Artesunate (ART) is a derivative of artemisinin that
is characterized by high efficacy, rapid effects, low toxicity and
reduced susceptibility to drug resistance (5,6). At
present, ART is commonly used for the treatment of mild to severe
malaria worldwide (7). However,
accumulating evidence has shown that ART also displays anticancer
properties, in addition to its antimalarial effect (8). For instance, ART has been reported to
induce apoptosis and autophagy in human bladder cancer cells
(9,10). Moreover, it can induce cell cycle
arrest, reactive oxygen species (ROS) generation and ferroptosis in
renal cell carcinoma (11). In the
present review, the potential anticancer effects of ART and the
underlying mechanism of action involved are summarized. The aim was
to provide a theoretical basis for the further development of ART
and its derivatives for the treatment of cancer.
ART has a hydrophilic group, and the
1,2,4-endoperoxide bridge is responsible for the antimalarial
activity of the drug. ART acts on all stages of malaria parasite
circulation. ART also may penetrate the cell membranes and generate
ROS, and a small amount of ART reaches the mitochondria of the
parasite, where ART and ROS react with each other, leading to
mitochondrial dysfunction (8). ART
is the only artemisinin derivative with high water solubility, due
to the addition of the hemisuccinate group. ART is metabolized to
docosahexaenoic acid (DHA) as it enters the body (16–18). ART
induces the generation of ROS, increasing malondialdehyde levels
and decreasing the levels antioxidants such as superoxide dismutase
and catalase, thereby causing alkylation of the proteins of the
Plasmodium parasite (19). At
present, ART is mainly used for the treatment of malaria of all
types (20), for immune regulation
(in type 1 diabetes in NOD mice) (21), as well as for liver (22), breast (23) and lung (24–26)
cancer.
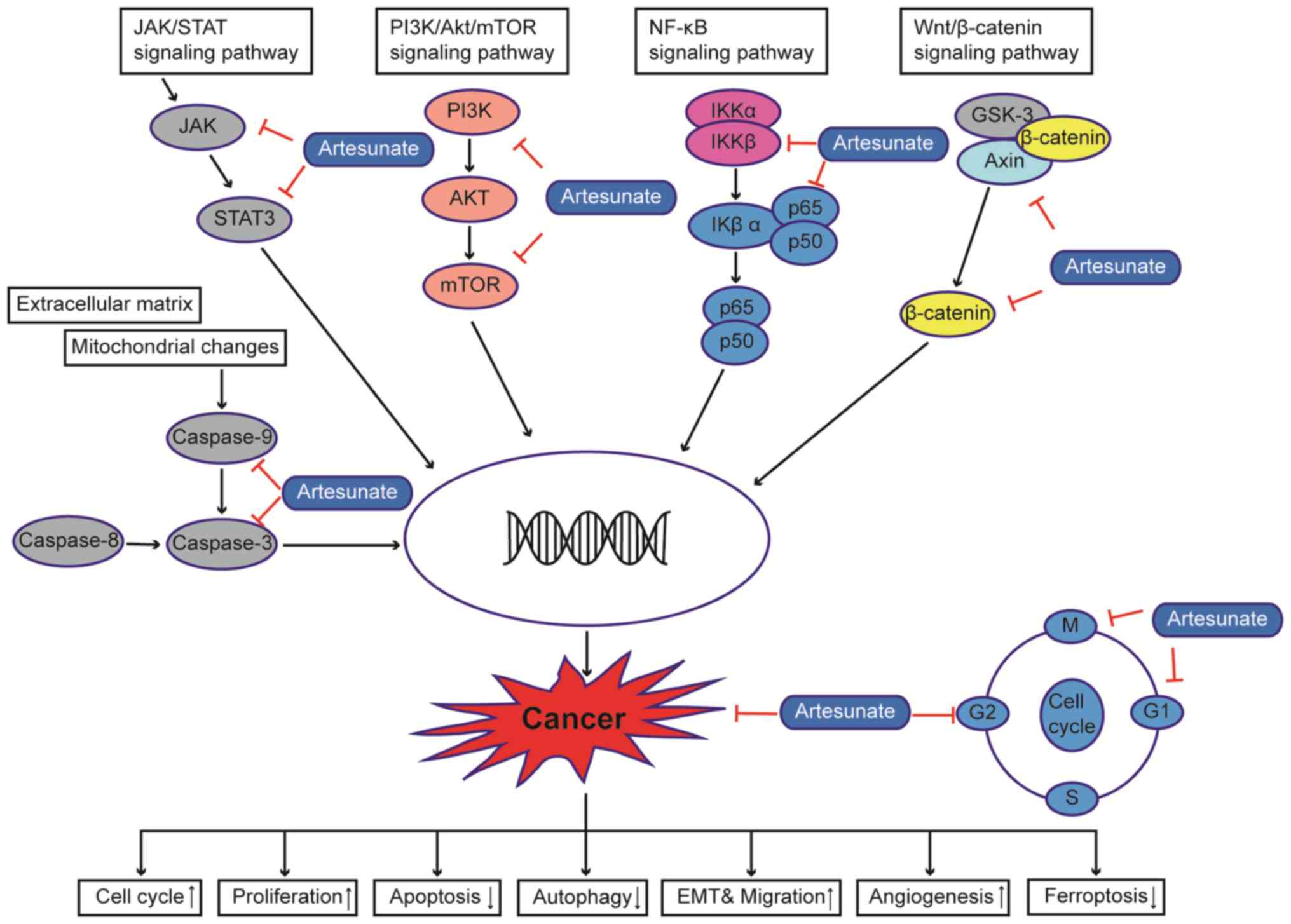
There is considerable evidence that ART can exert
anticancer effects on several types of cancer cells (6,15). ART
has been reported to induce apoptosis, differentiation and
autophagy in colorectal cancer cells by impairing angiogenesis
(27), inhibiting cell invasion and
migration (28), inducing cell cycle
arrest (11), upregulating ROS
levels, regulating signal transduction [for example, activating the
AMPK-mTOR-Unc-51-like autophagy activating kinase (ULK1) pathway in
human bladder cancer cells] (9) and
blocking immune escape (29). In
addition, ART has been shown to restore the sensitivity of a number
of cancer types to chemotherapeutic drugs by modulating various
signaling pathways; for example, ART can improve the apoptosis of
HCC by inhibiting the PI3K/AKT/mTOR pathway (30), and can increase liver cancer cell
sensitivity to sorafenib via suppression of the MEK/ERK pathway
(31) (Fig. 1).
Apoptosis is a type of programmed cell death that
does not elicit inflammatory responses (32). A number of studies have shown that
ART can induce apoptosis by activating the mitochondria-dependent
pathway, specifically by mediating the activation of caspase-3 and
−9 and the release of cytochrome c into the cytosol after
permeabilization of the mitochondrial membrane (33). Additionally, ART can induce HL-60
human acute promyelocytic leukemia cell and KG1a acute myeloid
leukemia cell death by regulating antiapoptotic proteins, such as
Bcl-2, as well as proapoptotic proteins, such as Bid and Bak,
through inhibition of the MEK/ERK and PI3K/Akt pathways (34). ART has also been demonstrated to
induce T helper 1 cell differentiation and promote apoptosis in
ovarian cancer cells via the microRNA (miR)-142/sirtuin 1 pathway
(35).
