Introduction
Hepatocellular carcinoma (HCC) is the fifth most
frequently diagnosed malignant cancer worldwide, with high
mortality rates (1). China accounts
for ~55% of all global incidences of primary liver cancer, and has
the third highest incidence rate of HCC worldwide, which has been
attributed to high infection rates of hepatitis B virus (2). Radical tumor resection and liver
transplantation are the most effective treatment options. However,
owing to the difficulty of early detection, patients with HCC often
lose the opportunity for surgical treatment due to local invasion
and distant metastasis (3).
Therefore, it is necessary to identify highly sensitive and
specific diagnostic biomarkers to aid the early detection and
prognostic prediction of patients with HCC.
Microfibril-associated protein 2 (MFAP2), also known
as microfibril-associated glycoprotein (MAGP) 1, interacts with
microfibrillin to regulate microfibril functioning, and is the most
widely distributed MAGP in most vertebrate proteins and microfiber
components (4,5). Previous studies have shown that the
gene that codes MFAP2 is located at 1p36.13, and is associated with
several malignant tumor types (6,7).
Consequently, MFAP2 is upregulated in breast cancer (8), thyroid cancer (9), melanoma (10), ovarian cancer (11) and gastric cancer (12,13), and
associated with the occurrence and progression of these tumors.
Previous studies have also shown that MFAP2 is upregulated in HCC
(14), but its role in the
occurrence and development of the disease is unclear. In order to
investigate the role of MFAP2 in the occurrence and development of
HCC, the aim of the present study was to evaluate the expression of
MFAP2 in liver cancer by analyzing the tumor specimens and detailed
clinical data of 94 patients. Based on the results from these
clinical data, the mechanism underlying the effects of MFAP2 in HCC
was also investigated.
Materials and methods
Datasets and prediction
Prediction of MFAP2 expression was performed in HCC
cases using datasets from The Cancer Genome Atlas (TCGA; http://portal.gdc.cancer.gov/projects/TCGA-LIHC).
The TCGA dataset (LICH), downloaded for differential gene
expression analysis in HCC, comprised 50 and 369 normal individuals
and patients with HCC, respectively).
Patient enrollment and follow-up
Paraffin-embedded samples were obtained from total
94 patients with HCC, who underwent tumor resection at the General
Surgery Department of the Anhui Provincial Hospital (Hefei, China)
between 2010 and 2019, were enrolled in the current study. All
patients were neither exposed to chemotherapy nor radiotherapy for
anticancer treatment before surgery. Clinical information,
including age, sex, tumor size and tumor node metastasis (TNM)
status were recorded. Patients were most often followed up for 3
years (range, 0.1–3 years). The present study also included the
fresh tissues of 21 patients who underwent tumor resection at the
hospital between January 2019 and January 2020. The present study
was performed in accordance with the Declaration of Helsinki, and
was approved by the Ethics Committee of Anhui Medical University.
The use of patient tissue samples and clinical information was
approved by the patient, and written informed consent was
obtained.
Immunohistochemistry
Tissues were fixed in 10% formalin for 9 h at room
temperature, embedded in paraffin, and cut into 4-µm thick sections
using a microtome. The sections were heated in a 60°C oven for 2 h
to soften the paraffin, and immediately transferred to xylene for
dewaxing. The samples were rehydrated in descending concentrations
of ethanol (100, 95 and 50%) and then washed with PBS. Antigen
retrieval was performed using sodium citrate solution in a
microwave oven, after which the sections were cooled, and then
exposed to 3% hydrogen peroxide to quench endogenous peroxidase
activity. The sections were washed three times with PBS, and then
incubated overnight with primary antibodies (Table I) in a humid chamber at 4°C.
Thereafter, slides containing tumor sections were washed 3 times
with PBS (for 5 min each), and incubated with secondary antibody
(Table I) at 37°C for 10 min.
Finally, the sections were incubated with 3-3 diaminobenzidine
(Beijing Solarbio Science & Technology Co., Ltd.), and the
nuclei counterstained with hematoxylin. Then the samples were
dehydrated with a gradient of alcohol (50, 95 and 100%) at room
temperature, cleared with xylene, sealed with neutral gum, dried
and observed using a light microscope (Nikon Corporation).
 | Table I.Antibodies used for western blotting
and immunohistochemistry. |
Table I.
Antibodies used for western blotting
and immunohistochemistry.
|
|
|
| Dilution |
|---|
|
|
|
|
|
|---|
| Antibody | Supplier | Cat. no. | Western blotting |
Immunohistochemistry |
|---|
| Rabbit
anti-MFAP2 | ProteinTech Group,
Inc. | AG24496 | 1:1,000 | 1:200 |
| Rabbit
anti-E-cadherin | ProteinTech Group,
Inc. | 20874-1-AP | 1:1,000 | 1:300 |
| Rabbit
anti-N-cadherin | ProteinTech Group,
Inc. | 22018-1-AP | 1:1,000 | 1:100 |
| Rabbit
anti-vimentin | ProteinTech Group,
Inc. | 10366-1-AP | 1:1,000 | 1:200 |
| Rabbit
anti-VEGFA | ProteinTech Group,
Inc. | 66828-1-Ig | 1:1,000 | – |
| Mouse anti-rabbit
IgG | OriGene
Technologies, Inc. | ZB-2301 | 1:5,000 | – |
Immunohistochemical results were interpreted using a
semi-quantitative evaluation system (15,16).
Briefly, staining intensity was scored as 0=absent, 1=weak,
2=medium, and 3=strong, while the percentage of the stained area
was scored as <10=1, 10–50=2, and >50%=3. A total score of
<3 was considered low expression, and ≥3 was regarded as high
expression. All sections were scored in 5 fields under a high-power
lens field, and an average score calculated. All sections were
independently evaluated by two pathologists.
Western blot analysis
RIPA cell/tissue lysis buffer (Beyotime Institute of
Biotechnology), containing a protease inhibitor mixture, was used
to lyse the HCC and adjacent tissues. The contents were centrifuged
at 13,000 × g for 15 min, and the protein-rich supernatant was
collected. Protein concentration was determined using the BCA
method (Beyotime Institute of Biotechnology); ~40 µg/lane protein
was separated by 10% SDS-PAGE, and then transferred to a
polyvinylidene fluoride membrane using the wet transfer method. The
membranes were blocked with 5% skimmed milk antigen, at room
temperature for 1 h, and then incubated overnight with primary
antibodies (Table I) at 4°C. The
membranes were washed 3 times with Tris buffered saline/Tween
(0.1%) and then incubated at room temperature with secondary
antibody (Table I) for 1.5 h.
