Introduction
Osteosarcoma (OS) accounts for 5% of tumors in
children and is the most common bone malignant tumor originating
from mesenchymal stem cells (1).
Furthermore, OS often occurs in teenagers or children under the age
of 20 and is characterized by rapid growth and early metastasis
(2). Recently, conventional
therapies, including neoadjuvant chemotherapy, surgery and
radiotherapy, have been used as treatment strategies for OS, and
the 5-year survival rate of patients with osteosarcoma has improved
from ~20% before the 1980s to currently ~70% (3,4);
However, half of the patients do not survive for longer than 10
year (5,6). In addition, the prognosis of OS remains
poor due to drug resistance, tumor recurrence and metastasis
(7). It is therefore crucial to
better understand the pathogenesis of OS to uncover potential
targets of tumor growth and metastasis regulation, which have
become essential for OS prevention and treatment in recent
years.
Previous studies have demonstrated that long
non-coding RNAs (lncRNAs) play significant roles in gene regulation
and genomic stability maintenance (8), and that they affect numerous cellular
processes, including cell proliferation, migration, invasion,
apoptosis and differentiation (9,10).
Numerous studies have reported that lncRNAs are involved in the
occurrence and development of various types of tumor (11–13). The
lncRNA C5orf66-antisense 1 (C5orf66-AS1), a recently discovered
lncRNA, has been reported to be associated with the pathogenesis of
different types of tumor. For example, Rui et al (14) demonstrated that C5orf66-AS1 can
promote cervical cancer cell proliferation via the microRNA
(miRNA/miR)-637/ring finger protein 1 axis. Lu et al
(15) demonstrated that C5orf66-AS1
prevents oral squamous cell carcinoma cell proliferation and
metastasis. However, the role and underlying mechanism of
C5orf66-AS1 in OS are yet to be elucidated.
It is well established that lncRNAs participate in
the pathogenesis of diseases via interaction with miRNAs (16). miRNAs represent another class of
endogenous small non-coding RNAs ~22 nucleotides in length, which
can regulate various cell processes, including cell proliferation,
apoptosis, invasion and differentiation (17,18). In
recent years, certain studies have confirmed that miRNAs serve a
key role in tumorigenesis, including in OS (19–21).
Previous studies reported that miR-149-5p plays an inhibitory role
in numerous types of cancer. For example, Luo et al
(22) confirmed that miR-149-5p can
regulate oral squamous cell carcinoma cell proliferation and
invasion by targeting transforming growth factor β2. Furthermore,
Ye and Chen (23) demonstrated that
miR-149-5p can inhibit medullary thyroid carcinoma cell
proliferation and invasion by targeting ARF GTPase-activating
protein 1. However, the expression and role of miR-149-5p in OS
remains unknown.
The present study aimed to identify the possible
roles of C5orf66-AS1 in OS cell proliferation and invasion and to
determine its underlying mechanisms.
Materials and methods
Clinical specimen collection
A total of 20 OS tissues and adjacent non-cancerous
tissues (2 cm from the tumor lesion) were collected from patients
(age range, 21–68 years; female to male ratio, 12:8) with OS who
underwent surgical treatment at the Huangshi Central Hospital
(Huangshi, China). All tissues were immediately frozen and stored
in liquid nitrogen or at −80°C. None of the patients received any
radiotherapy or chemotherapy prior to the surgery. The study
procedures were approved by the Ethics Committee of Huangshi
Central Hospital. Written informed consent was obtained from each
patient and all patients agreed to the use of their specimens in
this study.
Cell culture
The non-cancerous osteoblast hFOB1.19 cell line and
the human OS (HOS) cell lines MG63 and U2OS were purchased from the
American Type Culture Collection (ATCC). Cells were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 15% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin/streptomycin and placed at 37°C in a humidified
incubator containing 5% CO2. 293T cells (ATCC) were
cultured in DMEM supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C with 5% CO2.
Reverse transcription-quantitative
(RT-q)PCR
The expression levels of matrix metalloproteinase 9
(MMP-9), Bcl-2, Bax, miR-149-5p and C5orf66-AS1 were evaluated
using RT-qPCR. Total RNA was extracted from OS tissues, adjacent
non-cancerous tissues and HOS cell lines using an
TRIzol® Plus RNA Purification Kit (cat. no. 12183555;
Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Total RNA was reversed transcribed into
cDNA using the PrimeScript RT Reagent kit (Takara Biotechnology
Co., Ltd.) according to the manufacturer's protocol, and RT-qPCR
analysis was conducted using the SYBR PrimeScript RT-PCR kit
(Takara Biotechnology Co., Ltd.) using the ABI 7500 Real-Time PCR
system (Agilent Technologies, Inc.). The following thermocycling
conditions were used for qPCR: Initial denaturation at 95°C for 10
min, followed by 37 cycles of denaturation at 95°C for 15 sec,
annealing at 55°C for 40 sec and extension at 72°C for 34 sec. The
relative expression levels were normalized to endogenous control
GAPDH or U6 (24). The sequences of
the primers were as follows: GAPDH forward,
5′-CTTTGGTATCGTGGAAGGACTC-3′ and reverse,
5′-GTAGAGGCAGGGATGATGTTCT-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; miR-149-5p forward,
5′-CCCTCATTCTGTGCCACACTCCAGCTGGG-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; C5orf66-AS1 forward
5′-GGTCGGGCTTTTTCTTCCCA-3′ and reverse, 5′-GCCGCGGGAATGTCTTTATT-3′;
MMP-9 forward 5′-AGACCTGGGCAGATTCCAAAC-3′ and reverse,
5′-CGGCAAGTCTTCCGAGTAGT-3′; Bcl-2 forward
5′-AGTAAACCTGGACGGTGAAGTGATTG-3′ and reverse,
5′-CCAGGTAACAAAACCCCACA-3′; and Bax forward
5′-CAAGACCAGGGTGGTTGG-3′ and reverse, 5′-CACTCCCGCCACAAAGAT-3′.
Cell transfection
The control-small interfering (si)RNA,
C5orf66-AS1-siRNA, inhibitor control (5′-CAGUACUUUUGUGUAGUACAA-3′)
and miR-149-5p inhibitor (5′-GGGAGUGAAGACACGGAGCCAGA-3′) were
synthesized by Shanghai GenePharma Co., Ltd. U2OS cells were
transfected with 1 µM control-siRNA, 1 µM C5orf66-AS1-siRNA, 100 nM
inhibitor control, 100 nM miR-149-5p inhibitor, 1 µM
C5orf66-AS1-siRNA + 100 nM inhibitor control, or 1 µM
C5orf66-AS1-siRNA + 100 nM miR-149-5p inhibitor at 37°C for 48 h
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. At 48 h
following transfection, transfection efficiency was determined
using RT-qPCR analysis.