Autophagy is a conserved, self-degrading system that
is essential for maintaining cell homeostasis under stress
conditions, and which has been demonstrated to serve an important
role in cancer in association with a family of autophagy-related
proteins (LC3B) (36). ART can
induce autophagy and increase the levels of CD155 in uterine corpus
endometrial carcinoma (UCEC) cells. Moreover, it also regulates the
interaction between CD155 and its receptor on the NK92 natural
killer cell line by upregulating the co-stimulator CD226 and
downregulating the co-inhibitor TIGIT, thereby enhancing the
cytotoxicity of these cells. Thus, ART has a dual anticancer effect
on UCEC cells (37). ART also
induces autophagy by upregulating ROS production and activating the
AMP-activated protein kinase/mTOR/ULK1 pathway in human bladder
cancer cells (9).
Blood vessels provide oxygen and a nutrient supply
for the growth of tumors, which also facilitate the proliferation,
migration and subsequent invasion of malignant tumor cells in the
long term (47). Angiogenesis is a
dynamic and complex process that is regulated by a variety of
mechanisms. Inhibition of angiogenesis has become a therapeutic
strategy for pancreatic cancer (48), breast cancer (49) and ovarian cancer (50). Chen et al (51) demonstrated that ART could
downregulate the expression of VEGF and angiopoietin-1 in RPMI8226
myeloma cells, decrease the activation of ERK1and inhibit
angiogenesis. Their study indicated that ART possessed a potential
anti-myeloma effect, which was mediated by the inhibition of
angiogenesis.
Ferroptosis is a recently identified form of
regulated cell death, which is characterized by iron overload,
lipid ROS accumulation and lipid peroxidation (55). Evidence suggests that ferroptosis is
closely associated with the occurrence, development and inhibition
of cancer (56). Zhang et al
(26) demonstrated that ART could
upregulate the mRNA levels of transferrin receptor (a positive
regulator of ferroptosis), thus inducing apoptosis and ferroptosis
in A549 non-small cell lung cancer (NSCLC) cells. Li et al
(57) showed that ART enhanced the
anticancer effects of low-dose sorafenib (a novel multi-targeted
oral drug for the treatment of gastroenteric tumors) against Huh7,
SNU-449, and SNU-182 HCC cell lines in vitro and against a
Huh7 cell xenograft model in BALB/c nude mice. In addition,
ART-induced lysosome activation synergizes with the pro-oxidative
effects of sorafenib to sequentially promote lysosomal cathepsin
B/L activation, ferritin degradation, lipid peroxidation and
ferroptosis (57).
Previous studies have reported that ART exerted
minimal toxicity, was cost-effective and was effective for treating
different types of cancer (Table I)
(58–78). The potential anticancer properties of
ART in different types of cancer are discussed below.
Central nervous system tumors comprise a group of
malignancies that originate from tissues or structures of the
central nervous system and exhibit an incidence of 5.6 per 100,000
person-years in children under the age of 19 (85–87). ART
can selectively downregulate the expression of survivin and induce
the DNA damage response in glial cells to increase cell apoptosis
and cell cycle arrest, resulting in increased sensitivity to
radiotherapy (88). Previously, Wei
et al (67) found that ART
affected the nuclear localization of sterol regulatory
element-binding protein 2 (SREBP2) by decreasing the expression of
3-hydroxy-3-methylglutaryl-CoA reductase and inhibited the
mevalonate pathway, which in turn influenced the metabolism of
glioma cells. In addition, ART disrupted the interaction between
P53 and SREBP2 (which negatively regulates P53 and inhibits
senescence), upregulated the expression of P21 and induced
senescence in the U251, U87, U138 and SK-N-SH human glioma cell
lines (67). The combination of ART
and rapamycin (a specific inhibitor of mTOR) has been shown to
synergistically decrease translation-controlled tumor protein
(TCTP) expression and enhance the cytotoxicity of malignant
peripheral nerve sheath tumor (MPNST) cells via mTOR-TCTP positive
feedback loop, the results also suggested that TCTP may be a new
target for the treatment of neurofibromatosis type 1-associated
tumors and MPNSTs (89).
Thyroid cancer is the most common cancer in the
endocrine system, and its incidence is increasing worldwide
(90). Anaplastic thyroid carcinoma
(ATC) is an aggressive malignancy that is almost always fatal and
lacks effective systemic treatment options. It is highly resistant
to chemotherapy due to its undifferentiated and aggressive
characteristics (91,92). Ma and Fei (91) showed that ART could inhibit growth
and induce apoptosis in ATC cells (8505C, 8505C-r, KAT-4-r and
KAT-4), by suppressing mitochondrial respiration and acting
synergistically with chemotherapy drug doxorubicin without
affecting glycolysis. Thus, ART led to oxidative stress and damage
in ATC cells. Their work suggested that ART was a potential
complement to the treatment of ATC, particularly cases with
chemoresistance (91).
Ovarian cancer is the seventh most common type of
malignant neoplasm in women and the eighth cause of mortality
(102–104). Most patients with ovarian cancer
are typically diagnosed at an advanced stage of the disease
(105). Ovarian cancer is treated
with platinum chemotherapy following surgical resection (106). However, the recurrence rate is high
(107,108) and the survival rates of ovarian
cancer with International Federation of Gynecology and Obstetrics
stage III and IV are only 10–30% (109). ART has been found to significantly
reduce the expression of VEGF in the HO-8910 human ovarian cancer
cell line, as well as that of KDR/flk-1 (VEGF receptor) in
endothelial cells and HO-8910 cells, thereby significantly
inhibiting angiogenesis in a dose-dependent form. Additionally, ART
resulted in reduced xenograft tumor growth in nude mice, with no
clear toxicity to the animal (110). ART could reduce the total amount of
RAD51 and the formation of RAD51 foci in ovarian cancer cells
sustaining DNA damage. Moreover, the downregulation of RAD51
conferred ovarian cancer cells an increased sensitivity to
cisplatin (111). ART combined with
cisplatin can synergistically induce DNA double-strand breaks and
inhibit the proliferation of the HO8910 and SKOV-3 human ovarian
cancer cell lines (111). ART
induced the production of ROS and reduced proliferation in HEY1,
HEY2 and SKOV-3 ovarian cancer cells, which were associated with
downregulation in the expression levels of regulatory proteins of
the cell cycle, including cyclin D3, CDKs (CDK4, CDK2, and CDK1),
Rb, E2F-1 and CDC25C, while the tumor suppressor p21WAF1/CIP1, as
well as phosphorylated Chk2 kinase which is important in the DNA
damage response and an inhibitor of the CDC25 phosphatases were
upregulated (70).
Esophageal cancer (EC) is a common malignancy and
has a high incidence rate in China (112). Although therapeutic approaches have
improved, the 5-year survival of EC is <20% (113). ART can induce apoptosis and cell
cycle arrest in the Eca109 and Ec9706 EC cell lines by upregulating
Bax and caspase-3 and reducing mitochondrial membrane potential, as
well as Bcl-2 and CDC25A expression in a concentration-dependent
manner (72). In addition, an in
vivo study showed that ART produced a dose-dependent
Eca109-transplanted tumor regression in Balb/c nude mice, with
little side effects. These results revealed that CDC25A was a
molecular target of ART and that ART could inhibit the growth of EC
cells by inducing apoptosis and G0/G1cell
cycle arrest (72). Fei et al
(42) demonstrated that ART
inhibited the proliferation of EC cells, enhanced radiosensitivity
of TE-1 cells in vitro and enhanced the effect of apoptosis
induced by irradiation in TE-1 cells by upregulating P21 and
downregulating the expression of cyclin D1, RAD51, RAD54, Ku70 and
Ku86 protein of irradiated TE-1 cells. Moreover, ART also could
aggravate DNA damage of EC cells and prolong the formation of
γ-H2AX foci induced by IR in TE-1 cells. The results indicated that
ART may be a promising radiosensitizer for the treatment of EC. In
another study, Wang et al (73) found that ART can reverse doxorubicin
resistance in EC by downregulating the expression of ATP-binding
cassette G2 in Eca109 cells. ART was reported to inhibit the
proliferation, migration and invasion of KYSE-150 esophageal
squamous cell carcinoma cells by suppressing cell elasticity and
increasing adhesion; ART also may increase the apoptosis rate by
altering the cytoskeleton of KYSE-150 cells (114).
