Introduction
Most pancreatic cancer-associated deaths are
attributable to pancreatic adenocarcinoma (PAAD) globally (1). PAAD is a common cause of
cancer-associated mortality and only <4% of patients can live
longer than 5 years following diagnosis (2,3).
Furthermore, the incidence of PAAD is continuously increasing, and
it is estimated that PAAD will become the second leading cause of
cancer-associated mortality in the United States by 2030 (4). Pancreaticoduodenectomy is currently the
only potentially curative therapeutic approach for PAAD (5,6). Most
types of cancer can usually be cured at local stages by surgical
resection (5,6). However, resections of PAAD tumor
usually fail to achieve long-term survival in most cases (5,6). It is
therefore crucial to develop novel therapeutic approaches for
PAAD.
Genetic factors play pivotal roles in the
development and progression of PAAD (7). As a cell-surface glycoprotein, CD44
serves a crucial role in cell-cell interactions (8). CD44 is a multifunctional cellular
factor that participates in diverse biological processes, such as
recirculation, hematopoiesis, lymphocyte activation and tumor
metastasis (9). In PAAD, CD44
interacts with multiple oncogenic or tumor suppressive regulators
to promote cancer progression (10).
In addition, certain tumor suppressive microRNAs (miRNAs),
including miR-34a, targets CD44 to suppress cancer metastasis
(11). It has been reported that
miRNAs can interact with other non-coding RNA molecules, such as
long non-coding RNA (lncRNA; >200 nucleotides in length), to
participate in cancer biology (12).
Imatinib-upregulated lncRNA (IUR) is a novel lncRNA that has been
reported as a tumor suppressor in leukemia (13). Preliminary microarray data from our
laboratory revealed the downregulation of IUR in PAAD tissues and
its positive correlation with miR-34a (data not shown). IUR may
therefore have crosstalk with both miR-34a and CD44 in PAAD. Thus,
the present study aimed to investigate the interactions among IUR,
miR-34a and CD44 in PAAD, and to determine their effects on PAAD
cell invasion and migration.
Materials and methods
Patient information
The present study included 58 patients with PAAD (28
men and 30 women; age range, 23–66 years; mean age, 45±16.1 years)
selected from 122 patients with PAAD who were admitted at The
Second Affiliated Hospital of Xi'an Jiaotong University between
January 2012 and January 2014. All patients were followed up from
the date of admission until January 2019 to record their survival
conditions. Follow-up was performed every month through outpatient
visit and/or telephone calls. All PAAD cases were confirmed by
histopathological analysis. The inclusion criteria were as follows:
i) Patients newly diagnosed with PAAD; ii) patients with no other
clinical disorders; and iii) patients who received no treatment for
any disease six months prior to admission. The exclusion criteria
were as follows: i) Patients with recurrent PAAD; ii) patients for
whom therapies for PAAD were initiated; ii) patients with other
severe clinical disorders, and iii) stage IV patients. According to
the American Joint Committee on Cancer staging systems (14), there were 20, 23 and 15 cases at
stage I, II and III, respectively. This study was approved by The
Ethics Committee of The Second Affiliated Hospital of Xi'an
Jiaotong University. All patients were informed with the details of
this study and signed written informed consent.
PAAD tissues and cells
The human PAAD cell lines Capan-2 and HPAC (American
Type Culture Collection) were used in this study. Cells were
cultured in McCoy's 5a Medium (Thermo Fisher Scientific, Inc.)
containing 10% FBS (Thermo Fisher Scientific, Inc.) and placed at
37°C in a humidified incubator containing 5% CO2. Biopsy
was performed on all 58 patients with PAAD to collect PAAD lesion
tissue and adjacent non-cancer tissue samples within 3 cm around
tumors. All tissue samples were stored in liquid nitrogen before
use. Histopathological examination was performed to test all tissue
specimens. Non-cancer tissues contained <1% cancer cells and
PAAD tissues contained >95% cancer cells.