Finally, an enhanced chemiluminescent reagent (Beyotime Institute
of Biotechnology) was used to visualize the protein bands, which
were quantified using Image Lab v3.1 software (Bio-Rad
Laboratories, Inc.).
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was isolated from HCC tissues and MHCC97H
cells using the RNAeasy™ Plus kit (Beyotime Institute of
Biotechnology). A total of 1 µg RNA was then converted to
complementary DNA using the SuperScript IV reverse transcription
kit (Thermo Fisher Scientific, Inc.) according to manufacturer's
instructions. Levels of MFAP2 expression were determined via qPCR
using the SYBR-Green ProFlex PCR system (Thermo Fisher Scientific,
Inc.); the following thermocycling conditions were used: Initial
denaturation at 95°C for 5 min, followed by 40 cycles of 95°C for
30 sec, 60°C for 30 sec and 72°C for 30 sec. The corresponding
primers are presented in Table II.
Relative gene expression was determined using the 2−ΔΔCq
method (17), relative to that of
GAPDH.
| Table II.Quantitative PCR primer and siRNA
sequences. |
Table II.
Quantitative PCR primer and siRNA
sequences.
| Primer/siRNA | Sequence |
|---|
| MFAP2 | Forward:
5′-TCCGCCGTGTGTACGTCATT-3′ |
|
| Reverse:
5′-CTGGCCATCACGCCACATTT-3′ |
| GAPDH | Forward:
5′-TTGGTATCGTGGAAGGACTCA-3′ |
|
| Reverse:
5′-TGTCATCATATTTGGCAGGTT-3′ |
| MFAP2-siRNA | Sense:
5′-CCCACUAUAGCGACCAGAUTT-3′ |
|
| Antisense:
5′-AUCUGGUCGCUAUAGUGGGTT-3′ |
| NC-siRNA | Sense:
5′-CCAUACACAGGCCUUGCAATT-3′ |
|
| Antisense:
5′-UUGCAAGGCCUGUGUAUGGTT-3′ |
Cell lines and transfection
The human HCC MHCC97H cell line and human umbilical
vein endothelial cells (HUVECs) were acquired from the General
Surgery Laboratory of Anhui Provincial Hospital. The MHCC97H cell
line was cultured using RPMI-1640 medium containing 10%, fetal
bovine serum (FBS) and penicillin/streptomycin (100 U/ml) (all
Gibco; Thermo Fisher Scientific, Inc.). HUVECs were cultured using
endothelial cell medium (ECM; ScienCell Research Laboratories,
Inc.). Both cell lines were maintained at 37°C with 5%
CO2 in a humidified incubator.
According to the manufacturer's protocol, on
reaching 60% confluency, Lipofectamine® 2000 (Gibco;
Thermo Fisher Scientific, Inc.) was used to transfect the cells
with MFAP2 small interfering RNA (siMFAP2 and MFAP2-siRNA) and
negative control (NC)-siRNA (150 pmol/well), Shanghai Biosun
Sci&Tech Co; Table II). The
cells were then cultured at 37°C in a low-serum medium (Opti-MEM;
Gibco; Thermo Fisher Scientific, Inc.) for 6 h. The medium was then
replaced with complete medium containing 10% FBS, and the cells
were incubated for a further 6 h prior to subsequent
experimentation.
Cell counting kit 8 (CCK-8) assay
The CCK-8 kit (Dojindo Molecular Technologies, Inc.)
was used to assess the proliferative capacity of MHCC97H cells
following MFAP2-knockdown. A 96-well plate was seeded with
3×103 cells/well, and 10 µl CCK-8 reagent was added at
24, 48 and 72 h. After incubating for a further hour, the OD value
of each well was measured at 450 nm.
Transwell assay
A total of 1×105 MHCC97H cells were
seeded into the upper chamber of a Transwell insert (pore size,
8-µm) containing Matrigel (Costar; Corning, Inc.), with 200 µl
serum-free medium; 500 µl complete medium was added to the lower
chamber, and the cells were cultured at 37°C for 24 h. The
migratory cells were fixed with 4% formaldehyde at room temperature
for 20 min, and then stained with 0.2% crystal violet at room
temperature for 20 min. The cells were counted in five random
microscopic fields using a light microscope (magnification,
×200).
Wound-healing assay
A total of 3×105 MHCC97H cells per well
were seeded into a 6-well plate and cultured for 24 h at 37°C. The
cells were then cultured in serum-free basal medium for 6 h, and a
1,000-µl pipette tip was used to mark out the cell-free area, prior
to continued culture for an additional 24 h. Images were captured
under a light microscope (Nikon Corporation; magnification, ×100).
The area of the cell-free zone was recorded, and the wound area was
determined using the following formula: (Initial wound area-final
wound area)/initial wound area × 100%.
Tube-formation experiment
Pre-chilled Matrigel (100 µl/well) was added to a
24-well plate and incubated at 37°C for 1 h. Then, 1×104
HUVECs were seeded into each well with 200 µl ECM medium, as well
as 200 µl supernatant from MHCC97H cells cultured in serum-free
medium (obtained by centrifugation at 12,000 × g for 3 min at 4°C).
After 7 h, images were captured using a light microscope (Nikon
Corporation; magnification, ×100), and analyzed using ImageJ
software (V.1.8.0; National Institutes of health).
Statistical analysis
The χ2 test was used to compare
immunohistochemistry scores, and χ2 test or Fisher's
exact test were used to determine the association between
clinicopathological characteristics and MFAP2 expression. One-way
analysis of variance and Tukey's post-hoc test were used for
multiple comparisons of means, and a two-tailed unpaired Student's
t test was used to compare the differences between two groups. For
data conforming to normal distribution and homogeneity of variance,
the differences between two paired groups (normal vs. tumor
tissues) were assessed using paired Student's t-test. Spearman's
rank correlation was used to evaluate the association between the
expression profiles of MFAP2 and epithelial-mesenchymal transition
(EMT)-related proteins. Kaplan-Meier survival curves were used to
analyze the disease-free survival (DFS) and overall survival (OS)
of patients with HCC; differences were assessed using the log-rank
test. Cox proportional hazard regression models were used to obtain
unadjusted and the adjusted hazard ratio (HR) and 95% confidence
interval (CI) values for survival in univariate and multivariate
analysis. All statistical analyses were performed using SPSS 22
(IBM Corp) and GraphPad Prism 14 (GraphPad Software, Inc.). Data
are expressed as the mean ± SD, and P<0.05 was considered to
indicate a statistically significant difference.
