Introduction
Gastric cancer has the fifth highest incidence among
all cancers according to GLOBOCAN 2020 data. Globally, it is the
fourth most frequent cause of death among malignancies (1). Although the incidence of gastric cancer
is decreasing, it remains a significant public health problem
(2). Gastric cancer is commonly
diagnosed at an advanced stage, except in Japan and Korea where
routine screening is performed (3,4). The
efficacy of chemotherapy for advanced-stage gastric cancer has
increased the survival rate in recent years; however, the degree of
improvement is unsatisfactory (5,6).
The Hippo signaling pathway is essential in the
control of organ size. It consists of a series of serine/threonine
kinases and scaffolding proteins that regulate the subcellular
localization and activity of the effector proteins yes-associated
protein (YAP) and transcriptional co-activator with a PDZ-binding
domain (TAZ) (7). Overexpression of
YAP is observed frequently in a variety of cancer types (8). YAP expression is elevated in gastric
adenocarcinomas, and its knockdown inhibits gastric cancer cell
proliferation (9). High levels of
YAP in the nucleus have been linked to chemoresistance in various
cancer types (10–13). In gastric cancer cells, YAP is
associated with cisplatin and trastuzumab resistance (14,15).
Upregulation of YAP correlates with progression, metastasis and
poor prognosis in patients with gastric carcinoma (16). Moreover, the expression of TAZ, a YAP
paralog, is elevated in gastric signet ring cell carcinoma
(17).
Targeting YAP and TAZ, individually or in
combination, is likely to have a clinical impact on gastric cancer.
Verteporfin (VP), a benzoporphyrin derivative, is used in
photodynamic therapy (PDT) for exudative age-related macular
degeneration; specifically, against choroidal neovascularization
(18). We have recently reported the
efficacy of VP in PDT using gastric cancer cells (19). Other previous studies propose that VP
may inhibit cancer cell growth in the absence of photoactivation by
inhibiting the YAP/TAZ transcriptional enhanced associate domain
(YAP/TAZ-TEAD) complex (20–22). In the present study, the aim was to
investigate the effects of VP on two gastric cancer cell lines:
MKN-45 (TAZ-dominant) and MKN-74 (YAP-dominant).
Materials and methods
Clinical samples and datasets
The gene expression patterns of YAP and TAZ in
cancer and normal tissue were compared on the Gene Expression
Profiling Interactive Analysis (GEPIA) server developed by the
Zhang Lab at Peking University (http://gepia.cancer-pku.cn/). The survival analysis of
YAP and TAZ was performed on the Kaplan-Meier Plotter online tool
headquartered at the Semmelweis University in Budapest (http://kmplot.com/analysis/index.php?p=service&cancer=gastric).
Tissue images are obtained from the Human Protein Atlas (https://www.proteinatlas.org/).
Reagents and antibodies
VP (cat. no. SML0534) was purchased from
Sigma-Aldrich; Merck KGaA. The following primary antisera were used
for the immunoblotting analysis: Anti-β-actin antibody (cat. no.
13E5; Cell Signaling Technology, Inc.), anti-YAP/TAZ antibody (cat.
no. D24E4; Cell Signaling Technology, Inc), anti-Survivin antibody
(cat. no. EP2880Y; Abcam), anti-CTGF antibody (cat. no. L-20; Santa
Cruz Biotechnology, Inc.), and anti-CYR61/CCN1 antibody (cat. no.
ab24448; Abcam). The following primary monoclonal antibodies were
used for immunofluorescence: Anti-YAP antibody (cat. no. sc-101199;
Santa Cruz Biotechnology, Inc.), anti-TAZ antibody (cat. no.
ab84927; Abcam), and anti-CD31 antibody (cat. no. ab28364; Abcam).
Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (cat.
no. ab6789; Abcam) and goat anti-rabbit IgG H&L (cat. no.
ab97051; Abcam) were purchased for western blotting. Goat
anti-mouse IgG H&L (Alexa Fluor® 488; cat. no.
ab150113; Abcam) was purchased for immunofluorescence.
Cell lines and cultures
Kato III (derived from a signet ring cell carcinoma
with no further information), NUGC-4 (derived from an
adenocarcinoma, poorly differentiated, signet ring cell carcinoma
with lymph node metastasis in a 32-year-old female), MKN-45-Luc
(derived from a poorly differentiated adenocarcinoma in a
62-year-old female), MKN-74/CMV-Luc cells (derived from a
moderately differentiated tubular adenocarcinoma with liver
metastasis in a 37-year-old male), and PANC-1 (TAZ-dominant
pancreatic cancer cell line) were obtained from the JCRB Cell Bank.
The Kato III cell line was grown without antibiotics in Roswell
Park Memorial Institute (RPMI)-1640 (FUJIFILM Wako Pure Chemical
Corporation) medium with Eagle's minimal essential medium (E-MEM;
FUJIFILM Wako Pure Chemical Corporation), supplemented with 10%
fetal bovine serum (FBS; Biosera) and 1% L-glutamine solution
(FUJIFILM Wako Pure Chemical Corporation). The other cell lines
were cultured without antibiotics in RPMI-1640 medium, supplemented
with 10% fetal bovine serum and 1% L-glutamine solution. The cells
were cultured in a humidified incubator with 5% CO2 at
37°C. Dimethyl sulfoxide (DMSO) was used as the solvent for VP, and
all the negative control groups were treated with a DMSO volume the
same as the highest concentration of VP solution. Aluminum foil was
used to protect cells treated with VP from light exposure, and all
experiments with samples containing VP were performed in the
dark.
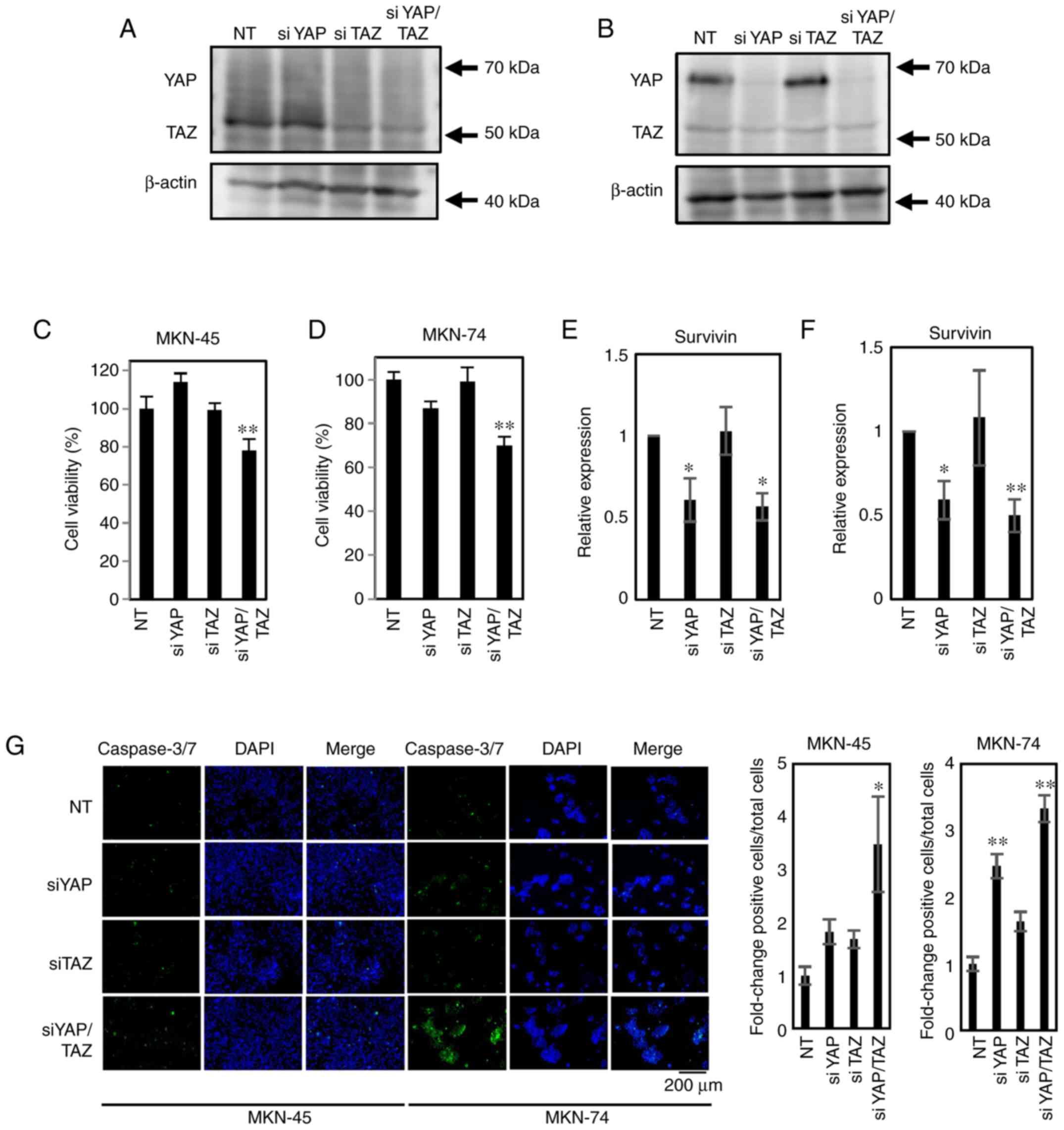
RNA interference and transfection
YAP and TAZ were transiently knocked down in the
MKN-74 and MKN-45 cell lines with validated siRNAs
[L-012200-00-0005 ON-TARGETplus Human YAP1 (cat. no. 10413)
siRNA-SMARTpool, L-016083-00-0005 ON-TARGETplus Human WW domain
containing transcription regulator 1 (WWTR1; also known as TAZ;
cat. no. 25937) siRNA-SMARTpool] from GE Healthcare Dharmacon
(Horizon Discovery Group). Universal non-targeting control pool of
four siRNAs (D-001810-10-05 ON-TARGETplus Non-targeting Pool) from
GE Healthcare Dharmacon (Horizon Discovery Group) were used for the
negative control group (NT). All the siRNA sequences used in the
present study are listed in Table
II. The cells were grown on 96-well or 6-well plates and
transfected (final concentration, 60 nM) with Lipofectamine RNAiMAX
reagent per the manufacturer's instructions (Thermo Fisher
Scientific, Inc.), incubated at 37°C for 96 h, and used for MTS
assays, RT-qPCR and caspase-3/7 assays. The efficiency and
specificity of the knockdown were assayed by western blotting.
