Introduction
Ovarian cancer (OC) is the 7th most diagnosed cancer
among women worldwide and the 10th most common in China (1). Additionally, ~90% of all OCs are
epithelial in terms of origin, that is, epithelial OC (EOC)
(2). Since EOC is a chemo-responsive
tumour, which responds well to platinum, including cisplatin, the
standard therapy for EOC remains surgery combined with
chemotherapy(3,4). Platinum-taxanes doublet is the standard
treatment in EOC (4). However, the
prognosis of patients with EOC remains poor (2,5,6). Firstly, almost 80% of patients develop
tolerance to standard platinum therapy and will recur with a
resistant disease (5). Secondly, the
5-year overall survival rate has remained at only 30–40% worldwide
since 1995 (2). Furthermore, >70%
of patients are diagnosed with advanced-stage disease and would
suffer from multiple metastases, migration and invasion of the
tumour, which eventually lead to death (6).
Although a meta-analysis has provided evidence of
the benefits of combination-platinum over single-agent platinum
(7), platinum combinations
inevitably lead to greater toxicity and similar antitumour
molecular mechanisms, which are more likely to lead to the
development of platinum resistance (8). Therefore, searching for novel chemical
drugs with diverse antitumour molecular mechanisms that could be
combined with platinum is an improved selection approach for EOC
combination chemotherapy (9).
Nigericin is an H+, K+ and
Pb2+ ionophore derived from Streptomyces
hygroscopicus (10,11). It is widely used as an antibiotic
agent in chickens (12). Nigericin
has exhibited potent anticancer activities in several types of
cancer (12,13), as well as in resistant cancer stem
cells (14,15). Nigericin sensitizes cancer cells and
exhibits a synergistic effect with other anticancer drugs, thereby
leading to improved efficacy of these drugs (16). This enhanced efficacy is mostly
associated with nigericin's ability to reduce internal pH in an
acidic tumour microenvironment (10,13). Our
previous study demonstrated that nigericin could inhibit
epithelial-mesenchymal transition (EMT) during cell invasion and
metastasis via the canonical Wnt/β-catenin signalling pathway
(17), and may be a chemotherapy
option combined with platinum for patients with EOC.
However, whether chemotherapy that combines
nigericin and platinum has better therapeutic effects than single
chemical drug therapy and the molecular mechanism of nigericin
triggered by the combination chemotherapy remain unclear.
Therefore, the present study aimed to explore the aforementioned
questions in human EOC cells.
Materials and methods
Cell lines and cell culture
Human SK-OV-3 and A2780 EOC cell lines were used in
the present study. SK-OV-3 cells were obtained from The Cell Bank
of Type Culture Collection of The Chinese Academy of Sciences, and
A2780 cells were purchased from American Type Culture Collection.
SK-OV-3 cells were cultured in McCoy's 5A medium (Thermo Fisher
Scientific, Inc.), while A2780 cells were cultured in RPMI-1640
medium (Thermo Fisher Scientific, Inc.). Both media were
supplemented with 10% FBS (Thermo Fisher Scientific, Inc.), 100
U/ml penicillin and 100 U/ml streptomycin (Sigma-Aldrich; Merck
KGaA). Both cell lines were cultured at 37°C in a humidified
atmosphere with 5% CO2. The cells were routinely grown
until they reached 80% confluence and then subcultured or plated
for experiments.
Drugs and antibodies
Nigericin and cisplatin were obtained from
Sigma-Aldrich; Merck KGaA. Nigericin (25 mM stock) was dissolved in
DMSO. Cisplatin (1 mM stock) was dissolved in normal saline (0.9%
NaCl). Both stock solutions were stored as aliquots at −20°C. DMSO
served as a vehicle control. The following primary antibodies were
used at the dilution recommended by the manufacturer (1:1,000) and
purchased from Cell Signaling Technology, Inc.: Rabbit anti-Axin-2
(cat. no. 2151), rabbit anti-MMP7 (cat. no. 71031), rabbit
anti-vimentin (cat. no. 5741), rabbit anti-tubulin (cat. no. 2148),
rabbit anti-β-catenin (cat. no. 25362), rabbit anti-slug (cat. no.
9585), rabbit anti-E-cadherin (cat. no. 3195) and rabbit anti-small
ubiquitin-like modifier 1 (SUMO1; cat. no. 4930). Mouse anti-GSK3β
(cat. no. sc-377213) was used at a 1:200 dilution, and purchased
from Santa Cruz Biotechnology, Inc. Goat anti-mouse IgG (HRP)
(ZB-2305) and Goad anti-rabbit IgG (HRP) (ZB-2301) were used at a
dilution of 1:2,500, and were purchased from ZSGB-BIO, Inc.
(http://www.zsbio.com/).
Wound healing assay
SK-OV-3 cells were plated into 6-well plates and
allowed to reach confluence at 90%. Subsequently, a 200-µl pipette
tip was used to scratch constant-diameter wounds in the confluent
monolayers. Cells were rinsed twice with PBS and incubated with
serum-free medium for 24 h at 37°C to weaken the interference
effects of cell proliferation. Cells in different wells were
treated with different concentrations of drugs (nigericin or
cisplatin) in the medium (5 and 20 µmol/ml), shown in each figure.
The wounds were imaged at 0 and 24 h after scratching under an
inverted microscope (NIB-100F; Ningbo Yongxin OPTICS CO.,
LTD.).
Transwell assay
Boyden chambers with 8-µm pore filters were used.
SK-OV-3 and A2780 cells treated with DMSO or combined drugs at
different concentrations in McCoy's 5A medium and RMPI-1640 medium,
respectively, without FBS, were plated in the upper chambers, which
were pre-coated with 25% Matrigel in medium at 37°C for 4 h. The
lower chambers were filled with culture medium supplemented with
10% FBS. After incubation for 24 h at 37°C, cells were fixed in 4%
paraformaldehyde for 20 min at room temperature, then non-migrating
cells in the upper chamber were completely removed using a cotton
swab. Cells that migrated to the lower surface of the membrane were
stained with 0.5% crystal violet for 3 min at room temperature.
These cells were using an inverted microscope and quantified by
counting the number of stained nuclei in five random fields using
ImageJ (1.42; National Institutes of Health). Each assay was
repeated twice.
Cell viability assay
The viability of EOC cells treated with nigericin in
combination with cisplatin was analysed using a Cell Counting Kit-8
(CCK-8; cat. no. CK04; Dojindo Molecular Technologies, Inc.)
according to the manufacturer's protocol. Briefly, SK-OV-3 and
A2780 cells were plated into a 96% well plate at a density of 4,000
cells/well in a final volume of 100 µl medium and cultured under
normal conditions for 24 h. Subsequently, nigericin (5 µM) and
cisplatin (5 µM or 20 µM) were added to each well of the plates
using DMSO as a vehicle control. Subsequently, 10 µl CCK-8 reagent
was added to each well post-incubation for 48 h, and the cells were
incubated and shielded from light for 1 h at 37°C. Cell viability
was calculated by measuring the absorbance at 450 nm using a
microplate reader (Bio-Rad Laboratories, Inc.). Growth inhibition
was calculated as a percentage of the vehicle control. Experiments
were performed three times and the data are presented as the mean ±
SD of five wells per treatment.