Dual-luciferase reporter assay
StarBase (http://starbase.sysu.edu.cn/) was used to identify the
relationship between C5orf66-AS1 and miR-149-5p. The
3′-untranslated region (UTR) of C5orf66-AS1 containing the target
sequence of miR-149-5p was amplified using RT-PCR and subsequently
inserted into a pmirGLO vector (Promega Corporation) to form the
reporter vector C5orf66-AS1-wild-type (C5orf66-AS1-WT). An
alternative expressing vector named C5orf66-AS1-mutant
(C5orf66-AS1-MUT) was also constructed by inserting the mutated
binding site into the pmirGLO vector (Promega Corporation).
C5orf66-AS1-WT or C5orf66-AS1-MUT and miR-149-5p mimic (sense,
5′-UCUGGCUCCGUGUCUUCACUCCC-3′; anti-sense,
5′-GAGUGAAGACACGGAGCCAGAUU-3′; Shanghai GenePharma Co., Ltd) or
mimic control (sense, 5′-UUCUCCGAACGUGUCACGUTT-3′; anti-sense,
5′-ACGUGACACGUUCGGAGAATT-3′; Shanghai GenePharma Co., Ltd.) were
co-transfected into 293T cells using Lipofectamine 3000 and
incubated for 48 h. The relative luciferase activity was detected
by Dual-Luciferase® Reporter assay system (Promega
Corporation) following the manufacturer's protocol. All firefly
luciferase activities were normalized to Renilla luciferase
activity.
Apoptosis evaluation by flow
cytometry
U2OS cells were transfected with control-siRNA,
C5orf66-AS1-siRNA, C5orf66-AS1-siRNA+inhibitor control, or
C5orf66-AS1- siRNA+miR-149-5p inhibitor at 37°C for 48 h, and U2OS
cell apoptosis was detected using a Annexin-V/PI Apoptosis
Detection kit (Beyotime Institute of Biotechnology). Briefly, 200
µl Annexin V-FITC and 10 µl PI were added into the U2OS cell
(106 cells) suspension for 30 min at 37°C in the dark
according to the manufacturer's protocol. The apoptotic cell rate
was determined using a flow cytometer (BD Biosciences) and
quantified using FlowJo software (version 7.2.4; FlowJo LLC).
Cell Counting Kit-8 (CCK-8) assay
CCK-8 assay (Beyotime Institute of Biotechnology)
was used to assess cell proliferation. After transfection, U2OS
cells (104 cells per well) were seeded in 96-well plates
overnight. CCK-8 reagent (10 µl) was added to each well and cells
were cultured at 37°C for 2 h. The optical density was measured at
a wavelength of 450 nm using a microplate reader following the
manufacturer's protocol.
Transwell migration and invasion
assay
U2OS cells were transfected with control-siRNA,
C5orf66-AS1-siRNA, C5orf66-AS1-siRNA+inhibitor control, or
C5orf66-AS1-siRNA+ miR-149-5p inhibitor at 37°C for 48 h. U2OS
cells (2×104 cells) were cultured in serum-free
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) and seeded
into the upper chamber of the Transwell (pore size, 8 µm; Corning,
Inc.). RPMI-1640 medium containing 10% FBS was added to the lower
chamber. Following incubation at 37°C in a 5% CO2
atmosphere for 48 h, cells remaining on the upper membrane were
scraped with cotton swabs, and the migrated and invaded cells on
the lower chamber were fixed with 4% paraformaldehyde at room
temperature for 30 min and stained with 0.1% crystal violet at room
temperature for 30 min. The migratory and invasive capacities of
the cells on the lower side of the membrane were visualized and
quantified using an inverted microscope (magnification, ×100; Nikon
Corporation). Transwell chambers were precoated with Matrigel (BD
Biosciences) at 37°C for 30 min for the invasion assay only.
Western blotting
U2OS cells were transfected with control-siRNA,
C5orf66-AS1-siRNA, C5orf66-AS1-siRNA+ inhibitor control, or
C5orf66-AS1-siRNA+miR-149-5p inhibitor at 37°C for 48 h. U2OS cells
were lysed using RIPA lysis buffer (Beyotime Institute of
Biotechnology) at 4°C. Proteins were quantified using a BCA Protein
Assay kit (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. Proteins (40 µg per lane) were
separated by 10% SDS-PAGE and transferred onto PVDF membranes.
After blocking with 5% skimmed milk in PBS-0.1% supplemented with
Tween-20 at room temperature for 1 h, membranes were incubated with
primary antibodies against GAPDH (cat. no. ab9485; 1:1,000; Abcam),
MMP-9 (cat. no. ab76003; 1:1,000; Abcam), Bcl-2 (cat. no. ab32124;
1:1,000; Abcam) and Bax (cat. no. ab182733; 1:1,000; Abcam)
overnight at 4°C. The membranes were washed with PBS-0.1%
supplemented with Tween-20 and incubated with a goat anti-rabbit
IgG H&L (HRP) pre-adsorbed antibody (cat. no. ab97080; 1:2,000;
Abcam) at room temperature for 1 h. Bands were detected using
enhanced chemiluminescence substrate (Pierce; Thermo Fisher
Scientific, Inc.) and semi-quantified using ImageJ software version
1.46 (National Institutes of Health).
Statistical analysis
Each experiment was performed three times. Data were
presented as the means ± standard deviation and analyzed using
GraphPad Prism 5.0 (GraphPad Software, Inc.). Differences among
groups were estimated using paired Student's t-test or one-way
ANOVA followed by Tukey's post hoc test. P<0.05 was considered
to indicate a statistically significant difference.