Results
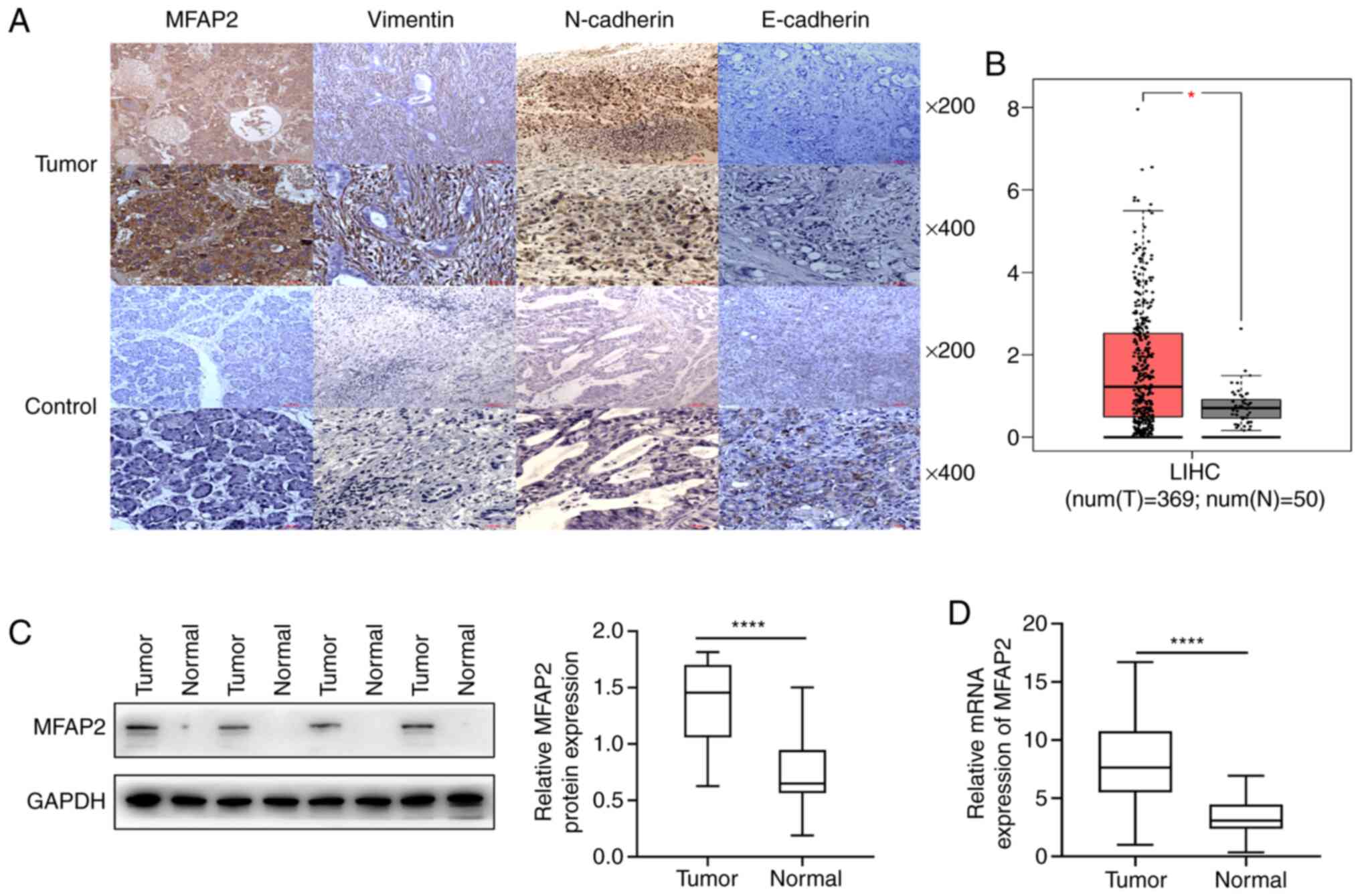
MFAP2 is upregulated in HCC
Immunohistochemistry results revealed that MFAP2 was
significantly upregulated in 76.6% of HCC tissues (72 of 94;
Fig. 1A and Table III). The expression of MFAP2 in HCC
tissues from TCGA was also higher than that in normal tissues
(P<0.05; Fig. 1B). A comparison
of patterns of MFAP2 expression in 21 pairs of fresh carcinoma and
liver tissues revealed a significant upregulation in liver cancer,
relative to adjacent normal tissues (P<0.001; Fig. 1C). RT-qPCR corroborated the western
blots findings, as evidenced by a significant upregulation in MFAP2
mRNA expression in fresh HCC compared with normal tissues
(P<0.001; Fig. 1D). These results
indicate that MFAP2 is upregulated in HCC.
| Table III.Differential expression of MFAP2 in
HCC tissues and corresponding paracarcinoma tissues (n=94). |
Table III.
Differential expression of MFAP2 in
HCC tissues and corresponding paracarcinoma tissues (n=94).
|
| MFAP2
expression |
|
|---|
|
|
|
|
|---|
| Tissue | Low, n (%) | High, n (%) | P-value |
|---|
| HCC | 22 (23.4) | 72 (76.6) | <0.05 |
| Paracarcinoma | 51 (54.3) | 43 (45.7) |
|
MFAP2 expression is positively
correlated with that of EMT marker proteins
Immunohistochemistry results revealed high
N-cadherin protein expression in 74.5% (70 of 94) in HCC specimens
(Fig. 1 and Table IV). Spearman's correlation analysis
revealed a significant positive correlation between MFAP2
expression and that of N-cadherin (r=0.425, P=0.021; Table IV). High vimentin expression was
also detected in HCC tissues (77/94, 81.9%; Fig. 1), which was also positively
correlated with MFAP2 (r=0.393, P=0.037; Table IV). By contrast, E-cadherin, an
epithelial phenotype-related protein, was downregulated in HCC
tissues (Fig. 1A), and further
showed a significant negative correlation with MFAP2 expression
(r=−0.204, P=0.049; Table IV).
These results suggest that the expression of MFAP2 is positively
correlated with the expression of EMT marker proteins.
| Table IV.Association between MFAP2 expression
and that of E-cadherin, N-cadherin and vimentin in hepatocellular
carcinoma. |
Table IV.