Transient transfection
The pcDNA3.1 vector was used as backbone to
construct IUR and CD44 expression vectors by Guangzhou RiboBio Co.,
Ltd. Negative control (NC) miRNA (5′-UGCGACGUUGGACGUGACGAAU-3′) and
miR-34a mimic (5′-UGGCAGUGUCUUAGCUGGUUGU-3′) were synthesized by
Sangon Biotech Co., Ltd. Capan-2 and HPAC cells were harvested at
70–80% confluence and 5×105 cells were transfected with
10 nM plasmids (pcDNA3.1-IUR, pcDNA3.1-CD44 vectors or empty
pcDNA3.1 vector as NC) or 30 nM miR-34a mimic (or negative control
miRNA as NC). All transfections were performed through transient
transfections using Lipofectamine® 2000 (Sangon Biotech
Co., Ltd.) for 6 h at 37°C. Cells were collected at 24 h
post-transfection for subsequent experiments. Untransfected cells
were used as the control (C) group.
Total RNA and miRNA extractions
The weight of tissues ranged from 0.016 to 0.021 g.
Tissue specimens (0.015 g) were ground in liquid nitrogen. Capan-2
cells were harvested and counted, and 1×106 cells were
collected. Total RNA and miRNA were extracted from tissues and
cells using Trizol® (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturers' instructions.
During the precipitation and washing step, 85% ethanol was used to
precipitate miRNA.
Reverse transcription quantitative
(RT-q) PCR
RNA samples were digested with DNase I to remove
genomic DNA. Tetro Reverse Transcriptase (Bioline) was used to
perform reverse transcriptions (1 µg RNA per reaction, 25°C for 10
min, 52°C for 30 min and 85°C for 10 min). qPCR reaction mixtures
were prepared using BlazeTaq™ SYBR-Green qPCR Mix (GeneCopoeia,
Inc.). GAPDH was used as the endogenous control to measure the
expression levels of IUR and CD44.
For miRNA qPCR, MystiCq® microRNA cDNA
Synthesis Mix (Sigma-Aldrich; Merck KGaA) was used to perform all
reverse transcription. The qPCR reaction mixtures were prepared
using miScript SYBR Green PCR Kit (Qiagen China Co., Ltd). U6 was
used as the endogenous control to measure the expression level of
miR-34a. All qPCR reactions were performed three times.
The primer sequences were as follows: IUR, forward
5′-AGCGGTTTCCTCTTGTTTGTG-3′, reverse 5′-CTTTTGGGTGAGAAAACAAGCC-3′;
CD44, forward 5′-ACCTGCCCAATGCCTTTGATGGA-3′, reverse
5′-CAAAGCCAAGGCCAAGAGGGATG-3′; GAPDH, forward
5′-GCTTTCTTTCCTTTCGCGCT-3′, reverse 5′-TTTGCGGTGGAAATGTCCTT-3′; and
miR-34a, forward 5′-TGGCAGTGTCTTAGCTGGT-3′. Universal miRNA reverse
primer and U6 forward primer were provided in the miScript
SYBR-Green PCR Kit. PCR reactions were performed as follows: 95°C
for 1 min, and then 95°C for 10 sec and 61°C for 40 sec.
The relative expression levels were normalized to
endogenous control and were expressed as 2−ΔΔCq
(15).
Western blotting
Capan-2 cells were harvested and counted. Cells
(1×106) were lysed using RIPA solution on ice (Sangon
Biotech Co., Ltd.). All protein samples were denatured in boiled
water for 5 min. Proteins (20 µg) were separated by 12% SDS-PAGE
and transferred onto PVDF membranes. Membranes were blocked using
5% skimmed milked in PBS at 23°C for 1 h and were incubated with
primary antibodies against CD44 (1:1,000; cat. no. ab157107; Abcam)
and GAPDH (1:1,000; cat. no. ab37168; Abcam) at 4°C overnight.
