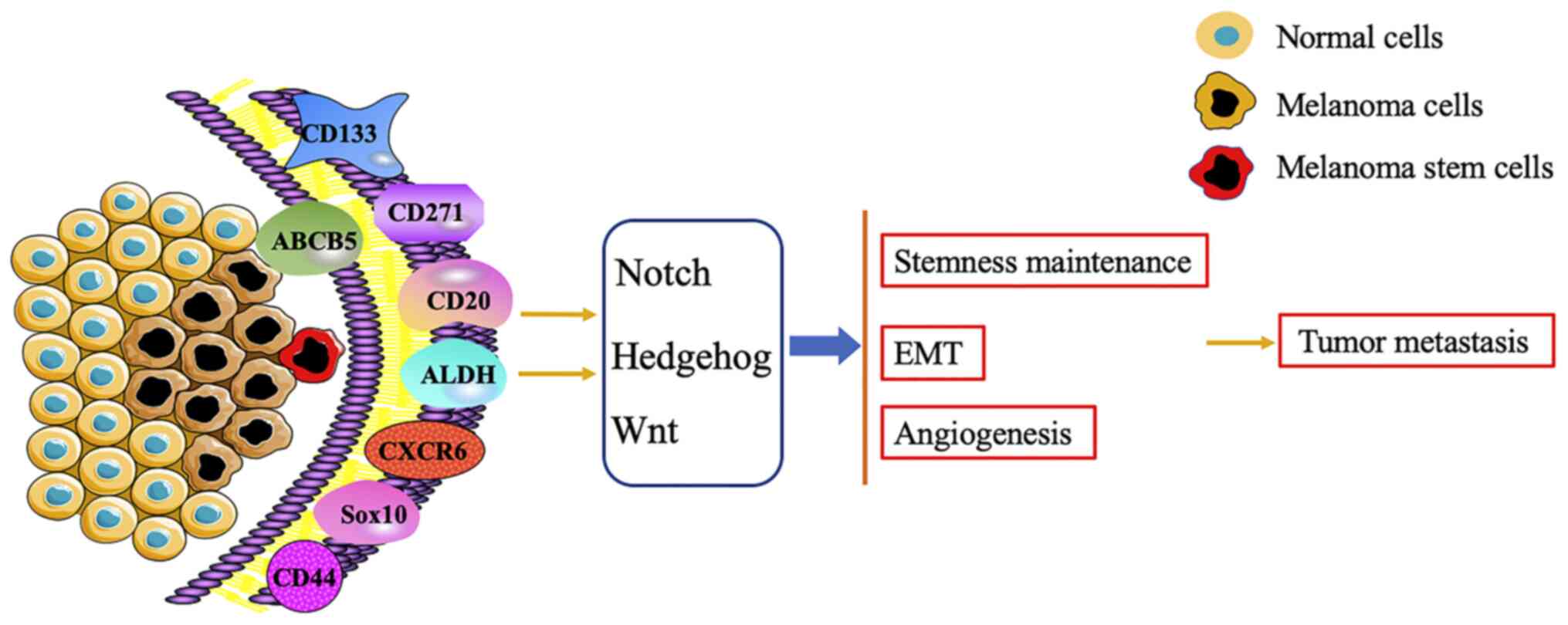
Melanoma is a malignant tumor of melanocytes that
typically arises in the skin. It is highly malignant, prone to
metastasis and recurrence, accounting for ~75% of skin
cancer-associated mortality (1). The
treatment of advanced or metastatic melanoma is particularly
challenging, and there is a high tendency for patients to relapse
and become resistant to current therapeutic agents (2). Although molecular targeted therapy and
immunotherapy have been reported to prolong the survival time of
patients, most patients will develop drug resistance within one
year (3–5), resulting in melanoma metastasis. The
existence of melanoma stem cells (MSCs) is one of the potential
causes of melanoma invasion and metastasis.
Cancer stem cells (CSCs) have been shown to be an
integral part of solid tumors (6).
Furthermore, CSCs exhibit distinctive and remarkable capacities of
self-renewal, differentiation and proliferation, which are believed
to have a key role in all aspects of carcinogenesis, including
tumor recurrence and metastasis (7,8).
Previous studies have demonstrated that the essence of tumor
metastasis is the transfer and homing of CSCs (6,9). In the
last decade, with the rise of CSCs, several lines of evidence
suggested that CSCs may be at the origin of tumor metastasis
(10–12). Interestingly, the CSC subpopulation
is responsible for many aspects of tumorigenesis and has been
reported to serve a crucial role in melanoma development,
progression, drug resistance and metastasis (13,14). The
fact that the CSCs are resistant to chemotherapy also explains that
traditional anticancer drugs can only inhibit or narrow the tumor,
but not completely eradicate it, leading to tumor metastasis and
recurrence (15,16). In addition, CSCs have been reported
to express a variety of biomarkers, such as CD34, aldehyde
dehydrogenase 1 (ALDH1), CD271, CD44 and lysine demethylase 5B
(JARID1B); however, none of these markers have been shown to be
CSCs-specific (17–19). Several potential biomarkers of CSCs
have been demonstrated to be expressed by certain human solid
tumors such as melanoma (16),
including CD133, ATP binding cassette subfamily B member 5 (ABCB5),
CD271, CD20 and ALDH (Table I).
Although the mechanism of MSCs promoting tumor metastasis and
recurrence has not been fully elucidated (13,20,21), the
activation of the signaling pathways, including Notch, Hedgehog and
Wnt, is modulated by these biomarkers to maintain the
characteristic of MSCs, thus promoting angiogenesis and
epithelial-mesenchymal transition (EMT) of melanoma and
accelerating tumor metastasis (7,20,22)
(Fig. 1). It is therefore important
to determine the role of MSCs in the invasion and metastasis of
melanoma.
As a surface protein with unknown function, CD133
(prominin-1) can be expressed on human melanoma, but hardly be
detected in normal skin (23). It
has been reported that CD133, a stem cell-related surface antigen,
is closely related to tumor proliferation and progression in
various types of tumor, including melanoma (15,24,25).