 | Table II.Sequences of siRNA
oligonucleotides. |
Table II.
Sequences of siRNA
oligonucleotides.
| Name | Target
sequences |
|---|
| Non-targeting |
5′-UGGUUUACAUGUCGACUAA-3′ |
| Pool (NT) |
5′-UGGUUUACAUGUUGUGUGA-3′ |
|
|
5′-UGGUUUACAUGUUUUCUGA-3′ |
|
|
5′-UGGUUUACAUGUUUUCCUA-3′ |
| Human YAP1 |
5′-GCACCUAUCACUCUCGAGA-3′ |
| siRNA |
5′-UGAGAACAAUGACGACCAA-3′ |
| SMART pool |
5′-GGUCAGAGAUACUUCUUAA-3′ |
| (si-YAP) |
5′-CCACCAAGCUAGAUAAAGA-3′ |
| Human WWTR1 |
5′-CCGCAGGGCUCAUGAGUAU-3′ |
| siRNA |
5′-GGACAAACACCCAUGAACA-3′ |
| SMART pool |
5′-AGGAACAAACGUUGACUUA-3′ |
| (si-TAZ) |
5′-CCAAAUCUCGUGAUGAAUC-3′ |
Cell viability assay
VP-induced changes in gastric cancer cell viability
were assessed using an MTS assay. MKN-45 and MKN-74 cells were
seeded into 96-well plates at a density of 5×104
cells/ml (200 µl/well) and incubated at 37°C for 24 h. Cells were
then treated with 10, 15 and 20 µM of VP and incubated at 37°C for
24, 48 and 72 h, respectively. Cell viability was measured by MTS
{3-(4,5-dimethylthiazol-2-yl)-5-[3-car-boxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium]
assay as follows: 100 µl of the culture medium and 20 µl of the
proliferation assay solution of a CellTiter 96® AQueous
One Solution Cell Proliferation assay (cat. no. G3580; Promega
Corporation) were combined and incubated at 37°C for 1 h.
Absorbance was then measured at 490 nm using a microplate reader
(Viento nano; Sumitomo Dainippon Pharma Co., Ltd.) with an
All-in-One Microplate reader Software (Gen 5 ver 2.00.18; BioTek),
and the viability of the sample relative to control cells was
calculated.
Western blot analysis
MKN-45 and MKN-74 cells were treated with 15 µM of
VP and incubated at 37°C for 24 h. Whole-cell lysates were
collected by adding ice-cold RIPA Lysis and Extraction Buffer (cat.
no. 89900; Thermo Fisher Scientific, Inc.) containing cOmplete™
ULTRA Tablets, EASYpack Protease Inhibitor Cocktail and PhoSTOP
(cat. nos. 05892970001 and 4906845001; Roche Diagnostics). Cells
were collected by scraping. Samples were vortexed and lysed on ice
for ~20 min. The lysed cells were centrifuged at 12,000 × g for 15
min at 4°C to remove cellular debris. Supernatants were transferred
to clean tubes, and protein concentrations were determined by a
Pierce 660 nm Protein Assay Reagent (cat. no. 1861426; Thermo
Fisher Scientific, Inc.). Proteins (30–50 µg/lane) were resolved by
4–20% SDS-PAGE and transferred to nitrocellulose membranes for
western blotting. Membranes were incubated with primary antibodies
at 4°C overnight in 5% BSA-TBS Tween. Primary antibody dilutions
were 1:1,000 unless otherwise indicated. Membranes were then washed
for 30 min in TBS-0.05% Tween. Membranes were incubated with
secondary antibodies (1:5,000) for 1 h at room temperature.
Immunoreactive proteins were visualized using Clarity Western ECL
substrate (cat. no. 1705061; Bio-Rad Laboratories, Inc.) and an
image analyzer (LAS-3000 mini; Fujifilm Co., Ltd.) with an Image
Reader (LAS-3000 UV mini ver2.2; Fujifilm Co., Ltd.). When multiple
proteins were evaluated, membranes were stripped using Restore PLUS
Western Blot Stripping Buffer (Thermo Fisher Scientific, Inc.)
before primary antibody incubation.
Crystal violet staining
VP-induced changes in gastric cancer cell viability
were also assessed by crystal violet (CV; cat. no. 031-04851;
FUJIFILM Wako Pure Chemical Corporation) staining. MKN-45 and
MKN-74 cells were seeded onto 6-well plates at a density of
3×105 cells/ml (1 ml/well) and incubated at 37°C for 24
h. Cells were treated with 10 and 15 µM of VP and incubated at 37°C
for 24 h. Cells were then fixed for 5 min with 4% paraformaldehyde
(PFA) and stained for 30 min at room temperature with 0.05% CV.
Samples were then washed twice with tap water and allowed to drain
in an inverted position for ~2 min. Staining was recorded by
photography, and then a volume of methanol equivalent to one-third
to one-half the total well volume was added and solubilize the dye
for 30 min at room temperature (20–25°C). The absorbance at 540 nm
was measured with aliquots transferred to a fresh plate using a
microplate reader (Viento nano; Sumitomo Dainippon Pharma Co.,
Ltd.) with an All-in-One Microplate reader Software (Gen 5 ver
2.00.18; BioTek).