Membranes were then incubated with the goat anti-rabbit
immunoglobulin G-HRP secondary antibody (1:1,000; cat. no. ab6721;
Abcam) at 23°C for 2 h. Signals were developed using RapidStep™ ECL
detection reagent (Sigma-Aldrich; Merck KGaA). The data were
analyzed via densitometry using ImageJ v1.48 software (National
Institutes of Health) and normalized to expression of the internal
control GAPDH.
Cell migration and invasion
assays
Capan-2 and HPAC cells were harvested and counted.
Cells (3×104) were mixed with 1 ml McCoy's 5a Medium
containing 1% FBS to prepare cell suspensions. The Transwell upper
chamber was filled with 0.1 ml serum-free cell suspension, and the
Transwell lower chamber was filled with McCoy's 5a Medium
supplemented with 20% FBS. Prior to invasion assay, membranes were
coated with Corning® Matrigel® matrix
(Corning) at 37°C for 6 h. Transwell chambers were incubated at
37°C for 12 h. Subsequently, the membranes were washed with PBS and
cells were stained with 1% crystal violet (Sigma-Aldrich; Merck
KGaA) at room temperature for 10 min. The stained cells were
counted under a light microscope.
Statistical analysis
GraphPad Prism 6 (GraphPad Software, Inc.) was used
for data analysis. Western blotting, RT-qPCR, cell invasion and
cell migration assays were repeated three times. Data are presented
as the mean ± standard deviation. Differences between two groups
were compared using paired t-tests. Comparisons between three
groups or more were done using one-way ANOVA followed by Tukey's
post hoc test. Correlation analysis was performed by linear
regression. The 58 patients with PAAD were divided into high- and
low-expression groups using the median expression level of IUR in
PAAD tissues as cutoff value (cutoff value, 2.11). KM plotter
(https://kmplot.com/analysis/) was used
for survival analysis, and survival curves were compared using
log-rank test. P<0.05 was considered to indicate a statistically
significant difference.
Results
IUR is downregulated in PAAD tissues
and low expression of IUR is associated with the poor survival of
patients with PAAD
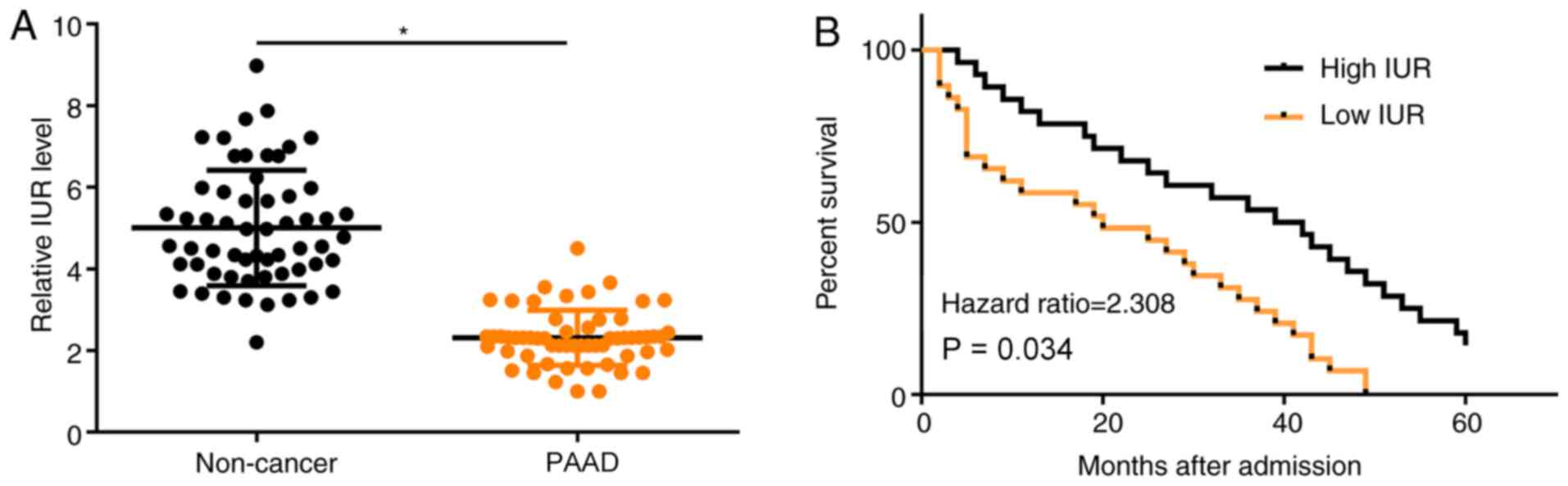
The expression of IUR in PAAD and adjacent non-tumor
tissues was assessed by RT-qPCR. The results demonstrated that IUR
expression was significantly decreased in PAAD tissues compared
with non-cancer tissues (Fig. 1A;
P<0.05). Survival curves were obtained patients with high and
low IUR expression. It was observed that patients in the
low-expression group had a significantly lower overall survival
rate compared with patients with high IUR expression (Fig. 1B; P<0.05).