Furthermore, CD133+ melanoma cells exhibit 411
upregulated genes, which are associated with angiogenesis, adhesion
and migration (26). Melanoma
CD133+ CSCs have the potential to initiate tumor
progression. In addition, CD133 can activate MAPK signal pathway
through notch receptor 1 (Notch1), regulate the expression of
vascular endothelial growth factor (VEGF) and matrix
metalloproteinases and promote the interaction of tumor endothelial
cells, leading to the increase of tumor angiogenesis and lung
metastasis (26). Furthermore, the
expression of CD133, p-p38 and p-MEK3/6 in metastatic melanoma is
significantly higher than that in paracancerous tissues (26,27),
suggesting that Notch1 and its MAPK signaling pathway network might
be considered as potential targets for MSCs-mediated melanoma
targeted therapy (28,29). Furthermore, it is generally known
that tumor angiogenesis is an essential factor for tumor growth and
metastasis. In particular, CD133+ and ABCB5+
MSCs are involved in the formation of perivascular niches (30). When RNAi is used to block the
expression of CD133, the tumorigenicity of stem cells in
vivo is significantly decreased and the expression of CD144 and
ABCB5 that are closely related to the vascular microenvironment is
downregulated (30). However,
ABCB5− cells lose the ability to form CD144+
angiogenic mimicry. It has been reported that
CD133+/ABCB5+ MSCs exist in CD144+
angiogenic mimicry, suggesting that CD133+ MSCs could
promote tumor metastasis by increasing the formation of angiogenic
mimicry and specific vascular microenvironment (30). Zimmerer et al (31) demonstrated by fluorescence microscopy
that CD133+ melanoma D10 cells xenograft into nude mice
can trigger an important angiogenesis process. A clonal dominance
of a CD133+ population exists within the hierarchy of
cells in cutaneous tissues from patients that have undergone
successive progressive stages of melanoma, from primary to
metastatic lesions (32). In
addition, in the CD133− melanoma cells subpopulation,
exposure to taxol induces the activation of apoptosis
signal-regulating kinase1/c-jun-N-terminal kinase, p38 and ERK
pathways and Bax expression; however, in CD133+ cells,
taxol only enhances the activity of the ERK pathway (33). Furthermore, it was demonstrated that
the expression of CD133+ in patients with recurrent and
metastatic melanoma is twice higher than in patients with primary
melanoma (32). Mechanistically,
CD133 downregulation in human metastatic melanoma cells can
decrease the capacity of sphere-forming and the metastasis
potential of melanocytes (28). In
addition, CD133, is closely related to the expression of certain
tumor associated antigens and could thus serve as a potential
target for immunotherapy (34,35).
CD133 may therefore be considered as a predictive marker of
melanoma and as a potential therapeutic target of high-risk
melanoma.
ABCB5 is a member of the ATP binding cassettes (ABC)
transporter family and a regulator of cell membrane potential.
ABCB5 can regulate the fusion of normal skin progenitor cells and
is considered as one marker of MSCs (36,37). It
is now well accepted that ABC transporters mediate multidrug
resistance through drug efflux in cancer cells, which is usually
related to cancer stem cells (38–40).
In vivo genetic lineage tracking demonstrated a specific
capacity of ABCB5+ sub-populations for self-renewal and
differentiation, as ABCB5+ cancer cells generate both
ABCB5+ and ABCB5− progeny whereas
ABCB5− tumor populations give rise, at lower rates,
exclusively to ABCB5− cells (37). Subsequently, ABCB5, a marker of MSCs,
has high tumorigenicity potential and is co-expressed with other
MSCs markers, such as CD133 (39,41,42).
ABCB5+ tumor cells detected in human melanoma patients
show a primitive molecular phenotype and correlate with clinical
melanoma progression (43). Analysis
of some clinical data demonstrated that overexpression of ABCB5 can
promote tumor progression, and that ABCB5 expression is usually low
in pigmented nevus subpopulation but high in primary and metastatic
melanoma cell subpopulation (37,44). In
serial human-to-mouse xenotransplantation experiments,
ABCB5+ melanoma cells possess greater tumorigenic
capacity than ABCB5− bulk populations (44,45). Ma
and Frank (45) reported that
ABCB5+ melanoma cells exist in the peripheral blood of
patients with melanoma. Subsequently, transplanting these cells to
Nod/SCID/IL2 mice can induce distant metastasis, and the degree of
metastasis is directly proportional to the number of
ABCB5+ melanoma cells transplanted.
CD271, also known as low affinity nerve growth
factor receptor or p75NTR, is a characteristic marker of MSCs
(36,49). High Expression of CD271 has been
reported in numerous human neural-crest-derived tissues and in some
human cancers, including melanomas (50–52).
Previous studies demonstrated that CD271+ melanoma cells
have a higher tumorigenicity potential than CD271− cells
and are involved in the metastasis of melanoma in vivo,
especially in peripheral nerves (49,53–55).
CD271 is the most reliable cell surface marker for the
identification of melanoma heterogeneous subsets (49). In addition, not only CD271 marks
dedifferentiated melanoma cells emerging, for instance, through
TGFβ-mediated EMT, BRAF inhibitor-induced reprogramming (56) or in response to immunotherapies
(57), but it is also functionally
involved in promoting low rates of proliferation and high
metastatic capacity (58). In fully
immunocompromised mouse models, including
NOD/SCID/IL2rγnull mice, melanoma cells expressing the
neurotrophin receptor CD271 have a higher tumor-initiation capacity
than CD271− cells, although the negative fraction is
also able to generate tumors in this mouse model (49). Furthermore, CD271+
melanoma cells are prone to form liver and lung metastasis in mice,
whereas CD271− melanoma cells rarely form metastasis
(59). In human samples, the
proportion of CD271/ SRY-Box transcription factor 10 (Sox10)
positive cells is significantly increased in metastatic melanoma
compared with primary melanoma cells (49,60).
Interestingly, CD271 inactivation not only results in decreased
melanoma cell survival, but also in increased sensitivity to BRAF
inhibitor treatment, suggesting that CD271 might confer therapy
resistance (61). Schnegg et
al (62) demonstrated that
CD133+ and CD271+ MSCs accumulate in the
perivascular niche that melanoma cells with formation of angiogenic
mimicry positively express CD271. Similarly, in the human uveal
melanoma cell line c918 cultured in 2D and 3D cultures, evaluation
of CD271 expression through immunofluorescence showed that CD271 is
expressed on the tumor cells that form the vasculogenic mimicry
(63). It was confirmed that
melanoma cells that form the vasculogenic mimicry acquire the tumor
stem cell-like phenotype and participate in angiogenesis (63).
A previous study reported that the number of MSCs
positive for ABCB5, CD271 and receptor activator of nuclear factor
κ B in circulating tumor cells (CTCs) of patients with advanced
melanoma is significantly increased (64). In addition, CTCs are highly enriched
in MSCs, which is a vital parameter inducing the formation of
distant secondary tumors (64).
Furthermore, melanoma-associated antigens, such as melanoma antigen
recognized by T-cells 1, are less exposed to CD271 and ABCB5
positive melanoma cells, which supports the hypothesis that
melanoma cells expressing the MSCs makers can escape the attack of
host immune system (65). A previous
study demonstrated that CD271 overexpression in melanoma cells
inhibits the production of melanoma-specific cytotoxic T
lymphocytes (CTLs), and that interferon-γ from CTLs subsequently
triggers the expression of CD271 in melanoma cells, downregulating
therefore the production of melanoma antigens (66). Similarly, it was reported that
melanoma cells highly expressing CD271 are associated with high
tumor metastasis potential and poor prognosis of patients (67).