Colony formation assay
SK-OV-3 and A2780 cells were plated into a 6% well
plate at a density of 1,000 cells/well and grown under normal
conditions for 12 h. Then, cells were treated with nigericin and
cisplatin at 5 or 20 µM, as shown in each figure. Following
incubation under normal conditions for 14 days, the cells were
fixed with 4% paraformaldehyde for 20 min at room temperature, and
then stained with 0.5% crystal violet (Beyotime Institute of
Biotechnology) for 30 min at room temperature. Subsequently, the
number of cell colonies was counted under an inverted
microscope.
Luciferase reporter assay
The TOPflash/FOPflash reporter assay was used for
monitoring the activity of the Wnt/β-catenin signalling pathway in
SK-OV-3 and A2780 cells. The pTOPflash vector (cat. no. D2501) and
pFOPflash vector (cat. no. D2503) were purchased from Beyotime
Biotechnology and constructed based on pGL6-TA, according to the
product specifications. The pTOPflash vector contains a firefly
luciferase gene under the control of two T cell factor (TCF)
response elements with three repeats located upstream of minimal TA
viral promoter. The negative control pFOPflash vector contains a
firefly luciferase gene under the control of two mutant TCF
response elements with three repeats located upstream of minimal TA
viral promoter. SK-OV-3 and A2780 cells (5×104
cells/well) were seeded into 48-well plates and incubated overnight
before transfection. The cells were co-transfected with reporter
plasmid (200 ng pTOPflash or pFOPflash) and control vector (20 ng
pRL-TK, cat. no. D2760, Beyotime Biotechnology) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). At 4 h post-transfection, the culture medium was
changed for complete medium and treated with nigericin and/or
cisplatin at 5 or 20 µM . The SK-OV-3 or A2780 cells were cultured
for 48 h in complete medium. Afterwards, the luciferase activity
was measured using the Dual-Luciferase® Reporter Assay
System (Promega Corporation) according to the manufacturer's
protocol, and normalized for transfection efficiency using the
corresponding Renilla luciferase activity. Reporter activity
was calculated as a ratio of TOPflash to FOPflash. The experiment
was performed in triplicate.
Transient knockdown
Transient gene/protein expression knockdown in
SK-OV-3 cells was established by transfection with 50 nM
non-specific [negative control small interfering (siRNA):
GCAAGCTGACCCTGAAGTT] or specific siRNA molecules targeting slug
(slug-siRNA-1: 5′-CAAACGACTTTGCAACTCC-3′; slug-siRNA-2:
5′-CCTCTTGGCATACTCCTCT-3′), which were purchased from Generay
biotech Co. Ltd. Cells were transfected with siRNAs using
Lipofectamine® RNAiMAX Reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) as recommended by the manufacturer. Cells
were analysed after 48 h at 37°C. The cells were then harvested for
RT-qPCR and western blot analysis.
Lentivirus-mediated
overexpression
Slug-overexpressing SK-OV-3 and A2780 EOC cell lines
were established via lentivirus transfection. A lentiviral vector
encoding slug (LV-slug) and a negative control vector (LV-vector)
were purchased from OBiO Technology (Shanghai) Corp., Ltd., and
carried an enhanced green fluorescent protein reporter gene, EGFP.
The lentiviral vector used for overexpression was named
pSLenti-CMV-EGFP-3×FLAG-PGK-Puro-WPRE according to the
manufacturer's introduction. For slug exogenous overexpression,
lentivirus containing LV-slug or the LV-vector were transfected
into SK-OV-3 and A2780 cells using Polybrene (5.0 µg/ml) from OBiO
Technology according to the manufacturer's protocol. Briefly, cells
were seeded into a 6-well plate (1×105 cells/well) for
adherence. Then, the cells were transfected with lentivirus vectors
(LV-slug or LV-vector) at a multiplicity of infection of 30, in the
presence of 5 µg/ml Polybrene at 37°C. After 72 h of transduction,
medium containing puromycin (0.2 mg/ml) was added to select stably
transduced cells, and stable cell lines were screened for 3 weeks.
Slug upregulation efficiency was assessed by western blotting.
Western blotting
Cells were washed with 1X PBS three times, and the
total cell proteins were extracted using RIPA lysis buffer
(Beyotime Institute of Biotechnology) with freshly added proteinase
inhibitor cocktail and phosphatase inhibitor (Sigma-Aldrich; Merck
KGaA) on ice for 15 min and centrifuged at 12,700 × g for 20 min at
4°C. BCA (Sigma-Aldrich; Merck KGaA) was used to determine the
concentration of total protein of EOC cells. For western blot
analysis, lysates (20 µg/lane) were subjected to 12% SDS-PAGE, and
then the fractionated proteins were transferred to a PVDF membrane
(ISEQ00010/IPVH00010; Millipore). The membrane was blocked in 1X
TBS with 0.5% Tween-20 (TBST) with 5% non-fat milk for 1 h at room
temperature and then incubated with primary antibodies against axin
2, vimentin, tubulin, β-catenin, slug, E-cadherin, SUMO and GSK3β,
overnight at 4°C. Next day, the membrane was washed with 1X TBST
three times and incubated with the appropriate HRP-conjugated
secondary antibodies for 1 h at room temperature. Positive
immunoreactive proteins were detected using the Novex™ ECL
Chemiluminescent Substrate Reagent Kit (Thermo Fisher Scientific,
Inc.) to visualize signals and bands. The tubulin band served as a
reference control.
Immunoprecipitation (IP) assay
In the IP assay, SK-OV-3 cells were cultured in
10-cm dishes and incubated with nigericin or cisplatin (as shown in
figures) for 48 h at 4°C. Then, the cells were lysed with IP-buffer
[Tris-Cl(pH 7.5) 50 mmol/l, NaCl 150 mmol/l, NP-40 1%, EDTA 5
mmol/l, PMSF 1 mmol/l, protease inhibitor (aprotinin 2 µg/ml,
leupeptin 2 µg/ml, and pepstatin 7 µg/ml)] on ice for 30 min, and
centrifuged at 12,700 × g for 10 min at 4°C. Then, the supernatant
was transferred to new tubes immediately, and 50% protein G agarose
was added with ratio of 100 µl for a 1 ml sample. The samples were
shaken on horizontal shaker for 10 min at 4°C, to eliminate
non-specific binding proteins. Next, add 1 µg anti-slug antibody
(cat. no. 9585; Cell Signaling Technology, Inc.) and 50% protein G
agarose (with ratio of 100 µl for a 1 ml sample solution) were
added, and incubated on a horizontal shaker overnight at 4°C,
centrifuged at 3,000 g for 10 min at 4°C, and the supernatants were
discarded. The agarose was then collected and washed twice with IP
buffer, then collected by centrifugation (3,000 × g for 10 min at
4°C), and detected by western blotting using anti-SUMO antibodies
(cat. no. 4930; CST).
