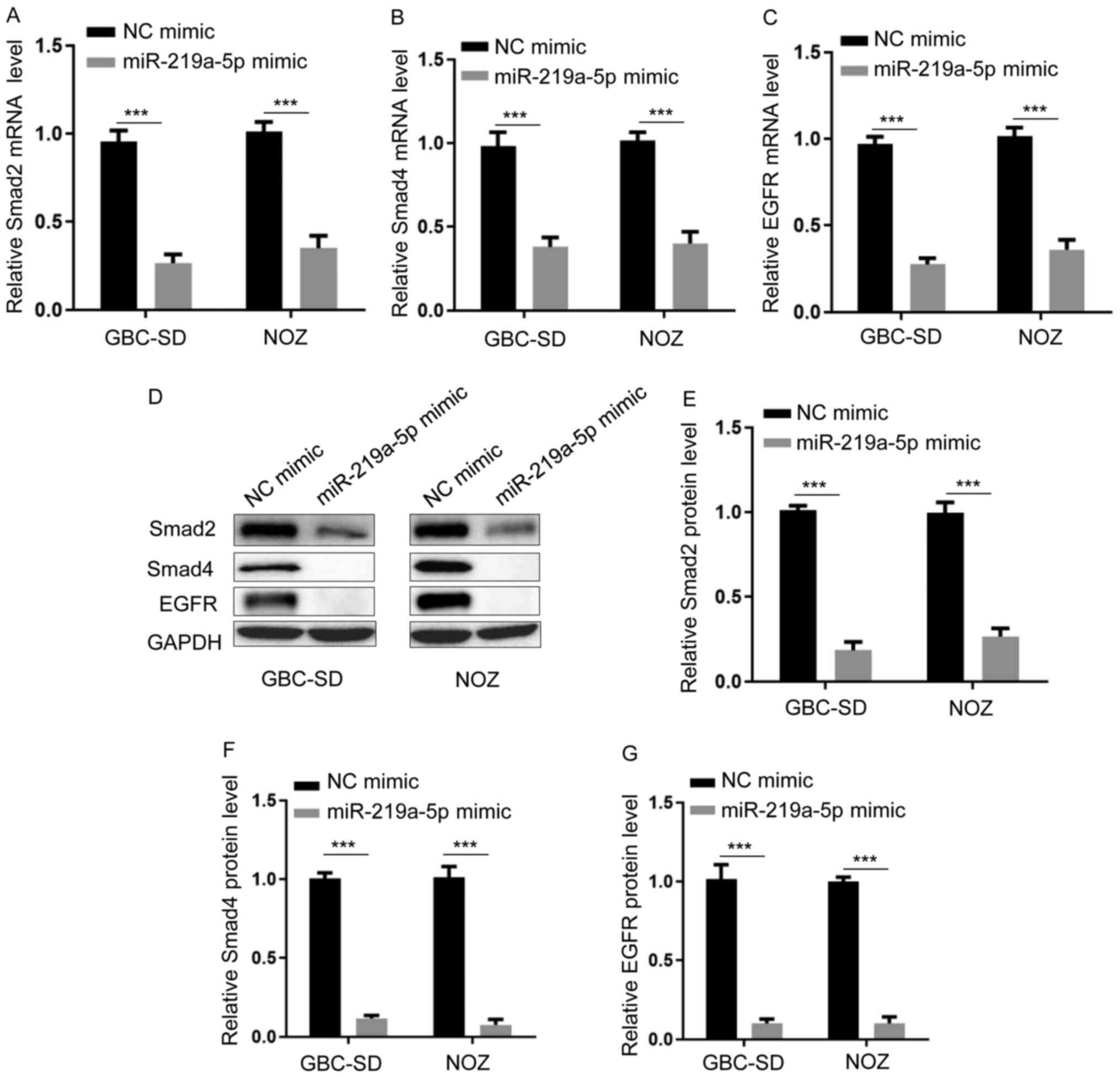
TargetScan software was used to detect Smad2/4 and EGFR expression.
RT-qPCR analysis demonstrated that transfection with miR-219a-5p
mimic inhibited the expression levels of Smad2, Smad4 and EGFR
(Fig. 3A-C) (Fig. 3A, GBC-SD, miR-219a-5p mimic vs. NC
mimic, P=0.0003; NOZ, miR-219a-5p mimic vs. NC mimic, P=0.0005.