Association between MFAP2 expression
and that of E-cadherin, N-cadherin and vimentin in hepatocellular
carcinoma.
|
| MFAP2 expression,
n |
|
|
|---|
|
|
|
|
|
|---|
|
Immunoreactivity | Low | High | r-value | P-value |
|---|
| E-cadherin
expression |
|
|
|
|
|
Low | 14 | 60 | −0.204 | 0.049 |
|
High | 8 | 12 |
|
|
| N-cadherin
expression |
|
|
|
|
|
Low | 13 | 11 |
0.425 | 0.021 |
|
High | 9 | 61 |
|
|
| Vimentin
expression |
|
|
|
|
|
Low | 10 | 7 |
0.393 | 0.037 |
|
High | 12 | 65 |
|
|
Association between MFAP2 expression
and patient clinical features
Significant associations between MFAP2 expression
and clinical characteristics were assessed across the 94 patients
with HCC analyzed (Table V). There
was no correlation between MFAP2 expression and age, sex, tumor
size, tumor nodule number, tumor capsula, liver cirrhosis, HBeAg
status and Child-Pugh grade. However, MFAP2 expression was
positively correlated with other clinical parameters, such as
vascular invasion (P=0.016), Edmondson grade (P=0.006), TNM status
(P=0.027) and AFP level (P=0.0006). These data indicate that the
expression of MFAP2 is associated with a number of clinical
features, and subsequently, with poor patient prognosis.
| Table V.MFAP2 expression status in relation
to clinicopathologic features in 94 patients with hepatocellular
carcinoma. |
Table V.
MFAP2 expression status in relation
to clinicopathologic features in 94 patients with hepatocellular
carcinoma.
|
|
| MFAP2 expression,
n |
|
|
|---|
|
|
|
|
|
|
|---|
| Parameter | Patients, n | Low | High |
χ2-value | P-value |
|---|
| Age |
|
|
|
|
|
|
≤60 | 69 | 14 | 55 | 1.404 | 0.236 |
|
>60 | 25 | 8 | 17 |
|
|
| Sex |
|
|
|
|
|
|
Male | 59 | 11 | 48 | 2.003 | 0.157 |
|
Female | 35 | 11 | 24 |
|
|
| Tumor size, cm |
|
|
|
|
|
| ≤5 | 32 | 10 | 22 | 1.666 | 0.197 |
|
>5 | 62 | 12 | 50 |
|
|
| Tumor nodule
number |
|
|
|
|
|
|
Single | 76 | 17 | 59 | 0.238 | 0.626 |
|
Multiple | 18 | 5 | 13 |
|
|
| Vascular
invasion |
|
|
|
|
|
|
Yes | 33 | 3 | 30 | 5.812 | 0.016 |
| No | 61 | 19 | 42 |
|
|
| Tumor capsula |
|
|
|
|
|
|
Absent | 38 | 10 | 28 | 0.302 | 0.583 |
|
Present | 56 | 12 | 44 |
|
|
| Edmondson
grade |
|
|
|
|
|
|
I–II | 53 | 18 | 35 | 7.556 | 0.006 |
|
III–IV | 41 | 4 | 37 |
|
|
| Cirrhosis |
|
|
|
|
|
|
Absent | 31 | 9 | 22 | 0.817 | 0.366 |
|
Present | 63 | 13 | 50 |
|
|
| HBeAg status |
|
|
|
|
|
|
Positive | 68 | 14 | 54 | 1.088 | 0.297 |
|
Negative | 26 | 8 | 18 |
|
|
| Child-Pugh
grade |
|
|
|
|
|
| A | 86 | 20 | 66 | 0.12 | 1.000 |
| B | 8 | 2 | 6 |
|
|
| TNM stage |
|
|
|
|
|
|
I–II | 58 | 18 | 40 | 4.918 | 0.027 |
|
III–IV | 36 | 4 | 32 |
|
|
| AFP (ng/ml) |
|
|
|
|
|
|
≤20 | 20 | 14 | 6 | 27.56 | 0.0006 |
|
>20 | 74 | 8 | 66 |
|
|
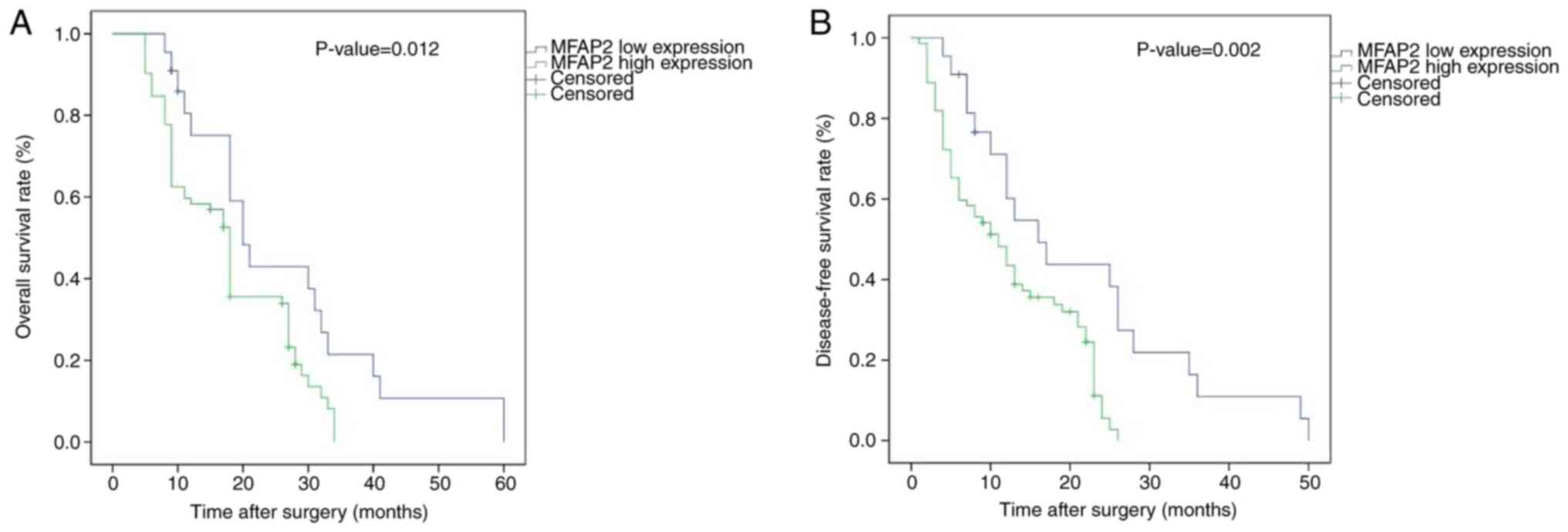
Survival analyses
Kaplan-Meier curves were generated, and the log-rank
test revealed that patients with high MFAP2 expression exhibited
shorter DFS and OS times than those with low MFAP2 expression
(Fig. 2). The median DFS and OS for
patients with high MFAP2 expression were 10 and 17 months,
respectively, whereas those for patients with low MFAP2 expression
were 12.5 (DFS) and 19 (OS) months. Results from univariate
analysis showed that high MFAP2 expression, high vascular invasion,
Edmundson grade, TNM stage and AFP level were predictors for poor
OS and DFS in patients with HCC (Table
VI). On the other hand, multivariate survival analysis revealed
that MFAP2 expression was an independent predictor for DFS
(HR=3.323; 95% CI=1.652–6.216; P<0.001) and OS (HR=3.895; 95%
CI=1.955–7.029; P<0.001).
| Table VI.Univariate and multivariate analysis
of factors associated with overall and Disease-free survival. |
Table VI.