Results
C5orf66-AS1 is upregulated in OS
tissues and cells
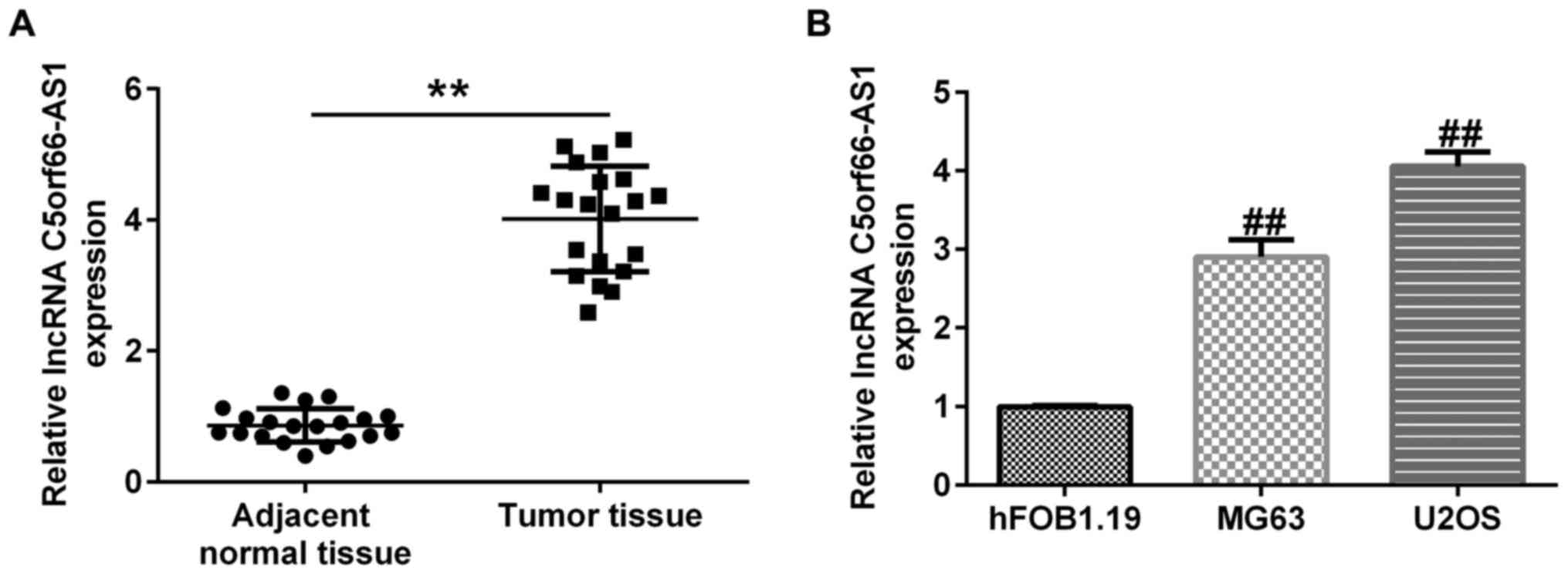
The expression of C5orf66-AS1 was evaluated in OS
tissues, adjacent non-cancerous tissues and HOS cells using
RT-qPCR. As presented in Fig. 1A,
C5orf66-AS1 was upregulated in OS tissues, compared with adjacent
normal tissues. Furthermore, a higher expression level of
C5orf66-AS1 was reported in U2OS and MG63 cells when compared with
hFOB1.19 cells (Fig. 1B). These
findings indicated that C5orf66-AS1 may be considered as a
regulator of OSCC tumorigenesis.
miR-149-5p directly interacts with
C5orf66-AS1
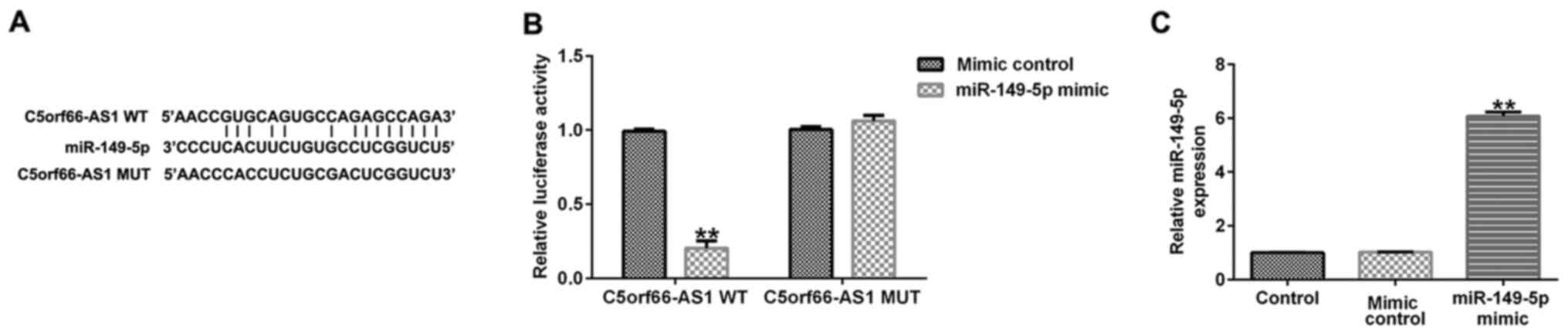
To investigate the potential mechanisms by which
C5orf66-AS1 may regulate the development of OS, StarBase software
was used to determine the target sites of C5orf66-AS1. The results
demonstrated that C5orf66-AS1 was a potential target of miR-149-5p
(Fig. 2A). In addition, the
luciferase reporter assay revealed that miR-149-5p mimic
significantly decreased the luciferase activity of the WT
C5orf66-AS1 3′-UTR construct, but had no significant effect on the
C5orf66-AS1 3′-UTR-MUT reporter (Fig.
2B). Furthermore, miR-149-5p mimic transfection increased
miR-149-5p expression in 293T cells, compared with the mimic
control group. These findings indicated that C5orf66-AS1 was a
direct target of miR-149-5p.