IUR expression positively correlates
with miR-34a expression but negatively correlated with CD44
expression
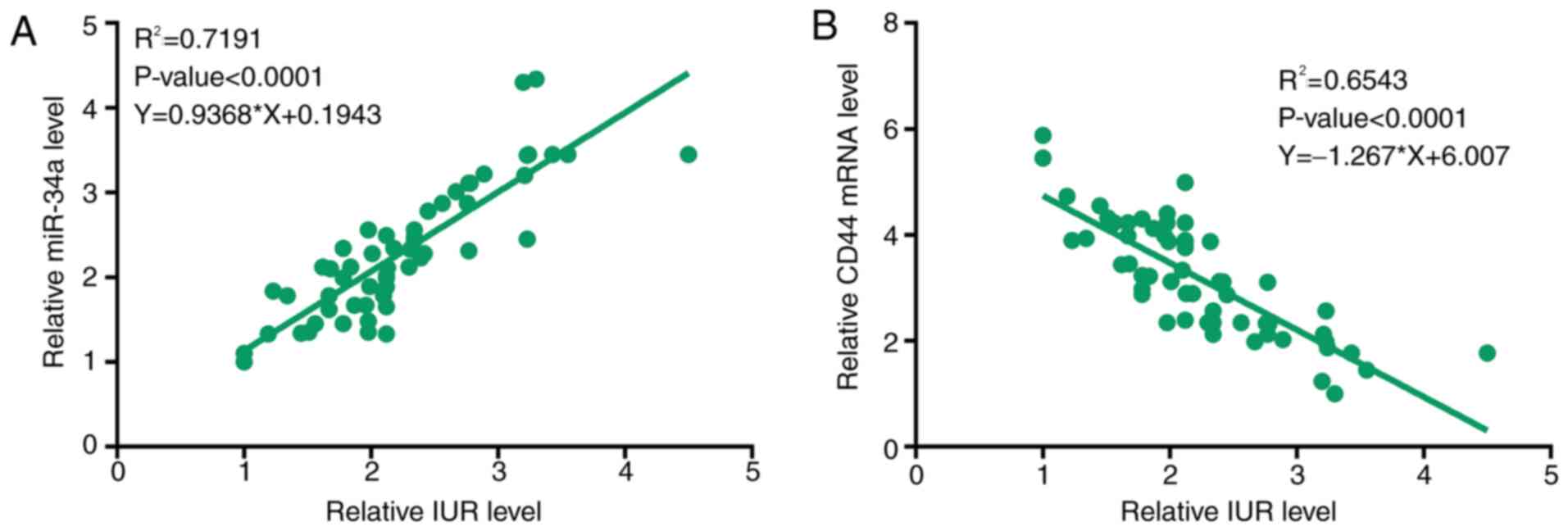
The expression of miR-34a and CD44 in PAAD tissue
samples was evaluated by RT-qPCR. Correlation analysis demonstrated
that the expression of IUR was positively correlated with the
expression of miR-34a (Fig. 2A), but
negatively correlated with the expression of CD44 (Fig. 2B) in PAAD tissues.
