Introduction
Gastric cancer is a common malignant tumor of the
digestive system. According to the report released by the World
Health Organization International Agency for Research on Cancer,
there were 9.6 million cancer deaths and 18.1 million new cancer
cases in 2018 worldwide, and gastric cancer ranks third with
respect to mortality and fifth in terms of incidence (1). The incidence and mortality rates of
gastric cancer are also high in China, which are about 2-fold
higher than the global average (1,2). Several
genes, including tumor suppressors and oncogenes, are involved in
the tumorigenesis and progression of gastric cancer.
Overexpression, amplification and rearrangement of oncogenes can
lead to the abnormal activation of cell signaling pathways, promote
cell proliferation and differentiation, inhibit cell apoptosis and
enhance cell invasion and metastasis (3,4).
Stomatin-like protein 2 (SLP-2) is a plasma membrane-associated
protein originally identified in 2000 (5). It belongs to the highly conserved
stomatin protein family and is expressed in several human tissue
types, such as the stomach, lung, intestine, spleen and gallbladder
(5,6). Previous studies have shown that SLP-2
is upregulated and acts as an oncogene in a variety of human cancer
types, including lung, endometrial, breast and colon cancer
(7–9). However, there are few reports on the
expression and the role of SLP-2 in gastric cancer. To the best of
our knowledge, only one study has revealed that SLP-2 was
upregulated in gastric cancer tissues compared with the
adjacent-normal gastric epithelium, which suggested that high-level
SLP-2 expression may contribute to the progression and poor
prognosis of gastric cancer (7).
The aim of the present study was to examine the
expression levels of SLP-2 in gastric cancer cell lines and to
investigate the effect and mechanism of action of SLP-2 on the
apoptosis and autophagy of gastric cancer cells. The findings may
provide insight into the clinical treatment of gastric cancer.
Materials and methods
Cell lines and cell culture
The normal gastric mucosa cell line GES-1 (cat. no.
BNCC353464), gastric adenocarcinoma cell line AGS (cat. no.
BNCC309318) and MKN-45 (cat. no. BNCC337682) and gastric tubular
adenocarcinoma cell line NCI-N87 (cat. no. BNCC312834) were
purchased from BNBIO. NCI-N87, GES-1 and MKN-45 cells were cultured
in RPMI-1640 medium (cat. no. KGM31800S; Nanjing KeyGen Biotech
Co., Ltd.), whereas AGS cells were cultured in F12 medium (cat. no.
KGM21700S; Nanjing KeyGen Biotech Co., Ltd.), containing 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) and 100 U/ml penicillin and
streptomycin. The cells were maintained at 37°C in a humidified
atmosphere with 5% CO2.
Cell transfection
Small interfering RNA (siRNA) against SLP-2
(siRNA-1, −2 and −3) and an siRNA negative control (NC) were
purchased from ZHBY Biotech Co., Ltd and were transfected into
NCI-N87 cells using Lipofectamine™ 3000 transfection reagent (cat.
no. L3000015; Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol. The sequences of all siRNAs used in
the present study are as follows: SLP-2 siRNA-1 sense,
5′-GUGCAGAGUCUCAAGGAAATT-3′ and antisense,
5′-UUUCCUUGAGACUCUGCACTT-3′; SLP-2 siRNA-2 sense,
5′-AGAAAAGGCUGAACAGAUATT-3′ and antisense,
5′-UAUCUGUUCAGCCUUUUCUTT-3′; SLP-2 siRNA-3 sense,
5′-GGCCAAGGCUAAAGCUGAATT-3′ and antisense,
5′-UUCAGCUUUAGCCUUGGCCTT-3′; and siRNA NC sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′ (reverse). Briefly, the cell culture
medium was replaced with serum-free medium before transfection. A
total of 2.5 µg siRNA or 5 µl Lipofectamine® 3000 was
separately diluted in 125 µl of Opti-MEM (Gibco; Thermo Fisher
Scientific, Inc.), then incubated at room temperature for 5 min.
The diluted plasmids were then added to the diluted Lipofectamine
3000 and incubated at room temperature for 15 min. The complex was
added to the cells and incubated at 37°C for 4 h. Subsequently, the
cell culture medium were replaced with fresh pre-warmed medium.
After incubation for 48 h, the cells were harvested for reverse
transcription-quantitative (RT-qPCR) and western blot analysis.
RT-qPCR
Total RNA was extracted from the cells using the
Ultrapure RNA kit (cat. no. CW0581M; CWBio). The concentration and
the purity [optical density (OD)260/OD280] of the RNA were
determined using an NP80 UV–Vis spectrophotometer (NanoPhotometer;
Implen GmbH). The RNA was then reverse-transcribed to cDNA using
the HiScript II Q RT SuperMix for qPCR with gDNA wiper (cat. no.
R223-01; Vazyme Biotech Co., Ltd.). The reverse transcription
reaction was conducted at 50°C for 15 min, and then 85°C for 5 sec.
The qPCR step was performed using 2X SYBR-Green PCR Master Mix
(cat. no. A4004M; Xiamen Life Internet Technology Co., Ltd.) on a
CFX connect™ Real-Time PCR detection system (Bio-Rad Laboratories,
Inc.). The reaction was set up follows: i) RNase-free distilled
H2O, 9.5 µl; ii) cDNA, 1 µl; iii) forward primer, 1 µl;
iv) reverse primer, 1 µl; and v) 2X SYBR-Green PCR Master Mix, 12.5
µl. The primers were obtained from Anhui General Biosystems Co.,
Ltd., and their sequences are shown in Table I. The thermocycling conditions were
included a pre-denaturation at 95°C for 10 min followed by 40
cycles of denaturation at 95°C for 10 sec, annealing at 57–62°C for
30 sec and extension at 72°C for 30 sec. GAPDH was used as the
internal control. The relative expression levels of SLP-2, ANXA2,
β-catenin, Bcl-2 and Beclin-1 were calculated using the 2-ΔΔCq
method (10).
 | Table I.Primer sequences and annealing
temperatures. |
Table I.