Univariate and multivariate analysis
of factors associated with overall and Disease-free survival.
| A, Univariate
analysis |
|---|
|
|---|
|
| Overall
survival | Disease-free
survival |
|---|
|
|
|
|
|---|
| Parameter | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age, ≤60 vs. >60
years | 0.923 | 0.552–1.469 | 0.796 | 0.935 | 0.556–1.478 | 0.789 |
| Sex, male vs.
female | 0.956 | 0.587–1.523 | 0.778 | 0.964 | 0.590–1.511 | 0.812 |
| Tumor size, ≤5 vs.
>5 cm | 0.962 | 0.481–1.874 | 0.912 | 0.945 | 0.519–1.727 | 0.853 |
| Tumor nodules,
single vs. multiple | 1.326 | 0.831–2.127 | 0.246 | 1.285 | 0.827–2.012 | 0.266 |
| Vascular invasion,
yes vs. no | 0.635 | 0.236–0.843 | 0.016 | 0.543 | 0.313–0.914 | 0.014 |
| Tumor capsula,
absent vs. present | 0.594 | 0.329–1.026 | 0.068 | 0.654 | 0.367–1.026 | 0.091 |
| Edmondson grade,
I–II vs. III–IV | 2.627 | 1.567–4.765 | <0.001 | 2.357 | 1.428–4.458 | 0.003 |
| Cirrhosis, absent
vs. present | 1.252 | 0.629–2.433 | 0.484 | 1.117 | 0.541–2.110 | 0.727 |
| HBeAg status,
positive vs. negative | 0.852 | 0.423–1.621 | 0.652 | 0.721 | 0.323–1.435 | 0.336 |
| Child-Pugh grade, A
vs. B | 1.061 | 0.623–1.811 | 0.799 | 1.146 | 0.686–1.915 | 0.603 |
| TNM stage, I–II vs.
III–IV | 2.222 | 1.375–3.589 | 0.001 | 2.331 | 1.440–3.774 | 0.001 |
| AFP, ≤20 vs. >20
U/ml | 1.934 | 1.074–3.483 | 0.028 | 1.759 | 1.000–3.095 | 0.052 |
| MFAP2 expression,
high vs. low | 2.003 | 1.150–3.488 | 0.014 | 2.151 | 1.228–3.768 | 0.007 |
|
| B, Multivariate
analysis |
|
|
| Overall
survival | Disease-free
survival |
|
|
|
|
|
Parameter | HR | 95% CI | P-value | HR | 95% CI | P-value |
|
| Vascular invasion,
yes vs. no | 5.141 | 2.534–10.427 | <0.001 | 12.626 | 5.007–31.840 | <0.001 |
| Edmondson grade,
I–II vs. III–IV | 3.647 | 2.567–5.765 | <0.001 | 3.324 | 2.428–5.458 | 0.023 |
| TNM stage, I–II vs.
III–IV | 3.222 | 2.375–4.589 | <0.001 | 3.351 | 2.440–8.774 | <0.001 |
| AFP, ≤20 vs. >20
U/ml | 2.932 | 1.324–3.453 | 0.038 | 1.759 | 1.500–3.045 | 0.022 |
| MFAP2 expression,
high vs. low | 3.895 | 1.955–7.029 | <0.001 | 3.323 | 1.652–6.216 | <0.001 |
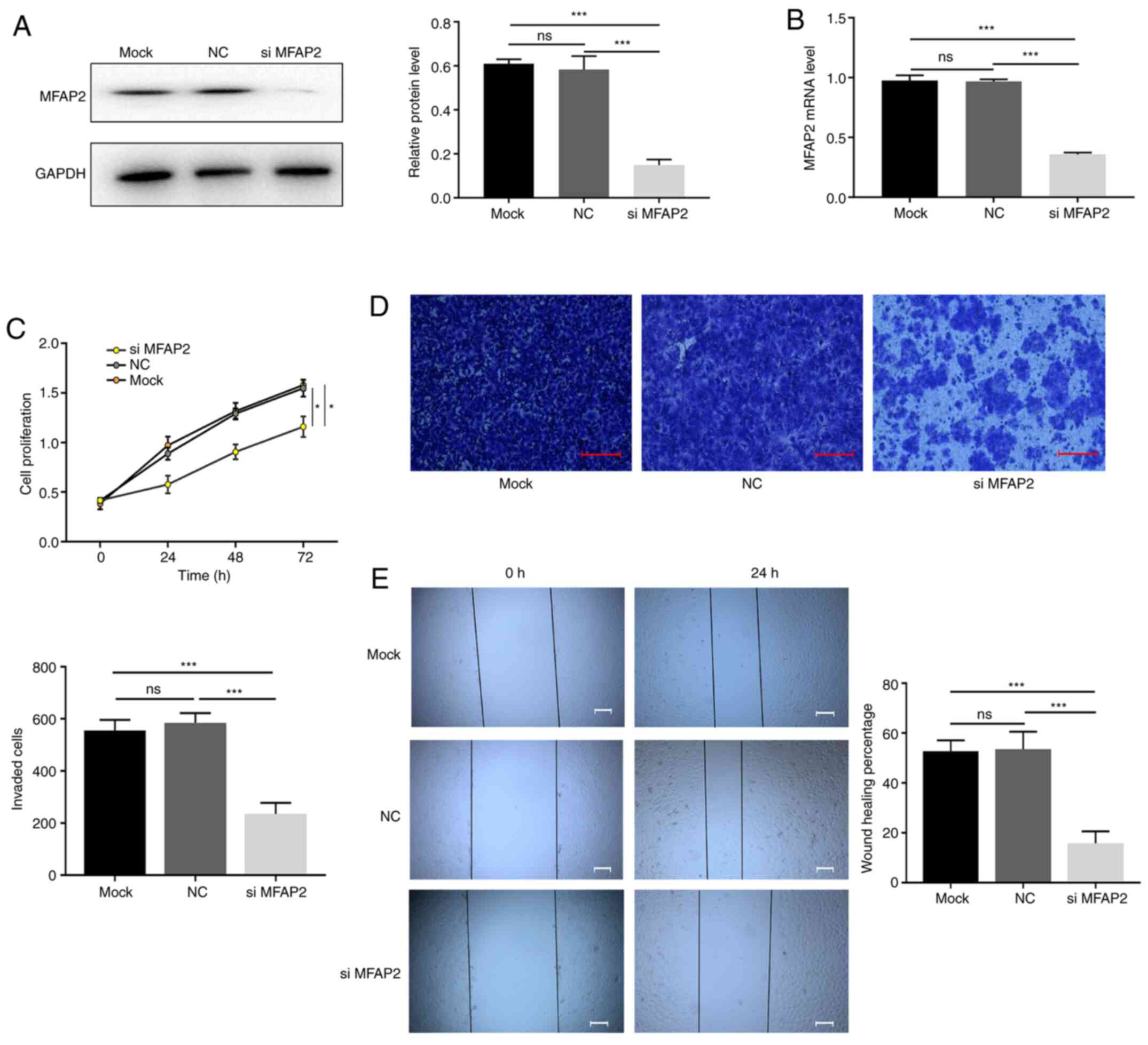
MFAP2-knockdown inhibits the
proliferation and invasiveness of MHCC97H cells
The effect of MFAP2 on the progression of HCC was
further evaluated using in vitro experiments. siRNA was used
to knock down MFAP2 in MHCC97H cells (Fig. 3A and B), after which cellular
proliferation and migration were evaluated. Compared with the NC
group, the CCK-8 assay results showed that knocking down MFAP2
significantly reduced the proliferation of MHCC97H cells (Fig. 3C). Transwell and wound-healing
experiments revealed that following MFAP2-knockdown, the migration
and invasion ability of MHCC97H cells was also reduced (Fig. 3D and E). These data indicate that
MFAP2-knockdown inhibited the proliferation and invasiveness of
MHCC97H cells.