Gastric cancer is the fourth leading cause of
cancer-related mortality in the world, with a 5-year survival rate
of <40% (115,116). ART can inhibit the proliferation of
the gastric cancer cell lines SGC-7901, BGC-823 and AGS in a
concentration-dependent manner, BGC-823 cells treated with ART
exhibited calcium overload, downregulated expression levels of VEGF
and upregulated expression levels of calpain-2 (117). ART treatment can also inhibit the
proliferation of the SGC-7901 gastric adenocarcinoma cell line and
induce apoptosis; the mechanism may be associated with Bax and
caspase-3 upregulation and CDC25A and Bcl-2 downregulation
(74). In addition, ART could
prevent the growth of Helicobacter pylori and gastric cancer
cells, inhibit the adhesion of Helicobacter pylori to these
cells and reduce Helicobacter pylori-enhanced ROS
production. Moreover, ART significantly reduces the number of tumor
nodules and tumor size in a gastric cancer mouse model by
inhibiting the NF-κB signaling pathway (118).
Colorectal cancer (CRC) is one of the most common
types of cancer worldwide and has incidence and mortality rates
globally (119,120). ART was found to inhibit CRC
proliferation and promote apoptosis in a dose-dependent manner to
significantly suppress the growth of colorectal tumors, decrease
the physiological activity of cancer and delay spontaneous liver
metastasis in the CLY CRC cell line. These anticancer effects were
associated with the membrane translocation of β-catenin and the
inhibition of unrestricted Wnt/β-catenin signaling (121). In addition, ART can reverse the
immunosuppression by downregulating the concentrations of TGF-β1
and IL-10 in Colon26 and RKO CRC cells (122). Jiang et al (76) found that ART induced apoptosis by
increasing the protein levels of cleaved caspase-3, poly-ADP ribose
polymerase (PARP), caspase-9 and Bax protein levels, while
decreasing the levels of LC3 and beclin-1 in HCT116 colon cancer
cells. ART can reduce the levels of oxidative stress and
inflammatory markers, downregulate cyclo-oxygenase-2, induce nitric
oxide (NO) synthase, NF-κB and IF-1β and reduce the risk of colon
cancer (77).
Lung cancer is the most common cancer in the world
and the leading cause of cancer death (123), which has an overall 5-year survival
rate of ~15% (124). Despite
advances in treatment, progressive NSCLC still severely limits
survival and requires new therapeutic compounds (125). ART can significantly inhibit the
invasion and migration of NSCLC cells (H1395, A549, LXF289 and H460
cells) by downregulating the transcription of urokinase-type
plasminogen activator, MMP-2 and MMP-7, whilst inhibiting AP-1 and
NF-κB-transactivation (126). In
addition, ART promotes radiosensitivity in A549 cells in
vitro and in vivo, possibly by inducing cell cycle
arrest at the G2/M phase through the NO signaling
pathway (127). Wang et al
(128) found that ART could inhibit
cell migration by upregulating the expression of the epithelial
marker E-cadherin in A549 and H1975 NSCLC cell lines. In another
study, ART could inhibit the invasion of A549 cells, and the
mechanism may be associated with the reduced expression of
intercellular adhesion molecule-1 and MMP-9 (129). Furthermore, ART inhibits the
proliferation of A549 and H1299 cells by arresting the cell cycle
at the G1 phase and suppresses lung tumor progression by
inhibiting the Wnt/β-catenin pathway (130). In A549 cells, ART combined with
cisplatin blocks the cell cycle at the G2/M phase and
induces apoptosis by upregulating the expression of Bax, p53, p21,
caspase-3, caspase-7 and caspase-9, whilst synergistically
regulating the activity of the MAPK pathway by downregulating
p-P38, p-JNK and p-ERK levels, which results in potentiated effects
against cancer cell proliferation on A549 cells (131).
Liver cancer is highly malignant and insensitive to
cytotoxic chemotherapy, and is associated with a very poor patient
prognosis (132,133). ART can activate caspase-3, increase
the Bax/Bcl-2 ratio and PARP, whilst downregulating mouse double
minute 2, which leads to induced apoptosis on human hepatocellular
carcinoma (HCC) cells but had little effect on normal cells
(134). The anticancer effects of
ART nanoliposomes on human HepG2 cells was stronger than those
mediated by ART active pharmaceutical ingredient at the same
concentration (135). ART may
function as a potential inhibitor of STAT3 in HCC cells to regulate
STAT3 targets, including caspase-3, Bcl-xl and survivin, interfere
with STAT3 dimerization and inhibition of both constitutive and
IL-6-inducible STAT3, leading to cell apoptosis in vitro
(78). Jing et al (30) also revealed that ART could inhibit
phosphorylation of AKT and mTOR significantly, and induce apoptosis
in HCC (SK-hep1 and SM-7721 cell lines) by inhibiting the
PI3K/AKT/mTOR pathway. In addition, ART combined with sorafenib
(which is a novel multi-targeted oral drug for the treatment of
cancer) further increased the apoptosis of HCC cells by dual
inhibition of both RAF/RAF/MAPK pathway and PI3K/AKT/mTOR pathway.
Thus, the study identified a potential treatment strategy combining
ART with sorafenib for the treatment of advanced HCC.
In a previous study, ART has been reported to induce
lactate dehydrogenase release and cell death in necrosis-sensitive
cholangiocarcinoma (136). Wang
et al (137) found that ART
could significantly inhibit proliferation in the Burkitt lymphoma
Raji cell line, where it induced apoptosis and autophagy. The
combination of ART and bromocriptine can synergistically promote
apoptosis by inhibiting miR-200c expression and increasing that of
PTEN in lactinomas (61). Chauhan
et al (138) found that ART
induced ROS production and subsequent cell death in a
receptor-interacting protein 1-dependent manner in human renal
carcinoma. ART exerted a potent antiproliferative effect on
polyomavirus-positive Merkel cell carcinoma (MCC) cells with good
overall tolerance and induced ferroptosis (139). In addition, ART also significantly
suppressed the growth of established MCC tumors in xenotransplanted
mice, suggesting that ART may be used for the treatment of MCC
(138). In another study, ART
blocked the Wnt/catenin pathway to inhibit the proliferation,
migration and invasion of uveal melanoma cells (primary 92.1 and
metastatic Omm2.3 UM cells), mainly by suppressing the
phosphorylation of GSK3β at Ser9 and decreasing the protein levels
of β-catenin and its downstream targets (c-Myc and cyclin D1)
(140). Wang et al (141) found that ART decreased androgen
receptor (AR) expression, increased the expression and the
catalytic activity of DNA methyltransferase3b (DNMT3b) in 22rv1
cells either in transplanted mice or in vitro. ART can
suppress tumor growth of prostatic cancer cells through AR-DNMT3b
pathway, suggesting it may be used for the treatment of prostate
cancer in the future. Yang et al (142) found that ART induced mitochondrial
dysfunction and cell apoptosis in the WERI-Rb1 and Y79 human
retinoblastoma cell lines and in the ARPE-19 human retinal pigment
epithelium cell line by upregulating Kruppel-like factor 6
expression, increasing the Bax/Bcl-2 ratio, promoting the release
of cytochrome c and stimulating the cleavage of caspase-9
and −3. Roh et al (143)
demonstrated that ART could induce ferroptosis in head and neck
(HNC) cells via cellular glutathione depletion and ROS
accumulation, and ART sensitivity decreased in some
cisplatin-resistant HNCs as a result of Nrf2-ARE pathway
activation. Berköz et al (144) suggested that ART treatment could
decrease cell migration, invasion and colony formation in the A375
human melanoma cell line, possibly by inhibiting STAT3, Src
activation and the protein expression of STAT3-associated
molecules, including MMP-2, MMP-9, myeloid-cell leukemia 11,
Bcl-xl, VEGF and Twist.