Statistical analysis
Statistical analyses were performed using a
statistical software package (SPSS 19.0; IBM Corp.). Data are
presented as the mean ± standard deviation. Statistical
significances were determined by one-way ANOVA with Tukey's post
hoc test for multiple comparisons. Unpaired Student's t-test was
used to determine the statistical significance for two independent
groups (cells treated with combined nigericin and cisplatin vs.
cells treated with cisplatin only). P<0.05 was considered to
indicate a statistically significant difference.
Results
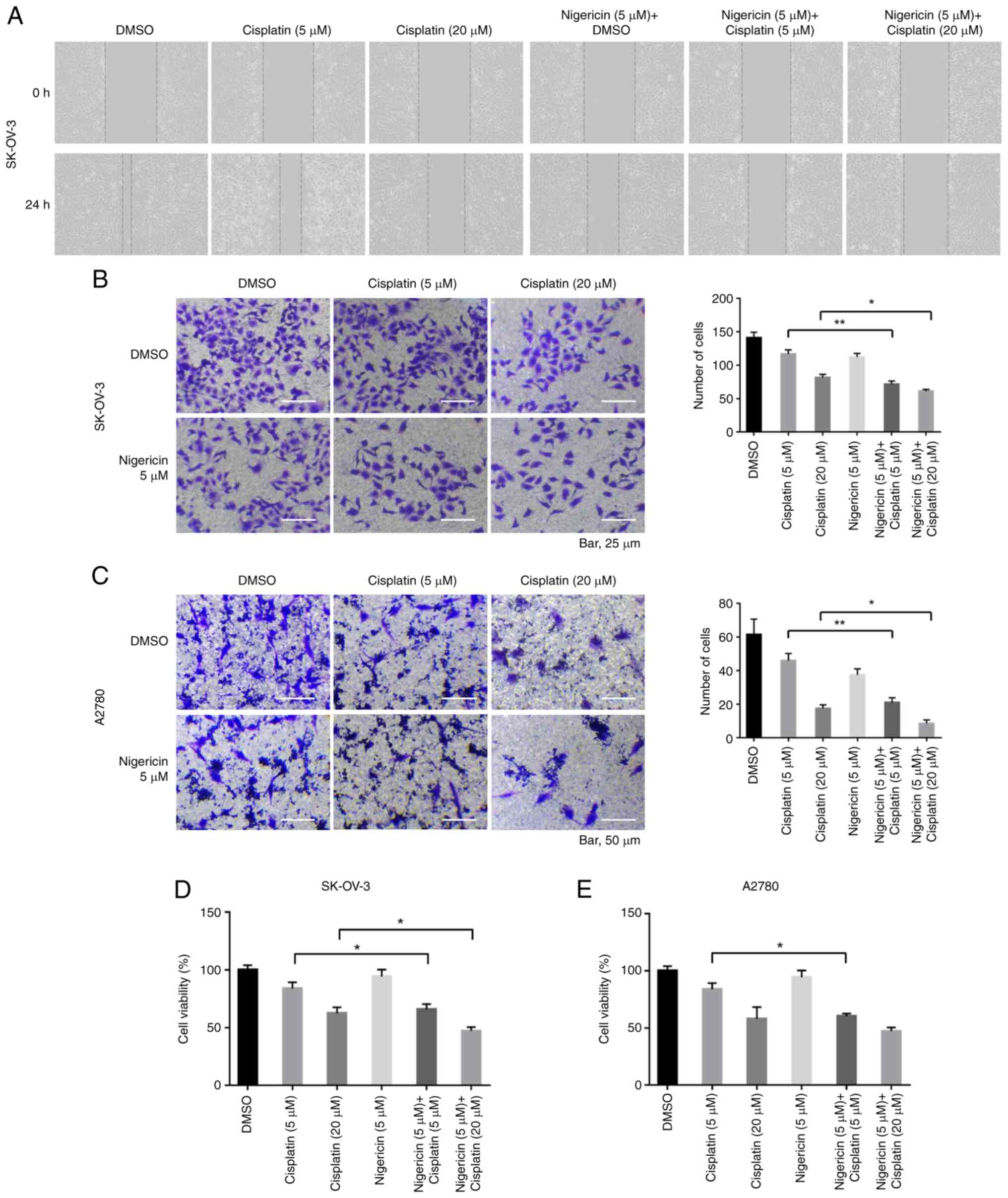
Nigericin combined with cisplatin
enhances the inhibitory effect of cisplatin on the migration of EOC
cells
To explore the effects of nigericin on promoting the
inhibitory effect of cisplatin on the migration of EOC cells,
nigericin combined with cisplatin was used to treat EOC cells, and
wound healing and Transwell assays were performed. The results
revealed that the inhibitory effect on the migration of EOC cells
after the addition of nigericin was more significant compared with
that of only cisplatin (5 µM nigericin and 5 µM cisplatin vs. 5 µM
cisplatin; 5 µM nigericin and 20 µM cisplatin vs. 20 µM cisplatin;
Fig. 1A-C). Moreover, the promoting
effect of nigericin on the inhibition of proliferation caused by
cisplatin was significant compared with cisplatin treatment alone
(Fig. 1D and E). These data
suggested that nigericin might interact with/affect the migration
pathway.