miR-149-5p is downregulated in OS
tissues and cells
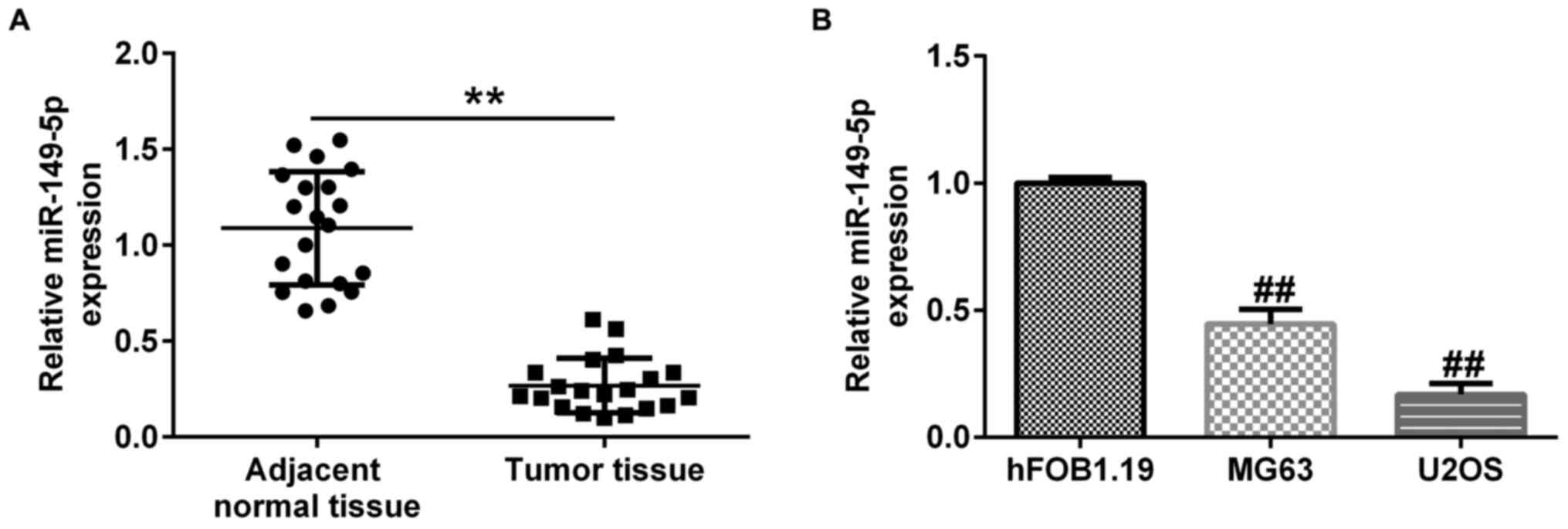
The expression of miR-149-5p in OS tissues and cells
was evaluated using RT-qPCR analysis. The results demonstrated that
miR-149-5p expression was significantly lower in OS tissues
compared with adjacent normal tissues (Fig. 3A). Furthermore, downregulation of
miR-149-5p was observed in MG63 and U2OS cells compared with
hFOB1.19 cells (Fig. 3B). All the
above results indicated that C5orf66-AS1 may regulate the
progression of OS by targeting miR-149-5p.
C5orf66-AS1 negatively regulates the
expression of miR-149-5p in OS cells
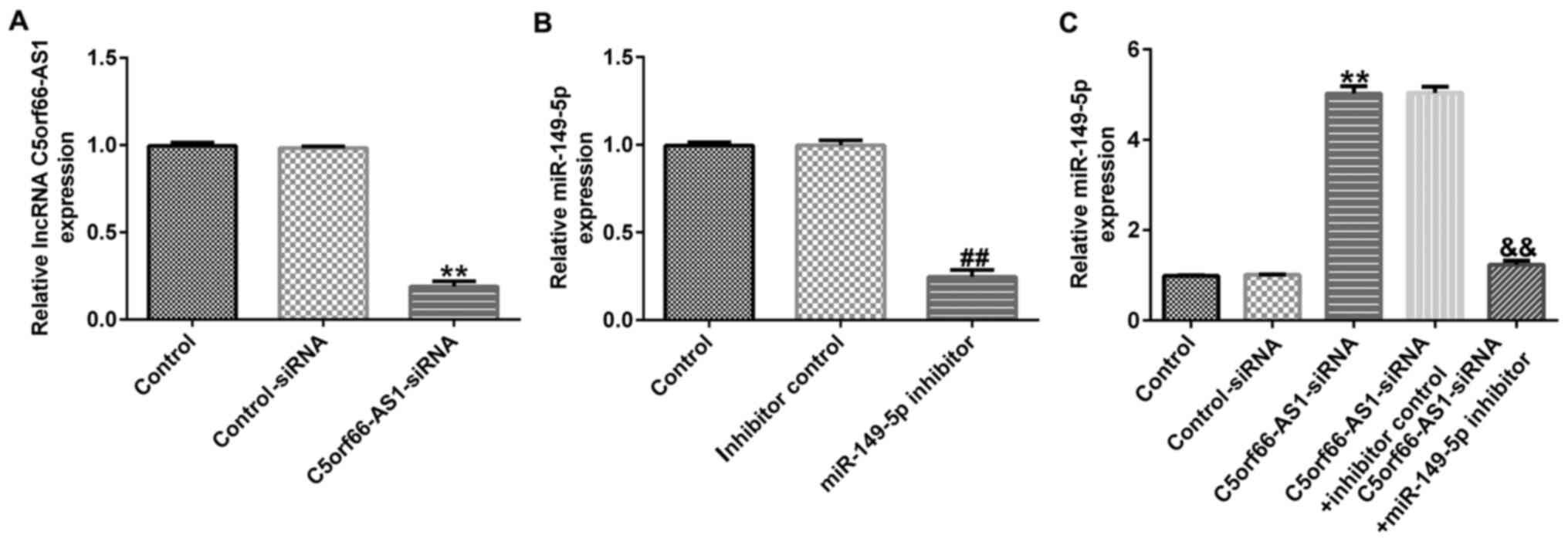
To further elucidate the regulatory correlation
between C5orf66-AS1 and miR-149-5p in OS, control-siRNA,
C5orf66-AS1-siRNA, inhibitor control or miR-149-5p inhibitor were
transfected into OS cells for 48 h. The results from RT-qPCR
demonstrated that C5orf66-AS1-siRNA inhibited C5orf66-AS1
expression in U2OS cells (Fig. 4A).
Furthermore, miR-149-5p expression was downregulated in miR-149-5p
inhibitor-transfected cells, compared with the inhibitor control
(Fig. 4B). Higher expression level
of miR-149-5p was also observed in C5orf66-AS1-siRNA transfected
cells compared with that in the control-siRNA group, whereas this
high expression level was significantly reversed following
transfection with miR-149-5p inhibitor (Fig. 4C). These findings suggested that
C5orf66-AS1 negatively regulated the expression of miR-149-5p in OS
cells.