IUR downregulates CD44 by upregulating
miR-34a
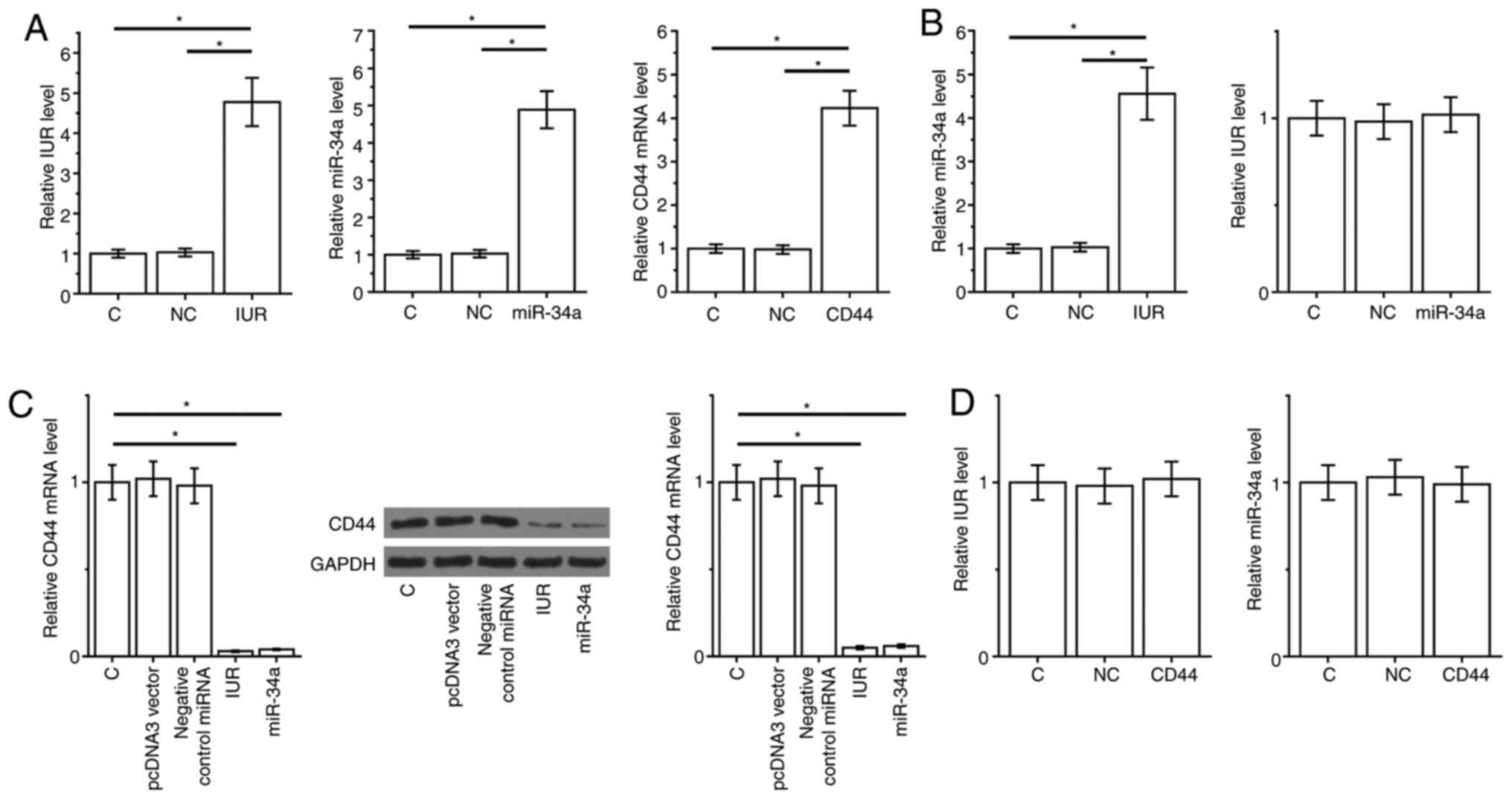
The IUR expression vector, miR-34a mimic and CD44
expression vector were transfected into Capan-2 cells. At 24 h
post-transfection, the expression of IUR, miR-34a and CD44 was
significantly upregulated compared with the C and NC groups
(Fig. 3A; P<0.05). Furthermore,
overexpression of IUR resulted in miR-34a upregulation, while
overexpression of miR-34a did not affect the expression of IUR
(Fig. 3B). In addition, IUR
overexpression or miR-34a led to downregulated expression of CD44
in PAAD cells (Fig. 3C; P<0.05).