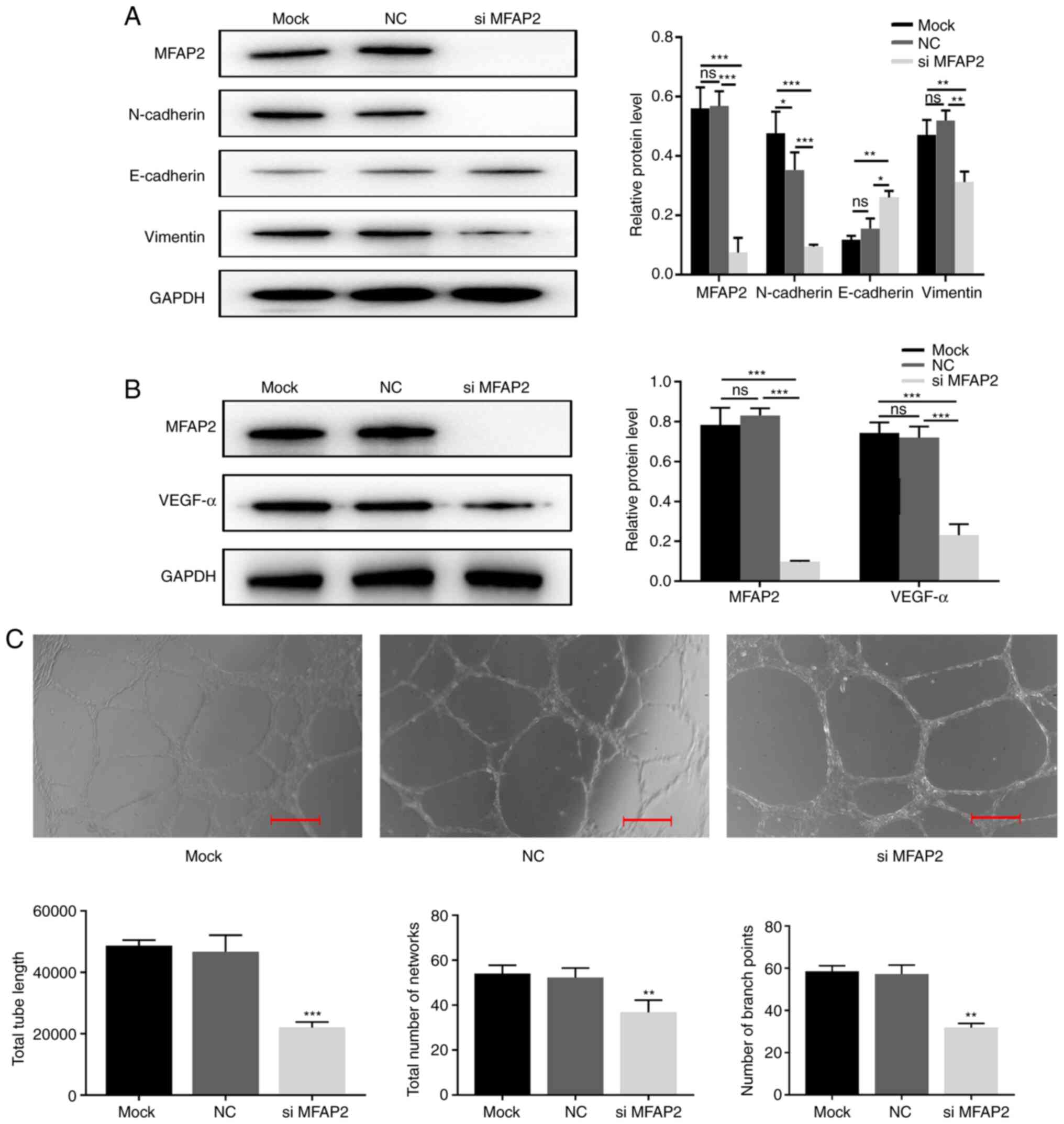
Effects of MFAP2-knockdown on EMT
As shown in Table
IV, the expression of MFAP2 in human HCC tissues was correlated
with EMT-related protein expression. Since EMT is a key event that
affects tumor differentiation and metastasis, the effect of MFAP2
on EMT was further investigated by detecting EMT-specific proteins
(including E-cadherin, N-cadherin and vimentin). After silencing
the expression of MFAP2 in MHCC97H cells, compared with the NC and
the Mock group, western blot analyses showed that the expression of
E-cadherin increased, while the expression of N-cadherin and
vimentin decreased, indicating that MFAP2 is essential for the EMT
of MHCC97H cells (Fig. 4A and
B).
Effects of MFAP2 on tube
formation
Vascular endothelial growth factor A (VEGFA) is the
most important VEGF in tumors (18).