Cancer is one of the most life-threatening diseases.
With the increasing prevalence of cancer, the development of
anticancer agents has become a key field of clinical and scientific
research. Developments in medical science and technology have
enabled the extraction of bioactive components from Traditional
Chinese medicines for research due to their reported anticancer
effects and lack of adverse reactions. ART has been demonstrated to
be effective against leukemia, breast cancer, gastrointestinal
tumors and other types of cancer (8,23,145).
Importantly, since it is a drug that is already being used for the
treatment of malaria, ART has a reliable safety record for clinical
use. Although the amount of clinical data regarding the use of ART
as an anticancer drug remains limited, preliminary results have
been encouraging in terms of efficacy and tolerance (22). Combination therapy should be a key
consideration in the future. In addition, development of modified
derivatives of ART after structural modifications or modifying the
treatment regimen to optimize the efficacy and toxicity profile are
also possible directions for future research.
To conclude, existing information provides evidence
supporting the use of ART as an anticancer agent. However, data
from systematic in vivo animal and human studies are
required to improve our understanding of the anticancer effects and
mechanism of action of ART in the future.
Not applicable.
The present study was supported by the Health
Commission of Hubei Province scientific research project (grant no.
WJ2021Q015), the College Students Innovation Program of Yangtze
University (grant no. Yz2020338) and the Health Commission of Hubei
Province Scientific Research Project (grant no. WJ2019-17).
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the present
study.
LL designed and supervised the study. JH, YZ and FW
reviewed the references. XY wrote the manuscript. YZ, FW and JZ
contributed to the table and figure. XY and LL revised the
manuscript. All authors read and approved the final manuscript.
Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Miller KD, Fidler-Benaoudia M, Keegan TH,
Hipp HS, Jemal A and Siegel RL: Cancer statistics for adolescents
and young adults, 2020. CA Cancer J Clin. 70:443–459. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD, Fuchs HE and Jemal
A: Cancer statistics, 2021. CA Cancer J Clin. 71:7–33. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cao W, Chen HD, Yu YW, Li N and Chen WQ:
Changing profiles of cancer burden worldwide and in China: A
secondary analysis of the global cancer statistics 2020. Chin Med J
(Engl). 134:783–791. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ishikawa C, Senba M and Mori N: Evaluation
of artesunate for the treatment of adult T-cell leukemia/lymphoma.
Eur J Pharmacol. 872:1729532020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Slezakova S and Ruda-Kucerova J:
Anticancer activity of artemisinin and its derivatives. Anticancer
Res. 37:5995–6003. 2017.PubMed/NCBI
|
|
7
|
Cen YY, Zao YB, Li P, Li XL, Zeng XX and
Zhou H: Research progress on pharmacokinetics and pharmacological
activities of artesunate. Zhongguo Zhong Yao Za Zhi. 43:3970–3978.
2018.(In Chinese). PubMed/NCBI
|
|
8
|
Khanal P: Antimalarial and anticancer
properties of artesunate and other artemisinins: Current
development. Monatsh Chem. Mar 30–2021.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou X, Chen Y, Wang F, Wu H, Zhang Y, Liu
J, Cai Y, Huang S, He N, Hu Z and Jin X: Artesunate induces
autophagy dependent apoptosis through upregulating ROS and
activating AMPK-mTOR-ULK1 axis in human bladder cancer cells. Chem
Biol Interact. 331:1092732020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao F, Vakhrusheva O, Markowitsch SD,
Slade KS, Tsaur I, Cinatl J Jr, Michaelis M, Efferth T, Haferkamp A
and Juengel E: Artesunate impairs growth in cisplatin-resistant
bladder cancer cells by cell cycle arrest, apoptosis and autophagy
induction. Cells. 9:26432020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Markowitsch SD, Schupp P, Lauckner J,
Vakhrusheva O, Slade KS, Mager R, Efferth T, Haferkamp A and
Juengel E: Artesunate inhibits growth of sunitinib-resistant renal
cell carcinoma cells through cell cycle arrest and induction of
ferroptosis. Cancers (Basel). 12:31502020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Klaunig JE: Oxidative stress and cancer.
Curr Pharm Des. 24:4771–4778. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lichota A and Gwozdzinski K: Anticancer
activity of natural compounds from plant and marine environment.
Int J Mol Sci. 19:35332018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chekem L and Wierucki S: Extraction of
artemisinin and synthesis of its derivates artesunate and
artemether. Med Trop (Mars). 66:602–605. 2006.(In French).
PubMed/NCBI
|
|
15
|
Efferth T: From ancient herb to modern
drug: Artemisia annua and artemisinin for cancer therapy.
Semin Cancer Biol. 46:65–83. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei T and Liu J: Anti-angiogenic
properties of artemisinin derivatives (review). Int J Mol Med.
40:972–978. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ho WE, Peh HY, Chan TK and Wong WS:
Artemisinins: Pharmacological actions beyond anti-malarial.
Pharmacol Ther. 142:126–139. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang J, Sun X, Wang L, Wong YK, Lee YM,
Zhou C, Wu G, Zhao T, Yang L, Lu L, et al: Artesunate-induced
mitophagy alters cellular redox status. Redox Biol. 19:263–273.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alagbonsi AI, Salman TM, Sulaiman SO,
Adedini KA and Kebu S: Possible mechanisms of the hypoglycaemic
effect of artesunate: Gender implication. Metabol Open.
10:1000872021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Venturini E, Zammarchi L, Bianchi L,
Montagnani C, Tersigni C, Bortone B, Chiappini E and Galli L:
Efficacy and safety of intravenous artesunate in children with
severe imported malaria. Pediatr Infect Dis J. 39:e2202020.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Z, Shi X, Liu J, Shao F, Huang G, Zhou
Z and Zheng P: Artesunate prevents type 1 diabetes in NOD mice
mainly by inducing protective IL-4-producing T cells and regulatory
T cells. FASEB J. 33:8241–8248. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li H, Xu K, Pian G and Sun S: Artesunate
and sorafenib: Combinatorial inhibition of liver cancer cell
growth. Oncol Lett. 18:4735–4743. 2019.PubMed/NCBI
|
|
23
|
Pirali M, Taheri M, Zarei S, Majidi M and
Ghafouri H: Artesunate, as a HSP70 ATPase activity inhibitor,
induces apoptosis in breast cancer cells. Int J Biol Macromol.
164:3369–3375. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Z, Wang Q, He T, Li W, Liu Y, Fan Y,
Wang Y, Wang Q and Chen J: The combination of artesunate and
carboplatin exerts a synergistic anti-tumour effect on non-small
cell lung cancer. Clin Exp Pharmacol Physiol. 47:1083–1091. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao Y, Liu J and Liu L: Artesunate
inhibits lung cancer cells via regulation of mitochondrial membrane
potential and induction of apoptosis. Mol Med Rep. 22:3017–3022.
