Introduction
Renal cell carcinoma (RCC; kidney cancer) is the
third most common malignant tumor in the genitourinary system
worldwide (1). There are ~338,000
new cases worldwide each year (2,3). Despite
advancements in the medical treatment for RCC in recent decades,
including the development anti-angiogenic agents, receptor targeted
therapy and immune checkpoint inhibition, the median overall
survival rate for patients is still not satisfactory (4). There is evidence that the 5-year
survival rate for patients with distant metastases is only 10%
(5). Due to its high levels of
resistance to chemotherapy and radiotherapy, radical surgery is
still the only therapy for localized renal cell cancer (6). Therefore, it is urgent to develop new
treatment targets and effective treatment methods.
Lycorine is a natural compound of
pyrrolo[de]phenanthridine ring-type alkaloid extracted from the
Amaryllidaceae genera (7). This
compound has various biological effects including anti-virus
(8), anti-malaria (9), anti-inflammation and anti-tumor
effects, with mild side effects (10,11).
Although the potential targets and mechanisms of action of lycorine
are still unclear, it is widely described as a candidate anti-tumor
drug, due to its high anti-tumor activity (12). Previous studies have shown that
lycorine has a selective cytotoxic effect on various cancer types
(13–16). Moreover, a previous study reported
that lycorine can target MEK2, induce autophagy-associated
apoptosis, and enhance vemurafenib activity in colorectal cancer.
However, hardly any research focused on its function in RCC
(17).
Ferroptosis is a form of programmed cell death
identified in recent years (18).
The main morphological changes associated with ferroptosis include
the shrinkage of mitochondria and mitochondrial ridge reduction
(18). Ferroptosis can be induced by
some specific small molecule compounds (such as erastin) and some
common clinical drugs (such as sorafenib and artemisinin) (19,20). The
formation of ferroptosis is mainly ascribed to initial lipid
peroxidation without the activation of caspases and the release of
reactive oxygen species (ROS) in cells, which is closely related to
the increase of iron ions in cells (21). Therefore, the entire ferroptosis
process can be impaired by the inhibitor of lipid peroxidation,
such as ferrostatin-1 (22).
Ferroptosis plays an important role in the occurrence of a variety
of tumors, including RCC (23).
Therefore, activating ferroptosis in tumors has great therapeutic
potential, which will provide new insight into the clinical
treatment of RCC. The present study aimed to investigate the
association between Lycorine and ferroptosis on the proliferation
of human renal cancer cells carcinoma (RCC) cells and its specific
anticancer mechanism.
Materials and methods
Cell culture and reagents
The human RCC cell lines, 786-O, A498 and Caki-1
cell lines were purchased from American Type Culture Collection,
the human normal renal proximal tubular cell line HK-2 was
purchased from Shanghai Bioleaf Biotech Co. Cells were cultured in
DMEM medium (HyClone; Cytiva) supplemented with 10% fetal calf
serum (Invitrogen; Thermo Fisher Scientific, Inc.) and
penicillin/streptomycin (100 mg/ml; Thermo Fisher Scientific, Inc.)
at 37°C in a 5% CO2-humidified incubator. Lycorine
(purity >98%) was purchased from MedChemExpress (cat. no.
HY-N0288). A 50 mM stock solution was prepared in dimethyl
sulfoxide (DMSO; Sigma-Aldrich; Merck KGaA), stored at −20°C and
diluted as needed in cell culture medium.
Cell viability assay
Cell viability was determined using MTS assays. RCC
or HK-2 cells (5×103 cells/well) were treated with
various concentrations of lycorine (0, 0.05, 0.1, 1, 5, 10, 20 and
50 µM) for 48 h, unless otherwise stated. The Aqueous One Solution
(Promega Corporation) was used for incubation for 120 min at 37°C
according to the manufacturer's instructions, and the absorption of
490 nm was measured. Three independent experiments were carried out
in triplicate.
Colony formation assay
Cells were seeded into a 6-well plate
(5×103 cells/well) and cultured for 24 h. Cells were
then treated with lycorine (10 µM). On day 8, colonies were fixed
in 3.7% paraformaldehyde for 15 min at 37°C, stained with 0.1%
crystal violet for 30 min at 37°C and counted manually.