In melanoma, numerous subpopulations with the
capacity of self-renewal, differentiation, tumorigenicity and/or
drug resistance have been described (49,68–70),
including one subpopulation expressing the B cell marker CD20
(36,71–73).
Further characterization with respect to melanoma-associated
antigen indicated that a more primitive melanoma phenotype revealed
overexpression of CD20 (16).
Importantly, CD20 was initially identified on a small percentage of
human melanoma cells when cultured in embryonic stem cell medium
and found on nonadherent spheres. These CD20+ melanoma
cells with the ability of self-renewal and differentiation followed
the definition of tumor stem cells. Consistent with the view that
cancer stem cells occupy a small part of tumors, CD20+
cells only account for ~2% of the total number of melanoma cells
(74). However, CD20+
melanoma cells were demonstrated to be highly tumorigenic in
vivo following xenotransplantation, suggesting that these cells
exhibit tumor-initiating capacity (16). A previous study reported that the
melanoma cells WM115 in the non-adherent form (melanoma spheroid
cells) express a higher level of CD20 compared with adherent WM115
cells and that it is more likely to develop tumor when melanoma
spheroid cells are transplanted into mice (16). Similarly to all stem cells, melanoma
spheroid cells are also capable of proliferation, differentiation
and self-renewal (16). In the late
stage of metastatic melanoma, the effect of targeted therapy with
anti-CD20 monoclonal antibody is more significant than that of
non-targeted therapy and can even achieve a clinical complete
response (75–77). In melanoma patients resistant to
chemotherapeutic drugs, intratumoral injections of rituximab, which
is the specific antibody against CD20, induces a regression of
tumor growth, accompanied by a significant decrease in serum levels
of inflammatory markers (77).
Although there is no clear evidence that CD20+ MSCs are
directly involved in melanoma metastasis, the stem cell-like
characteristics of CD20+ melanoma cells have been
confirmed, and their high tumorigenicity and migration ability are
the main reasons for tumor progression (16,76).
CD20 may therefore be considered as a new target for melanoma
treatment in the future.
ALDH represents a group of isoenzymes that can
oxidize acetaldehyde to acetic acid. The enzymatic activity of ALDH
has been used to identify stem or progenitor cells from various
malignancies including breast, colon and lung cancers (78–80).
According to previous studies, ALDH, which is a marker in many CSCs
(78), is associated with multidrug
resistance and immune tolerance of different types of solid tumor
(78,81–83), can
inhibit oxidative stress and enhance resistance to chemotherapeutic
drugs, such as oxazolidine, taxanes and platinum drugs (84–86). It
has been reported that melanoma cells with high expression of ALDH
exhibit MSCs characteristics (87,88).
Furthermore, ALDH-positive melanoma cells are more resistant to
chemotherapeutic agents, and silencing ALDH1A using small
interfering RNA can sensitize melanoma cells to drug-induced cell
death (88). ALDHhigh
cells (melanoma cells with high expression of ALDH) can produce
more melanoma clone spheres than ALDHlow cells (melanoma
cells with low expression of ALDH) (87); however, the ability of melanoma
formation in vivo is significantly inhibited following ALDH
silencing (88). For example,
following downregulation of aryl hydrocarbon receptor (AHR) and/or
ALDH1 in mouse melanoma B16F10 cells through retroviral
transduction, Contador et al (89) demonstrated that ALDH1 downregulation
could inhibit the metastasis of melanoma cells without AHR
expression, reduce the number of
CD133+/CD29+/CD44+ cells and the
size of melanospheres, confirming that overactivation of ALDH1
could promote the progression of melanoma in the context of AHR
deficiency. Similarly, ALDH1 silencing in melanoma cells using
short hairpin (sh)RNA significantly delays the appearance and
growth of xenograft melanoma and dramatically decreases the number
and load of metastases in mice (90). These studies confirm that targeting
ALDH1 may be considered as an effective strategy for the treatment
of advanced melanoma.
Sox10 is a key nuclear transcription factor involved
in the malignant transformation of melanocytes that has the
potential to be a marker of MSCs (91). The positive rate of Sox10 in sentinel
lymph node micrometastasis of melanoma is close to 100%, which is
significantly higher than other melanoma markers responsible for
melanoma metastasis, such as S100, HMB45 and Melania (91). Furthermore, it has also been reported
that the stem cell function of CD271+ melanoma cells may
be correlated to the CD271/Sox10 interaction network, although the
underlying mechanism remains unclear (59,92).
High levels of organic cation transporter (OCT)3/4, Nanog homeobox
(Nanog) and Sox10 are found in CD133+ transgenic mice
and human melanoma cells, and were demonstrated to promote tumor
neovascularization, indicating that Sox10 and other MSCs markers
can regulate each other and accelerate tumor progression (26). However, Sox10 silencing in human
melanoma cells suppresses neural crest stem cell properties,
inhibits cell proliferation and survival and completely abolishes
in vivo tumor formation (93). Sox10 may therefore represent a
promising target for the treatment of congenital naevi and melanoma
in human patients.
C-X-C motif chemokine receptor 6 (CXCR6) is also an
important marker of MSCs. Compared with ATP binding cassette
subfamily G member 2 (ABCG2)+ melanoma cells,
CXCR6+ melanoma cells can produce larger tumors in a
shorter time, and ABCG2+ and CXCR6+
double-positive cells have a higher tumorigenic potential than
ABCG2+ or CXCR6+ single-positive melanoma
cells in promoting tumor metastasis (94).
CD44 has been used as a specific marker of MSCs in
preclinical previous studies (95,96). It
has been reported that blocking insulin-like growth factor-1 can
prevent the metastatic and EMT processes of melanoma cells by
downregulating the stem cell markers Sox2, OCT3/4, CD44 CD133 and
deleting stem cell functional characteristics (97). CD44 may therefore serve an important
role in melanoma metastasis.
The process of angiogenesis is an important hallmark
of the growth and progression of tumors, including melanoma
(98). Evidence has shown that tumor
vessels are derived from capillaries and veins in the host tissue,
following activation by pro-angiogenic factors of pre-existing
endothelial cells migrating into the tumor and developing into new
vessel networks (99). The
endothelial progenitor cells found in the peripheral circulation
also contribute to tumor angiogenesis by differentiating and
proliferating in a local tumor (100). Similarly, mesenchymal stem cells
promote the growth and angiogenesis of tumors thanks to their
self-renewal capacity, long-term viability and differentiation
potential toward diverse cell types (101–104).
For example, CD133+ glioma cells secrete higher levels
of VEGF than CD133− cells, which indicates that tumor
stem cells promote tumor angiogenesis (105). Furthermore, MSCs, which are
involved in tumor angiogenesis, can accelerate melanoma metastasis.