In the present study, the expression of VEGFA was significantly
reduced following MFAP2-knockdown (Fig.
4B). Next, an in vitro HUVEC tube-formation assay was
used to directly determine the effect of MFAP2 on angiogenesis.
Compared with the NC group, the supernatant of MFAP2-knockout
MHCC97H cells reduced the average tube length, network and branch
point number of HUVECs (Fig. 4C).
These findings indicated that silencing MFAP2 reduces blood vessel
formation.
Discussion
MFAP2, a component of microfibers which was
originally identified in ligaments (19), interacts with numerous elastic fiber
components, such as tropoelastin and decorin, hence is considered a
component of elastin (20). Previous
studies have shown that an interaction between MFAP2 and fibrillin
1 can regulate the number of osteoclasts and bone resorption
(21). In addition, MFAP2 has been
shown to play an important role in tissue homeostasis and
differentiation (5–7). However, disease-induced fibrillin
mutations may alter the ability of myofibrils to bind MFAP2, and
lead to the occurrence and deterioration of the disease. Although
some reports have described MFAP2 upregulation in HCC (14), its exact role therein remains
unclear.
The present study revealed that MFAP2 was
upregulated in liver cancer, relative to normal adjacent tissues.
These results were consistent with those from TCGA database.
Furthermore, correlation analyses revealed a significant positive
correlation between MFAP2 upregulation and vascular invasion, high
Edmondson grade, elevated serum AFP and TNM stage in HCC tumors. In
addition, results from Kaplan-Meier survival analysis indicated
that patients with high MFAP2 expression showed poorer OS and DFS,
relative to those with low expression, and Cox regression analysis
also confirmed that MFAP2 upregulation was an independent predictor
for poor OS and DFS in patients with HCC.
Through in vitro experimentation, silencing
MFAP2 was found to significantly reduce the proliferation and
invasion abilities of HCC cell lines, which indirectly indicates
that MFAP2 affects the progression of HCC. Tumor progression is a
complex process regulated by numerous genes over multiple steps,
whereas EMT is a process of self-repair that occurs after damage to
adult epidermal cells (6,19). During tumor cell development, EMT
causes the loss of epithelial characteristics and gain of
mesenchymal cell traits (22,23),
bestowing tumor cells with stronger invasion and migration
capabilities. Downregulation of E-cadherin, which is associated
with cellular adhesion, is considered the most significant feature
of EMT (24–26). At the same time, interstitial factors
associated with tumor cells, such as Snail1, Snail2, Twist and
MMP-2, are often upregulated (27,28). In
the present study, it was evident that MFAP2 knockdown was
accompanied by the upregulation of vimentin and E-cadherin, while
N-cadherin was downregulated as a result of MFAP2-knockdown. These
results indicate that MFAP2 may regulate the invasion and
metastasis of HCC via EMT.
In addition to EMT, angiogenesis also plays an
important role tumor invasion and metastasis (29). There are two distinct phases of tumor
growth, that is, from the avascular slow growth phase to the
vascular fast proliferation phase. Tumor angiogenesis enables HCC
and other solid tumors to obtain sufficient nutrients, which is a
key link to promote the rapid growth of tumors (30).
Angiogenesis stimulating factors include a class of
cytokines known as antigenic growth factors, which act to stimulate
the formation of blood vessels (31). At present, angiogenic factors such as
platelet-derived growth factor, fibroblast growth factor,
transforming growth factor and VEGFA have been discovered (32), of which VEGFA plays a key role in
tumor angiogenesis (33). When tumor
cells develop, their expression levels will generally rise quickly.
Studies have shown that VEGFA is associated with tumor invasiveness
and susceptibility (34). Currently,
systemic therapies, including molecularly targeted drugs and immune
checkpoint inhibitors, have become common complementary therapies
for patients with HCC. VEGF-targeted therapeutic drugs, including
sorafenib and lenvatinib, are often used to inhibit the formation
of blood vessels in liver cancer. In the present study, clinical
data revealed that MFAP2 was associated with tumor angiogenesis,
and in vitro experimentation indicated that MFAP2-knockdown
was accompanied by low expression levels of VEGF. Therefore, we
hypothesize that MFAP2 may affect the formation of tumor blood
vessels by influencing VEGF. Inhibitors of MFAP2 may therefore be
combined with VEGF inhibitors to retard the formation of tumor
blood vessels.
In summary, the results of the present study suggest
that MFAP2 upregulation promotes tumor progression, is associated
with poor patient prognosis, and is an independent prognostic
biomarker for poor prognosis in HCC. Through in vitro
experimentation, MFAP2 was shown to inhibit the proliferation and
migration of MHCC97H cells by affecting the EMT process. In
addition, MFAP2 reduced the formation of tumor blood vessels by
inhibiting the expression of VEGFA. These results indicate that the
use of certain specific inhibitors to block MFAP2 activity may
provide novel targets for the development of anti-angiogenic
therapies against liver cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NZ and WJ designed the study, and performed the
experiments alongside FS. WJ wrote the manuscript and analyzed the
data alongside FS. WJ and FS conceived the study, supervised the
experiments and revised the manuscript for important intellectual
content. All authors read and approved the final manuscript. ZN, FS
and WJ confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Anhui Medical University (approval no. 2019-SH-010).
All patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shen JD, Fu SZ, Ju LL, Wang YF, Dai F, Liu
ZX, Ji HZ, Shao JG and Bian ZL: High expression of
ubiquitin-conjugating enzyme E2A predicts poor prognosis in
hepatocellular carcinoma. Oncol Lett. 15:7362–7368. 2018.PubMed/NCBI
|
|
3
|
Zhai X, Wang W, Ma Y, Zeng Y, Dou D, Fan
H, Song J, Yu X, Xin D, Du G, et al: Serum KIAA1199 is an
advanced-stage prognostic biomarker and metastatic oncogene in
cholangiocarcinoma. Aging (Albany NY). 12:23761–23777. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Penner AS, Rock MJ, Kielty CM and Shipley
JM: Microfibril-associated glycoprotein-2 interacts with
fibrillin-1 and fibrillin-2 suggesting a role for MAGP-2 in elastic
fiber assembly. J Biol Chem. 277:35044–35049. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Craft CS, Broekelmann TJ and Mecham RP:
Microfibril-associated glycoproteins MAGP-1 and MAGP-2 in disease.
Matrix Biol. 71-72:100–111. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Segade F, Suganuma N, Mychaleckyj JC and
Mecham RP: The intracellular form of human MAGP1 elicits a complex
and specific transcriptional response. Int J Biochem Cell Biol.