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
Total RNA was extracted from cultured cells using an
miRNeasy Mini kit (cat. no. 217004; Qiagen GmbH) and quantified
using a Biospec-nano spectrophotometer (Shimadzu Corporation). The
extracted RNA samples were stored at −80°C until use. cDNAs were
prepared from total RNA using a High-Capacity cDNA Reverse
Transcription kit (cat. no. 4374966; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols. Reverse
transcription reaction mixtures contained 2 µg of total RNA, 1X RT
buffer, 4 mM dNTP mix, 1X RT random primer, 50 units MultiScribe
reverse transcriptase and 20 units RNase inhibitor. Nuclease-free
water was added to adjust the reaction volume to 20 µl. The
reaction mixtures were incubated at 25°C for 10 min, followed by
37°C for 120 min and 85°C for 5 min. The real-time-PCR assays were
performed in 20 µl aliquots containing 1 µl RT products with 4 µl
LightCycler® FastStart DNA Master PLUS SYBR Green I
(cat. no. 03515869001; Roche Diagnostics), 0.2 µl (final
concentration 0.5 µM) of each primer and 14.6 µl nuclease-free
water. Analyses were run on a Real-Time PCR Light
Cycler® 1.5 Complete System (Roche Diagnostics). Thermal
cycling was initiated with a denaturation step at 95°C for 10 min,
followed by 45 cycles of 95°C for 10 sec, 60°C for 10 sec, and 72°C
for 10 sec. The cycle threshold (Cq) was recorded for each target
mRNA by LightCycler® Software version 3.5.28 (Roche
Diagnostics), and β-actin was used as the endogenous control for
data normalization. The relative expression was calculated using
the formula 2−ΔΔCq=2−(ΔCq, reagent treatment−ΔCq,
control) (23). All the primer
sequences used in the present study are listed in Table I.
| Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer
sequences |
|---|
|
Survivin | F:
5′-CAAGGACCACCGCATCTCTAC-3′ |
|
| R:
5′-AGTCTGGCTCGTTCTCAGTGG-3′ |
| CTGF | F:
5′-CAGTGTCTGACTTCGACAACGC-3′ |
|
| R:
5′-CCATCGGCGTGTTTGGAGTA-3′ |
| CYR61 | F:
5′-GAGTGGGTCTGTGACGAGGAT-3′ |
|
| R:
5′-GGTTGTATAGGATGCGAGGCT-3′ |
| β-actin | F:
5′-GCATCCTCACCCTGAAGTA-3′ |
|
| R:
5′-TGTGGTGCCAGATTTTCTCC-3′ |
Verteporfin uptake evaluation
An all-in-one fluorescence microscope (BZ-X800;
Keyence Corporation) equipped with an OP-87767 filter (excitation:
405 nm and fluorescence: 630 nm) was used to evaluate VP uptake.
MKN-45 and MKN-74 cells were seeded onto 96-well plates at a
density of 5×104 cells/ml (200 µl/well) and incubated at
37°C for 24 h. Cells were then treated with 15 µM of VP and
incubated at 37°C for 30 min, 1 or 2 h. Pixel intensities were
measured using ImageJ software ver1.52a (Wayne Rasband; National
Institutes of Health) using 20 random microscopic fields
(magnification, ×20), according to the manufacturer's instructions
(24).
Immunofluorescence
MKN-45 and MKN-74 cells were seeded on glass
coverslips in 6-well plates at a density of 3×105
cells/ml (1 ml/well) and incubated at 37°C for 24 h. Cells were
then treated with 15 µM of VP and incubated at 37°C for 24 h. Cells
were fixed with 4% PFA and permeabilized with 0.1% Triton X-100 and
then incubated in blocking buffer containing 5.0% BSA and 0.1%
glycine at room temperature (20–25°C). Cells were incubated with
the following primary antibodies in blocking buffer at 4°C
overnight; anti-YAP antibody (cat. no. sc-101199; Santa Cruz
Biotechnology, Inc.), anti-TAZ antibody (cat. no. ab84927; Abcam),
and anti-CD31 antibody (cat. no. ab28364; Abcam). Cells were washed
and then incubated with corresponding secondary antibody, Goat
anti-mouse IgG H&L (Alexa Fluor® 488; cat. no.
ab150113; Abcam) in blocking buffer for 1 h at room temperature in
the dark. Cells were washed, and the coverslips were mounted onto
slides using ProLong Antifade Gold reagent with
4′,6-diamidino-2-phenylindole (DAPI; Thermo Fisher Scientific,
Inc.). Slides were analyzed with an all-in-one fluorescence
microscope (BZ-X800; Keyence Corporation). Pixel intensities were
measured using ImageJ software ImageJ software ver1.52a (Wayne
Rasband; National Institutes of Health) using 20 random cells
(magnification, ×20), according to the manufacturer's
instructions.
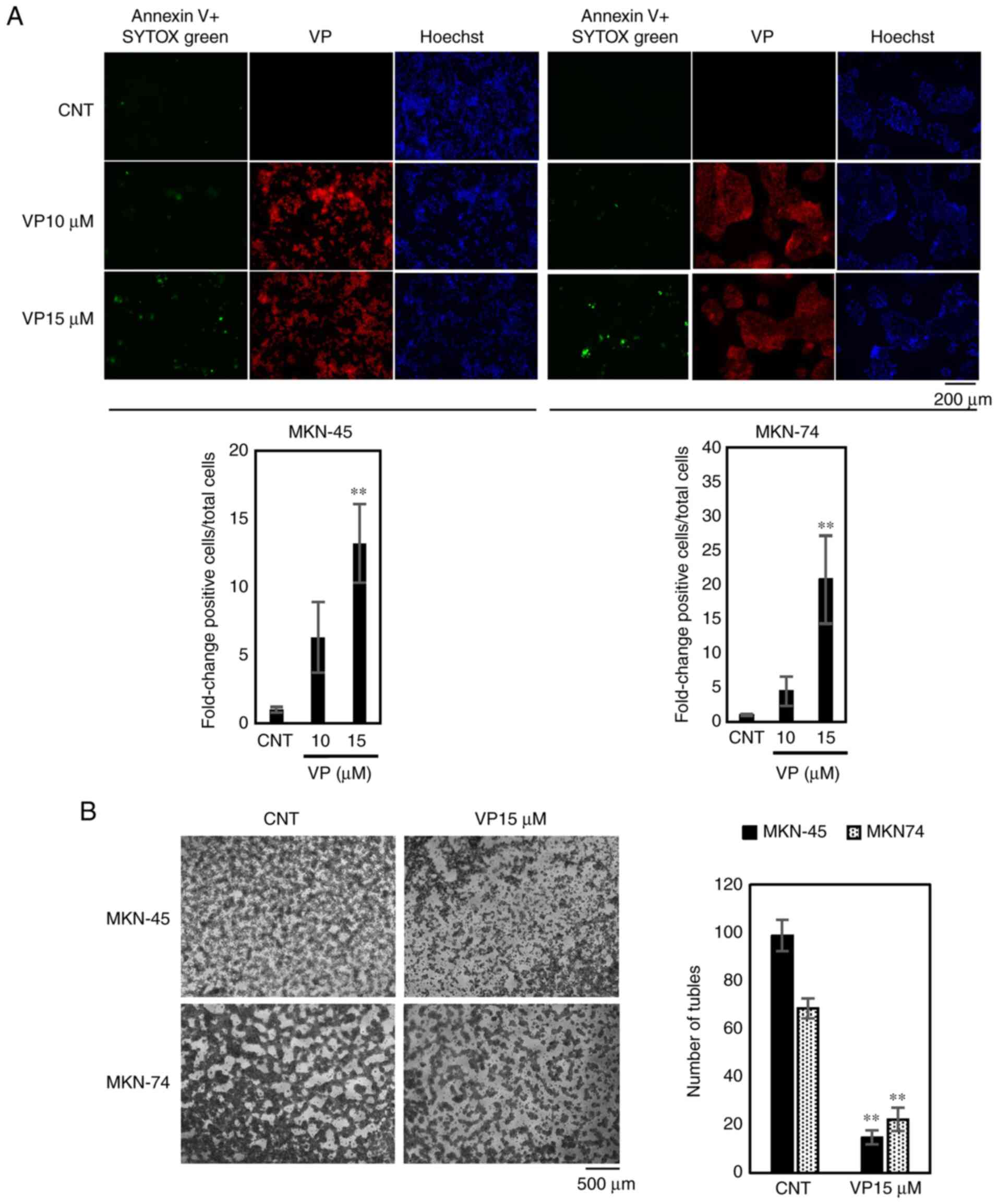
Matrigel-based tube formation
assay
An aliquot of growth factor-reduced Matrigel (BD
Biosciences) was warmed to room temperature, and then 50 µl/well
was transferred to 24-well plates on a horizontal surface to allow
even distribution. Plates were incubated for 15 min at 37°C. MKN-45
and MKN-74 cells (1×106) were resuspended with 1 ml
RPMI-1640 with 10% FBS and 20 µl of Matrigel and loaded onto the
Matrigel surface in a well. VP (15 µM) was added at the same time.