The renal and melanoma derived-CSCs are able to differentiate into
endothelial like cells when cultured in endothelial cell growth
specific medium (26,106). In addition, melanoma cells with
stem cell-like characteristics, particularly those locating at the
margin of the tumor, such as melanoma initiating cells, express and
deliver in the microenvironment several factors (including VEGF,
bFGF and PDGF) associated with angiogenesis (20). Since MSCs have high degree of
differentiation plasticity, they can contribute to the de
novo formation of tumor angiogenesis via a process named
vasculogenic mimicry (VM) (107).
Interestingly, tumor cells with abundant VM have a high plasticity,
and their ability to mimic vascular endothelial cells may be
related to the stemness of tumor cells (107,108).
It has been reported that increasing the expression of stem
cell-like genes in melanoma can improve the plasticity of tumor
cells (108,109). Furthermore, ABCB5+ MSCs
express specific endothelial and proangiogenic factors as well as
VE-cadherin, Tie2, VEGF and its receptors, which are specific
markers of VM (46). Consistently
with these observations, melanoma cells expressing the CD133 and
ABCG2 stem cells markers overexpress proangiogenic proteins, such
as VEGF and its receptor VEGFR-2, Tie2 and angiopoietin (27). Furthermore, CD271+ MSCs
were demonstrated to be associated with VM, through activation of
the VEGR receptor/PKC signaling pathway (64,110).
Thus, MSCs in angiogenesis contribute to melanoma growth and
metastasis.
The molecular markers of MSCs are of relative
specificity and can serve as targets of molecular targeted therapy.
They not only contribute to the removal of MSCs but also prevent
normal cells from being damaged, in order to achieve the highest
benefit of tumor therapy. For example, tubacin, which is an
inhibitor of histone deacetylase 6, can promote the release of
CD133+ exosomes in the human metastatic melanoma cells
FEMX–I, decrease the content of CD133 in cells and inhibit the
proliferation and clonogenesis of FEMX–I cells (111). In addition, monoclonal antibodies
against different epitopes of CD133 have exhibited a dose-dependent
cytotoxicity in FEMX–I cells (28,112).
Furthermore, CD133 monoclonal antibody can inhibit the
proliferation of FEMX–I melanoma cells and prevent the growth of
melanoma through its cytotoxic effect. It was also demonstrated
that CD133 downregulation by shRNA can decrease the appearance of
lung and spinal cord metastases from melanoma (28). In addition, andrographolide can block
the expression of notch1-dependent CD133 in melanoma cells and
decrease the activation of MAPK signaling pathway, leading thus to
inhibition of tumor growth, angiogenesis and metastasis (26). Similarly, the elimination of
chemoresistant ABCB5-positive cells may significantly inhibit the
overall growth of xenotransplanted melanomas by a selective
antibody (37). ABCB5 is involved in
the in vitro and in vivo survival of melanoma cells
following exposure to dacarbazine and the BRAF inhibitor
vemurafenib (113), and systemic
administration of anti-ABCB5 antibody inhibits melanoma
tumorigenesis in nude mice (37).
Furthermore, cuprous oxide nanoparticles can significantly decrease
the expression of Sox10 and CD271, which accelerates the apoptosis
and inhibits the tumorigenicity of CD271 overexpressing A375 and
WM266-4 cells (114). Lunasin can
decrease the expression level of ALDH, a marker of MSCs, and the
expression level of Nanog, a stem cell-related factor. Lunasin can
also promote the upregulation of microphthalmia associated
transcription factor that inhibits the colony-forming ability of
tumor cells and the growth of xenograft tumors, and can increase
the transformation from ALDHhigh to ALDHlow
melanoma cells (115). Furthermore,
magnolol can decrease the expression of CD271, CD166, JARID1B and
ABCB5 through reducing the expression level of notch 2, a
downstream target protein of HES-1, and of the cell cycle-related
protein cyclin D1, inhibiting therefore the proliferation of
melanoma cells and inducing their autophagy (116). In a clinical trial, rituximab, an
anti-CD20 monoclonal antibody, was used to treat CD20+
metastatic melanoma. The anti-CD20 monoclonal antibody can
eliminate CD20+ melanoma cells and increase the level of
peripheral B cells in patients with melanoma (77). In an in vivo and in
vitro study of melanoma cells, MSCs-induced angiogenesis
mimicry was shown be a potential biological target for some
anticancer compounds, such as the natural phytochemicals lupeol,
which can prevent tumor metastasis by inhibiting angiogenesis
(117). However, current research
mainly uses cell and animal models, and large-scale clinical trials
are required to support these conclusions.
The present review on MSCs provided some insights
into the metastasis, recurrence, drug resistance and treatment of
melanoma. MSCs serve a crucial role in the occurrence and
progression of melanoma, especially in tumor metastasis. MSCs can
promote tumor progression via MSCs-specific markers and the
subsequent regulation of related signaling pathways. It is
therefore crucial to fully understand the underlying mechanisms of
MSCs markers in the process of tumor metastasis, which would allow
the discovery of effective targets for the targeted therapy of
melanoma. By exploring MSCs markers and the related signal
transduction pathways, we believe that effective treatment
strategies to inhibit tumor metastasis or eradicate melanoma might
be discovered in the near future.
Not applicable.
This research was supported by the Project
Agreement for Science & Technology Development, Jilin Province
(grant no. 20200404135YY), the Youth Fund of National Natural
Science Foundation of China (grant no. 81801562), the Clinical
Research Foundation of First Hospital of Jilin University and the
Eighth Youth Developmental Foundation of First Hospital of Jilin
University (grant no. JDYY82017014).
Not applicable.
QY drafted the initial manuscript, and edited and
critically revised the manuscript. HJ and DW gave guidance on the
conception and design of the review. XS and SL were involved in
conceiving and designing the review. All authors have read and
approved the final manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Lo JA and Fisher DE: The melanoma
revolution: From UV carcinogenesis to a new era in therapeutics.
Science. 46:945–949. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Holderfield M, Deuker MM, McCormick F and
McMahon M: Targeting RAF kinases for cancer therapy: BRAF-mutated
melanoma and beyond. Nat Rev Cancer. 14:455–467. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chapman PB, Hauschild A, Robert C, Haanen
JB, Ascierto P, Larkin J, Dummer R, Garbe C, Testori A, Maio M, et
al: Improved survival with vemurafenib in melanoma with BRAF V600E
mutation. N Engl J Med. 364:2507–2516. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Flaherty KT, Robert C, Hersey P, Nathan P,
Garbe C, Milhem M, Demidov LV, Hassel JC, Rutkowski P, Mohr P, et
al: Improved survival with MEK inhibition in BRAF-mutated melanoma.
N Engl J Med. 367:107–114. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Christiansen SA, Khan S and Gibney GT:
Targeted therapies in combination with immune therapies for the
treatment of metastatic melanoma. Cancer J. 23:59–62. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nandy SB and Lakshmanaswamy R: Cancer stem
cells and metastasis. Prog Mol Biol Transl Sci. 151:137–176. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Parmiani G: Melanoma cancer stem cells:
Markers and functions. Cancers (Basel). 8:342016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fink J, Andersson-Rolf A and Koo BK: Adult
stem cell lineage tracing and deep tissue imaging. BMB Rep.