Primer sequences and annealing
temperatures.
| Primer name | Primer sequences,
5–3 | Primer length,
bp | Product length,
bp | Annealing
temperature, °C |
|---|
| SLP-2 | F:
GCTGCTGACTGCTGGGGTAT | 20 | 236 | 60.5 |
| SLP-2 | R:
CCTGCTGCCTGATTTATCTGTT | 22 |
|
|
| ANXA2 | F:
ACCTGGAGACGGTGATTTTG | 20 | 263 | 58.1 |
| ANXA2 | R:
CTCTGCTCTTCTACCCTTTGC | 21 |
|
|
| β-catenin | F:
TCTGAGGACAAGCCACAAGAT | 21 | 109 | 60.0 |
| β-catenin | R:
CAAGTCCAAGATCAGCAGTCTCATT | 25 |
|
|
| Bcl-2 | F:
TGAGTTCGGTGGGGTCAT | 18 | 189 | 57.3 |
| Bcl-2 | R:
CAGGAGAAATCAAACAGAGGC | 21 |
|
|
| Bax | F:
GGATGCGTCCACCAAGAA | 18 | 193 | 57.0 |
| Bax | R:
AAAGTAGAAAAGGGCGACAAC | 21 |
|
|
| Beclin-1 | F:
AATAACTTCAGGCTGGGTCG | 20 | 141 | 58.2 |
| Beclin-1 | R:
AGGAACAAGTCGGTATCTCTGAA | 23 |
|
|
| GAPDH | F:
TGACTTCAACAGCGACACCCA | 21 | 121 | 61.5 |
| GAPDH | R:
CACCCTGTTGCTGTAGCCAAA | 21 |
|
|
Western blot analysis
RIPA cell lysis buffer (cat. no. C1053; Applygen
Technologies, Inc.) was used to extract total protein from NCI-N87
cells. Protein concentration was measured using a BCA Assay kit
(cat. no. CW0014S; CWBio). A total of 30 µg protein was loaded per
lane, separated by 10% SDS-PAGE, and then transferred onto
nitrocellulose membranes at 300 mA for 80 min. After blocking with
5% non-fat milk (cat. no. P1622; Applygen Technologies, Inc.) at
4°C overnight, the membranes were incubated at 4°C overnight with
primary antibodies (dilution, 1:500), including rabbit anti-SLP-2
(cat. no. 10348-1-AP; ProteinTech Group, Inc.), rabbit anti-Annexin
A2 (ANXA2; cat. no. 11256-1-AP; ProteinTech Group, Inc.), rabbit
anti-Bax (cat. no. 50599-2-lg; ProteinTech Group, Inc.), rabbit
anti-Beclin-1 (cat. no. 11306-1-AP; ProteinTech Group, Inc.),
rabbit anti-β-catenin (cat. no. ab32572; Abcam), mouse anti-Bcl-2
(cat. no. ab692; Abcam), rabbit anti-LC3-I/LC3-II (cat. no.
bS-8878R; Biosynthesis Biotechnology Inc.) and mouse anti-GAPDH
(cat. no. TA-08; ZSGB-BIO). The membranes were washed, then
incubated with horseradish peroxidase-conjugated secondary
antibodies at 1:1,000 dilution, including goat anti-mouse IgG (H +
L) (cat. no. ZB-2305; ZSGB-BIO) and goat anti-rabbit IgG (H + L)
(cat. no. ZB-2301; ZSGB-BIO) at room temperature for 2 h. The band
signals were developed using ECL solution (Thermo Fisher
Scientific, Inc.). The Chemi Doc™ XRS+ system (Bio-Rad
Laboratories, Inc.) was used for visualization. Protein expression
was quantified by using ImageJ software (version 18.0; National
Institutes of Health).
Cell Counting Kit-8 (CCK-8)
Cells were seeded into 96-well plates
(4×104 cells per well), and cell proliferation was
detected using a CCK-8 kit according to the manufacturer's
instructions. The cells in each well were incubated with 10 µl
CCK-8 reagent at 37°C for 2 h. The OD was detected in each well at
450 nm using a WD-2102B microplate reader (Beijing Liuyi
Biotechnology Co., Ltd.).
Flow cytometry analysis of
apoptosis
Early and late apoptosis was detected using an
Annexin V-FITC/PI Apoptosis Kit (cat. no. AP101-100; Multi Sciences
Lianke Biotech, Co., Ltd.). A total of 1–3×106 cells
were collected and centrifuged with 1 ml PBS (twice at 503 × g, 4°C
for 3 min) The cells were resuspended in 1.5 ml Binding Buffer and
added to three tubes (one for the the blank control, the other two
as single staining with 5 µl Annexin V-FITC or 10 µl PI-PE
solution), and incubated at room temperature for 10 min. The blank
control tube was used to adjust the voltage of FSC, SSC and the
fluorescence channel. Under these voltage conditions, the
single-stained tube was used to adjust the compensation of the
fluorescence channel. After adjusting the parameters, the samples
were resuspended in Binding Buffer, and 5 µl Annexin V-FITC and 10
µl PI-PE solution were added. After incubation at room temperature
for 10 min in the dark, 200 µl ice-cold 1X Binding Buffer was added
and mixed well. Flow cytometric data were acquired using a
NovoCyte™ 2060R flow cytometer (ACEA Biosciences) with NovoExpress
software (version 1.2.5; Agilent Technologies, Inc.).