Each well was visualized under a microscope after 3 days at 37°C.
The average number of tubules counted manually in 10 random fields
from each well was calculated.
Quantification of cell death
Cell death was evaluated by staining with an Annexin
V-FITC Apoptosis Detection Kit Plus (BioVision, Inc.) and Hoechst
33342 stain (Thermo Fisher Scientific, Inc.). MKN-45 and MKN-74
cells were seeded into 96-well plates at a density of
5×104 cells/ml (200 µl/well) and incubated at 37°C for
24 h. Cells were then treated with 15 µM VP and incubated at 37°C
for 24 h. Annexin-V detection was performed according to the
manufacturer's protocol. Plates were analyzed with an all-in-one
fluorescence microscope (BZ-X800; Keyence Corporation). Dead cells
were quantified by calculating the ratio of Annexin V and SYTOX
green-positive nuclei and Hoechst 33342-positive nuclei (total
nuclei) in 20 random microscopic fields (magnification, ×20).
Quantification of caspase 3/7
activity
MKN-45 and MKN-74 cells were cultured on glass
coverslips. When the cultures achieved the desired confluency
(~50%), they were treated with siRNAs as described above. Caspase
activity was evaluated by staining (incubated at 37°C for 30 min)
with a Cell Event Caspase-3/7 Green Detection Reagent (Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
Coverslips were mounted with ProLong Gold Antifade reagent with
DAPI (Thermo Fisher Scientific, Inc.) and fixed overnight at room
temperature in the dark. Slides were analyzed with an all-in-one
fluorescence microscope (BZ-X800; Keyence Corporation).
Caspase-3/7-positive cells were quantified by calculating the ratio
of caspase-3/7-positive to DAPI-positive nuclei (total nuclei) in
10 random microscopic fields (magnification, ×20).
Statistical analysis
All data represent ≥ three independent experiments
using cells from a minimum of three separate isolations.
Skewness-Kurtosis was used to check the distribution of the data.
The Mann-Whitney U test was used to assess the statistical
significance of differences between two groups. Kruskal-Wallis test
was used for comparisons between groups and Dunn's test was applied
for multiple comparisons. Survival curves were calculated using the
Kaplan-Meier method on the Kaplan-Meier Plotter online tool.
Statistical analyses were performed using StatFlex software
(Windows ver. 6.0; Artech LLC), except for the Log-rank test for
survival analysis using the Kaplan-Meier Plotter online tool.
P<0.05 was considered to indicate a statistically significant
difference and all data are expressed as the mean ± SEM.
Results
YAP and TAZ are prognostic factors of
gastric cancer
YAP and TAZ expression levels in gastric cancer
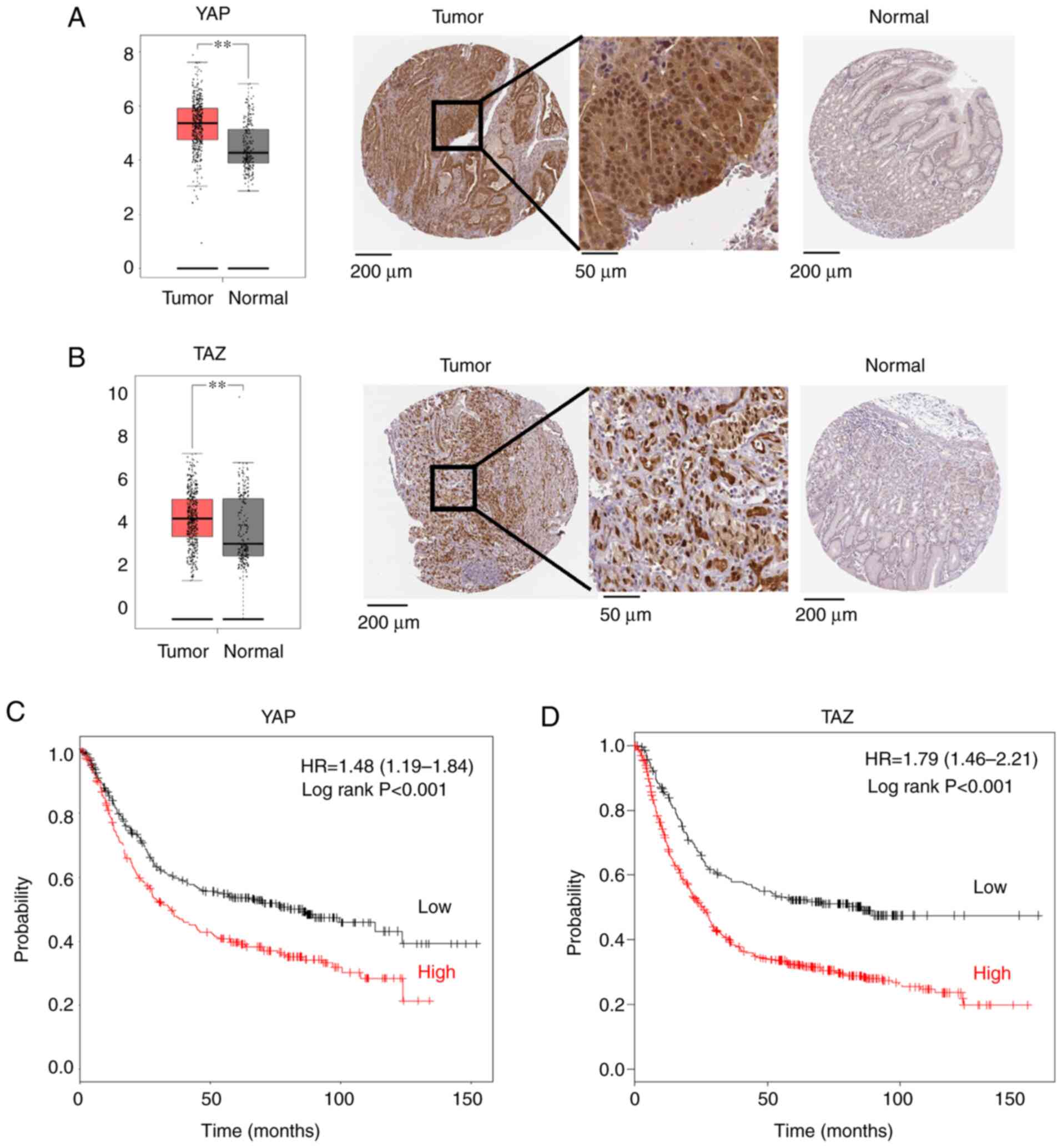
cells are significantly higher than those in normal tissues
(Fig. 1A and B). A higher expression
of YAP or TAZ in gastric cancer cells is associated with a
significantly poorer prognosis than cancer cells with lower
expression levels (Fig. 1C and
D).