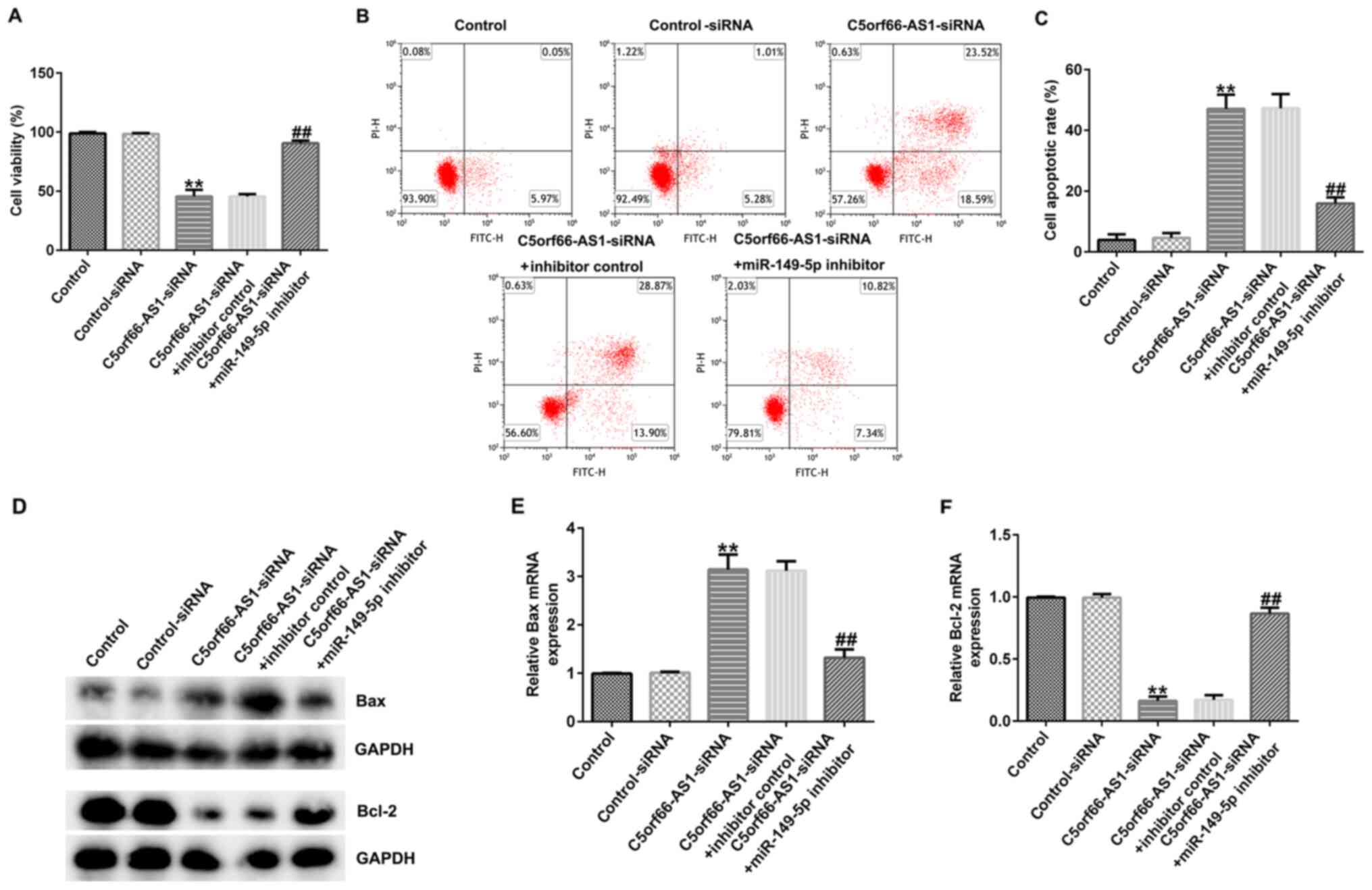
C5orf66-AS1-siRNA inhibits OS cell
viability and stimulates OS cell apoptosis by upregulating
miR-149-5p
The biological behaviors of U2OS cells co-regulated
by C5orf66-AS1 and miR-149-5p were explored. Control-siRNA,
C5orf66-AS1-siRNA, inhibitor control or miR-149-5p inhibitor were
transfected into U2OS cells for 48 h. The results from CCK-8 assay
demonstrated that C5orf66-AS1-siRNA decreased U2OS cell viability
(Fig. 5A). Furthermore, an increased
apoptotic rate of U2OS cells was observed in the C5orf66-AS1-siRNA
group compared with the control-siRNA group (Fig. 5B and C). In addition, the expression
levels of apoptosis-related proteins were detected. As presented in
Fig. 5D-F, C5orf66-AS1-siRNA
enhanced Bax protein and mRNA expression levels (Fig. 5D and E) and decreased Bcl-2 protein
and mRNA expression levels (Fig. 5D and
F) in U2OS cells compared with the control-siRNA group.
However, these observations were reversed following transfection
with miR-149-5p inhibitor. These results suggested that C5orf66-AS1
knockdown inhibited OS cell viability and promote OS cell apoptosis
by regulating miR-149-5p, which may inhibit the development of
OS.
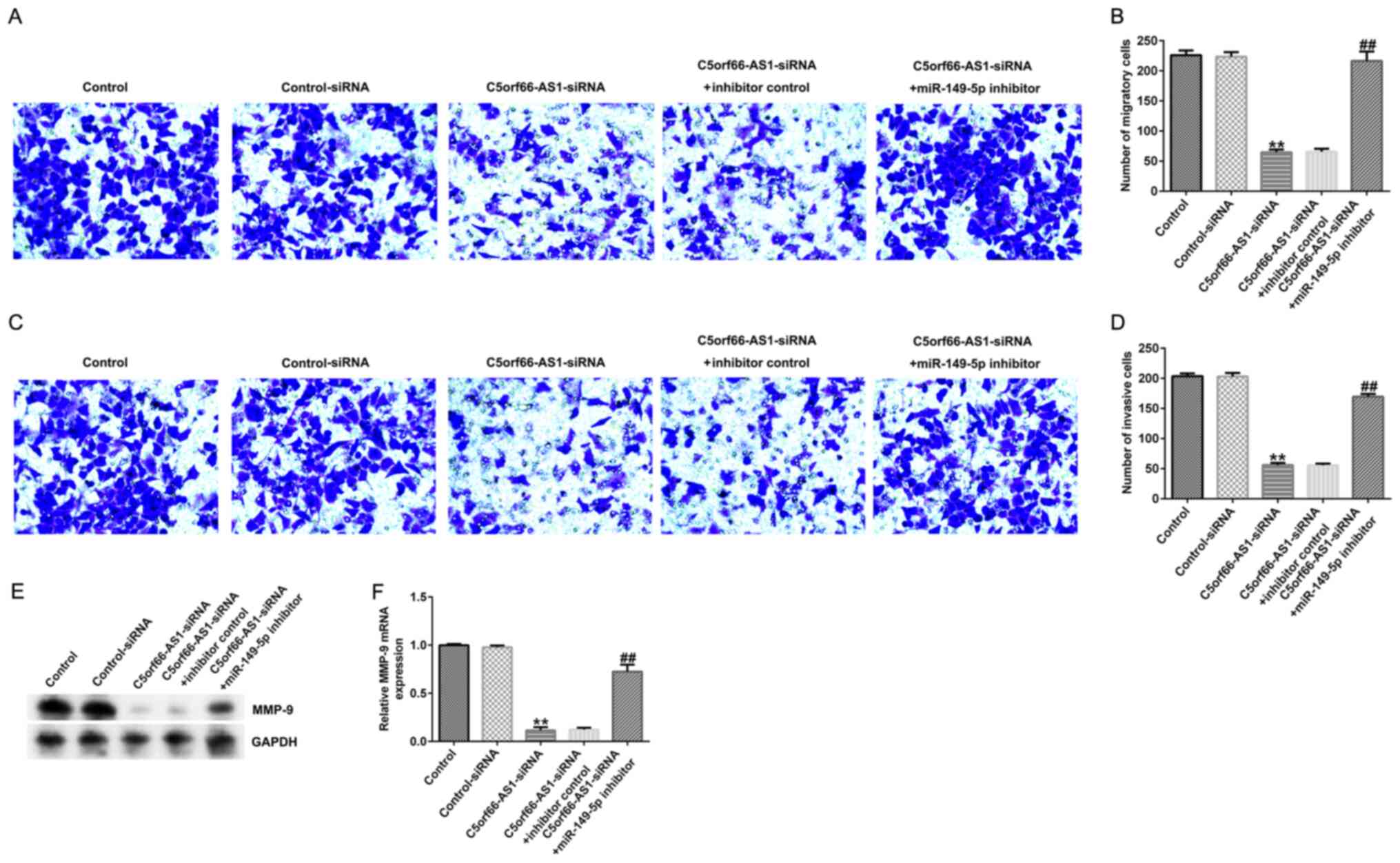
C5orf66-AS1-siRNA inhibits U2OS cell
migration and invasion via miR-149-5p upregulation
To further determine the role of C5orf66-AS1 in
regulating OS cell migration and invasion, Transwell assays were
used to evaluate the role of C5orf66-AS1 on U2OS cell migration and
invasion. The results demonstrated that C5orf66-AS1-siRNA decreased
U2OS cell migration (Fig. 6A and B)
and invasion (Fig. 6C and D)
compared with the control-siRNA group. Furthermore, the results
from western blotting and RT-qPCR demonstrated that MMP-9 was
downregulated in U2OS cells following C5orf66-AS1-siRNA
transfection (Fig. 6E and F). These
observations were reversed following cell transfection with
miR-149-5p inhibitor. These findings suggested that C5orf66-AS1
knockdown may inhibit OS cell proliferation, migration and invasion
via miR-149-5p upregulation.
Discussion
OS is the most common primary bone tumor in children
and adolescents and accounts for ~20% of all primary bone cancers
(1). OS is the second leading cause
of cancer-associated mortality in children (25,26). In
the recent years, multiple therapies have greatly improved the
treatment of OS, including radiopharmaceuticals (27), chemotherapy and radiotherapy
(28). However, the prognosis
remains poor for patients with OS. Thus, in-depth investigation
into the pathogenesis and underlying mechanisms of OS may provide
additional clinical evidence for the development of novel treatment
methods. The prognostic value of small molecules, including miRNAs
and lncRNAs have been demonstrated in OS. Huang et al
(29) confirmed that the lncRNA
small nucleolar RNA host gene 4 can promote OS proliferation and
migration by sponging miR-377-3p. Furthermore, Yang et al
(30) revealed that lncRNA-BC050642
could promote OS cell proliferation, stimulate colony formation and
inhibit cell apoptosis.
In the recent years, lncRNAs, which serve vital
roles in human diseases, have been widely investigated in
association with tumorigenesis and cancer progression. Recently,
Zhou et al (31) identified
C5orf66-AS1 as a potential biomarker for predicting early gastric
cancer and gastric carcinogenesis. Yu et al (32) reported that C5orf66-AS1 inhibits the
development and invasion of pituitary null cell adenomas. However,
the involvement of C5orf66-AS1 in the tumorigenesis of OS is yet to