2020.PubMed/NCBI
|
|
26
|
Zhang Q, Yi H, Yao H, Lu L, He G, Wu M,
Zheng C, Li Y, Chen S, Li L, et al: Artemisinin derivatives inhibit
non-small cell lung cancer cells through induction of ROS-dependent
apoptosis/ferroptosis. J Cancer. 12:4075–4085. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Verma S, Das P and Kumar VL:
Chemoprevention by artesunate in a preclinical model of colorectal
cancer involves down regulation of β-catenin, suppression of
angiogenesis, cellular proliferation and induction of apoptosis.
Chem Biol Interact. 278:84–91. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma JD, Jing J, Wang JW, Yan T, Li QH, Mo
YQ, Zheng DH, Gao JL, Nguyen KA and Dai L: A novel function of
artesunate on inhibiting migration and invasion of fibroblast-like
synoviocytes from rheumatoid arthritis patients. Arthritis Res
Ther. 21:1532019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qian P, Zhang YW, Zhou ZH, Liu JQ, Yue SY,
Guo XL, Sun LQ, Lv XT and Chen JQ: Artesunate enhances γδ
T-cell-mediated antitumor activity through augmenting γδ T-cell
function and reversing immune escape of HepG2 cells.
Immunopharmacol Immunotoxicol. 40:107–116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jing W, Shuo L, Yingru X, Min M, Runpeng
Z, Jun X and Dong H: Artesunate promotes sensitivity to sorafenib
in hepatocellular carcinoma. Biochem Biophys Res Commun. 519:41–45.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He W, Huang X, Berges BK, Wang Y, An N, Su
R and Lu Y: Artesunate regulates neurite outgrowth inhibitor
protein B receptor to overcome resistance to sorafenib in
hepatocellular carcinoma cells. Front Pharmacol. 12:6158892021.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu X, Lai Y and Hua ZC: Apoptosis and
apoptotic body: Disease message and therapeutic target potentials.
Biosci Rep. 39:BSR201809922019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wong YK, Xu C, Kalesh KA, He Y, Lin Q,
Wong WSF, Shen HM and Wang J: Artemisinin as an anticancer drug:
Recent advances in target profiling and mechanisms of action. Med
Res Rev. 37:1492–1517. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen S, Gan S, Han L, Li X, Xie X, Zou D
and Sun H: Artesunate induces apoptosis and inhibits the
proliferation, stemness, and tumorigenesis of leukemia. Ann Transl
Med. 8:7672020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen X, Zhang XL, Zhang GH and Gao YF:
Artesunate promotes Th1 differentiation from CD4+ T cells to
enhance cell apoptosis in ovarian cancer via miR-142. Braz J Med
Biol Res. 52:e79922019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Peng J, Yuan C, Wu Z, Wang Y, Yin W, Lin
Y, Zhou L and Lu J: Upregulation of microRNA-1 inhibits
proliferation and metastasis of breast cancer. Mol Med Rep.
22:454–464. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang J, Zhou L, Xiang JD, Jin CS, Li MQ
and He YY: Artesunate-induced ATG5-related autophagy enhances the
cytotoxicity of NK92 cells on endometrial cancer cells via
interactions between CD155 and CD226/TIGIT. Int Immunopharmacol.
97:1077052021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hayes JD, Dinkova-Kostova AT and Tew KD:
Oxidative stress in cancer. Cancer Cell. 38:167–197. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kohan R, Collin A, Guizzardi S, Tolosa de
Talamoni N and Picotto G: Reactive oxygen species in cancer: A
paradox between pro- and anti-tumour activities. Cancer Chemother
Pharmacol. 86:1–13. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kirtonia A, Sethi G and Garg M: The
multifaceted role of reactive oxygen species in tumorigenesis. Cell
Mol Life Sci. 77:4459–4483. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Srinivas US, Tan BWQ, Vellayappan BA and
Jeyasekharan AD: ROS and the DNA damage response in cancer. Redox
Biol. 25:1010842019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fei Z, Gu W, Xie R, Su H and Jiang Y:
Artesunate enhances radiosensitivity of esophageal cancer cells by
inhibiting the repair of DNA damage. J Pharmacol Sci. 138:131–137.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Petricciuolo M, Davidescu M, Fettucciari
K, Gatticchi L, Brancorsini S, Roberti R, Corazzi L and Macchioni
L: The efficacy of the anticancer 3-bromopyruvate is potentiated by
antimycin and menadione by unbalancing mitochondrial ROS production
and disposal in U118 glioblastoma cells. Heliyon. 6:e057412020.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yu X, Wang X, Wang X, Zhou Y, Li Y, Wang
A, Wang T, An Y, Sun W, Du J, et al: TEOA inhibits proliferation
and induces DNA damage of diffuse large b-cell lymphoma cells
through activation of the ROS-dependent p38 MAPK signaling pathway.
Front Pharmacol. 11:5547362020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang H, Li M, Zhu X, Zhang Z, Huang H and
Hou L: Artemisinin co-delivery system based on manganese oxide for
precise diagnosis and treatment of breast cancer. Nanotechnology.
Apr 28–2021.(Epub ahead of print).
|
|
46
|
Yao X, Zhao CR, Yin H, Wang K and Gao JJ:
Synergistic antitumor activity of sorafenib and artesunate in
hepatocellular carcinoma cells. Acta Pharmacol Sin. 41:1609–1620.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Viallard C and Larrivée B: Tumor
angiogenesis and vascular normalization: Alternative therapeutic
targets. Angiogenesis. 20:409–426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li S, Xu HX, Wu CT, Wang WQ, Jin W, Gao
HL, Li H, Zhang SR, Xu JZ, Qi ZH, et al: Angiogenesis in pancreatic
cancer: Current research status and clinical implications.
Angiogenesis. 22:15–36. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cao J, Liu X, Yang Y, Wei B, Li Q, Mao G,
He Y, Li Y, Zheng L, Zhang Q, et al: Decylubiquinone suppresses
breast cancer growth and metastasis by inhibiting angiogenesis via
the ROS/p53/BAI1 signaling pathway. Angiogenesis. 23:325–338. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Singh N, Badrun D and Ghatage P: State of
the art and up-and-coming angiogenesis inhibitors for ovarian
cancer. Expert Opin Pharmacother. 21:1579–1590. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen H, Shi L, Yang X, Li S, Guo X and Pan
L: Artesunate inhibiting angiogenesis induced by human myeloma
RPMI8226 cells. Int J Hematol. 92:587–597. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Andrade-Tomaz M, de Souza I, Rocha CRR and
Gomes LR: The role of chaperone-mediated autophagy in cell cycle
control and its implications in cancer. Cells. 9:21402020.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chen K, Shou LM, Lin F, Duan WM, Wu MY,
Xie X, Xie YF, Li W and Tao M: Artesunate induces G2/M cell cycle
arrest through autophagy induction in breast cancer cells.
Anticancer Drugs. 25:652–662. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Weng X, Zhu SQ and Cui HJ: Artesunate
inhibits proliferation of glioblastoma cells by arresting cell
cycle. Zhongguo Zhong Yao Za Zhi. 43:772–778. 2018.(In Chinese).
PubMed/NCBI
|
|
55
|
Wang Y, Wei Z, Pan K, Li J and Chen Q: The
function and mechanism of ferroptosis in cancer. Apoptosis.