Transmission electron microscopy of
autophagosomes
The cells were fixed in 2.5% glutaraldehyde at 4°C
overnight, and then washed three times with cold PBS. After fixing
in 1% osmic acid at 4°C for 1 h, and dehydration with gradient
alcohol and acetone, the cells were embedded in epoxy resin
overnight at room temperature, and placed in an oven at 60°C for 2
h. 60-nm sections were cut from the blocks, and then double-stained
with 2% uranyl acetate and lead citrate at room temperature for
5–10 min. The autophagosomes were observed under a transmission
electron microscope (HT7700; Hitachi, Ltd.).
Statistical analysis
GraphPad Prism7 software (GraphPad Software, Inc.)
was used for statistical analysis. The results are presented as the
mean ± standard deviation. Differences between groups were
evaluated using one-way ANOVA followed by the Least Significant
Difference test and Tukey's post hoc tests. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of SLP-2 in gastric cancer
cell lines
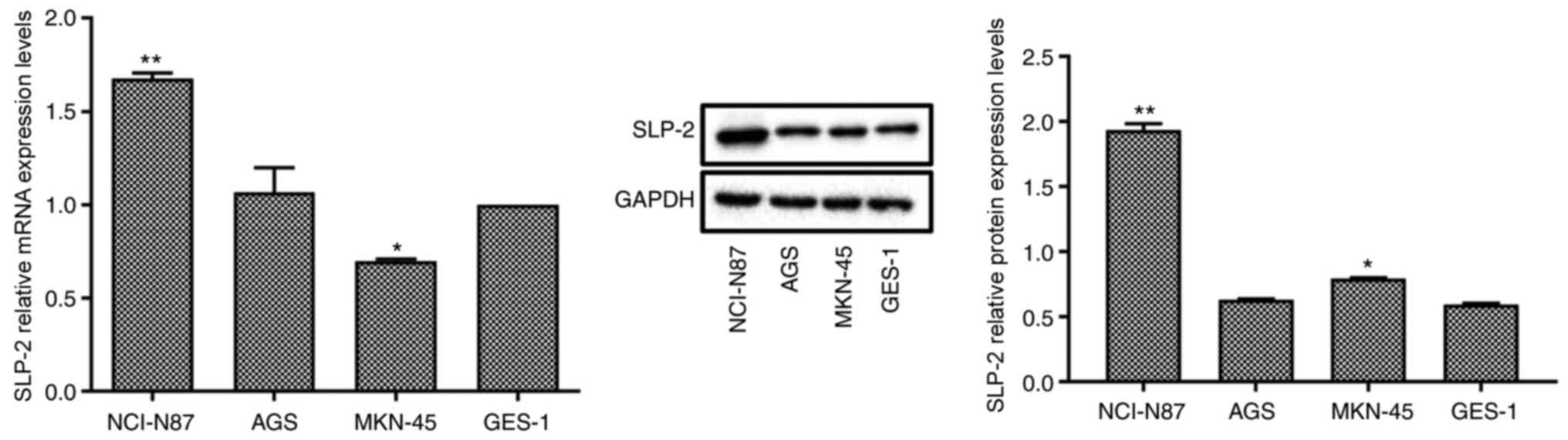
In order to select the gastric cancer cell line with
the highest expression levels of SLP-2, RT-qPCR and western blot
analysis were used to measure SLP-2 expression in different gastric
cancer cell lines (Fig. 1). Compared
with the normal cell line GES-1, the mRNA and protein expression
levels of SLP-2 significantly increased in NCI-N87 cell line, but
did not change significantly in AGS cell line. SLP-2 mRNA
expression level was decreased, but its protein expression level
was increased in the MKN-45 cell line. Therefore, NCI-N87 cell line
was selected for subsequent experiments.
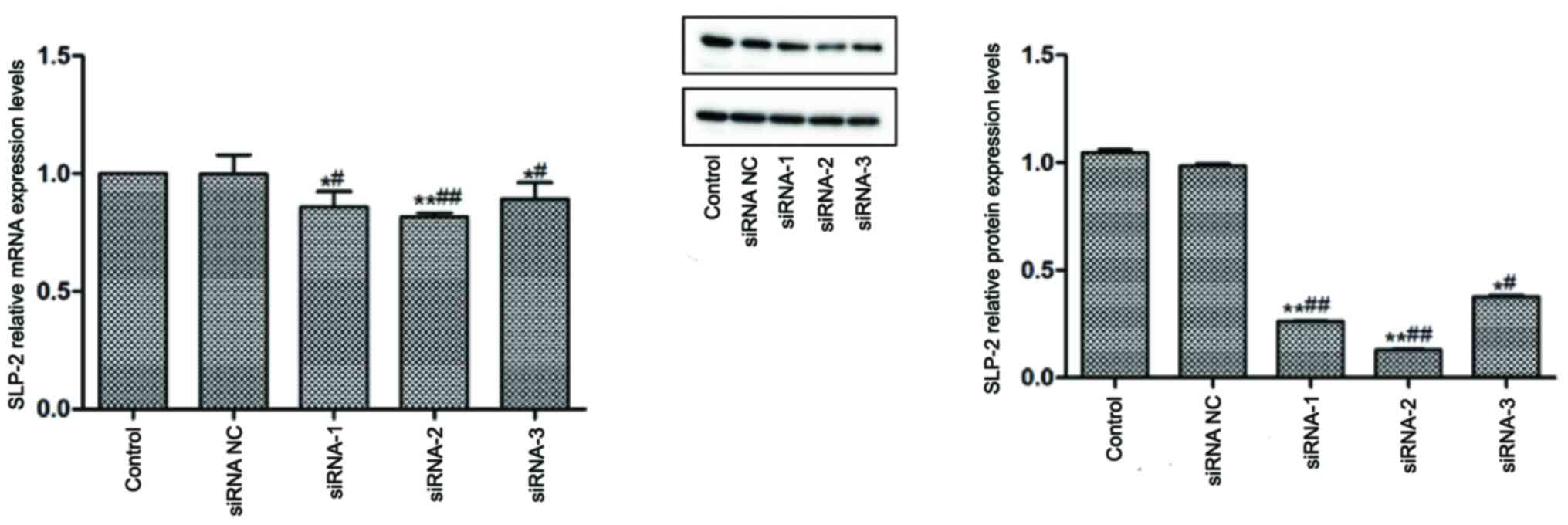
Efficiency of SLP-2 siRNA-mediated
silencing
In order to inhibit SLP-2 expression, three SLP-2
specific siRNA candidates were designed and transfected into
NCI-N87 cells. The silencing efficiency of these siRNA molecules
was detected using RT-qPCR and western blot analysis (Fig. 2). Compared with the control and siRNA
NC groups, siRNA-1, siRNA-2 and siRNA-3 significantly decreased the
mRNA and protein expression levels of SLP-2 in NCI-N87 cells.