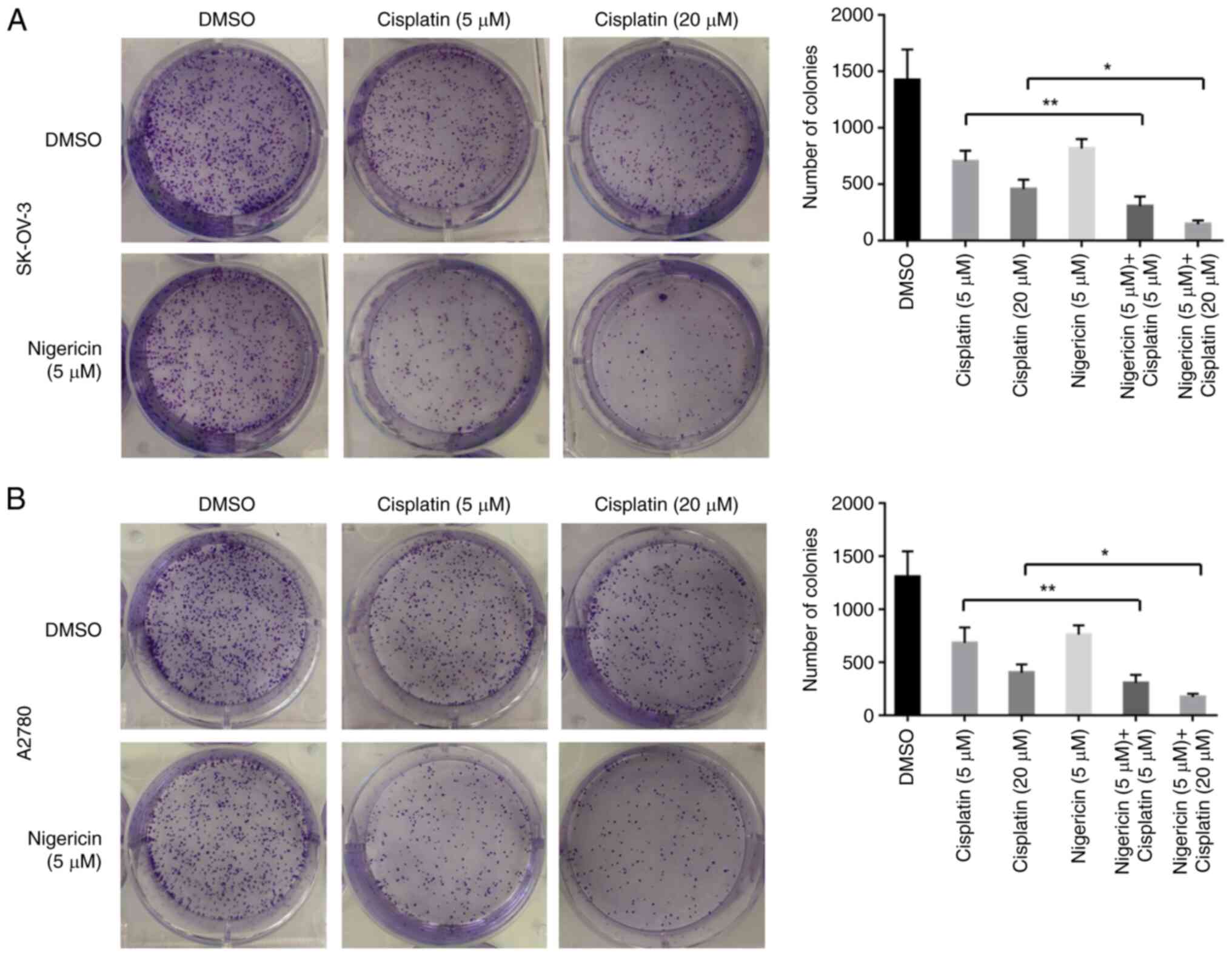
Nigericin combined with cisplatin
enhances the inhibitory effect of cisplatin on the colony formation
of EOC cells
The present study also used a colony formation assay
to examine the long-term effects of nigericin combined with
cisplatin on the proliferation of EOC cells. The results
demonstrated that nigericin and cisplatin could inhibit colony
formation. However, the combination of nigericin and cisplatin
exhibited more significant inhibitory effects than cisplatin only
on colony formation in SK-OV-3 and A2780 cells (Fig. 2). Overall, the combination of
nigericin and cisplatin caused a significant reduction in migration
and colony formation compared with the single cisplatin drug used
in EOC cells. These data suggested that the combination of
nigericin and cisplatin might inhibit the migration and
proliferation of EOC cells effectively.
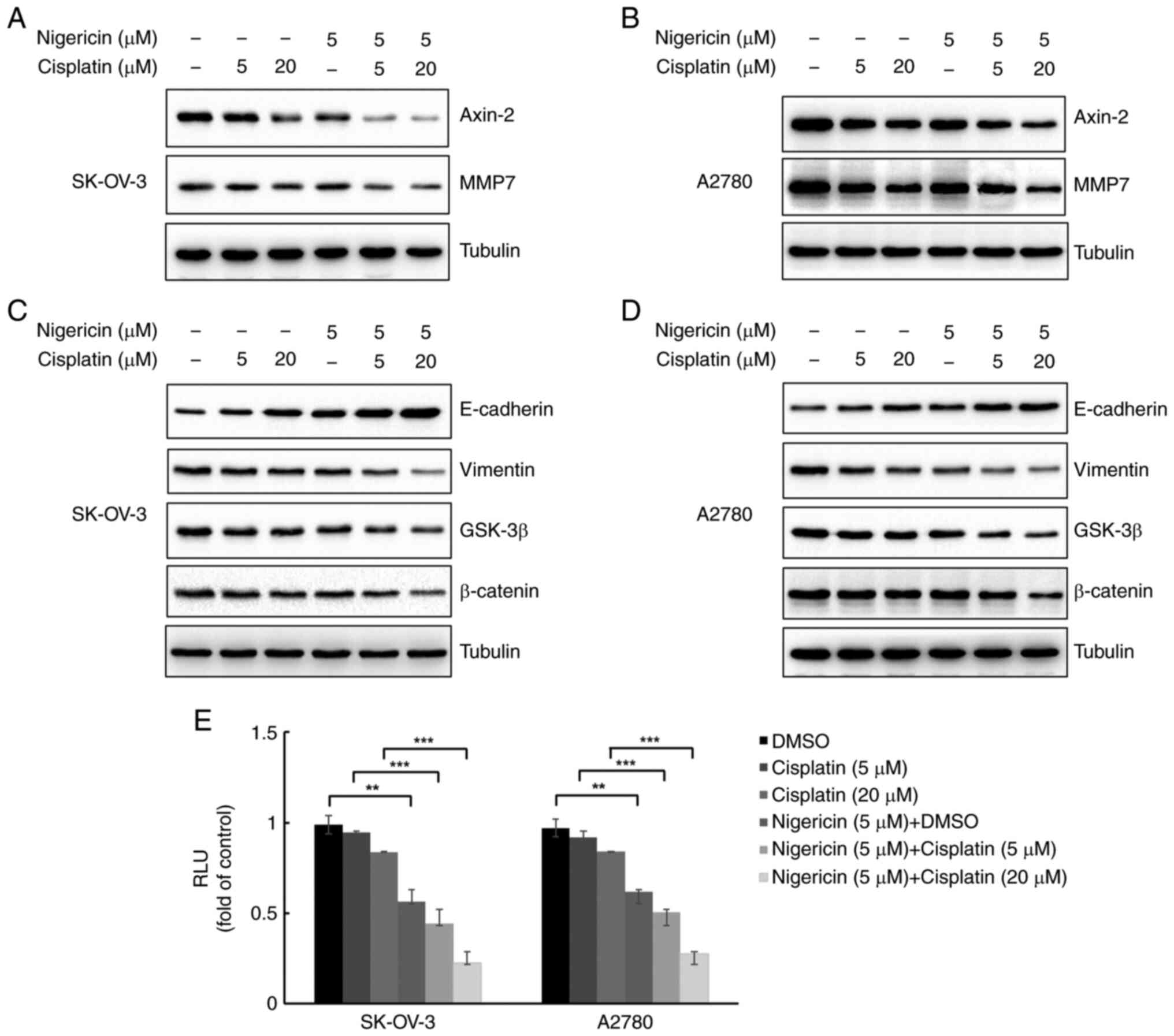
Nigericin combined with cisplatin
regulates MMP expression and inhibits the Wnt/β-catenin signalling
pathway in EOC cells
MMPs are usually overexpressed in EOC cells and
considered to be associated with metastasis (18). To determine whether nigericin
combined cisplatin influenced MMP expression in EOC cells, the
present study examined MMP7 expression in SK-OV-3 and A2780 cells
using western blotting. Furthermore, E-cadherin and vimentin are
markers of epithelial cells and mesenchymal cells, which serve
important roles in the EMT, and EMT is essential for metastasis of
EOC cells. A pervious study has demonstrated that nigericin could
increase the expression levels of E-cadherin and decrease vimentin
expression in EOC cells with a dose-response relationship via the
Wnt/β-catenin signalling pathway (12). The present study examined the
combined effect of nigericin and cisplatin on the Wnt/β-catenin
signalling pathway. The results demonstrated that 5 µM nigericin
enhanced the inhibitory effect of cisplatin (5 and 20 µM) on MMP7
and Axin-2 expression (Fig. 3A and
B). Nigericin (5 µM) also enhanced the inhibitory effect of
cisplatin (5 and 20 µM) on β-catenin and GSK-3β expression, thereby
indicating the involvement of nigericin and cisplatin in the
Wnt/β-catenin signalling pathway by inhibiting its activation
(Fig. 3C and D). Furthermore, when
combined with cisplatin, nigericin could increase the expression
levels of E-cadherin and decrease the expression levels of vimentin
in EOC cells (Fig. 3C and D).