be fully elucidated. The present study evaluated the expression of
C5orf66-AS1 in OS and adjacent non-cancerous tissues and in the HOS
cell lines MG63 and U2OS and the non-cancerous osteoblast cell line
hFOB1.19. C5orf66-AS1 was upregulated in OS tissues and HOS cell
lines, compared with the control groups. These findings indicated
that C5orf66-AS1 may be considered as a key factor in OS
tumorigenesis.
A previous study demonstrated that lncRNAs regulate
gene expression via an interaction with miRNAs in multiple cancer
types, including in OS (33). In the
present study, Starbase bioinformatics tool and a dual-luciferase
reporter assay predicted and further confirmed the target sites of
C5orf66-AS1. The results revealed that C5orf66-AS1 directly
interacts with miR-149-5p. In addition, miR-149-5p expression was
significantly higher in miR-149-5p mimic-transfected 293T cells
compared with the mimic control-transfected cells. miR-149-5p has
been found to act as a suppressor in various types of cancer
(34,35). In the present study, the expression
levels of miR-149-5p in 30 pairs of OS and adjacent non-cancerous
tissues, and in HOS and hFOB1.19 cell lines were determined using
RT-qPCR. Downregulation of miR-149-5p was observed in OS tissues
and HOS cell lines compared with the control groups. These results
suggested that C5orf66-AS1 may regulate the progression of OS by
targeting miR-149-5p.
To further illustrate the mechanism of C5orf66-AS1
in OS tumorigenesis, control-siRNA, C5orf66-AS1-siRNA, inhibitor
control or miR-149-5p inhibitor were transfected into U2OS cells
for 48 h. The results suggested that miR-149-5p expression was
downregulated in miR-149-5p inhibitor-transfected cells.
Furthermore, C5orf66-AS1 silencing increased miR-149-5p expression
and decreased C5orf66-AS1 expression levels, and miR-149-5p
expression increase was abolished by transfection with a miR-149-5p
inhibitor. Therefore, C5orf66-AS1 negatively regulated the
expression of miR-149-5p in OS cells.
A previous study reported that dysregulation of
oncogenes or tumor inhibitors is crucial in tumorigenesis and
cancer development (36). In
addition, previous studies demonstrated that lncRNAs regulate the
occurrence and development of numerous diseases and are closely
associated with tumor cell invasion and proliferation (37,38). The
effect of C5orf66-AS1 on HOS cell proliferation and apoptosis was
therefore investigated in the present study. Control-siRNA,
C5orf66-AS1-siRNA, inhibitor control or miR-149-5p inhibitor were
transfected into U2OS cells for 48 h. The results from the CCK-8
assay and flow cytometry indicated that C5orf66-AS1-siRNA inhibited
U2OS cell viability and increased U2OS cell apoptosis,
respectively, in the C5orf66-AS1-siRNA group, compared with the
control-siRNA group, thus demonstrating a potential tumor
growth-inhibiting effect. Apoptosis corresponds to the process of
cell self-destruction during certain physiological or pathological
conditions under the control of multiple genes (39). Bax and Bcl-2 are important mediators
of apoptosis and ultimately regulate apoptotic cell death (39). In the present study,
apoptosis-related gene expression was determined in U2OS cells
using RT-qPCR and western blot analysis. The results demonstrated
that C5orf66-AS1-siRNA decreased Bcl-2 expression and promoted Bax
expression compared with the control-siRNA group. However, all
these effects were reversed by the miR-149-5p inhibitor. These
findings suggested that C5orf66-AS1 downregulation regulated OS
cell viability and apoptosis by targeting miR-149-5p, which may
block the development of OS.