39:2303–2313. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mecham RP and Gibson MA: The
microfibril-associated glycoproteins (MAGPs) and the microfibrillar
niche. Matrix Biol. 47:13–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gong X, Dong T, Niu M, Liang X, Sun S,
Zhang Y, Li Y and Li D: lncRNA LCPAT1 upregulation promotes breast
cancer progression via enhancing MFAP2 transcription. Mol Ther
Nucleic Acids. 21:804–813. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dong SY, Chen H, Lin LZ, Jin L, Chen DX,
Wang OC and Ye ZQ: MFAP2 is a potential diagnostic and prognostic
biomarker that correlates with the progression of papillary thyroid
cancer. Cancer Manag Res. 12:12557–12567. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen Z, Lv Y, Cao D, Li X and Li Y:
Microfibril-associated protein 2 (MFAP2) potentiates invasion and
migration of melanoma by EMT and Wnt/β-catenin pathway. Med Sci
Monit. 26:e9238082020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Considine DPC, Jia G, Shu X, Schildkraut
JM, Pharoah PDP, Zheng W and Kar SP; Ovarian Cancer Association
Consortium, : Genetically predicted circulating protein biomarkers
and ovarian cancer risk. Gynecol Oncol. 160:506–513. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yao LW, Wu LL, Zhang LH, Zhou W, Wu L, He
K, Ren JC, Deng YC, Yang DM, Wang J, et al: MFAP2 is overexpressed
in gastric cancer and promotes motility via the MFAP2/integrin
α5β1/FAK/ERK pathway. Oncogenesis. 9:172020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun T, Wang D, Ping Y, Sang Y, Dai Y, Wang
Y, Liu Z, Duan X, Tao Z and Liu W: Integrated profiling identifies
SLC5A6 and MFAP2 as novel diagnostic and prognostic biomarkers in
gastric cancer patients. Int J Oncol. 56:460–469. 2020.PubMed/NCBI
|
|
14
|
Zhu X, Cheng Y, Wu F, Sun H, Zheng W,
Jiang W, Shi J, Ma S and Cao H: MFAP2 promotes the proliferation of
cancer cells and is associated with a poor prognosis in
hepatocellular carcinoma. Technol Cancer Res Treat.
19:15330338209775242020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhai LL, Cai CY, Wu Y and Tang ZG:
Correlation and prognostic significance of MMP-2 and TFPI-2
differential expression in pancreatic carcinoma. Int J Clin Exp
Pathol. 8:682–691. 2015.PubMed/NCBI
|
|
16
|
Zhai LL, Wu Y, Huang DW and Tang ZG:
Increased matrix metalloproteinase-2 expression and reduced tissue
factor pathway inhibitor-2 expression correlate with angiogenesis
and early postoperative recurrence of pancreatic carcinoma. Am J
Transl Res. 7:2412–2422. 2015.PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Claesson-Welsh L and Welsh M: VEGFA and
tumour angiogenesis. J Intern Med. 273:114–127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Craft CS: MAGP1, the extracellular matrix,
and metabolism. Adipocyte. 4:60–64. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gibson MA, Hatzinikolas G, Kumaratilake
JS, Sandberg LB, Nicholl JK, Sutherland GR and Cleary EG: Further
characterization of proteins associated with elastic fiber
microfibrils including the molecular cloning of MAGP-2 (MP25). J
Biol Chem. 271:1096–1103. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Clarke AW and Weiss AS:
Microfibril-associated glycoprotein-1 binding to tropoelastin:
multiple binding sites and the role of divalent cations. Eur J
Biochem. 271:3085–3090. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ju S, Wang F, Wang Y and Ju S: CSN8 is a
key regulator in hypoxia-induced epithelial-mesenchymal transition
and dormancy of colorectal cancer cells. Mol Cancer. 19:1682020.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang Z, Zhai X, Shi B, Luo D and Jin B:
KIAA1199 overexpression is associated with abnormal expression of
EMT markers and is a novel independent prognostic biomarker for
hepatocellular carcinoma. Onco Targets Ther. 11:8341–8348. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Solanas G, Porta-de-la-Riva M, Agustí C,
Casagolda D, Sánchez-Aguilera F, Larriba MJ, Pons F, Peiró S,
Escrivà M, Muñoz A, et al: E-cadherin controls beta-catenin and
NF-kappaB transcriptional activity in mesenchymal gene expression.
J Cell Sci. 121:2224–2234. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wendt MK, Tian M and Schiemann WP:
Deconstructing the mechanisms and consequences of TGF-β-induced EMT
during cancer progression. Cell Tissue Res. 347:85–101. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Willis BC and Borok Z: TGF-beta-induced
EMT: Mechanisms and implications for fibrotic lung disease. Am J
Physiol Lung Cell Mol Physiol. 293:L525–L534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Y, Ren H, Li J, Xue R, Liu H, Zhu Z,
Pan C, Lin Y, Hu A, Gou P, et al: Elevated HMGB1 expression induced
by hepatitis B virus X protein promotes epithelial-mesenchymal
transition and angiogenesis through STAT3/miR-34a/NF-κB in primary
liver cancer. Am J Cancer Res. 11:479–494. 2021.PubMed/NCBI
|
|
30
|
Zhou W, Yang L, Nie L and Lin H:
Unraveling the molecular mechanisms between inflammation and tumor
angiogenesis. Am J Cancer Res. 11:301–317. 2021.PubMed/NCBI
|
|
31
|
Jain S, Deore SV, Ghadi R, Chaudhari D,
Kuche K and Katiyar SS: Tumor microenvironment responsive
VEGF-antibody functionalized pH sensitive liposomes of docetaxel
for augmented breast cancer therapy. Mater Sci Eng C Mater Biol
Appl. 121:1118322021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kamiyama M and Augustin HG: Alternatively
spliced form of angiopoietin-2 as a new vascular rheostat. Cancer
Res. 81:35–37. 2021.PubMed/NCBI
|
|
33
|
Varone E, Decio A, Chernorudskiy A, Minoli
L, Brunelli L, Ioli F, Piotti A, Pastorelli R, Fratelli M, Gobbi M,
et al: The ER stress response mediator ERO1 triggers cancer
metastasis by favoring the angiogenic switch in hypoxic conditions.
Oncogene. 40:1721–1736. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Quan L, Ohgaki R, Hara S, Okuda S, Wei L,
Okanishi H, Nagamori S, Endou H and Kanai Y: Amino acid transporter
LAT1 in tumor-associated vascular endothelium promotes angiogenesis
by regulating cell proliferation and VEGF-A-dependent mTORC1
activation. J Exp Clin Cancer Res. 39:2662020. View Article : Google Scholar : PubMed/NCBI
|