However, overexpression of CD44 did not affect the expression of
IUR and miR-34a (Fig. 3D).
IUR inhibits Capan-2 and HPAC cell
invasive and migratory abilities via miR-34a and CD44
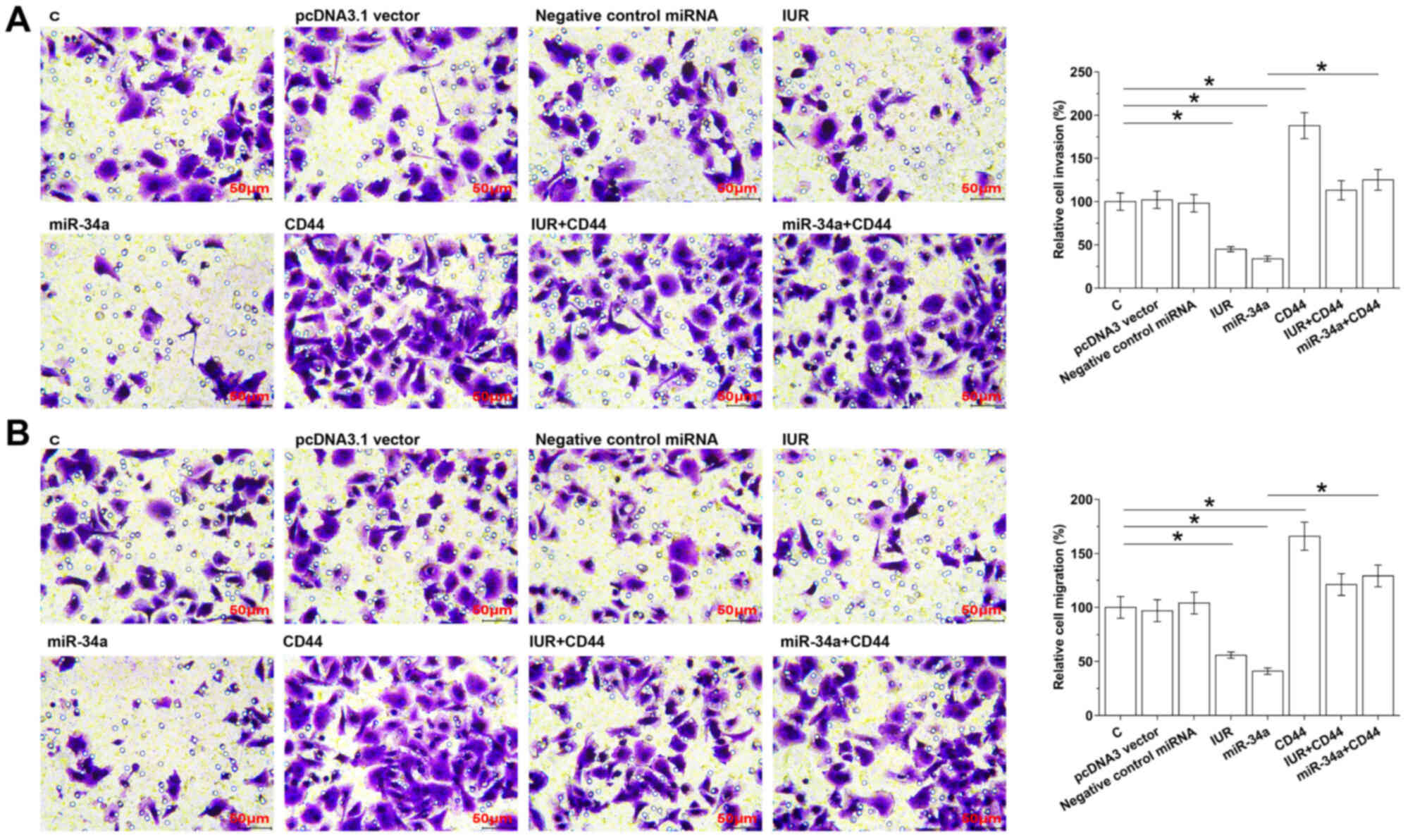
Cell invasion and migration assay results
demonstrated that, compared with the NC and C groups,
overexpression of IUR and miR-34a led to decreased invasive
(Fig. 4A) and migratory (Fig. 4B) abilities of Capan-2 cells
(P<0.05). Overexpression of CD44 played the opposite role and
attenuated the effect of IUR overexpression (P<0.05).