siRNA-2 was chosen for subsequent, as it displayed the highest
efficiency of SLP-2 silencing.
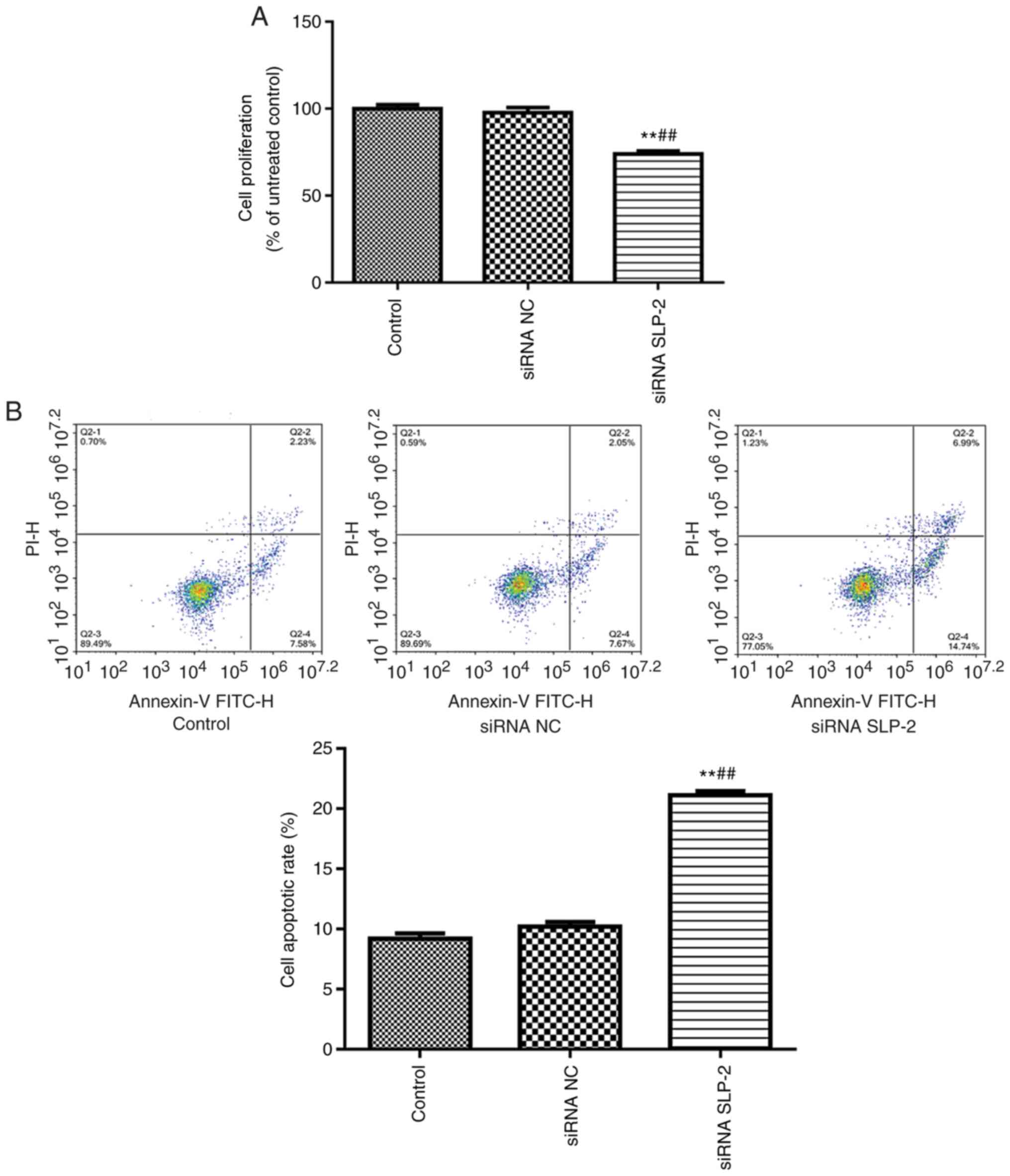
Effect of siRNA SLP-2 on the
proliferation and apoptosis of NCI-N87 cells
In order to examine the effect of siRNA SLP-2 on the
proliferation of NCI-N87 cells, CCK-8 was used to detect cell
proliferation (Fig. 3A). Compared
with the control and siRNA NC groups, siRNA SLP-2 significantly
inhibited the proliferation of NCI-N87 cells. In addition, the rate
of early and late apoptosis was evaluated using flow cytometry
(Fig. 3B). Compared with the control
and siRNA NC groups, the apoptotic rate significantly increased in
the siRNA SLP-2 group.