Migration and invasion are indicators of cancer
metastasis, including in OS. Zhang et al (40) reported that the lncRNA
differentiation antagonizing non-protein coding RNA promotes OS
migration and invasion via the miR-149/musashi RNA binding protein
2 axis. Chao et al (41) also
demonstrated the promotion of miR-552 in OS cell migration and
invasion (41). The effects of
C5orf66-AS1 on U2OS cell migration and invasion were therefore
investigated in the present study. The results demonstrated that
the migration and invasion of U2OS cells were inhibited by
C5orf66-AS1-siRNA, while this inhibition was abolished by the
miR-149-5p inhibitor. MMP-9, a vital proteolytic enzyme, has been
identified in cancer cells during malignant invasion and migration
(42). The present study
demonstrated that MMP-9 was downregulated in U2OS cells following
C5orf66-AS1-siRNA transfection, which was reversed by the
miR-149-5p inhibitor.
Taken together, the findings from the present study
suggested that C5orf66-AS1 knockdown had the potential to inhibit
OS cell proliferation and invasion by regulating miR-149-5p.
However, there were certain limitations to the current study.
Firstly, only one cell line (U2OS) was used to study the role of
C5orf66-AS1 in OS cells, and more OS cell lines should be used in
future experiments. Secondly, only one siRNA was used for
C5orf66-AS1 silencing; however, to prevent the off-target effect of
siRNA, two or more siRNAs for each gene should be used. In
addition, this study was just an in vitro preliminary study
on the role of C5orf66-AS1 in OS cells. To better understand the
role of C5orf66-AS1 in OS, further investigation is needed. For
example, the correlation between C5orf66-AS1 expression and the
clinicopathological characteristics of patients with OS should be
determined. In addition, in vivo investigation is required
to verify the function of C5orf66-AS1 in OS.
In summary, the results from the present study
indicated that C5orf66-AS1 expression was upregulated in OS.
C5orf66-AS1 silencing may prevent OS progression by inhibiting OS
cell proliferation and invasion via the regulation of miR-149-5p,
suggesting the anti-tumor function of C5orf66-AS1 in U2OS cells.
C5orf66-AS1 may therefore be considered as a potential therapeutic
target for the treatment of OS.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HZ contributed to the study design, data collection,
statistical analysis, data interpretation and manuscript
preparation. JS contributed to data collection, statistical
analysis and manuscript preparation. HZ and JS confirm the
authenticity of all the raw data. Both authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The study procedures were approved by the Ethics
Committee of Huangshi Central Hospital (Huangshi, China). Written
informed consent was obtained from each patient for the use of
their specimens in this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yap FHX, Amanuel B, Van Vliet C, Thomas M
and Wong D: Malignant transformation of fibrous dysplasia into
osteosarcoma confirmed with TP53 somatic mutation and mutational
analysis of GNAS gene. Pathology. 53:652–654. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Geary RL, Corrigan LR, Carney DN and
Higgins MJ: Osteosarcoma and second malignant neoplasms: A case
series. Ir J Med Sci. 188:1163–1167. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu ZJ, Tan JC, Qin X, Liu B and Yuan ZC:
Significance of circulating tumor cells in osteosarcoma patients
treated by neoadjuvant chemotherapy and surgery. Cancer Manag Res.
10:3333–3339. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hız M, Karaismailoglu B, Ulutas S,
Camurdan VB, Gorgun B and Oner Dincbas F: The effect of
preoperative radiotherapy on local control and prognosis in
high-grade non-metastatic intramedullary osteosarcoma of the
extremities. Arch Orthop Trauma Surg. 141:1083–1089. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang B and Zhang Y, Li R, Li J, Lu X and
Zhang Y: The efficacy and safety comparison of first-line
chemotherapeutic agents (high-dose methotrexate, doxorubicin,
cisplatin, and ifosfamide) for osteosarcoma: A network
meta-analysis. J Orthop Surg Res. 15:512020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rainusso N, Wang LL and Yustein JT: The
adolescent and young adult with cancer: State of the art - bone
tumors. Curr Oncol Rep. 15:296–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liao YX, Yu HY, Lv JY, Cai YR, Liu F, He
ZM and He SS: Targeting autophagy is a promising therapeutic
strategy to overcome chemoresistance and reduce metastasis in
osteosarcoma. Int J Oncol. 55:1213–1222. 2019.PubMed/NCBI
|
|
8
|
Zhou B, Li L, Qiu X, Wu J, Xu L and Shao
W: Long non-coding RNA ANRIL knockdown suppresses apoptosis and
pro-inflammatory cytokines while enhancing neurite outgrowth via
binding microRNA-125a in a cellular model of Alzheimer's disease.
Mol Med Rep. 22:1489–1497. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Long X, Li L, Zhou Q, Wang H, Zou D, Wang
D, Lou M and Nian W: Long non-coding RNA LSINCT5 promotes ovarian
cancer cell proliferation, migration and invasion by disrupting the
CXCL12/CXCR4 signalling axis. Oncol Lett. 15:7200–7206.
2018.PubMed/NCBI
|
|
10
|
Song B, Ye L, Wu S and Jing Z: Long
non-coding RNA MEG3 regulates CSE-induced apoptosis and
inflammation via regulating miR-218 in 16HBE cells. Biochem Biophys
Res Commun. 521:368–374. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang PS, Wang Z and Yang C: Dysregulations
of long non-coding RNAs - The emerging ‘lnc’ in environmental
carcinogenesis. Semin Cancer Biol. Apr 3–2021.(Epub ahead of
print). doi: 10.1016/j.semcancer.2021.03.029. View Article : Google Scholar
|
|
12
|
Bhan A, Soleimani M and Mandal SS: Long
Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rajagopal T, Talluri S, Akshaya RL and
Dunna NR: HOTAIR lncRNA: A novel oncogenic propellant in human
cancer. Clin Chim Acta. 503:1–18. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rui X, Xu Y, Jiang X, Ye W, Huang Y and
Jiang J: Long non-coding RNA C5orf66-AS1 promotes cell
proliferation in cervical cancer by targeting miR-637/RING1 axis.
Cell Death Dis. 9:11752018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu T, Liu H and You G: Long non-coding RNA
C5orf66-AS1 prevents oral squamous cell carcinoma through
inhibiting cell growth and metastasis. Int J Mol Med. 42:3291–3299.