Expression patterns of YAP and TAZ
differ among gastric cancer cell lines
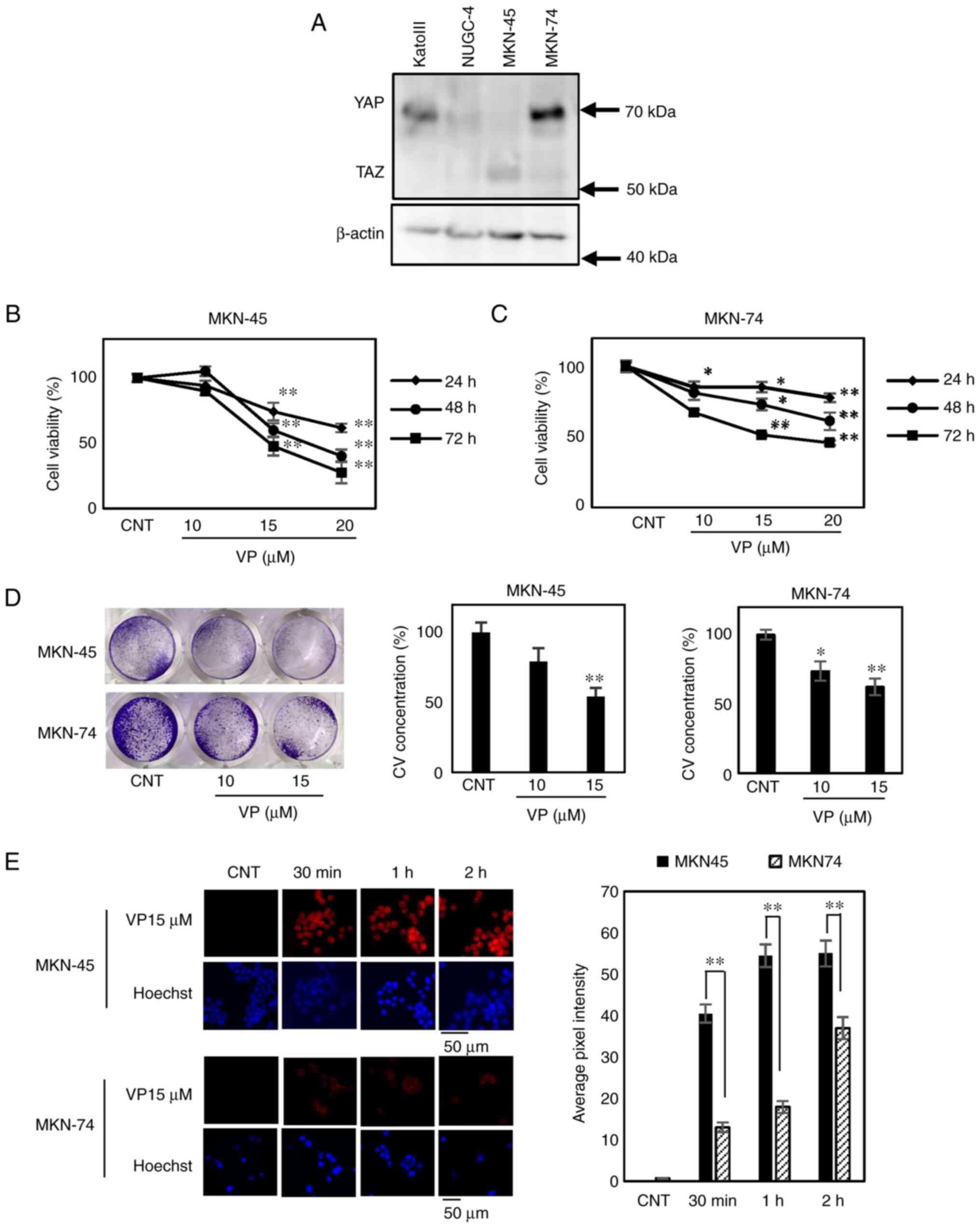
Initially, YAP and TAZ expression levels were
examined in four gastric cancer cell lines. Kato III, NUGC-4 and
MKN-74 cells exhibited a YAP-dominant expression pattern. By
contrast, the MKN-45 cell line showed a TAZ-dominant expression
pattern (Fig. 2A). Therefore, MKN-45
was selected as the TAZ-dominant cell line and MKN-74 as the
YAP-dominant cell line (Fig.
S1A).
VP suppresses MKN-45
proliferation
The effect of VP on proliferation in MKN-45 and
MKN-74 cells was examined using 10, 15 and 20 µM VP. VP treatment
suppressed proliferation in MKN-45 cells (Fig. 2B) more than it does in MKN-74 cells
at 24 h (Fig. 2C). In MKN-74 cells,
the same level of proliferation suppression was observed at 72 h.
VP treatment decreased the amount of CV staining in MKN-45 cells in
a dose-dependent manner over 24 h (Fig.
2D). VP uptake was faster in MKN-45 cells than MKN-74 cells
(Fig. 2E).
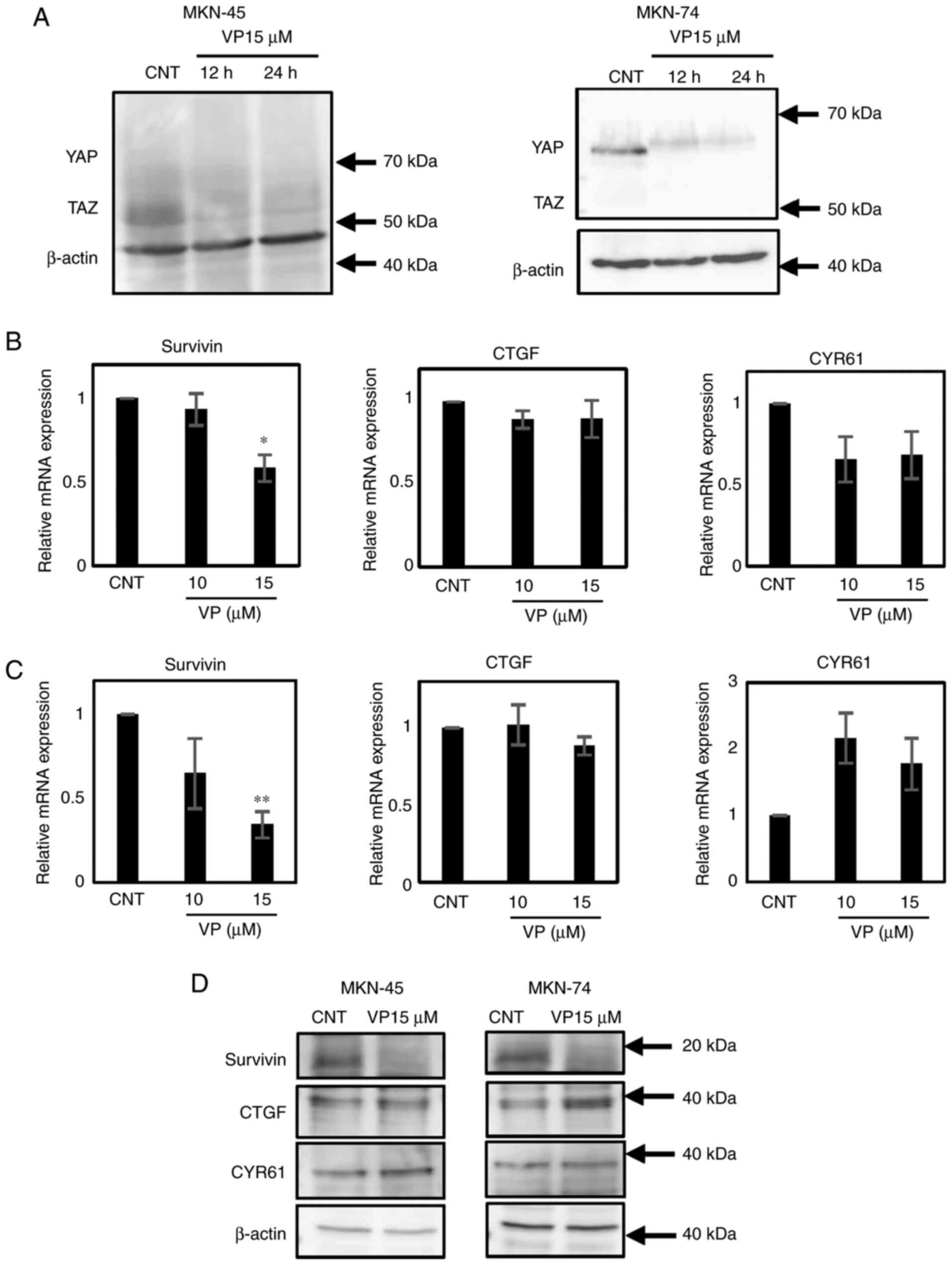
VP decreases YAP and TAZ proteins
after 12 h
Western blotting demonstrated that VP treatment for
12 h reduces the expression of YAP in MKN-74 cells and TAZ in
MKN-45 cells (Fig. 3A). PANC-1 cells
were used as a positive control for TAZ expression (Fig. S1A).
VP decreases Surivivin gene and
protein expression in MKN-45 and MKN-74 cells
VP treatment reduces the expression of downstream
Survivin gene in MKN-45 and MKN-74 cells at 24 h (Fig. 3B and C). Moreover, immunoblotting
confirmed that only Survivin expression is affected by VP treatment
at 24 h in both cell lines (Fig.
3D).