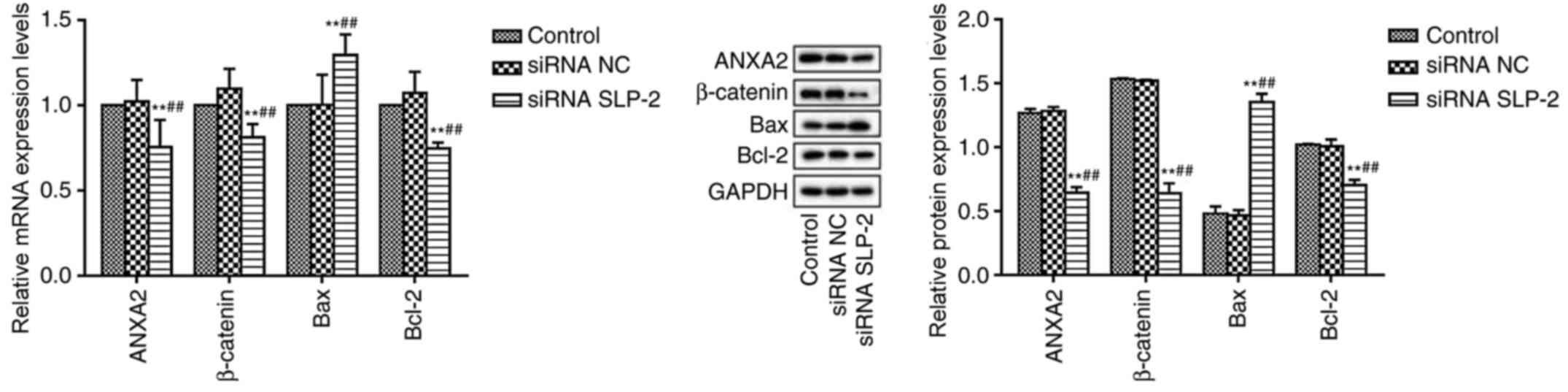
Effect of siRNA SLP-2 on the
expression of ANXA2, β-catenin, Bcl-2 and Bax in NCI-N87 cells
RT-qPCR and western blot analysis were used to
detect the expression levels of ANXA2, β-catenin, Bcl-2 and Bax in
NCI-N87 cells (Fig. 4). Compared
with the control and siRNA NC groups, the mRNA and protein
expression levels of ANXA2, β-catenin and Bcl-2 were significantly
reduced in the siRNA SLP-2 group, whereas the expression of Bax was
significantly increased.
Effect of siRNA SLP-2 on autophagy of
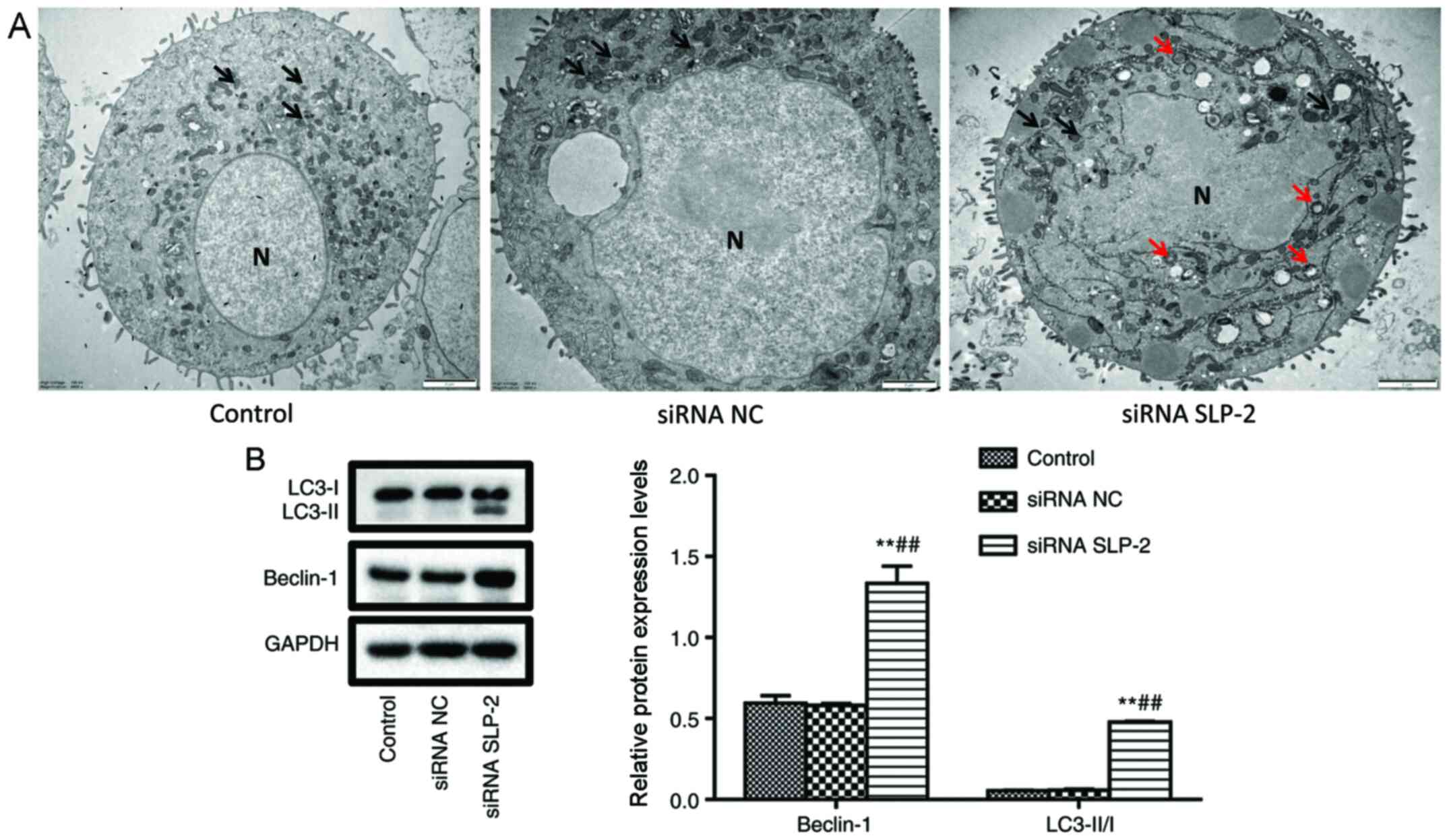
NCI-N87 cells
To investigate the effect of siRNA SLP-2 on the
autophagy of NCI-N87 cells, autophagosomes were observed by
transmission electron microscopy, and the expression of Beclin-1
and LC3-II/I was detected by western blot analysis. As shown in
Fig. 5A, compared with the control
and siRNA NC groups, the mitochondria were severely damaged, and a
large number of autophagosomes were observed in the siRNA SLP-2
group. Furthermore, the protein levels of Beclin-1 and LC3-II/I in
the siRNA SLP-2 group were significantly increased (Fig. 5B). These results indicated that siRNA
SLP-2 can induce autophagy in NCI-N87 cells.