Another PAAD cell line, HPAC, was used to repeat the
Transwell assays to further confirm the role of IUR in regulating
PAAD cell invasion and migration. Similarly, invasion (Fig. S1A) and migration (Fig. S1B) of HPAC cells were inhibited
after the overexpression of IUR and miR-34a (P<0.05).
Overexpression of CD44 played the opposite role and attenuated the
effects of IUR overexpression (Fig.
S1; P<0.05).
Discussion
The present study investigated the associations
among IUR, miR-34a and CD44 in PAAD. The results demonstrated that
IUR may be considered as an oncogenic lncRNA in PAAD and could
inhibit cancer cell invasion and migration by downregulating CD44
through the upregulation of miR-34a, which can directly target CD44
(11).
The function of IUR has only been characterized in
leukemia (13), in which IUR can
regulate the STAT5-CD71 pathway to inhibit tumorigenesis induced by
BCR-ABL1 (13). Our preliminary
microarray data revealed downregulation of IUR in PAAD. In the
present study, IUR was downregulated in PAAD and overexpression of
IUR resulted in decreased invasive and migratory abilities of PAAD
cells. IUR may therefore be considered as a tumor suppressor lncRNA
in PAAD.
The survival of patients with PAAD has been improved
in the last decades, although only <5% of patients live longer
than 5 years (16,17). In the present study, according to the
5-year follow-up, only four patients (6.9%) survived, which is
slightly higher than the average (4%) global level (2,3). This
could be due to the fact that this study only included patients at
clinical stages I–III. Due to the extremely poor survival of these
patients, accurate prognostic assignment is extremely important
(18). The present study
demonstrated that patients with low IUR expression in PAAD tissues
experienced significantly lower overall survival rate. Evaluation
of IUR expression may therefore assist the prognosis of patients
with PAAD.
It has been reported that miR-34a can target CD44 in
prostate cancer (11). In the
present study, expression of CD44 was downregulated following
miR-34a overexpression, both at the mRNA and protein levels.
miR-34a may therefore also target CD44 in PAAD. In addition, this
study reported that IUR could upregulate miR-34a. However, no
potential target of miR-34a on IUR was observed. Both IUR and
miR-34a are known to have crosstalk with the STAT signaling pathway
(13,19). Therefore, the STAT signaling pathway
may mediate the interaction between miR-34a and IUR. This will be
further investigated in the future.
In conclusion, the present study demonstrated that
IUR was upregulated in PAAD tissues compared with non-cancer tissue
samples. In addition, IUR may upregulate miR-34a in PAAD cells to
downregulate CD44, thereby inhibiting the invasive and migratory
abilities of PAAD cancer cells.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by The Science and
Technology Program of Shaanxi Province of China (grant no.