Fig. 3B, GBC-SD, miR-219a-5p mimic
vs. NC mimic, P=0.0004; NOZ, miR-219a-5p mimic vs. NC mimic,
P=0.0008. Fig. 3C, GBC-SD,
miR-219a-5p mimic vs. NC mimic, P=0.0005; NOZ, miR-219a-5p mimic
vs. NC mimic, P=0.0004). Similarly, western blot analysis
demonstrated that transfection with miR-219a-5p mimic decreased the
protein expression levels of Smad2, Smad4 and EGFR (Fig. 3D-G) (Fig.
3E, GBC-SD, miR-219a-5p mimic vs. NC mimic, P=0.0002; NOZ,
miR-219a-5p mimic vs. NC mimic, P=0.0003. Fig. 3F, GBC-SD, miR-219a-5p mimic vs. NC
mimic, P=0.0004; NOZ, miR-219a-5p mimic vs. NC mimic, P=0.0007.
Fig. 3G, GBC-SD, miR-219a-5p mimic
vs. NC mimic, P=0.0006; NOZ, miR-219a-5p mimic vs. NC mimic,
P=0.0005). Taken together, these results suggest that miR-219a-5p
modulates the TGF-β/Smad pathway and EGFR expression.
circMTO1 is closely associated with
GBC progression
To further determine whether circMTO1 plays a role
in GBC, GBC datasets were downloaded from TCGA database. As
expected, the results demonstrated that circMTO1 expression was
higher in patients with GBC compared with healthy individuals
(P=0.018; Fig. 4A). Notably,
patients with low circMTO1 expression had prolonged overall
survival times than those with high circMTO1 expression (P=0.0069;
Fig. 4B). In addition, patients with
high circMTO1 expression were associated with advanced disease
(stage II vs. stage I; P=0.024; stage IV vs. stage II, P=0.035;
Fig. 4C). Finally, we proposed the
working model for circMTO1-induced GBC progression via miR-219a-5p
(Fig. 4D). Collectively, these
results suggest that circMTO1 is closely associated with GBC
progression.
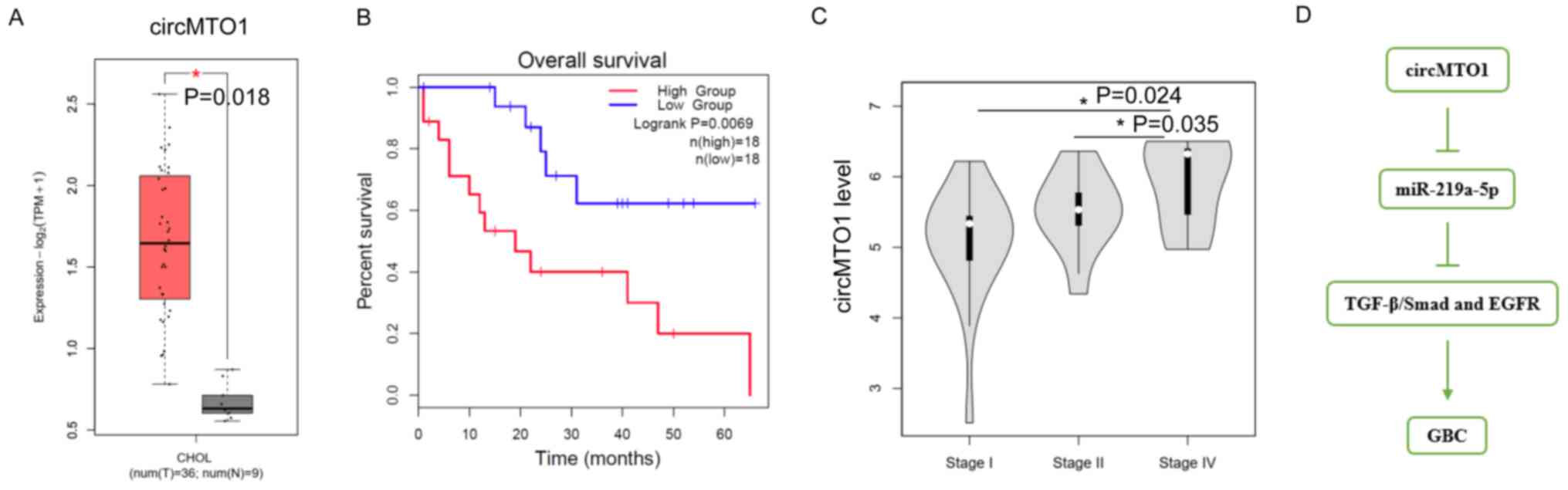 | Figure 4.circMTO1 is closely associated with
GBC progression. (A) circMTO1 expression in clinical normal tissues
(n=9) and GBC tissues (n=36). (B) Overall survival analysis of
patients with GBC, with high or low circMTO1 expression levels. (C)
circMTO1 expression was measured at different stages of GBC. Stage
I, the tumor has grown into the lamina propria or the muscle layer
(muscularis); stage II, the cancer has grown through the muscle
layer into the fibrous tissue on the side of the peritoneum or
liver, but has not invaded the liver, and stage IV, the tumor has
grown into one of the main blood vessels leading into the liver
(portal vein or hepatic artery) or it has grown into 2 or more
structures outside of the liver. (D) Mechanistic model of
circMTO1-induced GBC. *P<0.05. circMTO1, circular mitochondrial
translation optimization 1 homologue; GBC, gallbladder cancer; miR,
microRNA; TGF, transforming growth factor; EGFR, epidermal growth
factor receptor. |
Discussion
The results of the present study confirmed that
circMTO1 interacts with miR-219a-5p to promote the TGF-β/Smad
signaling pathway and EGFR expression in GBC cells. To the best of
our knowledge, the present study was the first to demonstrate that
circMTO1 may serve as an oncogene in patients with GBC. The results
demonstrated that circMTO1 expression was dysregulated in GBC cells
and tumors. In addition, circMTO1 was involved in GBC progression.