Transmission electron microscopy
(TEM)
Cells were washed with PBS, then fixed in 2.5%
phosphate-buffered glutaraldehyde (Sigma-Aldrich; Merck KGaA) and
2% osmium tetroxide for 3 h at 4°C. The cells were then stained
with 2% uranyl acetate at 37°C for 12 h. After dehydration and
polymerization, samples were cut into 70–90 nm ultrathin sections
using an ultramicrotome (EMUC7, Leica Microsystems, Inc.). Cells
were infiltrated in methanol/Lowicryl K4M (Wuhan Biofavor Biotech
Services Co., Ltd.; in a series 2:1, 1:1 and 1:2) at −30°C for 2 h.
Images were captured using a transmission electron microscope (FEI;
Thermo Fisher Scientific, Inc.).
Mitochondrial membrane potential (MMP)
assay
The mitochondrial membrane potential of the cells
was investigated by tetramethylrhodamine methyl ester (TMRM) (AAT
Bioquest, Inc.) staining. Briefly, cells were seeded in confocal
dishes (1×105/well) and cultured for 48 h. The cells
were washed twice with PBS, then incubated for 20 min at 37°C with
100 nM TMRM to stain the mitochondria. TMRM intensity was
determined using a confocal fluorescence microscope.
Western blot analysis
Total protein was extracted from cells using the
M-PER™ Mammalian Protein Extraction reagent (Thermo Fisher
Scientific, Inc.). The protein concentration was determined using
BCA method. Protein samples were added to an equal volume of 2X SDS
loading buffer and boiled for 5 min. Proteins (30 µg per lane) were
separated via SDS-PAGE on a 10% gel, then transferred to a
polyvinylidene difluoride membrane (Beijing Zhongshan Jinqiao
Biotechnology Co., Ltd.). The membrane was blocked with 5% skimmed
milk (Beyotime Institute of Biotechnology) at room temperature for
2 h and incubated with primary antibodies against GPX4 (1:3,000;
cat. no. sc-166570; Santa Cruz Biotechnology, Inc.), ACSL4
(1:2,000; cat. no. sc-365230; Santa Cruz Biotechnology, Inc.) or
anti-β-actin (1:500; cat. no. SA00001-9; ProteinTech Group, Inc.)
overnight at 2–8°C. Following primary antibody incubation,
membranes were incubated with a horseradish peroxidase-conjugated
secondary antibody (1:1,000; cat. no. SA00001-9; ProteinTech Group,
Inc.) at room temperature for 3 h. Protein bands were observed
using an Amersham ECL Prime Western Blotting Detection Reagent (GE
Healthcare Life Sciences).
5-hydroxyeicosatetraenoic acid (HETE)
assay
5-HETE, a ferroptotic marker (24), was assessed using a 5-HETE ELISA kit
(cat. no. CED739Ge; Wuhan USCN Business Co., Ltd.), according to
the manufacturer's protocol.
12/15HETE assay
12-HETE and 15-HETE are two ferroptotic markers
(25). 12- or 15-HETE levels were
determined using 12- or 15-HETE ELISA kits (cat. nos. ab133034 and
ab133035; Abcam), according to the manufacturer's protocols. Each
96-well plate includes control, blank, standard and sample wells,
each in duplicate. First, 100 µl of the appropriate diluent and 50
µl assay buffer were added to all wells except blank wells, and 100
µl of the appropriate diluent was added to the standard wells.
Subsequently, 50 µl 12- or 15-HETE alkaline phosphatase conjugate
was added to NSB, standard and sample wells and 50 µl 12- or
15-HETE antibody was added to the B0, standard wells and sample
wells. The plates were incubated for 2 h on a 56 × g plate shaker
at room temperature, the contents of the wells were then washed
three times. Subsequently, 5 µl 12/15-HETE alkaline phosphatase
conjugate was added to all wells except blank wells and 200 µl pNpp
substrate solution was added to each well. Plates were incubated at
37°C for 3 h without shaking. The reaction was then stopped by
adding 50 µl of stop solution to each well. The absorbance was
measured at 405 nm using a microplate reader, and the formula
provided by the manufacturer was used to calculate the average net
absorbance measurement for each well, corresponding to the 12- or
15-HETE levels.