Discussion
In the present study, the expression of SLP-2 was
examined in human gastric cancer cell lines (AGS, MKN-45 and
NCI-N87) and a human gastric epithelial cell line (GES-1). The AGS
cell line was originally derived from the resected tumor of a
patient with gastric adenocarcinoma (11). The NCI-N87 cell line was derived from
the metastatic site of a patient with gastric carcinoma (11). In the present study, SLP-2 expression
level was higher in a cell line originating from distant metastases
(NCI-N87) than in the cell line derived from a primary tumor (AGS),
indicating that SLP-2 exhibits differential expression in different
stages of gastric cancer, and that SLP-2 may be associated with the
pathogenesis of gastric cancer. Since the NCI-N87 cell line
expressed the highest levels of SLP-2 among all the tested cell
lines, it was used for subsequent loss-of-function experiments.
SLP-2 is a membrane-associated protein first
isolated from human erythrocytes by Wang and Morrow in 2000
(5). Later, Wang et al
(12) demonstrated that SLP-2 was
also a mitochondrial protein that played important roles in
maintaining calcium homeostasis (12). A number of studies have demonstrated
that SLP-2 is highly expressed in tumor tissue and is closely
related to the occurrence and development of tumors (7–9). For
instance, Zhang et al (13)
reported that SLP-2 was overexpressed in ovarian cancer tissue and
that its expression correlated with clinical stage, pathological
differentiation and lymph node metastasis of patients with ovarian
cancer, suggesting that SLP-2 may play a role in promoting the
invasion and metastasis of ovarian cancer. Moreover, Zhou et
al suggested that SLP-2 was highly expressed in colorectal
cancer tissues and cell lines and that positive expression of SLP-2
could be used as an independent risk factor for poor prognosis of
colorectal cancer (14). Silencing
of the SLP-2 gene significantly inhibits the proliferation,
migration and invasion of colorectal cancer cells (14). Liu et al detected the
expression of SLP-2 in primary gastric cancer tissue and matched
normal gastric tissue samples using RT-qPCR and western blot
analysis, which indicated that SLP-2 was highly expressed in
gastric cancer tissues (7).
Furthermore, they observed that high expression of SLP-2 was
closely related to the depth of invasion, lymph node metastasis,
distant metastasis and American Joint Committee on Cancer stage.
Cox regression analysis also indicated that the expression of SLP-2
was an independent prognostic factor for gastric cancer (7). These findings suggest that SLP-2 may be
an oncogene involved in the development of gastric cancer.
Apoptosis is an active, sequential and programmed
cell death process (15). Autophagy
is a survival process in which eukaryotic cells degrade damaged or
senescent organelles and proteins for cellular renewal (15). Crosstalk can occur between autophagy
and apoptosis (16–18). The activation of autophagy usually
protects cells from apoptosis (15,19). In
some circumstances, however, autophagy may promote cellular demise
by inducing apoptosis (15,19). Thus, the complex interplay between
autophagy and apoptosis determines the fate of cells. The BCL-2
protein family is a central regulator of autophagy and apoptosis
(20). The anti-apoptotic protein
Bcl-2 can form a heterodimer with the pro-apoptotic protein Bax,
resulting in the enhancement of the anti-apoptotic effect of Bcl-2
(21). Bax is also able to form
homodimers to promote apoptosis (21). The autophagy-related protein Beclin-1
can competitively bind with Bcl-2/Bcl-xl to promote autophagy
(21). The inhibition of apoptosis
and the induction of autophagy have been documented in several
types of tumor, including gastric cancer (15). In recent years, the development of
autophagy inhibitors, as well as inducers of tumor cell apoptosis
to a certain extent, has attracted increasing attention (16,22–24).
In the present study, CCK-8 and flow cytometry were
used to examine the effect of SLP-2 on the proliferation and
apoptosis of NCI-N87 cells. The results demonstrated that
downregulation of SLP-2 could significantly inhibit proliferation
and induce apoptosis of NCI-N87 cells. In addition, RT-qPCR and
western blot results indicated that SLP-2 silencing could
significantly downregulate Bcl-2 and upregulate Bax in these cells.
This finding is consistent with a previous study suggesting that
downregulation of SLP-2 can inhibit the proliferation and induce
the apoptosis of glioma cells by regulating the expression of Bcl-2
and Bax (25). To the best of our
knowledge, the role of SLP-2 on autophagy has not been reported to
date. Thus, this study is the first to reveal the effect of SLP-2
on autophagy of gastric cancer cells. Moreover, the present results
showed a large number of autophagosomes in siRNA SLP-2-transfected
cells, and siRNA SLP-2 upregulated Beclin-1 and LC3-I/II,
suggesting that the downregulation of SLP-2 may induce apoptosis of
NCI-N87 cells by promoting autophagy.