48:655–667. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ricci E, Mattei E, Dumontet C, Eaton CL,
Hamdy F, van der Pluije G, Cecchini M, Thalmann G, Clezardin P and
Colombel M: Increased expression of putative cancer stem cell
markers in the bone marrow of prostate cancer patients is
associated with bone metastasis progression. Prostate.
73:1738–1746. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Croker AK and Allan AL: Cancer stem cells:
Implications for the progression and treatment of metastatic
disease. J Cell Mol Med. 12:374–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li F, Tiede B, Massague J and Kang Y:
Beyond tumorigenesis: Cancer stem cells in metastasis. Cell Res.
17:3–14. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wickremesekera AC, Brasch HD, Lee VM,
Davis PF, Woon K, Johnson R, Tan ST and Itinteang T: Expression of
cancer stem cell markers in metastatic melanoma to the brain. J
Clin Neurosci. 60:112–116. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nguyen N, Couts KL, Luo Y and Fujita M:
Understanding melanoma stem cells. Melanoma Manag. 2:179–188. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klein WM, Wu BP, Zhao S, Wu H,
Klein-Szanto AJ and Tahan SR: Increased expression of stem cell
markers in malignant melanoma. Mod Pathol. 20:102–107. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fang D, Nguyen TK, Leishear K, Finko R,
Kulp AN, Hotz S, Van Belle PA, Xu X, Elder DE and Herlyn M: A
tumorigenic subpopulation with stem cell properties in melanomas.
Cancer Res. 65:9328–9337. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tirino V, Desiderio V, Paino F, De Rosa A,
Papaccio F, Fazioli F, Pirozzi G and Papaccio G: Human primary bone
sarcomas contain CD133+ cancer stem cells displaying
high tumorigenicity in vivo. FASEB J. 25:2022–2030. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Desiderio V, Papagerakis P, Tirino V,
Zheng L, Matossian M, Prince ME, Paino F, Mele L, Papaccio F,
Montella R, et al: Increased fucosylation has a pivotal role in
invasive and metastatic properties of head and neck cancer stem
cells. Oncotarget. 6:71–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Collins AT, Berry PA, Hyde C, Stower MJ
and Maitland NJ: Prospective identification of tumorigenic prostate
cancer stem cells. Cancer Res. 65:10946–10951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Marzagalli M, Raimondi M, Fontana F,
Montagnani Marelli M, Moretti RM and Limonta P: Cellular and
molecular biology of cancer stem cells in melanoma: Possible
therapeutic implications. Semin Cancer Biol. 59:221–235. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kumar D, Gorain M, Kundu G and Kundu GC:
Therapeutic implications of cellular and molecular biology of
cancer stem cells in melanoma. Mol Cancer. 16:72017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee N, Barthel SR and Schatton T: Melanoma
stem cells and metastasis: Mimicking hematopoietic cell
trafficking? Lab Invest. 94:13–30. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shakhova O and Sommer L: Testing the
cancer stem cell hypothesis in melanoma: The clinics will tell.
Cancer Lett. 338:74–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M, Peschle C and De Maria R: Identification and
expansion of human colon-cancer-initiating cells. Nature.
445:111–115. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang D, Tang DG and Rycaj K: Cancer stem
cells: Regulation programs, immunological properties and
immunotherapy. Semin Cancer Biol. 52:94–106. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kumar D, Kumar S, Gorain M, Tomar D, Patil
HS, Radharani NNV, Kumar TVS, Patil TV, Thulasiram HV and Kundu GC:
Notch1-MAPK signaling axis regulates CD133(+) cancer stem
cell-mediated melanoma growth and angiogenesis. J Invest Dermatol.
136:2462–2474. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Monzani E, Facchetti F, Galmozzi E,
Corsini E, Benetti A, Cavazzin C, Gritti A, Piccinini A, Porro D,
Santinami M, et al: Melanoma contains CD133 and ABCG2 positive
cells with enhanced tumourigenic potential. Eur J Cancer.
43:935–946. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rappa G, Fodstad O and Lorico A: The stem
cell-associated antigen CD133 (Prominin-1) is a molecular
therapeutic target for metastatic melanoma. Stem Cells.
26:3008–3017. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Houben R, Wischhusen J, Menaa F, Synwoldt
P, Schrama D, Brocker EB and Becker JC: Melanoma stem cells:
Targets for successful therapy? J Dtsch Dermatol Ges. 6:541–546.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lai CY, Schwartz BE and Hsu MY:
CD133+ melanoma subpopulations contribute to
perivascular niche morphogenesis and tumorigenicity through
vasculogenic mimicry. Cancer Res. 72:5111–5118. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zimmerer RM, Matthiesen P, Kreher F,
Kampmann A, Spalthoff S, Jehn P, Bittermann G, Gellrich NC and
Tavassol F: Putative CD133+ melanoma cancer stem cells
induce initial angiogenesis in vivo. Microvasc Res. 104:46–54.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sharma BK, Manglik V, O'Connell M,
Weeraratna A, McCarron EC, Broussard JN, Divito KA,
Simbulan-Rosenthal CM, Rosenthal DS and Zapas JL: Clonal dominance
of CD133+ subset population as risk factor in tumor
progression and disease recurrence of human cutaneous melanoma. Int
J Oncol. 41:1570–1576. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
El-Khattouti A, Selimovic D, Haikel Y,
Megahed M, Gomez CR and Hassan M: Identification and analysis of
CD133(+) melanoma stem-like cells conferring resistance to taxol:
An insight into the mechanisms of their resistance and response.
Cancer Lett. 343:123–133. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koshio J, Kagamu H, Nozaki K, Saida Y,
Tanaka T, Shoji S, Igarashi N, Miura S, Okajima M, Watanabe S, et
al: DEAD/H (Asp-Glu-Ala-Asp/His) box polypeptide 3, X-linked is an
immunogenic target of cancer stem cells. Cancer Immunol Immunother.
62:1619–1628. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gedye C, Quirk J, Browning J, Svobodova S,
John T, Sluka P, Dunbar PR, Corbeil D, Cebon J and Davis ID:
Cancer/testis antigens can be immunological targets in clonogenic
CD133+ melanoma cells. Cancer Immunol Immunother.