25:786–798. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu Z, Zhao Q, Zuo ZX, Yuan SQ, Yu K,
Zhang Q, Zhang X, Sheng H, Ju HQ, Cheng H, et al: Systematic
analysis of the aberrances and functional implications of
ferroptosis in cancer. iScience. 23:1013022020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li ZJ, Dai HQ, Huang XW, Feng J, Deng JH,
Wang ZX, Yang XM, Liu YJ, Wu Y, Chen PH, et al: Artesunate
synergizes with sorafenib to induce ferroptosis in hepatocellular
carcinoma. Acta Pharmacol Sin. 42:301–310. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hu LJ, Jiang T, Wang FJ, Huang SH, Cheng
XM and Jia YQ: Effects of artesunate combined with bortezomib on
apoptosis and autophagy of acute myeloid leukemia cells in vitro
and its mechanism. Zhonghua Xue Ye Xue Za Zhi. 40:204–208. 2019.(In
Chinese). PubMed/NCBI
|
|
59
|
Kim C, Lee JH, Kim SH, Sethi G and Ahn KS:
Artesunate suppresses tumor growth and induces apoptosis through
the modulation of multiple oncogenic cascades in a chronic myeloid
leukemia xenograft mouse model. Oncotarget. 6:4020–4035. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kumar B, Kalvala A, Chu S, Rosen S, Forman
SJ, Marcucci G, Chen CC and Pullarkat V: Antileukemic activity and
cellular effects of the antimalarial agent artesunate in acute
myeloid leukemia. Leuk Res. 59:124–135. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wang X, Du Q, Mao Z, Fan X, Hu B, Wang Z,
Chen Z, Jiang X, Wang Z, Lei N, et al: Combined treatment with
artesunate and bromocriptine has synergistic anticancer effects in
pituitary adenoma cell lines. Oncotarget. 8:45874–45887. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Karpel-Massler G, Westhoff MA, Kast RE,
Dwucet A, Nonnenmacher L, Wirtz CR, Debatin KM and Halatsch ME:
Artesunate enhances the antiproliferative effect of temozolomide on
U87MG and A172 glioblastoma cell lines. Anticancer Agents Med Chem.
14:313–318. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Berte N, Lokan S, Eich M, Kim E and Kaina
B: Artesunate enhances the therapeutic response of glioma cells to
temozolomide by inhibition of homologous recombination and
senescence. Oncotarget. 7:67235–67250. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lian S, Shi R, Huang X, Hu X, Song B, Bai
Y, Yang B, Dong J, Du Z, Zhang Y, et al: Artesunate attenuates
glioma proliferation, migration and invasion by affecting cellular
mechanical properties. Oncol Rep. 36:984–990. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Berdelle N, Nikolova T, Quiros S, Efferth
T and Kaina B: Artesunate induces oxidative DNA damage, sustained
DNA double-strand breaks, and the ATM/ATR damage response in cancer
cells. Mol Cancer Ther. 10:2224–2233. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Button RW, Lin F, Ercolano E, Vincent JH,
Hu B, Hanemann CO and Luo S: Artesunate induces necrotic cell death
in schwannoma cells. Cell Death Dis. 5:e14662014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wei S, Liu L, Chen Z, Yin W, Liu Y, Ouyang
Q, Zeng F, Nie Y and Chen T: Artesunate inhibits the mevalonate
pathway and promotes glioma cell senescence. J Cell Mol Med.
24:276–284. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Greenshields AL, Fernando W and Hoskin DW:
The anti-malarial drug artesunate causes cell cycle arrest and
apoptosis of triple-negative MDA-MB-468 and HER2-enriched SK-BR-3
breast cancer cells. Exp Mol Pathol. 107:10–22. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wen L, Liu L, Wen L, Yu T and Wei F:
Artesunate promotes G2/M cell cycle arrest in MCF7 breast cancer
cells through ATM activation. Breast Cancer. 25:681–686. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Greenshields AL, Shepherd TG and Hoskin
DW: Contribution of reactive oxygen species to ovarian cancer cell
growth arrest and killing by the anti-malarial drug artesunate. Mol
Carcinog. 56:75–93. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li B, Bu S, Sun J, Guo Y and Lai D:
Artemisinin derivatives inhibit epithelial ovarian cancer cells via
autophagy-mediated cell cycle arrest. Acta Biochim Biophys Sin
(Shanghai). 50:1227–1235. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Liu L, Zuo LF, Zuo J and Wang J:
Artesunate induces apoptosis and inhibits growth of Eca109 and
Ec9706 human esophageal cancer cell lines in vitro and in
vivo. Mol Med Rep. 12:1465–1472. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang L, Liu L, Chen Y, Du Y, Wang J and
Liu J: Correlation between adenosine triphosphate (ATP)-binding
cassette transporter G2 (ABCG2) and drug resistance of esophageal
cancer and reversal of drug resistance by artesunate. Pathol Res
Pract. 214:1467–1473. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wang L, Liu L, Wang J and Chen Y:
Inhibitory effect of artesunate on growth and apoptosis of gastric
cancer cells. Arch Med Res. 48:623–630. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhang P, Luo HS, Li M and Tan SY:
Artesunate inhibits the growth and induces apoptosis of human
gastric cancer cells by downregulating COX-2. Onco Targets Ther.
8:845–854. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Jiang F, Zhou JY, Zhang D, Liu MH and Chen
YG: Artesunate induces apoptosis and autophagy in HCT116 colon
cancer cells, and autophagy inhibition enhances the
artesunate-induced apoptosis. Int J Mol Med. 42:1295–1304.
2018.PubMed/NCBI
|
|
77
|
Kumar VL, Verma S and Das P: Artesunate
suppresses inflammation and oxidative stress in a rat model of
colorectal cancer. Drug Dev Res. 80:1089–1097. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ilamathi M, Santhosh S and
Sivaramakrishnan V: Artesunate as an anti-cancer agent targets
stat-3 and favorably suppresses hepatocellular carcinoma. Curr Top
Med Chem. 16:2453–2463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wojcicki AV, Kasowski MM, Sakamoto KM and
Lacayo N: Metabolomics in acute myeloid leukemia. Mol Genet Metab.
130:230–238. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Efferth T, Giaisi M, Merling A, Krammer PH
and Li-Weber M: Artesunate induces ROS-mediated apoptosis in
doxorubicin-resistant T leukemia cells. PLoS One. 2:e6932007.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wang Y, Yang J, Chen L, Wang J, Wang Y,
Luo J, Pan L and Zhang X: Artesunate induces apoptosis through
caspase-dependent and -independent mitochondrial pathways in human
myelodysplastic syndrome SKM-1 cells. Chem Biol Interact.
219:28–36. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Tan M, Rong Y, Su Q and Chen Y: Artesunate
induces apoptosis via inhibition of STAT3 in THP-1 cells. Leuk Res.
62:98–103. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Li Y, Feng L, Li Y, Jiang W, Shan N and
Wang X: Artesunate possesses anti-leukemia properties that can be
enhanced by arsenic trioxide. Leuk Lymphoma. 55:1366–1372. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Papanikolaou X, Johnson S, Garg T, Tian E,
Tytarenko R, Zhang Q, Stein C, Barlogie B, Epstein J and Heuck C:
Artesunate overcomes drug resistance in multiple myeloma by
inducing mitochondrial stress and non-caspase apoptosis.
Oncotarget. 5:4118–4128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Blessing MM and Alexandrescu S: Embryonal
tumors of the central nervous system: An update. Surg Pathol Clin.