Furthermore, luciferase reporter assays revealed that activation of
the Wnt/β-catenin pathway was markedly reduced by treatment with
combined nigericin (5 µM) and cisplatin (5 and 20 µM) compared with
cisplatin alone (5 and 20 µM) in both SK-OV-3 and A2780 cells
(Fig. 3E).
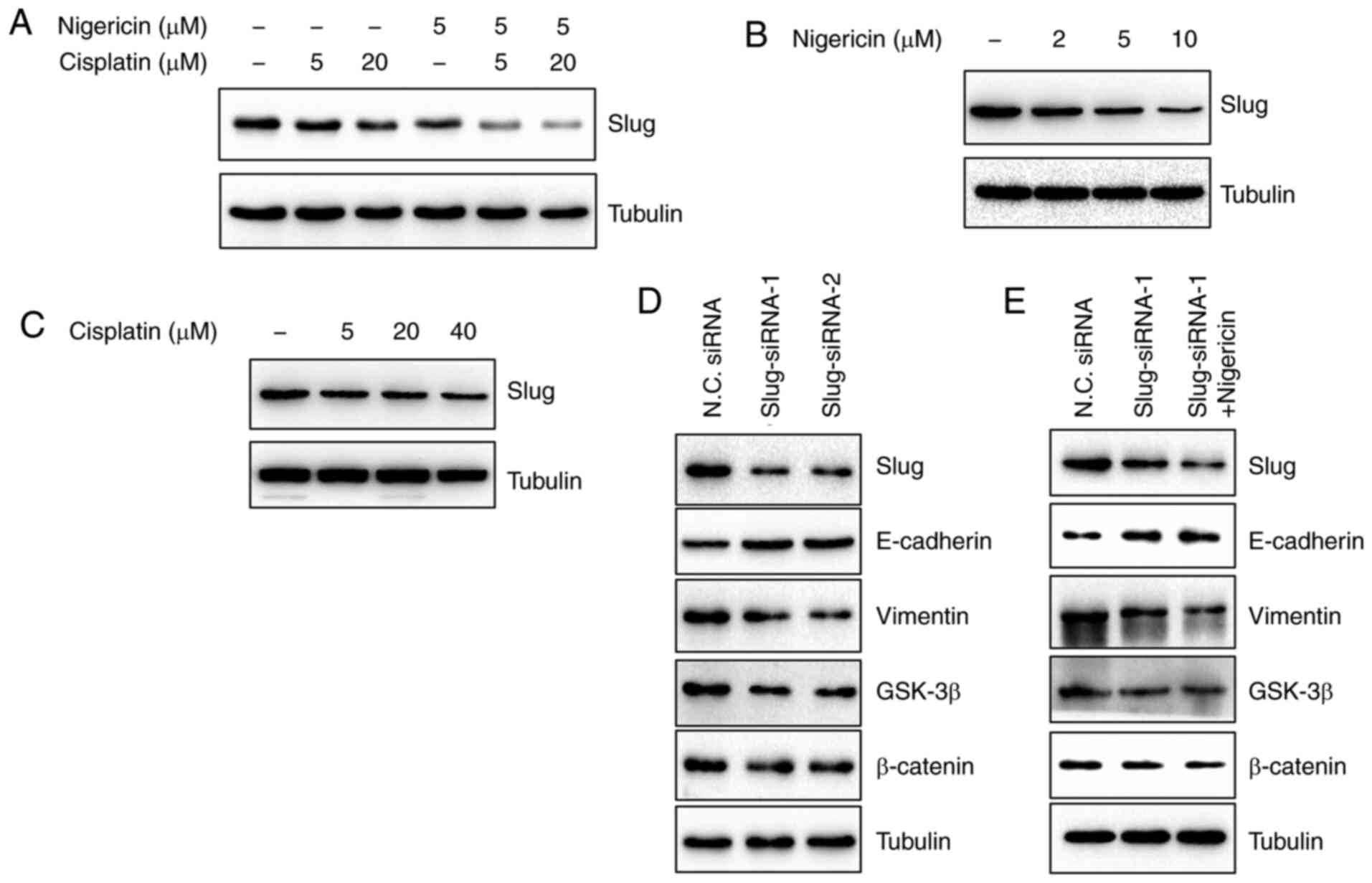
Nigericin combined with cisplatin
enhances the inhibition of the Wnt/β-catenin signalling pathway by
inhibiting slug expression
Slug is a transcription factor, which is increased
by the activation of the Wnt/β-catenin signalling pathway (19). To determine whether slug was
regulated by the combined effects of nigericin and cisplatin on the
Wnt/β-catenin signalling pathway, the present study firstly
examined slug expression following the treatment of SK-OV-3 cells
with nigericin and cisplatin by western blotting. The results
revealed that nigericin combined with cisplatin inhibited slug
expression (Fig. 4A). Subsequently,
the expression levels of slug in SK-OV-3 cells treated with
nigericin or cisplatin alone were detected, respectively. The
results revealed that slug expression was decreased following
treatment with nigericin in a concentration-dependent manner, while
slug protein expression was slightly altered by cisplatin (Fig. 4B and C). Furthermore, when siRNA was
used to suppress slug expression and the Wnt signalling pathway
(Fig. 4D), the inhibitory effect was
not as effective as that of treatment with 5 µM nigericin shown in
Fig. 2C. The inhibitory effect on
the Wnt signalling pathway was more obvious when 5 µM nigericin was
added (Fig. 4E).