Apoptosis and autophagy are mediated by multiple
signaling pathways, among which mTOR signaling pathway is the most
important (26,27). It has been reported that ANXA2 can
regulate autophagy in Pseudomonas aeruginosa infection
through the AKT1-mTOR-ULK1/2 signaling pathway (28). ANXA2 is a multifunctional
calcium-dependent phospholipid-binding protein that is widely
distributed in the membrane, cytoplasm and nucleus of eukaryotic
cells (29). ANXA2 plays important
roles in cytoskeleton remodeling, cell phenotype change and cell
movement by regulating the expression of connexin molecules
(30–32). The abnormal expression of ANXA2 is
closely associated type 2 diabetes, cancer and autoimmune diseases
(33–35). Yang et al (36) analyzed protein-protein interactions
in SLP-2-overexpressing A549 cells using immunoprecipitation and
proteomics and determined that ANXA2 interacted with SLP-2 and
β-catenin directly. Additional experiments demonstrated that the
knockout of the SLP-2 gene inhibited SLP-2/ANXA2/β-catenin cascade
formation, reduced the translocation of cytoplasmic β-catenin into
the nucleus and suppressed the expression of the downstream target
gene, survivin, thereby inhibiting the proliferation of non-small
cell lung cancer cells (36). Zhou
et al demonstrated that the cell viability, migration and
invasion of SW620 significantly decreased following SLP-2 gene
silencing (14). In addition,
silencing of the SLP-2 gene in SW620 cells inhibited the expression
of metastasis-associated genes and the activation of Wnt/β-catenin
signaling pathway (14). In the
present study, the expression levels of ANXA2 and β-catenin were
determined in NCI-N87 cells. The results demonstrated that SLP-2
silencing could significantly downregulate the expression of ANXA2
and β-catenin. This finding is consistent with a previous report in
NSCLC cells (36) and suggests that
SLP-2 may affect the apoptosis and autophagy of gastric cancer
cells by regulating ANXA2/β-catenin signaling.
A limitation of this study is that the regulatory
effect of SLP-2 on the ANXA2/β-catenin signaling pathway was only
determined in NCI-N87 cells. In addition, ANXA2/β-catenin signaling
inhibitors could be used in future studies to confirm the
hypothesis that this signaling pathway mediates the effect of SLP-2
on gastric cancer cell apoptosis and autophagy.
In conclusion, SLP-2 silencing significantly induced
apoptosis and autophagy, as well as inhibited the proliferation of
NCI-N87 cells, and this effect may be mediated by inhibition of
ANXA2/β-catenin signaling. These findings revealed the effect and
the mechanism of action of SLP-2 in gastric cancer cells and
provided new potential avenues for the clinical treatment of
gastric cancer.
Acknowledgements
Not applicable.
Funding
This study was supported by The National Key Project
of Precision Medicine Research (grant no. 2017YFC0908300) and The
Qinghai Provincial Scientific Research Project (grant no.
2019-SF-L3).
Availability of data and materials
The data sets generated and analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
XW and SY designed the study and prepared the
manuscript. SY, YH, HZ, FW and LS conducted the experiments and
analyzed the data. All authors were substantially involved in the
research, acquisition of data, analysis and manuscript preparation.
All authors read and approved the final manuscript. XW and SY
confirmed the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li Z, Ying X, Shan F and Ji J: The
association of garlic with Helicobacter pylori infection and
gastric cancer risk: A systematic review and meta-analysis.
Helicobacter. 23:e125322018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mao QQ, Xu XY, Shang A, Gan RY, Wu DT,
Atanasov AG and Li HB: Phytochemicals for the prevention and
treatment of gastric cancer: Effects and Mechanisms. Int J Mol Sci.
21:212020. View Article : Google Scholar
|
|
4
|
Xiao S and Zhou L: Gastric cancer:
Metabolic and metabolomics perspectives (Review). Int J Oncol.
51:5–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y and Morrow JS: Identification and
characterization of human SLP-2, a novel homologue of stomatin
(band 7.2b) present in erythrocytes and other tissues. J Biol Chem.
275:8062–8071. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Deng H, Deng Y, Liu F, Chen J, Li Z, Zhao
K, Guan X and Liang W: Stomatin-like protein 2 is overexpressed in
cervical cancer and involved in tumor cell apoptosis. Oncol Lett.
14:6355–6364. 2017.PubMed/NCBI
|
|
7
|
Liu D, Zhang L, Shen Z, Tan F, Hu Y, Yu J
and Li G: Increased levels of SLP-2 correlate with poor prognosis
in gastric cancer. Gastric Cancer. 16:498–504. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang X, Hu Y, Shi H, Zhang C, Wang Z, Liu
X, Chen H, Zhang L and Cui D: The diagnostic value of TROP-2, SLP-2
and CD56 expression in papillary thyroid carcinoma. Eur Arch
Otorhinolaryngol. 275:2127–2134. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu Q, Li A, Wang L, He W, Zhao L, Wu C,
Lu S, Ye X, Zhao H, Shen X, et al: Stomatin-like protein 2 promotes
tumor cell survival by activating the JAK2-STAT3-PIM1 pathway,
suggesting a novel therapy in CRC. Mol Ther Oncolytics. 17:169–179.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Burgermeister E, Xing X, Röcken C, Juhasz
M, Chen J, Hiber M, Mair K, Shatz M, Liscovitch M, Schmid RM, et
al: Differential expression and function of caveolin-1 in human
gastric cancer progression. Cancer Res. 67:8519–8526. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Y, Cao W, Yu Z and Liu Z:
Downregulation of a mitochondria associated protein SLP-2 inhibits
tumor cell motility, proliferation and enhances cell sensitivity to
chemotherapeutic reagents. Cancer Biol Ther. 8:1651–1658. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Song X, Li C and Tian Y:
Expression and clinical significance of SLP-2 in ovarian tumors.