13:235–247. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Alegría-Loyola MA, Galnares-Olalde JA and
Mercado M: Tumors of the central nervous system. Rev Med Inst Mex
Seguro Soc. 55:330–340. 2017.(In Spanish). PubMed/NCBI
|
|
87
|
Francis SS, Wang R, Enders C, Prado I,
Wiemels JL, Ma X and Metayer C: Socioeconomic status and childhood
central nervous system tumors in California. Cancer Causes Control.
32:27–39. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Reichert S, Reinboldt V, Hehlgans S,
Efferth T, Rödel C and Rödel F: A radiosensitizing effect of
artesunate in glioblastoma cells is associated with a diminished
expression of the inhibitor of apoptosis protein survivin.
Radiother Oncol. 103:394–401. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Kobayashi D, Hirayama M, Komohara Y,
Mizuguchi S, Wilson Morifuji M, Ihn H, Takeya M, Kuramochi A and
Araki N: Translationally controlled tumor protein is a novel
biological target for neurofibromatosis type 1-associated tumors. J
Biol Chem. 289:26314–26326. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Zhao J, Wen J, Wang S, Yao J, Liao L and
Dong J: Association between adipokines and thyroid carcinoma: A
meta-analysis of case-control studies. BMC Cancer. 20:7882020.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ma L and Fei H: Antimalarial drug
artesunate is effective against chemoresistant anaplastic thyroid
carcinoma via targeting mitochondrial metabolism. J Bioenerg
Biomembr. 52:123–130. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Capdevila J, Wirth LJ, Ernst T, Ponce Aix
S, Lin CC, Ramlau R, Butler MO, Delord JP, Gelderblom H, Ascierto
PA, et al: PD-1 blockade in anaplastic thyroid carcinoma. J Clin
Oncol. 38:2620–2627. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Breast Cancer Expert Committee of National
Cancer Quality Control Center; Breast Cancer Expert Committee of
China Anti-Cancer Association; Cancer Drug Clinical Research
Committee of China Anti-Cancer Association, . Guidelines for
clinical diagnosis and treatment of advanced breast cancer in China
(2020 edition). Zhonghua Zhong Liu Za Zhi. 42:781–797. 2020.(In
Chinese). PubMed/NCBI
|
|
94
|
Barzaman K, Karami J, Zarei Z,
Hosseinzadeh A, Kazemi MH, Moradi-Kalbolandi S, Safari E and
Farahmand L: Breast cancer: Biology, biomarkers, and treatments.
Int Immunopharmacol. 84:1065352020. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Britt KL, Cuzick J and Phillips KA: Key
steps for effective breast cancer prevention. Nat Rev Cancer.
20:417–436. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Dong X, Bai X, Ni J, Zhang H, Duan W,
Graham P and Li Y: Exosomes and breast cancer drug resistance. Cell
Death Dis. 11:9872020. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Tran TH, Nguyen TD, Poudel BK, Nguyen HT,
Kim JO, Yong CS and Nguyen CN: Development and evaluation of
artesunate-loaded chitosan-coated lipid nanocapsule as a potential
drug delivery system against breast cancer. AAPS PharmSciTech.
16:1307–1316. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Tran TH, Nguyen AN, Kim JO, Yong CS and
Nguyen CN: Enhancing activity of artesunate against breast cancer
cells via induced-apoptosis pathway by loading into lipid carriers.
Artif Cells Nanomed Biotechnol. 44:1979–1987. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhang S, Yuan H, Guo Y, Wang K, Wang X and
Guo Z: Towards rational design of RAD51-targeting prodrugs:
platinumIV-artesunate conjugates with enhanced
cytotoxicity against BRCA-proficient ovarian and breast cancer
cells. Chem Commun (Camb). 54:11717–11720. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Raza A, Ghoshal A, Chockalingam S and
Ghosh SS: Connexin-43 enhances tumor suppressing activity of
artesunate via gap junction-dependent as well as independent
pathways in human breast cancer cells. Sci Rep. 7:75802017.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Li Z, Zhu YT, Xiang M, Qiu JL, Luo SQ and
Lin F: Enhanced lysosomal function is critical for paclitaxel
resistance in cancer cells: Reversed by artesunate. Acta Pharmacol
Sin. 42:624–632. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Kujawa KA and Lisowska KM: Ovarian
cancer-from biology to clinic. Postepy Hig Med Dosw (Online).
69:1275–1290. 2015.(In Polish). View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Aziz NB, Mahmudunnabi RG, Umer M, Sharma
S, Rashid MA, Alhamhoom Y, Shim YB, Salomon C and Shiddiky MJA:
MicroRNAs in ovarian cancer and recent advances in the development
of microRNA-based biosensors. Analyst. 145:2038–2057. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Gaona-Luviano P, Medina-Gaona LA and
Magaña-Pérez K: Epidemiology of ovarian cancer. Chin Clin Oncol.
9:472020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Yang Y, Qi S, Shi C, Han X, Yu J, Zhang L,
Qin S and Gao Y: Identification of metastasis and
prognosis-associated genes for serous ovarian cancer. Biosci Rep.
40:BSR201943242020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Kuroki L and Guntupalli SR: Treatment of
epithelial ovarian cancer. BMJ. 371:m37732020. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Rooth C: Ovarian cancer: Risk factors,
treatment and management. Br J Nurs. 22:S23–S30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Penson RT, Valencia RV, Cibula D, Colombo
N, Leath CA III, Bidziński M, Kim JW, Nam JH, Madry R, Hernández C,
et al: Olaparib versus nonplatinum chemotherapy in patients with
platinum-sensitive relapsed ovarian cancer and a germline BRCA1/2
mutation (SOLO3): A randomized phase III trial. J Clin Oncol.
38:1164–1174. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Yadav G, Vashisht M, Yadav V and Shyam R:
Molecular biomarkers for early detection and prevention of ovarian
cancer-a gateway for good prognosis: A narrative review. Int J Prev
Med. 11:1352020. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Chen HH, Zhou HJ, Wu GD and Lou XE:
Inhibitory effects of artesunate on angiogenesis and on expressions
of vascular endothelial growth factor and VEGF receptor KDR/flk-1.
Pharmacology. 71:1–9. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Wang B, Hou D, Liu Q, Wu T, Guo H, Zhang
X, Zou Y, Liu Z, Liu J, Wei J, et al: Artesunate sensitizes ovarian
cancer cells to cisplatin by downregulating RAD51. Cancer Biol
Ther. 16:1548–1556. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Fatehi Hassanabad A, Chehade R, Breadner D
and Raphael J: Esophageal carcinoma: Towards targeted therapies.
Cell Oncol (Dordr). 43:195–209. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Fan J, Liu Z, Mao X, Tong X, Zhang T, Suo
C and Chen X: Global trends in the incidence and mortality of
esophageal cancer from 1990 to 2017. Cancer Med. 9:6875–6887. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Shi R, Cui H, Bi Y, Huang X, Song B, Cheng
C, Zhang L, Liu J, He C, Wang F, et al: Artesunate altered cellular
mechanical properties leading to deregulation of cell proliferation
and migration in esophageal squamous cell carcinoma. Oncol Lett.