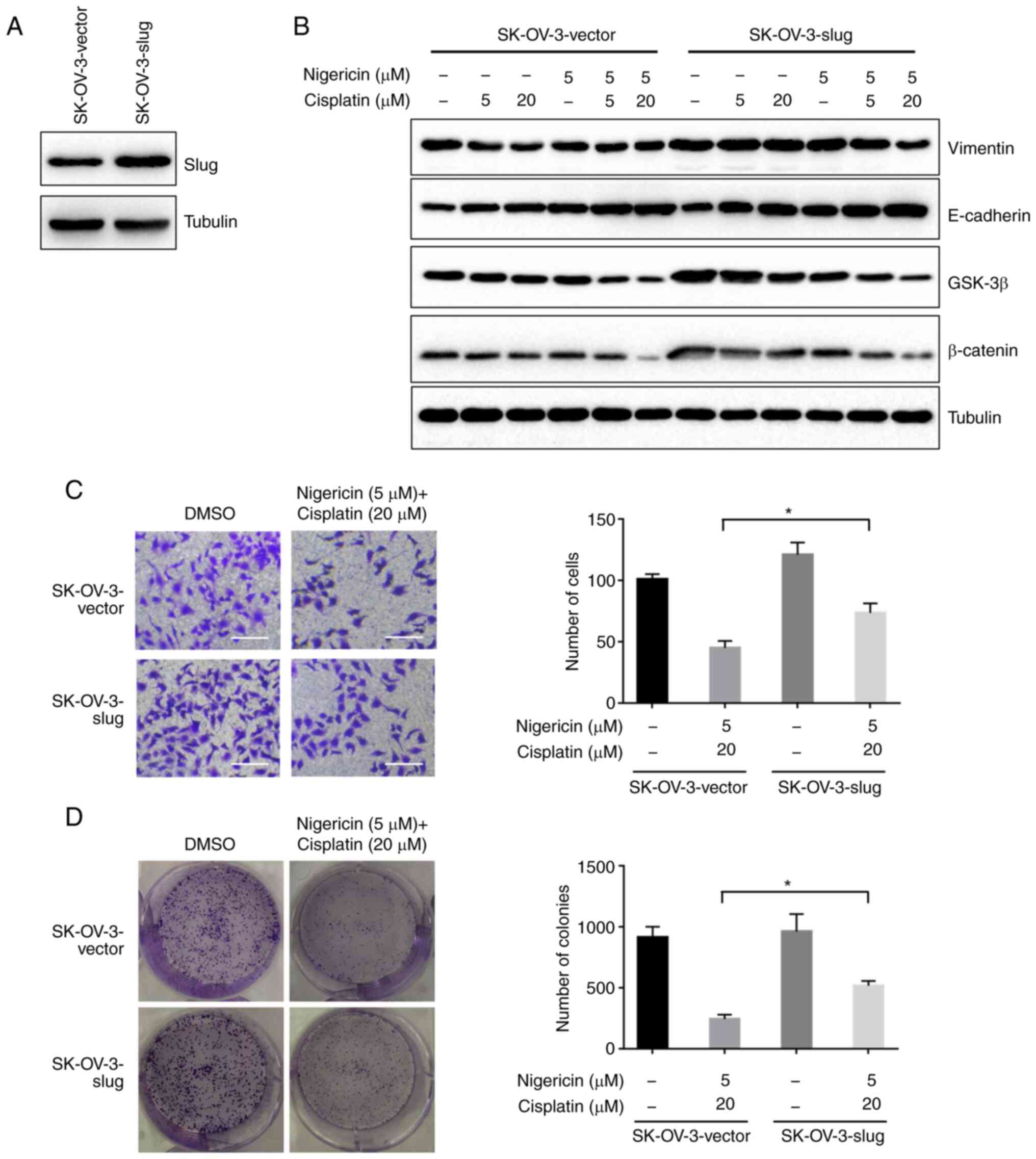
Subsequently, SK-OV-3-slug cells with stable
overexpression of slug were generated (Fig. 5A) to rescue the phenotype induced by
the combined therapy. The extent of the decreased expression of
vimentin, GSK-3β and β-catenin, and the increase in E-cadherin was
less in the SK-OV-3-slug cells, compared with SK-OV-3-vector cells
with the same nigericin and cisplatin treatment. It is suggested
that the inhibition of the combined effect on the Wnt/β-catenin
pathway could be reduced by the overexpression of slug (Fig. 5B). Furthermore, the inhibitory effect
on the migration and colony formation was also decreased (Fig. 5C and D). A2780 cells with stable
overexpression of slug were also generated (Fig. S1A). Similar results were observed in
migration and colony formation assays following slug overexpression
(Fig. S1B and C). These results
suggested a pivotal role of slug in the combined therapy of
nigericin and cisplatin suppressing EMT. They also implied the
important role served by nigericin in the suppression of EMT in the
combined therapy in EOC cells.
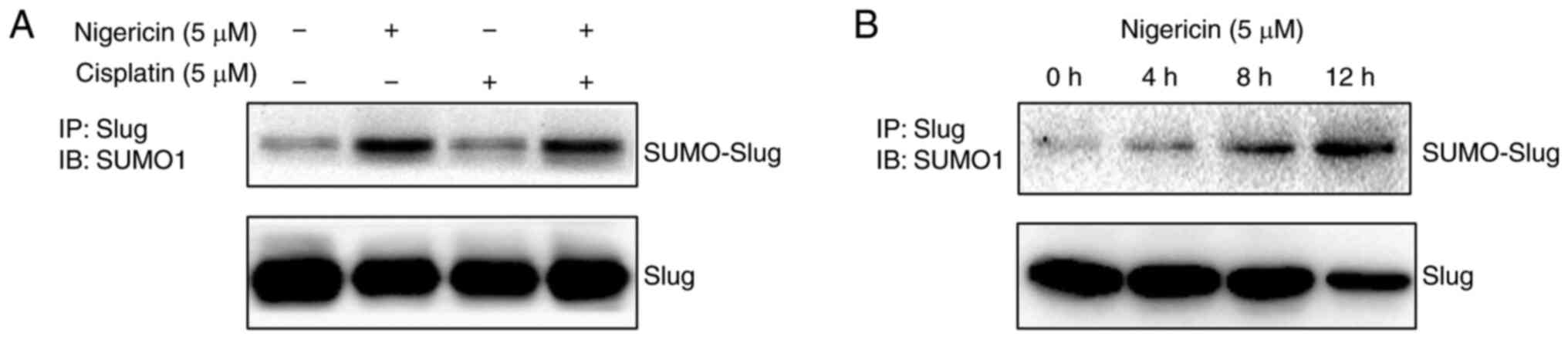
Nigericin inhibits slug expression by
enhancing its SUMOylation
Considering that nigericin could enhance the
knockdown effect of siRNA on slug, nigericin may regulate slug
expression in a post-transcriptional manner. Subsequently, the
present study investigated whether nigericin degraded slug through
SUMOylation in SK-OV-3 cells. The results demonstrated that
nigericin, but not cisplatin, promoted the SUMOylation of slug
(Fig. 6A) in a time-dependent manner
(Fig. 6B).