58:1635–1646. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lang D, Mascarenhas JB and Shea CR:
Melanocytes, melanocyte stem cells, and melanoma stem cells. Clin
Dermatol. 31:166–178. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schatton T, Murphy GF, Frank NY, Yamaura
K, Waaga-Gasser AM, Gasser M, Zhan Q, Jordan S, Duncan LM,
Weishaupt C, et al: Identification of cells initiating human
melanomas. Nature. 451:345–349. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Begicevic RR and Falasca M: ABC
transporters in cancer stem cells: Beyond chemoresistance. Int J
Mol Sci. 18:23622017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Frank NY, Margaryan A, Huang Y, Schatton
T, Waaga-Gasser AM, Gasser M, Sayegh MH, Sadee W and Frank MH:
ABCB5-mediated doxorubicin transport and chemoresistance in human
malignant melanoma. Cancer Res. 65:4320–4333. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Roesch A, Fukunaga-Kalabis M, Schmidt EC,
Zabierowski SE, Brafford PA, Vultur A, Basu D, Gimotty P, Vogt T
and Herlyn M: A temporarily distinct subpopulation of slow-cycling
melanoma cells is required for continuous tumor growth. Cell.
141:583–594. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang S, Tang L, Lin J, Shen Z, Yao Y, Wang
W, Tao S, Gu C, Ma J, Xie Y and Liu Y: ABCB5 promotes melanoma
metastasis through enhancing NF-κB p65 protein stability. Biochem
Biophys Res Commun. 492:18–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xiao J, Egger ME, McMasters KM and Hao H:
Differential expression of ABCB5 in BRAF inhibitor-resistant
melanoma cell lines. BMC Cancer. 18:6752018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
de Waard NE, Kolovou PE, McGuire SP, Cao
J, Frank NY, Frank MH, Jager MJ and Ksander BR: Expression of
multidrug resistance transporter ABCB5 in a murine model of human
conjunctival melanoma. Ocul Oncol Pathol. 1:182–189. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vasquez-Moctezuma I, Meraz-Rios MA,
Villanueva-Lopez CG, Magana M, Martinez-Macias R, Sanchez-Gonzalez
DJ, García-Sierra F and Herrera-González NE: ATP-binding cassette
transporter ABCB5 gene is expressed with variability in malignant
melanoma. Actas Dermosifiliogr. 101:341–348. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ma J and Frank MH: Isolation of
circulating melanoma cells. Methods Mol Biol. Sep 29–2015.(Epub
ahead of print). doi: https://doi.org/10.1007/7651_2015_300.
View Article : Google Scholar
|
|
46
|
Frank NY, Schatton T, Kim S, Zhan Q,
Wilson BJ, Ma J, Saab KR, Osherov V, Widlund HR, Gasser M, et al:
VEGFR-1 expressed by malignant melanoma-initiating cells is
required for tumor growth. Cancer Res. 71:1474–1485. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Schatton T, Schutte U, Frank NY, Zhan Q,
Hoerning A, Robles SC, Zhou J, Hodi FS, Spagnoli GC, Murphy GF and
Frank MH: Modulation of T-cell activation by malignant melanoma
initiating cells. Cancer Res. 70:697–708. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Eggermont AM and Robert C: New drugs in
melanoma: It's a whole new world. Eur J Cancer. 47:2150–2157. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Boiko AD, Razorenova OV, van de Rijn M,
Swetter SM, Johnson DL, Ly DP, Butler PD, Yang GP, Joshua B, Kaplan
MJ, et al: Human melanoma-initiating cells express neural crest
nerve growth factor receptor CD271. Nature. 466:133–137. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chesa PG, Rettig WJ, Thomson TM, Old LJ
and Melamed MR: Immunohistochemical analysis of nerve growth factor
receptor expression in normal and malignant human tissues. J
Histochem Cytochem. 36:383–389. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pietra G, Manzini C, Vitale M, Balsamo M,
Ognio E, Boitano M, Queirolo P, Moretta L and Mingari MC: Natural
killer cells kill human melanoma cells with characteristics of
cancer stem cells. Int Immunol. 21:793–801. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Truzzi F, Marconi A, Lotti R, Dallaglio K,
French LE, Hempstead BL and Pincelli C: Neurotrophins and their
receptors stimulate melanoma cell proliferation and migration. J
Invest Dermatol. 128:2031–2040. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nielsen PS, Riber-Hansen R and Steiniche
T: Immunohis tochemical CD271 expression correlates with melanoma
progress in a case-control study. Pathology. 50:402–410. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Guo R, Fierro-Fine A, Goddard L, Russell
M, Chen J, Liu CZ, Fung KM and Hassell LA: Increased expression of
melanoma stem cell marker CD271 in metastatic melanoma to the
brain. Int J Clin Exp Pathol. 7:8947–8951. 2014.PubMed/NCBI
|
|
55
|
Denkins Y, Reiland J, Roy M,
Sinnappah-Kang ND, Galjour J, Murry BP, Blust J, Aucoin R and
Marchetti D: Brain metastases in melanoma: Roles of neurotrophins.
Neuro Oncol. 6:154–165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Shaffer SM, Dunagin MC, Torborg SR, Torre
EA, Emert B, Krepler C, Beqiri M, Sproesser K, Brafford PA, Xiao M,
et al: Rare cell variability and drug-induced reprogramming as a
mode of cancer drug resistance. Nature. 546:431–435. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Holzel M and Tuting T:
Inflammation-induced plasticity in melanoma therapy and metastasis.
Trends Immunol. 37:364–374. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Restivo G, Diener J, Cheng PF, Kiowski G,
Bonalli M, Biedermann T, Reichmann E, Levesque MP, Dummer R and
Sommer L: low neurotrophin receptor CD271 regulates phenotype
switching in melanoma. Nat Commun. 8:19882017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Redmer T, Welte Y, Behrens D, Fichtner I,
Przybilla D, Wruck W, Yaspo ML, Lehrach H, Schäfer R and
Regenbrecht CR: The nerve growth factor receptor CD271 is crucial
to maintain tumorigenicity and stem-like properties of melanoma
cells. PLoS One. 9:e925962014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Prasmickaite L, Skrbo N, Hoifodt HK, Suo
Z, Engebraten O, Gullestad HP, Aamdal S, Fodstad Ø and Maelandsmo
GM: Human malignant melanoma harbours a large fraction of highly
clonogenic cells that do not express markers associated with cancer
stem cells. Pigment Cell Melanoma Res. 23:449–451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lehraiki A, Cerezo M, Rouaud F, Abbe P,
Allegra M, Kluza J, Marchetti P, Imbert V, Cheli Y, Bertolotto C,
et al: Increased CD271 expression by the NF-κB pathway promotes
melanoma cell survival and drives acquired resistance to BRAF
inhibitor vemurafenib. Cell Discov. 1:150302015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Schnegg CI, Yang MH, Ghosh SK and Hsu MY:
Induction of vasculogenic mimicry overrides VEGF-A silencing and
enriches stem-like cancer cells in melanoma. Cancer Res.