9:2249–2255. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Eusebi LH, Telese A, Marasco G, Bazzoli F
and Zagari RM: Gastric cancer prevention strategies: A global
perspective. J Gastroenterol Hepatol. 35:1495–1502. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Niu PH, Zhao LL, Wu HL, Zhao DB and Chen
YT: Artificial intelligence in gastric cancer: Application and
future perspectives. World J Gastroenterol. 26:5408–5419. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Zhou X, Sun WJ, Wang WM, Chen K, Zheng JH,
Lu MD, Li PH and Zheng ZQ: Artesunate inhibits the growth of
gastric cancer cells through the mechanism of promoting oncosis
both in vitro and in vivo. Anticancer Drugs. 24:920–927. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Su T, Li F, Guan J, Liu L, Huang P, Wang
Y, Qi X, Liu Z, Lu L and Wang D: Artemisinin and its derivatives
prevent Helicobacter pylori-induced gastric carcinogenesis
via inhibition of NF-κB signaling. Phytomedicine. 63:1529682019.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
La Vecchia S and Sebastián C: Metabolic
pathways regulating colorectal cancer initiation and progression.
Semin Cell Dev Biol. 98:63–70. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Johdi NA and Sukor NF: Colorectal cancer
immunotherapy: Options and strategies. Front Immunol. 11:16242020.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Li LN, Zhang HD, Yuan SJ, Tian ZY, Wang L
and Sun ZX: Artesunate attenuates the growth of human colorectal
carcinoma and inhibits hyperactive Wnt/beta-catenin pathway. Int J
Cancer. 121:1360–1365. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Cui C, Feng H, Shi X, Wang Y, Feng Z, Liu
J, Han Z, Fu J, Fu Z and Tong H: Artesunate down-regulates
immunosuppression from colorectal cancer Colon26 and RKO cells in
vitro by decreasing transforming growth factor β1 and
interleukin-10. Int Immunopharmacol. 27:110–121. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Bei Y, Chen X, Raturi VP, Liu K, Ye S, Xu
Q and Lu M: Treatment patterns and outcomes change in early-stage
non-small cell lung cancer in octogenarians and older: A SEER
database analysis. Aging Clin Exp Res. 33:147–156. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Huo KG, D'Arcangelo E and Tsao MS:
Patient-derived cell line, xenograft and organoid models in lung
cancer therapy. Transl Lung Cancer Res. 9:2214–2232. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Xu K, Zhang C, Du T, Gabriel ANA, Wang X,
Li X, Sun L, Wang N, Jiang X and Zhang Y: Progress of exosomes in
the diagnosis and treatment of lung cancer. Biomed Pharmacother.
134:1111112021. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Rasheed SA, Efferth T, Asangani IA and
Allgayer H: First evidence that the antimalarial drug artesunate
inhibits invasion and in vivo metastasis in lung cancer by
targeting essential extracellular proteases. Int J Cancer.
127:1475–1485. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Zhao Y, Jiang W, Li B, Yao Q, Dong J, Cen
Y, Pan X, Li J, Zheng J, Pang X and Zhou H: Artesunate enhances
radiosensitivity of human non-small cell lung cancer A549 cells via
increasing NO production to induce cell cycle arrest at G2/M phase.
Int Immunopharmacol. 11:2039–2046. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Wang JS, Wang MJ, Lu X, Zhang J, Liu QX,
Zhou D, Dai JG and Zheng H: Artesunate inhibits
epithelial-mesenchymal transition in non-small-cell lung cancer
(NSCLC) cells by down-regulating the expression of BTBD7.
Bioengineered. 11:1197–1207. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Chen X, Han K, Chen F, Wu C and Huang W:
Effects of artesunate on the invasion of lung adenocarcinoma A549
cells and expression of ICAM-1 and MMP-9. Zhongguo Fei Ai Za Zhi.
16:567–571. 2013.(In Chinese). PubMed/NCBI
|
|
130
|
Tong Y, Liu Y, Zheng H, Zheng L, Liu W, Wu
J, Ou R, Zhang G, Li F, Hu M, et al: Artemisinin and its
derivatives can significantly inhibit lung tumorigenesis and tumor
metastasis through Wnt/β-catenin signaling. Oncotarget.
7:31413–31428. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Li W, Ma G, Deng Y, Wu Q, Wang Z and Zhou
Q: Artesunate exhibits synergistic anti-cancer effects with
cisplatin on lung cancer A549 cells by inhibiting MAPK pathway.
Gene. 766:1451342021. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Anwanwan D, Singh SK, Singh S, Saikam V
and Singh R: Challenges in liver cancer and possible treatment
approaches. Biochim Biophys Acta Rev Cancer. 1873:1883142020.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Cheng Z, Wei-Qi J and Jin D: New insights
on sorafenib resistance in liver cancer with correlation of
individualized therapy. Biochim Biophys Acta Rev Cancer.
1874:1883822020. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Hou J, Wang D, Zhang R and Wang H:
Experimental therapy of hepatoma with artemisinin and its
derivatives: In vitro and in vivo activity, chemosensitization, and
mechanisms of action. Clin Cancer Res. 14:5519–5530. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Jin M, Shen X, Zhao C, Qin X, Liu H, Huang
L, Qiu Z and Liu Y: In vivo study of effects of artesunate
nanoliposomes on human hepatocellular carcinoma xenografts in nude
mice. Drug Deliv. 20:127–133. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Guragain D, Seubwai W, Kobayashi D,
Silsinivanit A, Vaeteewoottacharn K, Sawanyawisuth K, Wongkham C,
Wongkham S, Araki N and Cha'on U: Artesunate and chloroquine induce
cytotoxic activity on cholangiocarcinoma cells via different cell
death mechanisms. Cell Mol Biol (Noisy-le-grand). 64:113–118. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Wang ZC, Liu Y, Wang H, Han QK and Lu C:
Research on the relationship between artesunate and raji cell
autophagy and apoptosis of burkitt's lymphoma and its mechanism.
Eur Rev Med Pharmacol Sci. 21:2238–2243. 2017.PubMed/NCBI
|
|
138
|
Chauhan AK, Min KJ and Kwon TK:
RIP1-dependent reactive oxygen species production executes
artesunate-induced cell death in renal carcinoma caki cells. Mol
Cell Biochem. 435:15–24. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Sarma B, Willmes C, Angerer L, Adam C,
Becker JC, Kervarrec T, Schrama D and Houben R: Artesunate affects
T antigen expression and survival of virus-positive merkel cell
carcinoma. Cancers (Basel). 12:9192020. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Zheng L and Pan J: The anti-malarial drug
artesunate blocks Wnt/β-catenin pathway and inhibits growth,
migration and invasion of uveal melanoma cells. Curr Cancer Drug
Targets. 18:988–998. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Wang Z, Wang C, Wu Z, Xue J, Shen B, Zuo
W, Wang Z and Wang SL: Artesunate suppresses the growth of
prostatic cancer cells through inhibiting androgen receptor. Biol
Pharm Bull. 40:479–485. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Yang Y, Wu N, Wu Y, Chen H, Qiu J, Qian X,
Zeng J, Chiu K, Gao Q and Zhuang J: Artesunate induces
mitochondria-mediated apoptosis of human retinoblastoma cells by
upregulating Kruppel-like factor 6. Cell Death Dis. 10:8622019.
View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Roh JL, Kim EH, Jang H and Shin D: Nrf2
inhibition reverses the resistance of cisplatin-resistant head and
neck cancer cells to artesunate-induced ferroptosis. Redox Biol.
11:254–262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Berköz M, Özkan-Yılmaz F, Özlüer-Hunt A,
Krośniak M, Türkmen Ö, Korkmaz D and Keskin S: Artesunate inhibits
melanoma progression in vitro via suppressing STAT3 signaling
pathway. Pharmacol Rep. 73:650–663. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Mancuso RI, Foglio MA and Olalla Saad ST:
Artemisinin-type drugs for the treatment of hematological
malignancies. Cancer Chemother Pharmacol. 87:1–22. 2021. View Article : Google Scholar : PubMed/NCBI
|