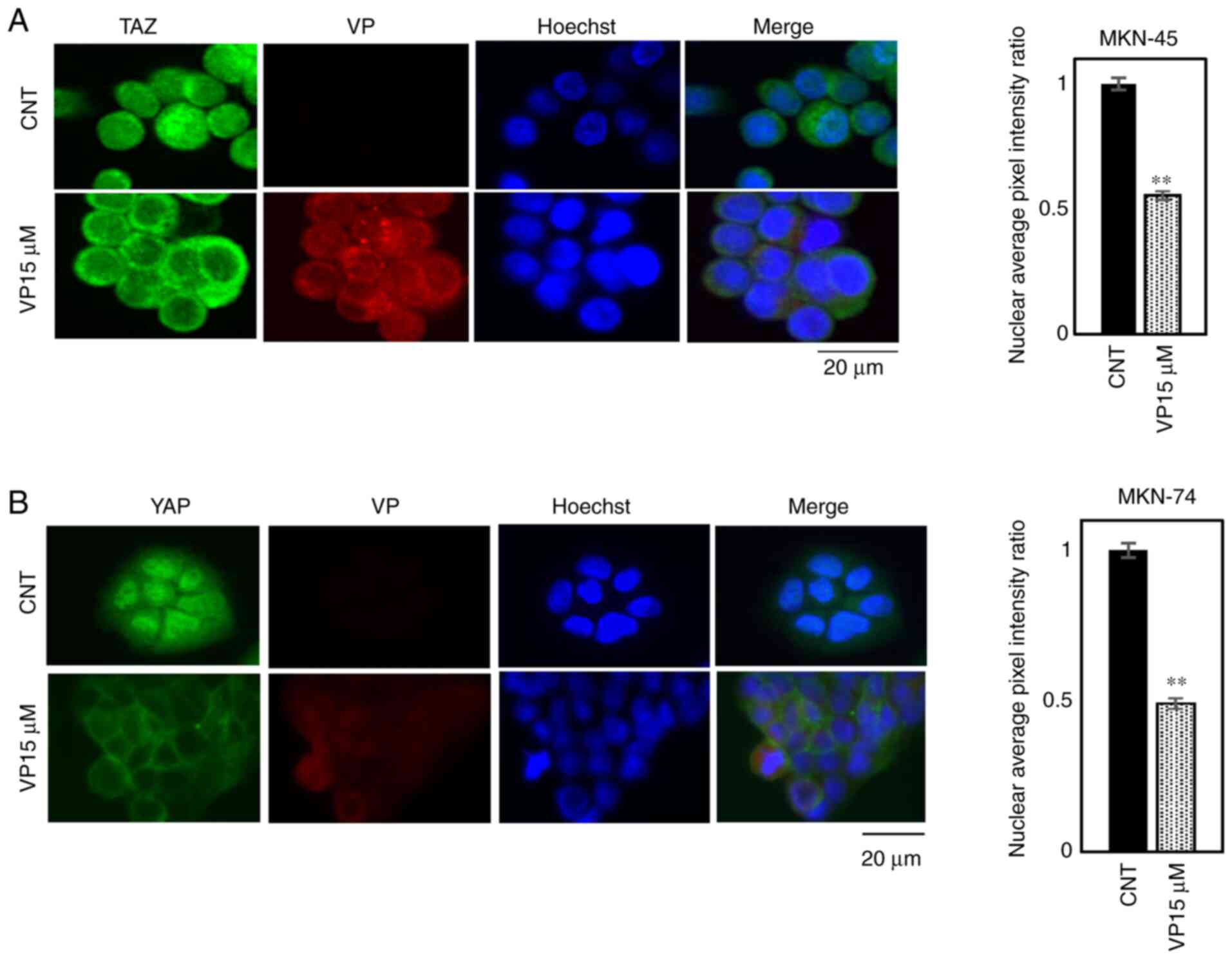
VP changes the localization of YAP and
TAZ from nuclear to cytosolic
Immunofluorescence analysis demonstrated that
exposure to 15 µM VP for 2 h changes YAP and TAZ localization from
the nucleus to the cytosol in MKN-45 and MKN-74 cells (Fig. 4A and B).
VP induces cell death in MKN-45 and
MKN-74 cells in a dose-dependent manner
Annexin V staining demonstrated that treatment with
15 µM of VP for 24 h induces cell death in MKN-45 and MKN-74 cells
in a dose-dependent manner (Fig.
5A).
VP suppresses vascular mimicry
MKN-45 and MKN-74 are CD31-positive cells that have
angiogenic and vasculogenic potential (Fig. S1B). Therefore, the vascular mimicry
of both cell lines was examined using tube formation assays. In
MKN-45 and MKN-74 cells, exposure to 15 µM of VP for 72 h
suppressed the level vascular mimicry, demonstrated by a decrease
in the number of tubules formed (Fig.
5B).
Concurrent YAP/TAZ knockdown decreases
cellular proliferation
RNA interference assays targeting YAP, TAZ and
YAP/TAZ in MKN-45 and MKN-74 cells demonstrated that simultaneous
knockdown (YAP/TAZ) is required to decrease cell proliferation
(Fig. 6A-D). YAP and YAP/TAZ
knockdown also decrease Survivin expression. Concurrent
YAP/TAZ knockdown increases caspase3/7 activity in both cell lines
(Fig. 6E-G).
Discussion
In the present study, it was demonstrated that VP
affects both YAP-dominant (MKN-74) and TAZ-dominant (MKN-45) cell
lines. Moreover, in the poorly differentiated, TAZ-dominant gastric
cancer cell line MKN-45, VP treatment elicited a more rapid
suppression than in MKN-74 cells.
VP is used in PDT as a photosensitizer to eliminate
abnormal blood vessels in the eyes of patients with conditions such
as macular degeneration (18). It
produces highly reactive, short-lived, singlet oxygen species when
stimulated by non-thermal red light at 689 nm. VP has also been
employed for PDT in oncology. A Phase I/II clinical trial of PDT
with VP was conducted in patients with locally advanced pancreatic
cancer in 2014 (25). However, light
penetration into abdominal tissue is an invasive process.
Therefore, the present study focused on gastric cancers because
these tumors are more accessible to PDT via endoscopy. Our previous
study reported the efficacy of VP in PDT using a gastric cancer
cell line (19). More recently, VP
has been reported to have an anti-cancer effect via YAP inhibition
in the absence of light activation (20–22,26,27).
Moreover, VP inhibited interactions between YAP/TAZ and the TEA
domain transcriptional factor (TEAD), and inhibited YAP function by
upregulating the 14-3-3σ sequestering of YAP in the cytoplasm
(28).
YAP and TAZ gene expression levels are elevated in a
subset of human gastric cancers, most of which are associated with
poor clinical outcomes (29–33). Choi et al (34) recently demonstrated that MYC
upregulation is a direct consequence of YAP activation in
YAP-activated human gastric cancer cells.
YAP and TAZ are usually discussed in the same vein.
However, in renal development, YAP-knockout (KO) mice are
embryonically lethal (35), whereas
TAZ-knockout mice develop severe cystic kidney disease (36). A study using YAP-KO, TAZ-KO and
YAP-/TAZ-KO cell lines generated by the CRISPR/Cas9 technique in
293A cells revealed that YAP inactivation has more significant
effects on cellular physiology than TAZ inactivation; namely, cell
spreading, volume, granularity, glucose uptake, proliferation and
migration (37). TAZ expression is
highly elevated in gastric signet ring cell carcinoma (17). Hayashi et al (38) reported an imbalance in TAZ and YAP
expression in gastrointestinal cancer cell lines. Another study
reported that TAZ accumulation is negatively regulated by YAP
abundance (39). The present results
suggest that concurrent suppression of YAP and TAZ inhibits cell
proliferation and that TAZ and YAP may serve distinct roles in
cancer progression.
In the present study, four gastric cancer cell lines
were used; one was TAZ-dominant and the other three were
YAP-dominant. Kato III and NUGC-4 are signet ring carcinoma cell
lines; however, YAP/TAZ dominance patterns do not depend on
histological classification. VP decreases YAP and TAZ expression
and subsequently reduces the expression of downstream genes,
predominantly Survivin. Survivin is a new member of the inhibitor
of apoptosis (IAP) family and is selectively upregulated in most
human cancers but not in normal tissues (40). Moreover, it exhibits anti-apoptotic
activity via caspase-3 and caspase-7 inhibition (41) and is a prognostic factor in gastric
cancer associated with lymphatic metastasis (42–44). A
correlation between YAP and Survivin in gastric cancer has been
demonstrated for tumorigenesis and lymph node metastasis (45). The current results indicate that VP
inhibits YAP/TAZ and induces apoptosis by increasing caspase-3 and
caspase-7 activity via decreasing Survivin expression in
gastric cancer cells.