Determination of GSH (reduced
glutathione)/GSSG (oxidized glutathione) and malondialdehyde (MDA)
levels
The levels of GSH and MDA in cell extracts were
determined. The Reduced glutathione (GSH) was analyzed using the
Reduced Glutathione Assay kit (Nanjing Jiancheng Bioengineering
Institute; cat. no. A006), according to the manufacturer's
instructions. Total glutathione/oxidized glutathione (GSH/GSSG) was
determined using the Total Glutathione/Oxidized Glutathione Assay
kit (Nanjing Jiancheng Bioengineering Institute; cat. no. A061).
MDA levels were determined using a lipid peroxidation (MDA) test
kit (Biovision, Inc.; cat. no. K739-100) according to the
manufacturer's instructions.
Statistical analysis
All data are presented as the mean ± SD. Statistical
analysis was carried out using SPSS 22.0 software (IBM Corp.). The
test methods were described in figure legends. Graphs were
generated using Excel (Microsoft Corporation) or GraphPad Prism
5.00 for Windows (GraphPad Software, Inc.). All experiments were
repeated at least three times. P<0.05 was considered to indicate
a statistically significant difference.
Results
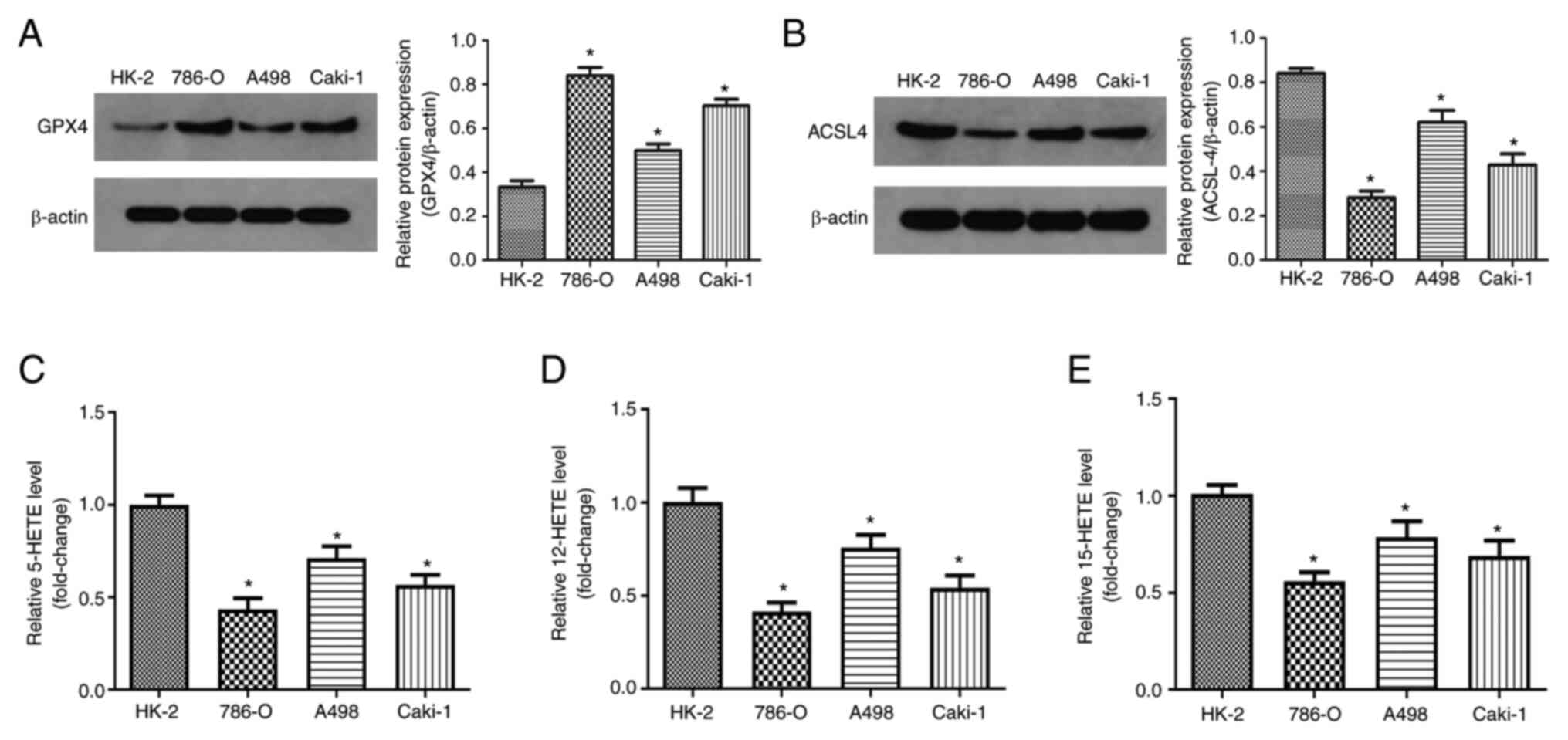
Determination of ferroptosis levels in
human RCC cells
Previous studies have indicated that RCC cells are
highly sensitive to ferroptosis (26). Therefore, in the present study,
ferroptosis levels were evaluated in human RCC cells. GPX4, 12-HETE
and 15-HETE participate in ferritin deposition as lipid peroxides.