Based on these findings, circMTO1 may serve as a biomarker or
target for diagnosis and treatment of GBC.
circRNAs are generated by back-splicing of 3′ and 5′
splice sites (23). As circRNAs are
ubiquitously expressed in several tissues, they play important
roles across various biological processes (24,25).
Several circRNAs, including circERBB2, circFOXP1 and circHIPK3,
have been reported to regulate GBC progression (26–28).
Consistent with these findings, the results of the present study
demonstrated that circMTO1 knockdown decreased cell viability and
tube formation of HDLECs, while promoting the apoptosis of GBC
cells. Notably, a few studies have reported that overexpression of
circMTO1 enhances the apoptosis of gastric cancer cells and
hepatoma cells (8,29). These findings oppose the results of
the present study, thus it was hypothesized that circMTO1 may exert
different roles in different types of cancer.
miRNAs play key roles in different types of human
cancer by modulating gene expression levels (16). It has been reported that circRNAs
sponge and decoy miRNAs in cancer cells (30). The results of the present study
demonstrated that circMTO1 negatively regulated miR-219a-5p
expression. Previous studies have verified that miR-92, miR-9 and
miR-6893 are downstream effectors in different types of cancer
(11,22,31).
Thus, the results of the present study broaden the interacting
spectrum of circMTO1. The results presented here demonstrated that
miR-219a-5p inhibitor attenuated circMTO1 knockdown-induced
apoptosis, which is consistent with previous studies (19,20).
Long et al (32) reported
that miR-219-5p targets B-cell lymphoma 2 (Bcl-2) to inhibit
melanoma growth and metastasis. Thus, it was hypothesized that
circMTO1 interacts with miR-219-5p to regulate GBC progression by
targeting anti-apoptotic genes, such as Bcl-2.
TGF-β signaling plays a key role in tumor
progression, including epithelial-to-mesenchymal transition (EMT),
via Smads and the MAPK pathway (33). A recent study suggested that
TGF-β/Smad signaling promotes GBC metastasis by upregulating the
miR-182/CADM1 axis (34). Smads
exert their roles via phosphorylation (35). The results of the present study
demonstrated that miR-219a-5p decreased Smad2/4 expression levels
at the transcriptional level. Thus, detection of Smad 2/4
phosphorylation levels were not necessary and significant. In
addition, miR-219a-5p downregulated EGFR expression in GBC cells,
which was consistent with a previous study, suggesting that
miR-219a-5p regulates EGFR to affect EMT of ovarian cancer cells
(36). Notably, Shen et al
(37) demonstrated that activation
of the EGFR signaling pathway promotes gallbladder cancer invasion
and metastasis. The authors reported that PLEK2 interacts with the
kinase domain of EGFR and suppresses EGFR ubiquitination by c-CBL,
which results in constitutive activation of EGFR signaling and
elevated expression of the downstream effector, CCL2.
In conclusion, the results of the present study
suggest that circMTO1 contributes to GBC progression via the
miR-219a-5p/Smad/EGFR axis. This novel mechanism can be used to
develop targeted therapeutic strategies for patients with GBC.
However, the present study is not without limitations. First,
further studies are required to determine the role of circMTO1 in
different models (other GBC cell lines and animal models).