Vascular mimicry (VM) is a concept proposed by
Maniotis et al (46) in 1999.
Tumors with vigorous VM are often aggressive and associated with a
poor prognosis (47). Evaluating the
therapeutic potential of inhibiting VM has critical clinical
importance. Sun et al (48)
demonstrated that VM is correlated with the differentiation, stage
and metastatic potential of tumors, in addition to less favorable
prognoses in clinical samples of gastric cancer. VP has been shown
to suppress VM by disrupting the YAP-TEAD complex (49,50).
CD31-positive cancer cells are reported to display VM (51), and the cell lines used in the present
study were CD31-positive (Fig.
S1B). Therefore, VM was evaluated in VP-treated cells using
tube formation assays and it was revealed that VP significantly
suppresses VM in MKN-45 and MKN-74 cells. Consequently, VP may
represent a promising target for anti-cancer therapy in
YAP-dominant or TAZ-dominant gastric cancers with poor prognostic
outcomes.
Kang et al (52) demonstrated that the
anti-proliferative effects of VP on gastric cancer cell lines are
associated with suppression of FAT1, another marker associated with
poor prognoses. Notably, more recent studies report that VP can
potentially treat chemo-resistant gastric cancer stem cells
(53,54). However, these studies did not
distinguish between YAP and TAZ and did not evaluate the effects of
VP on Survivin expression and vascular mimicry. In the present
study, characteristics that reveal underlying mechanisms of VP in
the suppression of gastric tumor cell growth were investigated.
One limitation of the present study was the high
concentration of VP required to achieve suppression. The plasma
concentration in human patients receiving liposomized VP
(Visudyne®) is ~2 µg/ml. The in vitro assays used
10 and 15 µM VP treatments, which equate to 5 and 7.5 µg/ml,
respectively, or 2–4 times more than a recommended clinical dose.
Liposomization can increase VP tissue uptake, leading to a high
concentration in the gastric cancer tissue.
In conclusion, VP has the potential to suppress
different types of gastric cancers via suppressing Survivin.
Further in vivo studies will be required to confirm the
potential of VP for use as a therapeutic agent.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The resources and facilities of the Faculty of
Medicine, Tottori University were used to conduct this study. This
research did not receive any specific grant from funding agencies
in the public, commercial or not-for-profit sectors.
Availability of data and materials
The datasets used and analyzed in this study are
available from the corresponding author upon reasonable
request.
Authors contributions
HT, ST, HY, TR, MY, and KT designed and implemented
the experiments. ST, TT, NT, and MT searched the literature and
analyzed and interpreted the data. ST, TT, NT, and MT were also
involved in writing, reviewing, and editing the manuscript. IH made
substantial contributions to the conception and design of the
project and gave final approval of the version to be published. HT
and ST confirmed the authenticity of all the raw data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN Estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sitarz R, Skierucha M, Mielko J, Offerhaus
GJA, Maciejewski R and Polkowski WP: Gastric cancer: Epidemiology,
prevention, classification and treatment. Cancer Manag Res.
10:239–248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takahashi T, Saikawa Y and Kitagawa Y:
Gastric cancer: Current status of diagnosis and treatment. Cancers
(Basel). 5:48–63. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sugano K: Screening of gastric cancer in
Asia. Best Pract Res Clin Gastroenterol. 29:895–905. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Japanese Gastric Cancer Association, .
Japanese gastric cancer treatment guidelines 2014 (ver. 4). Gastric
Cancer. 20:1–19. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wagner AD, Syn NL, Moehler M, Grothe W,
Yong WP, Tai BC, Ho J and Unverzagt S: Chemotherapy for advanced
gastric cancer. Cochrane Database Syst Rev.
8:CD0040642017.PubMed/NCBI
|
|
7
|
Pan D: Hippo signaling in organ size
control. Genes Dev. 21:886–897. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zanconato F, Cordenonsi M and Piccolo S:
YAP/TAZ at the roots of cancer. Cancer Cell. 29:783–803. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Xu ZP, Yang YC, Zhu JS, Zhou Z
and Chen WX: Expression of yes-associated protein in gastric
adenocarcinoma and inhibitory effects of its knockdown on gastric
cancer cell proliferation and metastasis. Int J Immunopathol
Pharmacol. 25:583–590. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zanconato F and Piccolo S: Eradicating
tumor drug resistance at its YAP -biomechanical roots. EMBO J.
35:459–461. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou Y, Wang Y, Zhou W, Chen T, Wu Q,
Chutturghoon VK, Lin B, Geng L, Yang Z, Zhou L and Zheng S: YAP
promotes multi-drug resistance and inhibits autophagy-related cell
death in hepatocellular carcinoma via the RAC1-ROS-mTOR pathway.
Cancer Cell Int. 19:1792019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song J, Xie LX, Zhang XY, Hu P, Long MF,
Xiong F, Huang J and Ye XQ: Role of YAP in lung cancer resistance
to cisplatin. Oncol Lett. 16:3949–3954. 2018.PubMed/NCBI
|
|
13
|
Song R, Gu D, Zhang L, Zhang X, Yu B, Liu
B and Xie J: Functional significance of Hippo/YAP signaling for
drug resistance in colorectal cancer. Mol Carcinog. 57:1608–1615.
2018. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu T, Sun L and Zhu X: Yes-associated
protein enhances proliferation and attenuates sensitivity to
cisplatin in human gastric cancer cells. Biomed Pharmacother.
105:1269–1275. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi J, Li F, Yao X, Mou T, Xu Z, Han Z,
Chen S, Li W, Yu J, Qi X, et al: The HER4-YAP1 axis promotes
trastuzumab resistance in HER2-positive gastric cancer by inducing
epithelial and mesenchymal transition. Oncogene. 37:3022–3038.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu X, Xin Y, Xiao Y and Zhao J:
Overexpression of YAP1 is correlated with progression, metastasis
and poor prognosis in patients with gastric carcinoma. Pathol Oncol
Res. 20:805–811. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yue G, Sun X, Gimenez-Capitan A, Shen J,
Yu L, Teixido C, Guan W, Rosell R, Liu B and Wei J: TAZ is highly
expressed in gastric signet ring cell carcinoma. Biomed Res Int.
2014:3930642014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cruess AF, Zlateva G, Pleil AM and
Wirostko B: Photodynamic therapy with verteporfin in age-related
macular degeneration: A systematic review of efficacy, safety,
treatment modifications and pharmacoeconomic properties. Acta
Ophthalmol. 87:118–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mae Y, Kanda T, Sugihara T, Takata T,
Kinoshita H, Sakaguchi T, Hasegawa T, Tarumoto R, Edano M, Kurumi
H, et al: Verteporfin-photodynamic therapy is effective on gastric
cancer cells. Mol Clin Oncol. 13:102020.PubMed/NCBI
|
|
20
|
Dasari VR, Mazack V, Feng W, Nash J, Carey
DJ and Gogoi R: Verteporfin exhibits YAP-independent
anti-proliferative and cytotoxic effects in endometrial cancer
cells. Oncotarget. 8:28628–28640. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Al-Moujahed A, Brodowska K, Stryjewski TP,
Efstathiou NE, Vasilikos I, Cichy J, Miller JW, Gragoudas E and
Vavvas DG: Verteporfin inhibits growth of human glioma in vitro
without light activation. Sci Rep. 7:76022017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong L, Lin F, Wu W, Liu Y and Huang W:
Verteporfin inhibits YAP-induced bladder cancer cell growth and
invasion via Hippo signaling pathway. Int J Med Sci. 15:645–652.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH image to imageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huggett MT, Jermyn M, Gillams A, Illing R,
Mosse S, Novelli M, Kent E, Bown SG, Hasan T, Pogue BW and Pereira
SP: Phase I/II study of verteporfin photodynamic therapy in locally
advanced pancreatic cancer. Br J Cancer. 110:1698–1704. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brodowska K, Al-Moujahed A, Marmalidou A,
Meyer Zu Horste M, Cichy J, Miller JW, Gragoudas E and Vavvas DG:
The clinically used photosensitizer Verteporfin (VP) inhibits
YAP-TEAD and human retinoblastoma cell growth in vitro without
light activation. Exp Eye Res. 124:67–73. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Feng J, Gou J, Jia J, Yi T, Cui T and Li
Z: Verteporfin, a suppressor of YAP-TEAD complex, presents
promising antitumor properties on ovarian cancer. Onco Targets
Ther. 9:5371–5381. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang C, Zhu X, Feng W, Yu Y, Jeong K, Guo
W, Lu Y and Mills GB: Verteporfin inhibits YAP function through
up-regulating 14-3-3σ sequestering YAP in the cytoplasm. Am J
Cancer Res. 6:27–37. 2015.PubMed/NCBI
|
|
29
|
Kang W, Tong JH, Chan AW, Lee TL, Lung RW,
Leung PP, So KK, Wu K, Fan D, Yu J, et al: Yes-associated protein 1
exhibits oncogenic property in gastric cancer and its nuclear
accumulation associates with poor prognosis. Clin Cancer Res.