2014K11-03-03-07) and The Natural Science Foundation of Shaanxi
Province of China (grant no. 2016JM8093). The funding bodies had no
role in the design of the study; collection, analysis and
interpretation of data and writing of the manuscript.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
RL and SM wrote the manuscript, performed literature
research and analyzed data. GZ, JL, JWL, XW and BH analyzed data,
performed the experiments and provided statistical analysis. JZ
performed literature research and was responsible for project
management and study design. RL, JZ, SM, GZ, JL JWL, XW and BH
confirm the authenticity of the data in the present manuscript. All
authors read and approved the final version.
Ethics approval and consent to
participate
The Ethics Committee of the Second Affiliated
Hospital of Xi'an Jiaotong University approved this study. All
procedures performed in studies involving human participants were
in accordance with the 1964 Declaration of Helsinki and its later
amendments or comparable ethical standards. Written informed
consent was obtained from all individual participants included in
the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Luberice K, Downs D, Sadowitz B, Ross S
and Rosemurgy A: Has survival improved following resection for
pancreatic adenocarcinoma? Am J Surg. 214:341–346. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig
AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and
deaths to 2030: The unexpected burden of thyroid, liver, and
pancreas cancers in the United States. Cancer Res. 74:2913–2921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nickel F, Haney CM, Kowalewski KF, Probst
P, Limen EF, Kalkum E, Diener MK, Strobel O, Müller-Stich BP and
Hackert T: Laparoscopic versus open pancreaticoduodenectomy: A
systematic review and meta-analysis of randomized controlled
trials. Ann Surg. 271:54–66. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Amundadottir LT: Pancreatic cancer
genetics. Int J Biol Sci. 12:314–325. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aruffo A, Stamenkovic I, Melnick M,
Underhill CB and Seed B: CD44 is the principal cell surface
receptor for hyaluronate. Cell. 61:1303–1313. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Senbanjo LT and Chellaiah MA: CD44: A
multifunctional cell surface adhesion receptor is a regulator of
progression and metastasis of cancer cells. Front Cell Dev Biol.
5:182017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang W, Zhang Y, Kane KT, Collins MA,
Simeone DM, di Magliano MP and Nguyen KT: CD44 regulates pancreatic
cancer invasion through MT1-MMP. Mol Cancer Res. 13:9–15. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu C, Kelnar K, Liu B, Chen X,
Calhoun-Davis T, Li H, Patrawala L, Yan H, Jeter C, Honorio S, et
al: The microRNA miR-34a inhibits prostate cancer stem cells and
metastasis by directly repressing CD44. Nat Med. 17:211–215. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jalali S, Bhartiya D, Lalwani MK,
Sivasubbu S and Scaria V: Systematic transcriptome wide analysis of
lncRNA-miRNA interactions. PLoS One. 8:e538232013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Xu Y, Wang G, Chen X, Liang W and Ni
H: Long non-coding RNA Mirt2 relieves lipopolysaccharide-induced
injury in PC12 cells by suppressing miR-429. J Physiol Biochem.
75:403–413. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Appel BL, Tolat P, Evans DB and Tsai S:
Current staging systems for pancreatic cancer. Cancer J.
18:539–549. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Von Hoff DD, Ervin T, Arena FP, Chiorean
EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et
al: Increased survival in pancreatic cancer with nab-paclitaxel
plus gemcitabine. N Engl J Med. 369:1691–1703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ilic M and Ilic I: Epidemiology of
pancreatic cancer. World J Gastroenterol. 22:9694–9705. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang JJ, Hu ZG, Shi WX, Deng T, He SQ and
Yuan SG: Prognostic significance of neutrophil to lymphocyte ratio
in pancreatic cancer: A meta-analysis. World J Gastroenterol.
21:2807–2815. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li H, Rokavec M and Hermeking H: Soluble
IL6R represents a miR-34a target: Potential implications for the
recently identified IL-6R/STAT3/miR-34a feed-back loop. Oncotarget.
6:14026–14032. 2015. View Article : Google Scholar : PubMed/NCBI
|