Discussion
Although platinum is still the first-line therapy
for post-operative chemotherapy (8),
it is a major obstacle in the effective treatment of EOC that
patients usually relapse with a tumour that is resistant to
chemotherapy with cisplatin or carboplatin (20,21).
Therefore, exploring appropriate drugs that could be combined with
platinum for post-operative chemotherapy instead of the single
platinum used in dealing with EOC is urgent (9). A previous study has demonstrated that
nigericin can inhibit the EMT process during cell invasion and
metastasis (17), indicating that
nigericin might be an effective chemotherapeutic drug in patients
with metastatic EOC. However, to the best of our knowledge, the
potential inhibitory effect of nigericin combined with cisplatin on
EOC cells has not been studied comprehensively.
In the present study, the combined use of nigericin
and cisplatin exhibited enhanced effects in the inhibition of the
migration and colony formation of EOC cells. This was mainly due to
the inhibitory effect of nigericin on the Wnt/β-catenin signalling
pathway. In addition, combination treatment exhibited more
significant inhibitory effects on the Wnt/β-catenin signalling
pathway compared with single drug treatment. In terms of the
molecular mechanism, nigericin inhibited slug expression by
promoting slug SUMOylation, which suppressed the activation of the
Wnt/β-catenin signalling pathway. In the rescue experiment, slug
overexpression reduced the inhibitory effect of nigericin on the
Wnt/β-catenin signalling pathway, as well as the inhibition of
migration and colony formation of EOC cells. Therefore, combination
treatment with two drugs could have an improved effect in EOC
therapy.
However, drugs with the same molecular mechanism are
not recommended (4). A meta-analysis
that used individual patient data from random control tests
compared single-agent platinum with a platinum-combination in
sensitive recurrent EOC (7,8,22–25). The
results revealed that combination-platinum chemotherapy may improve
the overall survival and progression-free survival; however,
platinum combinations inevitably cause greater toxicity in some
patients with cancer. Furthermore, combination-platinum
chemotherapy is only performed for platinum-sensitive EOC, which is
more likely to develop platinum resistance (8). The combination of different mechanism
drugs could inhibit tumour progress via more than one pathway,
which could be an improved strategy for drug development. The
present results suggested that slug might be the potential
molecular target of nigericin, and was also involved in the
combined effect of nigericin and cisplatin in EOC cells. Moreover,
the mechanism of cisplatin is generating unrepairable DNA lesions,
which may be different from that of nigericin.
Slug is an EMT-related gene that triggers the
initial phases of the EMT process (26–28) via
transcriptional reprogramming (29–31). As
a major molecular mechanism of epithelial cancer development, EMT
has been considered the footstone for metastasis and invasion
during the oncogenic process (32–34). The
present study not only demonstrated that nigericin could inhibit
the EMT process by increasing E-cadherin expression and reducing
vimentin expression by inhibiting slug, which was consistent with
previous studies (17,35), but also revealed the more significant
synergistic effect in the combined treatment with cisplatin.
Considering that slug is a transcription factor that can be
regulated by the Wnt/β-catenin signalling pathway (19), the present results suggested that
crosstalk might occur between the Wnt signalling cascade and the
DNA repair pathways. To the best of our knowledge, this report was
the first to address the synergistic effect of the combined
treatment with nigericin and cisplatin in cancer cells. Nigericin
could also enhance the SUMOylation of slug, which might be the
underlying mechanism of the synergistic effect. Aside from
ubiquitination, SUMOylation mediated by SUMO1 or SUMO2/3 is another
gene regulation pattern at the post-translational level in the
regulation of kinase activity, protein trafficking and stability
(36,37). On the one hand, pathogens could use
the characteristics of proteins to degrade important kinase or
transcription factors by enhancing their SUMOylation, which may
serve a key role in pathogen-related oncogenic transformation
(38). On the other hand, such
characteristics could also be used to develop drugs to inhibit
oncogene expression by enhancing SUMOylation. Although interesting
results have been revealed, the potential crosstalk of the related
pathways and the precise mechanism of the synergistic effect of
combined nigericin and cisplatin treatment remain to be
explored.
In conclusion, the present data revealed that
nigericin combined with cisplatin might enhance the in vitro
inhibitory effect of cisplatin on EOC metastasis by inhibiting slug
expression via the Wnt/β-catenin signalling pathway. In the future,
the underlying mechanisms of the synergistic and in vivo
effects of the combined therapy need to be elucidated in
detail.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Shandong
Natural Science Foundation (grant no. ZR2017PH029).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SY and SZ conceived and designed the experiments.
BZ, JL, SY and SZ confirm the authenticity of all the raw data. BZ,
CW, JL and SY performed the experiments. BZ, XL and BW analysed the
data. JL and SY contributed reagents/materials/analysis tools. BZ,
CW and XL wrote the manuscript. XL and SZ reviewed the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reid BM, Permuth JB and Sellers TA:
Epidemiology of ovarian cancer: A review. Cancer Biol Med. 14:9–32.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Colombo N, Parma G, Zanagnolo V and
Insinga A: Management of ovarian stromal cell tumors. J Clin Oncol.
25:2944–2951. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Damia G and Broggini M: Platinum
resistance in ovarian cancer: Role of DNA repair. Cancers (Basel).
11:1192019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tomao F, D'Incalci M, Biagioli E,
Peccatori FA and Colombo N: Restoring platinum sensitivity in
recurrent ovarian cancer by extending the platinum-free interval:
Myth or reality? Cancer. 123:3450–3459. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lheureux S, Gourley C, Vergote I and Oza
AM: Epithelial ovarian cancer. Lancet. 393:1240–1253. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Raja FA, Counsell N, Colombo N, Pfisterer
J, du Bois A, Parmar MK, Vergote IB, Gonzalez-Martin A, Alberts DS,
Plante M, et al: Platinum versus platinum-combination chemotherapy
in platinum-sensitive recurrent ovarian cancer: A meta-analysis
using individual patient data. Ann Oncol. 24:3028–3034. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Parmar MK, Ledermann JA, Colombo N, du
Bois A, Delaloye JF, Kristensen GB, Wheeler S, Swart AM, Qian W,
Torri V, et al: Paclitaxel plus platinum-based chemotherapy versus
conventional platinum-based chemotherapy in women with relapsed
ovarian cancer: The ICON4/AGO-OVAR-2.2 trial. Lancet.
361:2099–2106. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Borella F, Ghisoni E, Giannone G, Cosma S,
Benedetto C, Valabrega G and Katsaros D: Immune checkpoint
inhibitors in epithelial ovarian cancer: An overview on efficacy
and future perspectives. Diagnostics (Basel). 10:1462020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kaushik V, Yakisich JS, Kumar A, Azad N
and Iyer AKV: Ionophores: Potential use as anticancer drugs and
chemosensitizers. Cancers (Basel). 10:3602018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harned RL, Hidy PH, Corum CJ and Jones KL:
Nigericin a new crystalline antibiotic from an unidentified
Streptomyces. Antibiot Chemother (Northfield). 1:594–596.