17:2130–2139. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li P, Sun D, Li X, He Y, Li W, Zhao J,
Wang Y, Wang H and Xin Y: Elevated expression of Nodal and YAP1 is
associated with poor prognosis of gastric adenocarcinoma. J Cancer
Res Clin Oncol. 142:1765–1773. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hong SA, Son MW, Cho J, Jang SH, Lee HJ,
Lee JH, Cho HD, Oh MH and Lee MS: Low angiomotin-p130 with
concomitant high Yes-associated protein 1 expression is associated
with adverse prognosis of advanced gastric cancer. APMIS.
125:996–1006. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang S, Zhu L, Cao Y, Li L, Xie Y, Deng J
and Xiong J: Significant association of YAP1 and HSPC111 proteins
with poor prognosis in Chinese gastric cancer patients. Oncotarget.
8:80303–80314. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang L, Song X, Li X, Wu C and Jiang J:
Yes-Associated protein 1 as a novel prognostic biomarker for
gastrointestinal cancer: A meta-analysis. Biomed Res Int.
2018:40391732018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Choi W, Kim J, Park J, Lee DH, Hwang D,
Kim JH, Ashktorab H, Smoot D, Kim SY, Choi C, et al: YAP/TAZ
initiates gastric tumorigenesis via upregulation of MYC. Cancer
Res. 78:3306–3320. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Morin-Kensicki EM, Boone BN, Howell M,
Stonebraker JR, Teed J, Alb JG, Magnuson TR, O'Neal W and Milgram
SL: Defects in yolk sac vasculogenesis, chorioallantoic fusion, and
embryonic axis elongation in mice with targeted disruption of
Yap65. Mol Cell Biol. 26:77–87. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tian Y, Kolb R, Hong JH, Carroll J, Li D,
You J, Bronson R, Yaffe MB, Zhou J and Benjamin T: TAZ promotes PC2
degradation through a SCFbeta-Trcp E3 ligase complex. Mol Cell
Biol. 27:6383–6395. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Plouffe SW, Lin KC, Moore JL III, Tan FE,
Ma S, Ye Z, Qiu Y, Ren B and Guan KL: The Hippo pathway effector
proteins YAP and TAZ have both distinct and overlapping functions
in the cell. J Biol Chem. 293:11230–11240. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hayashi H, Higashi T, Yokoyama N, Kaida T,
Sakamoto K, Fukushima Y, Ishimoto T, Kuroki H, Nitta H, Hashimoto
D, et al: An imbalance in TAZ and YAP expression in hepatocellular
carcinoma confers cancer stem cell-like behaviors contributing to
disease progression. Cancer Res. 75:4985–4997. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Finch-Edmondson ML, Strauss RP, Passman
AM, Sudol M, Yeoh GC and Callus BA: TAZ protein accumulation is
negatively regulated by YAP abundance in mammalian cells. J Biol
Chem. 290:27928–27938. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Altieri DC: Survivin, versatile modulation
of cell division and apoptosis in cancer. Oncogene. 22:8581–8589.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tamm I, Wang Y, Sausville E, Scudiero DA,
Vigna N, Oltersdorf T and Reed JC: IAP-family protein survivin
inhibits caspase activity and apoptosis induced by Fas (CD95), Bax,
caspases, and anti-cancer drugs. Cancer Res. 58:5315–5320.
1998.PubMed/NCBI
|
|
42
|
Cerda-Opazo P, Valenzuela-Valderrama M,
Wichmann I, Rodríguez A, Contreras-Reyes D, Fernández EA,
Carrasco-Aviño G, Corvalán AH and Quest AFG: Inverse expression of
survivin and reprimo correlates with poor patient prognosis in
gastric cancer. Oncotarget. 9:12853–12867. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bertazza L, Mocellin S, Marchet A, Pilati
P, Gabrieli J, Scalerta R and Nitti D: Survivin gene levels in the
peripheral blood of patients with gastric cancer independently
predict survival. J Transl Med. 7:1112009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang J, Zhu Z, Sun Z, Sun X, Wang Z and
Xu H: Survivin gene expression increases gastric cancer cell
lymphatic metastasis by up-regulating vascular endothelial growth
factor-C expression levels. Mol Med Rep. 9:600–606. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Da C, Xin Y, Zhao J and Luo X:
Significance and relationship between Yes-associated protein and
survivin expression in gastric carcinoma and precancerous lesions.
World J Gastroenterol. 15:4055–4061. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Maniotis AJ, Folberg R, Hess A, Seftor EA,
Gardner LM, Pe'er J, Trent JM, Meltzer PS and Hendrix MJ: Vascular
channel formation by human melanoma cells in vivo and in vitro:
Vasculogenic mimicry. Am J Pathol. 155:739–752. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang S, Zhang D and Sun B: Vasculogenic
mimicry: Current status and future prospects. Cancer Lett.
254:157–164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun J, Sun B, Sun R, Zhu D, Zhao X, Zhang
Y, Dong X, Che N, Li J, Liu F, et al: HMGA2 promotes vasculogenic
mimicry and tumor aggressiveness by up-regulating Twist1 in gastric
carcinoma. Sci Rep. 7:22292017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bora-Singhal N, Nguyen J, Schaal C,
Perumal D, Singh S, Coppola D and Chellappan S: YAP1 regulates OCT4
activity and SOX2 expression to facilitate self-renewal and
vascular mimicry of stem-like cells. Stem Cells. 33:1705–1718.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wei H, Wang F, Wang Y, Li T, Xiu P, Zhong
J, Sun X and Li J: Verteporfin suppresses cell survival,
angiogenesis and vasculogenic mimicry of pancreatic ductal
adenocarcinoma via disrupting the YAP-TEAD complex. Cancer Sci.
108:478–487. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kim HS, Won YJ, Shim JH, Kim HJ, Kim J,
Hong HN and Kim BS: Morphological characteristics of vasculogenic
mimicry and its correlation with EphA2 expression in gastric
adenocarcinoma. Sci Rep. 9:34142019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kang MH, Jeong GS, Smoot DT, Ashktorab H,
Hwang CM, Kim BS, Kim HS and Park YY: Verteporfin inhibits gastric
cancer cell growth by suppressing adhesion molecule FAT1.
Oncotarget. 8:98887–98897. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Xiong J, Wang S, Chen T, Shu X, Mo X,
Chang G, Chen JJ, Li C, Luo H and Lee JD: Verteporfin blocks
Clusterin which is required for survival of gastric cancer stem
cell by modulating HSP90 function. Int J Biol Sci. 15:312–324.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Giraud J, Molina-Castro S, Seeneevassen L,
Sifré E, Izotte J, Tiffon C, Staedel C, Boeuf H, Fernandez S,
Barthelemy P, et al: Verteporfin targeting YAP1/TAZ-TEAD
transcriptional activity inhibits the tumorigenic properties of
gastric cancer stem cells. Int J Cancer. 146:2255–2267. 2020.
View Article : Google Scholar : PubMed/NCBI
|