These molecules have recently been recognized as direct markers of
ferroptosis (24). Thus, GPX4,
5-HETE, 12-HETE, 15-HETE were used as indicators of ferroptosis in
this study. A total of three typical malignant RCC cell lines
(786-O, A498 and Caki-1) and a human normal renal proximal tubule
cell line (HK-2) were used. Compared with the normal kidney cells
HK-2, the protein levels of GPX4 significantly increased in RCC
cells (Fig. 1A).
ACSL4 is considered to be the core regulatory gene
of ferroptosis (27). The expression
of ACSL4 in human RCC cells was significantly reduced compared with
normal human renal cells (Fig. 1B).
ELISA was used to detect 5-HETE, 12-HETE and 15-HETE levels. RCC
cells displayed. significantly lower levels of 5-HETE, 12-HETE and
15-HETE than normal cells (Fig.
1C-E). The decrease of ACSL4 expression may be related to
impaired ferroptosis in RCC cells. These results suggest that the
levels of ferroptosis in human RCC cells are reduced.
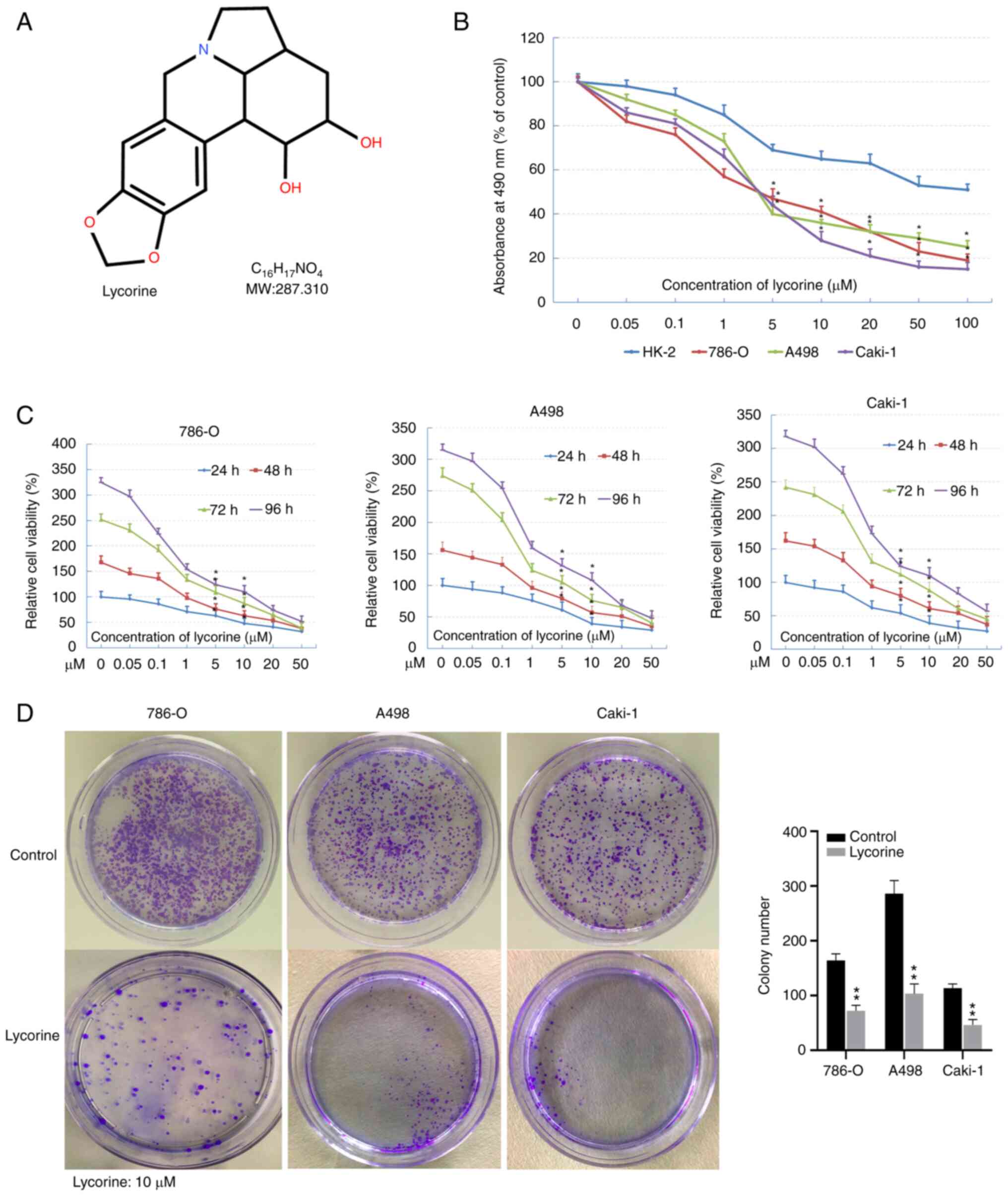
Lycorine inhibits the proliferation of
human RCC cells
The chemical structure of lycorine is shown in
Fig. 2A. MTS and colony formation