75:1682–1690. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Valyi-Nagy K, Kormos B, Ali M, Shukla D
and Valyi-Nagy T: Stem cell marker CD271 is expressed by
vasculogenic mimicry-forming uveal melanoma cells in
three-dimensional cultures. Mol Vis. 18:588–592. 2012.PubMed/NCBI
|
|
64
|
Gray ES, Reid AL, Bowyer S, Calapre L,
Siew K, Pearce R, Cowell L, Frank MH, Millward M and Ziman M:
Circulating melanoma cell subpopulations: Their heterogeneity and
differential responses to treatment. J Invest Dermatol.
135:2040–2048. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Civenni G, Walter A, Kobert N,
Mihic-Probst D, Zipser M, Belloni B, Seifert B, Moch H, Dummer R,
van den Broek M and Sommer L: Human CD271-positive melanoma stem
cells associated with metastasis establish tumor heterogeneity and
long-term growth. Cancer Res. 71:3098–3109. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Furuta J, Inozume T, Harada K and Shimada
S: CD271 on melanoma cell is an IFN-γ-inducible immunosuppressive
factor that mediates downregulation of melanoma antigens. J Invest
Dermatol. 134:1369–1377. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Roesch A: Melanoma stem cells. J Dtsch
Dermatol Ges. 13:118–124. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Clarke MF and Fuller M: Stem cells and
cancer: Two faces of eve. Cell. 124:1111–1115. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Held MA, Curley DP, Dankort D, McMahon M,
Muthusamy V and Bosenberg MW: Characterization of melanoma cells
capable of propagating tumors from a single cell. Cancer Res.
70:388–397. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Frank NY, Schatton T and Frank MH: The
therapeutic promise of the cancer stem cell concept. J Clin Invest.
120:41–50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Boonyaratanakornkit JB, Yue L, Strachan
LR, Scalapino KJ, LeBoit PE, Lu Y, Leong SP, Smith JE and Ghadially
R: Selection of tumorigenic melanoma cells using ALDH. J Invest
Dermatol. 130:2799–2808. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Pinc A, Somasundaram R, Wagner C, Hormann
M, Karanikas G, Jalili A, Bauer W, Brunner P,
Grabmeier-Pfistershammer K, Gschaider M, et al: Targeting CD20 in
melanoma patients at high risk of disease recurrence. Mol Ther.
20:1056–1062. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Akbulut H, Babahan C, Abgarmi SA, Ocal M
and Besler M: Recent advances in cancer stem cell targeted therapy.
Crit Rev Oncog. 24:1–20. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Yaiza JM, Gloria RA, Maria Belen GO, Elena
LR, Gema J, Juan Antonio M, María Ángel GC and Houria B: Melanoma
cancer stem-like cells: Optimization method for culture, enrichment
and maintenance. Tissue Cell. 60:48–59. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Schmidt P, Kopecky C, Hombach A, Zigrino
P, Mauch C and Abken H: Eradication of melanomas by targeted
elimination of a minor subset of tumor cells. Proc Natl Acad Sci
USA. 108:2474–2479. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Murphy GF, Wilson BJ, Girouard SD, Frank
NY and Frank MH: Stem cells and targeted approaches to melanoma
cure. Mol Aspects Med. 39:33–49. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Schlaak M, Schmidt P, Bangard C, Kurschat
P, Mauch C and Abken H: Regression of metastatic melanoma in a
patient by antibody targeting of cancer stem cells. Oncotarget.
3:22–30. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, et al: ALDH1 is a marker of normal and malignant human
mammary stem cells and a predictor of poor clinical outcome. Cell
Stem Cell. 1:555–567. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Huang EH, Hynes MJ, Zhang T, Ginestier C,
Dontu G, Appelman H, Fields JZ, Wicha MS and Boman BM: Aldehyde
dehydrogenase 1 is a marker for normal and malignant human colonic
stem cells (SC) and tracks SC overpopulation during colon
tumorigenesis. Cancer Res. 69:3382–3389. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Jiang F, Qiu Q, Khanna A, Todd NW, Deepak
J, Xing L, Wang H, Liu Z, Su Y, Stass SA and Katz RL: Aldehyde
dehydrogenase 1 is a tumor stem cell-associated marker in lung
cancer. Mol Cancer Res. 7:330–338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Carpentino JE, Hynes MJ, Appelman HD,
Zheng T, Steindler DA, Scott EW and Huang EH: Aldehyde
dehydrogenase-expressing colon stem cells contribute to
tumorigenesis in the transition from colitis to cancer. Cancer Res.
69:8208–8215. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Roudi R, Korourian A, Shariftabrizi A and
Madjd Z: Differential expression of cancer stem cell markers ALDH1
and CD133 in various lung cancer subtypes. Cancer Invest.
33:294–302. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Yoshida A, Hsu LC and Dave V: Retinal
oxidation activity and biological role of human cytosolic aldehyde
dehydrogenase. Enzyme. 46:239–244. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Singh S, Brocker C, Koppaka V, Chen Y,
Jackson BC, Matsumoto A, Thompson DC and Vasiliou V: Aldehyde
dehydrogenases in cellular responses to oxidative/electrophilic
stress. Free Radic Biol Med. 56:89–101. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Le Moguen K, Lincet H, Deslandes E,
Hubert-Roux M, Lange C, Poulain L, Gauduchon P and Baudin B:
Comparative proteomic analysis of cisplatin sensitive IGROV1
ovarian carcinoma cell line and its resistant counterpart
IGROV1-R10. Proteomics. 6:5183–5192. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Moreb JS, Gabr A, Vartikar GR, Gowda S,
Zucali JR and Mohuczy D: Retinoic acid down-regulates aldehyde
dehydrogenase and increases cytotoxicity of
4-hydroperoxycyclophosphamide and acetaldehyde. J Pharmacol Exp
Ther. 312:339–345. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Santini R, Vinci MC, Pandolfi S,
Penachioni JY, Montagnani V, Olivito B, Gattai R, Pimpinelli N,
Gerlini G, Borgognoni L and Stecca B: Hedgehog-GLI signaling drives
self-renewal and tumorigenicity of human melanoma-initiating cells.
Stem Cells. 30:1808–1818. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Luo Y, Dallaglio K, Chen Y, Robinson WA,
Robinson SE, McCarter MD, Wang J, Gonzalez R, Thompson DC, Norris
DA, et al: ALDH1A isozymes are markers of human melanoma stem cells
and potential therapeutic targets. Stem Cells. 30:2100–2113. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Contador-Troca M, Alvarez-Barrientos A,
Merino JM, Morales-Hernandez A, Rodriguez MI, Rey-Barroso J,
Barrasa E, Cerezo-Guisado MI, Catalina-Fernández I,
Sáenz-Santamaría J, et al: Dioxin receptor regulates aldehyde
dehydrogenase to block melanoma tumorigenesis and metastasis. Mol
Cancer. 14:1482015. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yue L, Huang ZM, Fong S, Leong S, Jakowatz
JG, Charruyer-Reinwald A, Wei M and Ghadially R: Targeting ALDH1 to
decrease tumorigenicity, growth and metastasis of human melanoma.