Oncol Lett. 17:4626–4632. 2019.PubMed/NCBI
|
|
14
|
Zhou C, Li Y, Wang G, Niu W, Zhang J, Wang
G, Zhao Q and Fan L: Enhanced SLP-2 promotes invasion and
metastasis by regulating Wnt/β-catenin signal pathway in colorectal
cancer and predicts poor prognosis. Pathol Res Pract. 215:57–67.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu JZ, Hu YL, Feng Y, Guo YB, Liu YF,
Yang JL, Mao QS and Xue WJ: Rafoxanide promotes apoptosis and
autophagy of gastric cancer cells by suppressing PI3K/Akt/mTOR
pathway. Exp Cell Res. 385:1116912019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang G, Zhang T, Sun W, Wang H, Yin F,
Wang Z, Zuo D, Sun M, Zhou Z, Lin B, et al: Arsenic sulfide induces
apoptosis and autophagy through the activation of ROS/JNK and
suppression of Akt/mTOR signaling pathways in osteosarcoma. Free
Radic Biol Med. 106:24–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nowak KL and Edelstein CL: Apoptosis and
autophagy in polycystic kidney disease (PKD). Cell Signal.
68:1095182020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kasprowska-Liśkiewicz D: The cell on the
edge of life and death: Crosstalk between autophagy and apoptosis.
Postepy Hig Med Dosw. 71:825–841. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
D'Arcy MS: Cell death: A review of the
major forms of apoptosis, necrosis and autophagy. Cell Biol Int.
43:582–592. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Edlich F: BCL-2 proteins and apoptosis:
Recent insights and unknowns. Biochem Biophys Res Commun.
500:26–34. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang Q, Bu W, Wang D, Xin Y, Xu X and Sun
H: Advance research on interaction between autophagy and apoptosis
and its influence in development of tumors. J Jilin Univ.
41:1303–1306. 2015.
|
|
22
|
Cao Y, Luo Y, Zou J, Ouyang J, Cai Z, Zeng
X, Ling H and Zeng T: Autophagy and its role in gastric cancer.
Clin Chim Acta. 489:10–20. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qian HR and Yang Y: Functional role of
autophagy in gastric cancer. Oncotarget. 7:17641–17651. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song J, Zhou Y, Gong Y, Liu H and Tang L:
Rottlerin promotes autophagy and apoptosis in gastric cancer cell
lines. Mol Med Rep. 18:2905–2913. 2018.PubMed/NCBI
|
|
25
|
Chen X, Wang P, Xie M, Zhao H and Wu H:
Study on mechanism of down-regulation of SLP-2 gene expression on
proliferation and apoptosis of glioma cells. Chin J Immunol.
34:55–59. 2018.(In Chinese).
|
|
26
|
Yu Z, Chen Y, Liang C and Eriocalyxin B:
Eriocalyxin B Induces apoptosis and autophagy involving
Akt/mammalian target of rapamycin (mTOR) pathway in prostate cancer
cells. Med Sci Monit. 25:8534–8543. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Wang C, Liu Y, You J and Su G:
Autophagy, lysosome dysfunction and mTOR inhibition in MNU-induced
photoreceptor cell damage. Tissue Cell. 61:98–108. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li R, Tan S, Yu M, Jundt MC, Zhang S and
Wu M: Annexin A2 regulates autophagy in Pseudomonas
aeruginosa infection through the Akt1-mTOR-ULK1/2 signaling
pathway. J Immunol. 195:3901–3911. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Karimi-Busheri F, Marcoux Y, Tredget EE,
Li L, Zheng J, Ghoreishi M, Weinfeld M and Ghahary A: Expression of
a releasable form of annexin II by human keratinocytes. J Cell
Biochem. 86:737–747. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shi H, Xiao L, Duan W, He H, Ma L, Da M,
Duan Y, Wang Q, Wu H, Song X, et al: ANXA2 enhances the progression
of hepatocellular carcinoma via remodeling the cell motility
associated structures. Micron. 85:26–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He H, Xiao L, Cheng S, Yang Q, Li J, Hou
Y, Song F, Su X, Jin H, Liu Z, et al: Annexin A2 enhances the
progression of colorectal cancer and hepatocarcinoma via
cytoskeleton structural rearrangements. Microsc Microanal.
25:950–960. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen CY, Lin YS, Chen CH and Chen YJ:
Annexin A2-mediated cancer progression and therapeutic resistance
in nasopharyngeal carcinoma. J Biomed Sci. 25:302018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sharma MC: Annexin A2 (ANX A2): An
emerging biomarker and potential therapeutic target for aggressive
cancers. Int J Cancer. 144:2074–2081. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang T, Wang Z, Niu R and Wang L: Crucial
role of Anxa2 in cancer progression: Highlights on its novel
regulatory mechanism. Cancer Biol Med. 16:671–687. 2019.PubMed/NCBI
|
|
35
|
Cardoso CM, de Jesus SF, de Souza MG,
Queiroz LD, Santos EM, Dos Santos EP, Oliveira LP, Santos CK, Sousa
Santos SH, et al: High levels of ANXA2 are characteristic of
malignant salivary gland tumors. J Oral Pathol Med. 48:929–934.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang CT, Li JM, Li LF, Ko YS and Chen JT:
Stomatin-like protein 2 regulates survivin expression in non-small
cell lung cancer cells through β-catenin signaling pathway. Cell
Death Dis. 9:4252018. View Article : Google Scholar : PubMed/NCBI
|