1951.PubMed/NCBI
|
|
12
|
Liu F, Li W, Hua S, Han Y, Xu Z, Wan D,
Wang Y, Chen W, Kuang Y, Shi J and Zhi Q: Nigericin exerts
anticancer effects on human colorectal cancer cells by inhibiting
wnt/β-catenin signaling pathway. Mol Cancer Ther. 17:952–965. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yakisich JS, Azad N, Kaushik V, O'Doherty
GA and Iyer AK: Nigericin decreases the viability of
multidrug-resistant cancer cells and lung tumorspheres and
potentiates the effects of cardiac glycosides. Tumour Biol.
39:10104283176943102017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Boesch M, Zeimet AG, Rumpold H, Gastl G,
Sopper S and Wolf D: Drug transporter-mediated protection of cancer
stem cells from ionophore antibiotics. Stem Cells Transl Med.
4:1028–1032. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deng CC, Liang Y, Wu MS, Feng FT, Hu WR,
Chen LZ, Feng QS, Bei JX and Zeng YX: Nigericin selectively targets
cancer stem cells in nasopharyngeal carcinoma. Int J Biochem Cell
Biol. 45:1997–2006. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hrgovic I, Glavic Z, Kovacic Z, Mulic S,
Zunic L and Hrgovic Z: Repeated administration of inhibitors for
ion pumps reduce markedly tumor growth in vivo. Med Arch. 68:76–78.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang W, Zhao Y, Yao S, Cui X, Pan W, Huang
W, Gao J, Dong T and Zhang S: Nigericin inhibits epithelial ovarian
cancer metastasis by suppressing the cell cycle and
epithelial-mesenchymal transition. Biochemistry (Mosc). 82:933–941.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Karam A and Dorigo O: MMPs in ovarian
cancer as therapeutic targets. Anticancer Agents Med Chem.
12:764–772. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nieszporek A, Skrzypek K, Adamek G and
Majka M: Molecular mechanisms of epithelial to mesenchymal
transition in tumor metastasis. Acta Biochim Pol. 66:509–520.
2019.PubMed/NCBI
|
|
20
|
Kulshrestha A, Katara GK, Ibrahim SA,
Riehl V, Sahoo M, Dolan J, Meinke KW, Pins MR and Beaman KD:
Targeting V-ATpase isoform restores cisplatin activity in resistant
ovarian cancer: Inhibition of autophagy, endosome function, and
ERK/MEK pathway. J Oncol. 2019:23438762019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Galluzzi L, Vitale I, Michels J, Brenner
C, Szabadkai G, Harel-Bellan A, Castedo M and Kroemer G: Systems
biology of cisplatin resistance: Past, present and future. Cell
Death Dis. 5:e12572014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pfisterer J, Plante M, Vergote I, du Bois
A, Hirte H, Lacave AJ, Wagner U, Stahle A, Stuart G, Kimmig R, et
al: Gemcitabine plus carboplatin compared with carboplatin in
patients with platinum-sensitive recurrent ovarian cancer: An
intergroup trial of the AGO-OVAR, the NCIC CTG, and the EORTC GCG.
J Clin Oncol. 24:4699–4707. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gonzalez-Martin AJ, Calvo E, Bover I,
Rubio MJ, Arcusa A, Casado A, Ojeda B, Balana C, Martinez E,
Herrero A, et al: Randomized phase II trial of carboplatin versus
paclitaxel and carboplatin in platinum-sensitive recurrent advanced
ovarian carcinoma: A GEICO (Grupo Espanol de Investigacion en
Cancer de Ovario) study. Ann Oncol. 16:749–755. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alberts DS, Liu PY, Wilczynski SP, Clouser
MC, Lopez AM, Michelin DP, Lanzotti VJ and Markman M; Southwest
Oncology G, : Randomized trial of pegylated liposomal doxorubicin
(PLD) plus carboplatin versus carboplatin in platinum-sensitive
(PS) patients with recurrent epithelial ovarian or peritoneal
carcinoma after failure of initial platinum-based chemotherapy
(Southwest Oncology Group Protocol S0200). Gynecol Oncol.
108:90–94. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bolis G, Scarfone G, Giardina G, Villa A,
Mangili G, Melpignano M, Presti M, Tateo S, Franchi M, Parazzini F,
et al: Carboplatin alone vs carboplatin plus epidoxorubicin as
second-line therapy for cisplatin-or carboplatin-sensitive ovarian
cancer. Gynecol Oncol. 81:3–9. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Savagner P, Yamada KM and Thiery JP: The
zinc-finger protein slug causes desmosome dissociation, an initial
and necessary step for growth factor-induced epithelial-mesenchymal
transition. J Cell Biol. 137:1403–1419. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Phillips S and Kuperwasser C: SLUG:
Critical regulator of epithelial cell identity in breast
development and cancer. Cell Adh Migr. 8:578–587. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ye X, Tam WL, Shibue T, Kaygusuz Y,
Reinhardt F, Ng Eaton E and Weinberg RA: Distinct EMT programs
control normal mammary stem cells and tumour-initiating cells.
Nature. 525:256–260. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fang JH, Zhou HC, Zhang C, Shang LR, Zhang
L, Xu J, Zheng L, Yuan Y, Guo RP, Jia WH, et al: A novel vascular
pattern promotes metastasis of hepatocellular carcinoma in an
epithelial-mesenchymal transition-independent manner. Hepatology.
62:452–465. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu X, Sun H, Qi J, Wang L, He S, Liu J,
Feng C, Chen C, Li W, Guo Y, et al: Sequential introduction of
reprogramming factors reveals a time-sensitive requirement for
individual factors and a sequential EMT-MET mechanism for optimal
reprogramming. Nat Cell Biol. 15:829–838. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: Emt: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lu D, Choi MY, Yu J, Castro JE, Kipps TJ
and Carson DA: Salinomycin inhibits Wnt signaling and selectively
induces apoptosis in chronic lymphocytic leukemia cells. Proc Natl
Acad Sci USA. 108:13253–13257. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Geiss-Friedlander R and Melchior F:
Concepts in sumoylation: A decade on. Nat Rev Mol Cell Biol.
8:947–956. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kho C, Lee A, Jeong D, Oh JG, Chaanine AH,
Kizana E, Park WJ and Hajjar RJ: SUMO1-dependent modulation of
SERCA2a in heart failure. Nature. 477:601–605. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chand V, John R, Jaiswal N, Johar SS and
Nag A: High-risk HPV16E6 stimulates hADA3 degradation by enhancing
its SUMOylation. Carcinogenesis. 35:1830–1839. 2014. View Article : Google Scholar : PubMed/NCBI
|