Secondly, although the present study demonstrated that miR-219a-5p
negatively regulated TGF-β signaling and EGFR, further studies are
required to determine whether other downstream targets are involved
in the circMTO1/miR-219a-5p axis-modulated GBC progression.
Acknowledgements
Not applicable.
Funding
The present study was supported by Lanzhou Science
and Technology Development Guide Project (grant no.
2019-ZD-21).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
PW, CZ and YD designed the present study and
performed the experiments. DL, LW and DZ analyzed the data and
prepared the figures. PW and LW drafted the initial manuscript. DL,
DZ and LW confirmed the authenticity of all the raw data. PW and YD
reviewed and edited the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shaffer EA: Gallbladder cancer: The
basics. Gastroenterol Hepatol (N Y). 4:737–741. 2008.PubMed/NCBI
|
|
2
|
Sachs TE, Akintorin O and Tseng J: How
should gallbladder cancer be managed? Adv Surg. 52:89–100. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kakaei F, Beheshtirouy S, Nejatollahi SM,
Zarrintan S and Mafi MR: Surgical treatment of gallbladder
carcinoma: A critical review. Updates Surg. 67:339–351. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lazcano-Ponce EC, Miquel JF, Munoz N,
Herrero R, Ferrecio C, Wistuba II, Alonso de Ruiz P, Aristi Urista
G and Nervi F: Epidemiology and molecular pathology of gallbladder
cancer. CA Cancer J Clin. 51:349–364. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Misra S, Chaturvedi A, Misra NC and Sharma
ID: Carcinoma of the gallbladder. Lancet Oncol. 4:167–176. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patop IL and Kadener S: circRNAs in
cancer. Curr Opin Genet Dev. 48:121–127. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo JU, Agarwal V, Guo H and Bartel DP:
Expanded identification and characterization of mammalian circular
RNAs. Genome Biol. 15:4092014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Song R, Li Y, Hao W, Yang L, Chen B, Zhao
Y, Sun B and Xu F: Circular RNA MTO1 inhibits gastric cancer
progression by elevating PAWR via sponging miR-199a-3p. Cell Cycle.
19:3127–3139. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ge Z, Li LF, Wang CY, Wang Y and Ma WL:
circMTO1 inhibits cell proliferation and invasion by regulating
Wnt/β-catenin signaling pathway in colorectal cancer. Eur Rev Med
Pharmacol Sci. 22:8203–8209. 2018.PubMed/NCBI
|
|
10
|
Li Y, Wan B, Liu L, Zhou L and Zeng Q:
Circular RNA circMTO1 suppresses bladder cancer metastasis by
sponging miR-221 and inhibiting epithelial-to-mesenchymal
transition. Biochem Biophys Res Commun. 508:991–996. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang X, Zhong B, Zhang W, Wu J and Wang
Y: Circular RNA circMTO1 inhibits proliferation of glioblastoma
cells via miR-92/WWOX signaling pathway. Med Sci Monit.
25:6454–6461. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang X, Lin YK, Lu ZL and Li J: Circular
RNA circ-MTO1 serves as a novel potential diagnostic and prognostic
biomarker for gallbladder cancer. Eur Rev Med Pharmacol Sci.
24:8359–8366. 2020.PubMed/NCBI
|
|
13
|
Shi CC, Pan LY, Peng ZY and Li JG:
circMTO1 attenuated acute kidney injury through regulating miR-337.
Inflammation. 43:1304–1311. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Braicu C, Zimta AA, Harangus A, Iurca I,
Irimie A, Coza O and Berindan-Neagoe I: The function of non-coding
RNAs in lung cancer tumorigenesis. Cancers (Basel). 11:6052019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boufraqech M, Nilubol N, Zhang L, Gara SK,
Sadowski SM, Mehta A, He M, Davis S, Dreiling J, Copland JA, et al:
miR30a inhibits LOX expression and anaplastic thyroid cancer
progression. Cancer Res. 75:367–377. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rao C, Miao X, Zhao G, Zhang C, Shen H,
Dong C and Yang M: miR-219a-5p enhances cisplatin sensitivity of
human non-small cell lung cancer by targeting FGF9. Biomed
Pharmacother. 114:1086622019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu K, Shi J, Mo D, Yang Y, Fu Q and Luo Y:
miR-219a-1 inhibits colon cancer cells proliferation and invasion
by targeting MEMO1. Cancer Biol Ther. 21:1163–1170. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wei T, Cheng S, Fu XN and Feng LJ:
miR-219a-5p enhances the radiosensitivity of non-small cell lung
cancer cells through targeting CD164. Biosci Rep.