2018.PubMed/NCBI
|
|
16
|
Tang XJ, Wang W and Hann SS: Interactions
among lncRNAs, miRNAs and mRNA in colorectal cancer. Biochimie.
163:58–72. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Shen S, Li Z, Li W and Weng X:
MIR-140-5p affects chondrocyte proliferation, apoptosis, and
inflammation by targeting HMGB1 in osteoarthritis. Inflamm Res.
69:63–73. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu WX, Liu Z, Deng F, Wang DD, Li XW, Tian
T, Zhang J and Tang JH: miR-145: A potential biomarker of cancer
migration and invasion. Am J Transl Res. 11:6739–6753.
2019.PubMed/NCBI
|
|
19
|
Gulino R, Forte S, Parenti R, Memeo L and
Gulisano M: MicroRNA and pediatric tumors: Future perspectives.
Acta Histochem. 117:339–354. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Llobat L and Gourbault O: Role of
microRNAs in human osteosarcoma: Future perspectives. Biomedicines.
9:4632021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Liu S, Shi J, Li J, Wang S, Liu H,
Zhao S, Duan K, Pan X and Yi Z: The Role of miRNA in the Diagnosis,
Prognosis, and Treatment of Osteosarcoma. Cancer Biother
Radiopharm. 34:605–613. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Luo K, He J, Yu D and Açil Y: miR-149-5p
regulates cisplatin chemosensitivity, cell growth, and metastasis
of oral squamous cell carcinoma cells by targeting TGFβ2. Int J
Clin Exp Pathol. 12:3728–3739. 2019.PubMed/NCBI
|
|
23
|
Ye X and Chen X: miR-149-5p inhibits cell
proliferation and invasion through targeting GIT1 in medullary
thyroid carcinoma. Oncol Lett. 17:372–378. 2019.PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang C, Ma K and Li WY: IL-6 promotes
cancer stemness and oncogenicity in U2OS and MG-63 osteosarcoma
cells by upregulating the OPN-STAT3 pathway. J Cancer.
10:6511–6525. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ozturk S, Gorgun C, Gokalp S, Vatansever S
and Sendemir A: Development and characterization of cancer stem
cell-based tumoroids as an osteosarcoma model. Biotechnol Bioeng.
117:2527–2539. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Anderson PM: Radiopharmaceuticals for
treatment of osteosarcoma. Adv Exp Med Biol. 1257:45–53. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Heng M, Gupta A, Chung PW, Healey JH,
Vaynrub M, Rose PS, Houdek MT, Lin PP, Bishop AJ, Hornicek FJ, et
al Japanese Musculoskeletal Oncology Group (JMOG); Soft Tissue
Osteosarcoma International Collaborative (STOIC), : The role of
chemotherapy and radiotherapy in localized extraskeletal
osteosarcoma. Eur J Cancer. 125:130–141. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang YF, Lu L, Shen HL and Lu XX: lncRNA
SNHG4 promotes osteosarcoma proliferation and migration by sponging
miR-377-3p. Mol Genet Genomic Med. 8:e13492020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang Y, Fei M, Zhou X, Li Y and Jin D: The
potential value of lncRNA-BC050642 in osteosarcoma origination and
outcomes. Artif Cells Nanomed Biotechnol. 47:1859–1866. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou Q, Li H, Jing J, Yuan Y and Sun L:
Evaluation of C5orf66-AS1 as a potential biomarker for predicting
early gastric cancer and its role in gastric carcinogenesis.
OncoTargets Ther. 13:2795–2805. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu G, Li C, Xie W, Wang Z, Gao H, Cao L,
Hao L and Zhang Y: Long non-coding RNA C5orf66-AS1 is downregulated
in pituitary null cell adenomas and is associated with their
invasiveness. Oncol Rep. 38:1140–1148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou Y, Yin L, Li H, Liu LH and Xiao T:
The lncRNA LINC00963 facilitates osteosarcoma proliferation and
invasion by suppressing miR-204-3p/FN1 axis. Cancer Biol Ther.
20:1141–1148. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiaobo Z, Qizhi H, Zhiping W and Tao D:
Down-regulated miR-149-5p contributes to preeclampsia via
modulating endoglin expression. Pregnancy Hypertens. 15:201–208.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu M, Xiao J, Chen M, Yuan L, Li J, Shen H
and Yao S: miR-149-5p promotes chemotherapeutic resistance in
ovarian cancer via the inactivation of the Hippo signaling pathway.
Int J Oncol. 52:815–827. 2018.PubMed/NCBI
|
|
36
|
Jiang J, Xie C, Liu Y, Shi Q and Chen Y:
Up-regulation of miR-383-5p suppresses proliferation and enhances
chemosensitivity in ovarian cancer cells by targeting TRIM27.
Biomed Pharmacother. 109:595–601. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Marín-Béjar O, Mas AM, González J,
Martinez D, Athie A, Morales X, Galduroz M, Raimondi I, Grossi E,
Guo S, Rouzaut A, Ulitsky I and Huarte M: The human lncRNA
LINC-PINT inhibits tumor cell invasion through a highly conserved
sequence element. Genome Biol. 18:1–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu Y, Yang Y, Li L, Liu Y, Geng P, Li G
and Song H: lncRNA SNHG1 enhances cell proliferation, migration,
and invasion in cervical cancer. Biochem Cell Biol. 96:38–43. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Russo A, Cardile V, Graziano ACE, Avola R,
Bruno M and Rigano D: Involvement of Bax and Bcl-2 in induction of
apoptosis by essential oils of three lebanese salvia species in
human prostate cancer cells. Int J Mol Sci. 19:E2922018. View Article : Google Scholar
|
|
40
|
Zhang W, Li JZ, Tai QY, Tang JJ, Huang YH
and Gao SB: lncRNA DANCR regulates osteosarcoma migration and
invasion by targeting miR-149/MSI2 axis. Eur Rev Med Pharmacol Sci.
24:6551–6560. 2020.PubMed/NCBI
|
|
41
|
Chao Y, Hu K, Wang X and Wang L:
MicroRNA-552 promotes migration and invasion of osteosarcoma
through targeting TIMP2. Biochem Biophys Res Commun. 511:63–68.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen T, Chen Z, Lian X, Wu W, Chu L, Zhang
S and Wang L: MUC 15 Promotes osteosarcoma cell proliferation,
migration and invasion through Livin, MMP-2/MMP-9 and Wnt/β-catenin
signal pathway. J Cancer. 12:467–473. 2021. View Article : Google Scholar : PubMed/NCBI
|