Melanoma Res. 25:138–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Willis BC, Johnson G, Wang J and Cohen C:
SOX10: A useful marker for identifying metastatic melanoma in
sentinel lymph nodes. Appl Immunohistochem Mol Morphol. 23:109–112.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Paratore C, Goerich DE, Suter U, Wegner M
and Sommer L: Survival and glial fate acquisition of neural crest
cells are regulated by an interplay between the transcription
factor Sox10 and extrinsic combinatorial signaling. Development.
128:3949–3961. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Shakhova O, Zingg D, Schaefer SM, Hari L,
Civenni G, Blunschi J, Claudinot S, Okoniewski M, Beermann F,
Mihic-Probst D, et al: Sox10 promotes the formation and maintenance
of giant congenital naevi and melanoma. Nat Cell Biol. 14:882–890.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Taghizadeh R, Noh M, Huh YH, Ciusani E,
Sigalotti L, Maio M, Arosio B, Nicotra MR, Natali P, Sherley JL and
La Porta CA: CXCR6, a newly defined biomarker of tissue-specific
stem cell asymmetric self-renewal, identifies more aggressive human
melanoma cancer stem cells. PLoS One. 5:e151832010. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Zhao F, Zhang R, Wang J, Wu D, Pan M, Li
M, Guo M and Dou J: Effective tumor immunity to melanoma mediated
by B16F10 cancer stem cell vaccine. Int Immunopharmacol.
52:238–244. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Dou J, He X, Liu Y, Wang Y, Zhao F, Wang
X, Chen D, Shi F and Wang J: Effect of downregulation of ZEB1 on
vimentin expression, tumour migration and tumourigenicity of
melanoma B16F10 cells and CSCs. Cell Biol Int. 38:452–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Le Coz V, Zhu C, Devocelle A, Vazquez A,
Boucheix C, Azzi S, Gallerne C, Eid P, Lecourt S and Giron-Michel
J: IGF-1 contributes to the expansion of melanoma-initiating cells
through an epithelial-mesenchymal transition process. Oncotarget.
7:82511–82527. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
RS K: Tumor angiogenesis. N Engl J Med.
358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Rafii S, Lyden D, Benezra R, Hattori K and
Heissig B: Vascular and haematopoietic stem cells: novel targets
for anti-angiogenesis therapy? Nat Rev Cancer. 2:826–835. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Huang WH, Chang MC, Tsai KS, Hung MC, Chen
HL and Hung SC: Mesenchymal stem cells promote growth and
angiogenesis of tumors in mice. Oncogene. 32:4343–4354. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Jeon ES, Lee IH, Heo SC, Shin SH, Choi YJ,
Park JH, Park DY and Kim JH: Mesenchymal stem cells stimulate
angiogenesis in a murine xenograft model of A549 human
adenocarcinoma through an LPA1 receptor-dependent mechanism.
Biochim Biophys Acta. 1801:1205–1213. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Otsu K, Das S, Houser SD, Quadri SK,
Bhattacharya S and Bhattacharya J: Concentration-dependent
inhibition of angiogenesis by mesenchymal stem cells. Blood.
113:4197–4205. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Sun B, Zhang S, Ni C, Zhang D, Liu Y,
Zhang W, Zhao X, Zhao C and Shi M: Correlation between melanoma
angiogenesis and the mesenchymal stem cells and endothelial
progenitor cells derived from bone marrow. Stem Cells Dev.
14:292–298. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Bao S, Wu Q, Sathornsumetee S, Hao Y, Li
Z, Hjelmeland AB, Shi Q, McLendon RE, Bigner DD and Rich JN: Stem
cell-like glioma cells promote tumor angiogenesis through vascular
endothelial growth factor. Cancer Res. 66:7843–7848. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Bussolati B, Bruno S, Grange C, Ferrando U
and Camussi G: Identification of a tumor-initiating stem cell
population in human renal carcinomas. FASEB J. 22:3696–3705. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Hendrix MJ, Seftor EA, Hess AR and Seftor
RE: Vasculogenic mimicry and tumour-cell plasticity: Lessons from
melanoma. Nat Rev Cancer. 3:411–421. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Seftor RE, Hess AR, Seftor EA, Kirschmann
DA, Hardy KM, Margaryan NV and Hendrix MJ: Tumor cell vasculogenic
mimicry: From controversy to therapeutic promise. Am J Pathol.
181:1115–1125. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Girouard SD and Murphy GF: Melanoma stem
cells: Not rare, but well done. Lab Invest. 91:647–664. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Vartanian A, Stepanova E, Grigorieva I,
Solomko E, Baryshnikov A and Lichinitser M: VEGFR1 and PKCα
signaling control melanoma vasculogenic mimicry in a VEGFR2
kinase-independent manner. Melanoma Res. 21:91–98. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chao OS, Chang TC, Di Bella MA, Alessandro
R, Anzanello F, Rappa G, Goodman OB and Lorico A: The HDAC6
inhibitor tubacin induces release of CD133(+) extracellular
vesicles from cancer cells. J Cell Biochem. 118:4414–4424. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Alamodi AA, Eshaq AM, Hassan SY, Al Hmada
Y, El Jamal SM, Fothan AM, Arain OM, Hassan SL, Haikel Y, Megahed M
and Hassan M: Cancer stem cell as therapeutic target for melanoma
treatment. Histol Histopathol. 31:1291–1301. 2016.PubMed/NCBI
|
|
113
|
Luo Y, Ellis LZ, Dallaglio K, Takeda M,
Robinson WA, Robinson SE, Liu W, Lewis KD, McCarter MD, Gonzalez R,
et al: Side population cells from human melanoma tumors reveal
diverse mechanisms for chemoresistance. J Invest Dermatol.
132:2440–2450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Yu B, Wang Y, Yu X, Zhang H, Zhu J, Wang
C, Chen F, Liu C, Wang J and Zhu H: Cuprous oxide
nanoparticle-inhibited melanoma progress by targeting melanoma stem
cells. Int J Nanomedicine. 12:2553–2567. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Shidal C, Al-Rayyan N, Yaddanapudi K and
Davis KR: Lunasin is a novel therapeutic agent for targeting
melanoma cancer stem cells. Oncotarget. 7:84128–84141. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Kaushik G, Venugopal A, Ramamoorthy P,
Standing D, Subramaniam D, Umar S, Jensen RA, Anant S and Mammen
JM: Honokiol inhibits melanoma stem cells by targeting notch
signaling. Mol Carcinog. 54:1710–1721. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Bhattacharyya S, Mitra D, Ray S, Biswas N,
Banerjee S, Majumder B, Mustafi SM and Murmu N: Reversing effect of
Lupeol on vasculogenic mimicry in murine melanoma progression.
Microvasc Res. 121:52–62. 2019. View Article : Google Scholar : PubMed/NCBI
|