40:BSR201927952020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan J, Bu X, Li Z, Wu J, Wang C, Li D,
Song J and Wang J: Screening the expression of several miRNAs from
TaqMan low density array in traumatic brain injury: miR-219a-5p
regulates neuronal apoptosis by modulating CCNA2 and CACUL1. J
Neurochem. 150:202–217. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maruyama T, Nishihara K, Umikawa M,
Arasaki A, Nakasone T, Nimura F, Matayoshi A, Takei K, Nakachi S,
Kariya KI and Yoshimi N: MicroRNA-196a-5p is a potential prognostic
marker of delayed lymph node metastasis in early-stage tongue
squamous cell carcinoma. Oncol Lett. 15:2349–2363. 2018.PubMed/NCBI
|
|
22
|
Chen M, Ai G, Zhou J, Mao M, Li H and Guo
J: circMTO1 promotes tumorigenesis and chemoresistance of cervical
cancer via regulating miR-6893. Biomed Pharmacother.
117:1090642019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N and
Kadener S: circRNA biogenesis competes with pre-mRNA splicing. Mol
Cell. 56:55–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li R, Jiang J, Shi H, Qian H, Zhang X and
Xu W: circRNA: A rising star in gastric cancer. Cell Mol Life Sci.
77:1661–1680. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yan Z, Xiao Y, Chen Y and Luo G: Screening
and identification of epithelial-to-mesenchymal transition-related
circRNA and miRNA in prostate cancer. Patho Res Pract.
216:1527842020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang X, He M, Huang S, Lin R, Zhan M,
Yang D, Shen H, Xu S, Cheng W, Yu J, et al: Circular RNA circERBB2
promotes gallbladder cancer progression by regulating
PA2G4-dependent rDNA transcription. Mol Cancer. 18:1662019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kai D, Yannian L, Yitian C, Dinghao G, Xin
Z and Wu J: Circular RNA HIPK3 promotes gallbladder cancer cell
growth by sponging microRNA-124. Biochem Biophys Res Commun.
503:863–869. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang S, Zhang Y, Cai Q, Ma M, Jin LY, Weng
M, Zhou D, Tang Z, Wang JD and Quan Z: Circular RNA FOXP1 promotes
tumor progression and Warburg effect in gallbladder cancer by
regulating PKLR expression. Mol Cancer. 18:1452019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang J, Tan Q, Wang W and Yu J: Mechanism
of the regulatory effect of overexpression of circMTO1 on
proliferation and apoptosis of hepatoma cells via miR-9-5p/NOX4
axis. Cancer Manag Res. 12:3915–3925. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kristensen LS, Hansen TB, Veno MT and
Kjems J: Circular RNAs in cancer: Opportunities and challenges in
the field. Oncogene. 37:555–565. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han D, Li J, Wang H, Su X, Hou J, Gu Y,
Qian C, Lin Y, Liu X, Huang M, et al: Circular RNA circMTO1 acts as
the sponge of microRNA-9 to suppress hepatocellular carcinoma
progression. Hepatology. 66:1151–1164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Long J, Menggen Q, Wuren Q, Shi Q and Pi
X: miR-219-5p inhibits the growth and metastasis of malignant
melanoma by targeting BCL-2. Biomed Res Int. 2017:90325022017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bao RF, Shu YJ, Hu YP, Wang XA, Zhang F,
Liang HB, Ye YY, Li HF, Xiang SS, Weng H, et al: miR-101 targeting
ZFX suppresses tumor proliferation and metastasis by regulating the
MAPK/Erk and Smad pathways in gallbladder carcinoma. Oncotarget.
7:22339–22354. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qiu Y, Luo X, Kan T, Zhang Y, Yu W, Wei Y,
Shen N, Yi B and Jiang X: TGF-β upregulates miR-182 expression to
promote gallbladder cancer metastasis by targeting CADM1. Mol
Biosyst. 10:679–685. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Syed V: TGF-β signaling in cancer. J Cell
Biochem. 117:1279–1287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang L, Yu M and Zhao S: lncRNA MEG3
modified epithelial-mesenchymal transition of ovarian cancer cells
by sponging miR-219a-5p and regulating EGFR. J Cell Biochem.
120:17709–17722. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shen H, He M, Lin R, Zhan M, Xu S, Huang
X, Xu C, Chen W, Yao Y, Mohan M and Wang J: PLEK2 promotes
gallbladder cancer invasion and metastasis through EGFR/CCL2
pathway. J Exp Clin Cancer Res. 38:2472019. View Article : Google Scholar : PubMed/NCBI
|