assays were used to study the anti-tumor activity of lycorine on
RCC. MTS assays indicated that lycorine significantly inhibited the
proliferation of RCC cells in a dose-dependent manner, with
IC50 values of 5–10 µM (Fig.
2B). In addition, the inhibitory effect of lycorine on the
three RCC cell lines was time-dependent (Fig. 2C). In the colony formation assay, 10
µM lycorine had a significant inhibitory effect on the colony
formation of the three RCC cell lines (Fig. 2D). These results indicated that
lycorine effectively inhibited the proliferation of RCC cells.
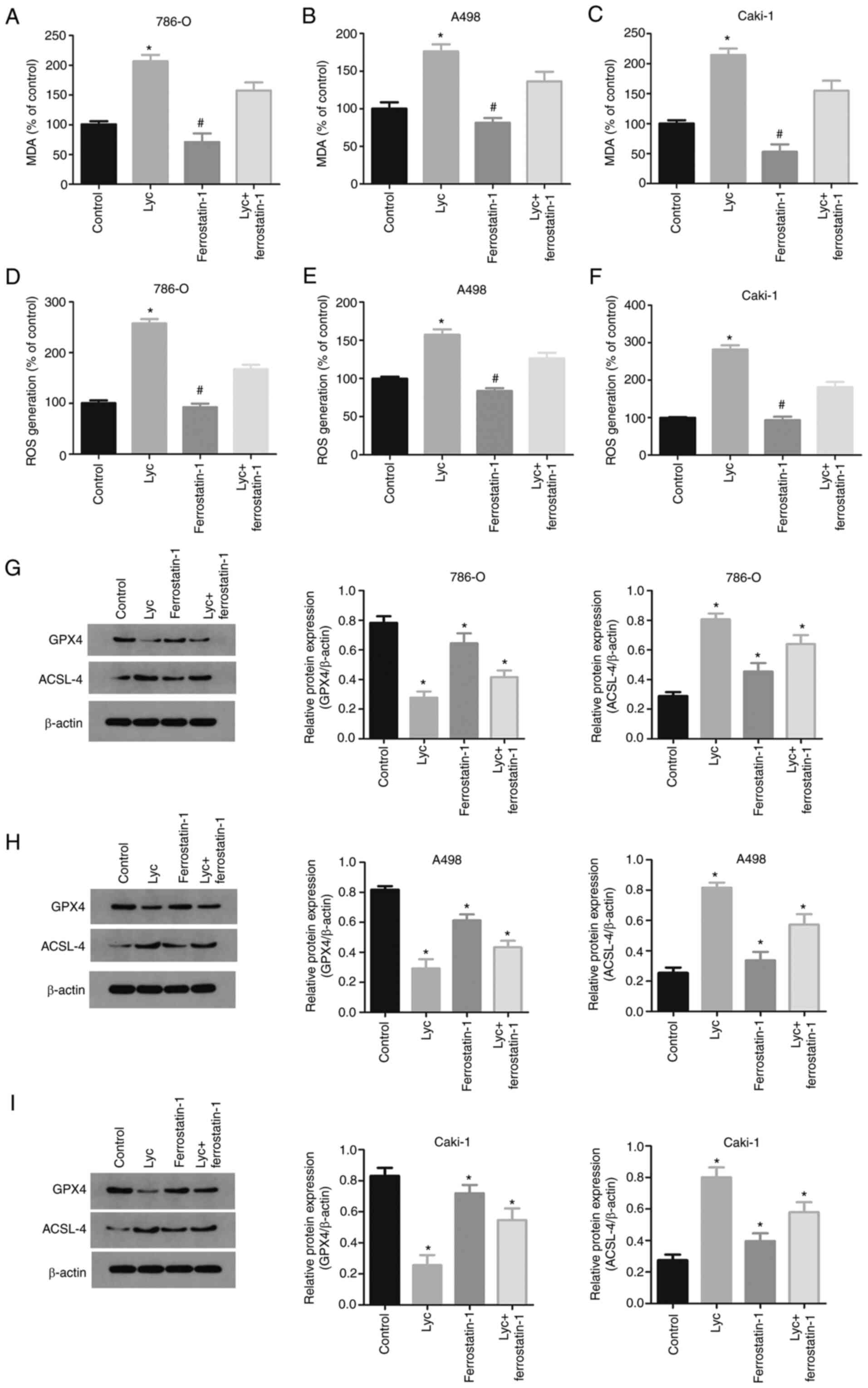
Lycorine induces ferroptosis in human
RCC cells by downregulating GPX4 and upregulating ACSL4
To determine whether the anti-tumor effect of
lycorine was related to ferroptosis, the lipid peroxidation levels
of lycorine were investigated in human RCC cells. Lipid
peroxidation is a key step during ferroptosis. MDA is a natural
by-product of lipid peroxidation and is commonly used as a marker
of lipid peroxidation (28). After
24 h of 10 µM lycorine treatment, the MDA levels and ROS generation
of the three RCC cell lines significantly increased compared with
the control group (Fig. 3A-F).
GSH and GSSG are important cellular antioxidant
systems that provide an inhibitory environment for the reduction of
oxidized substances. GPX4 is an important regulator of ferroptosis.
GPX4 deficiency causes a sharp increase in GSSG and a decrease in
the GSH/GSSG ratio. Moreover, loss of GPX4 activity can lead to
ferroptosis (29). In the present
study, after 24 h of treatment with lycorine, the expression of
GPX4 in RCC cells was significantly reduced, while the expression
of ACSL4 significantly increased (Fig.
3G-I). These results indicated that lycorine may induce
ferroptosis in human RCC cells.
The anti-tumor effect of lycorine on
RCC cells is blocked by ferroptosis inhibitors
Ferrostatin-1 is a specific inhibitor of ferroptosis
(30), which can reduce the
accumulation of intracellular ROS and cell death. It has been
reported that ferrostatin-1 can inhibit ferroptosis by
ferroptosis-inducers in vitro. In the present study,
ferrostatin-1 significantly reduced the lycorine-induced decrease
in GPX4 and increase in ACSL4 expression (Fig. 3G-I). Similarly, ferrostatin-1 reduced
the increase in MDA levels induced by lycorine (Fig. 3A-C).
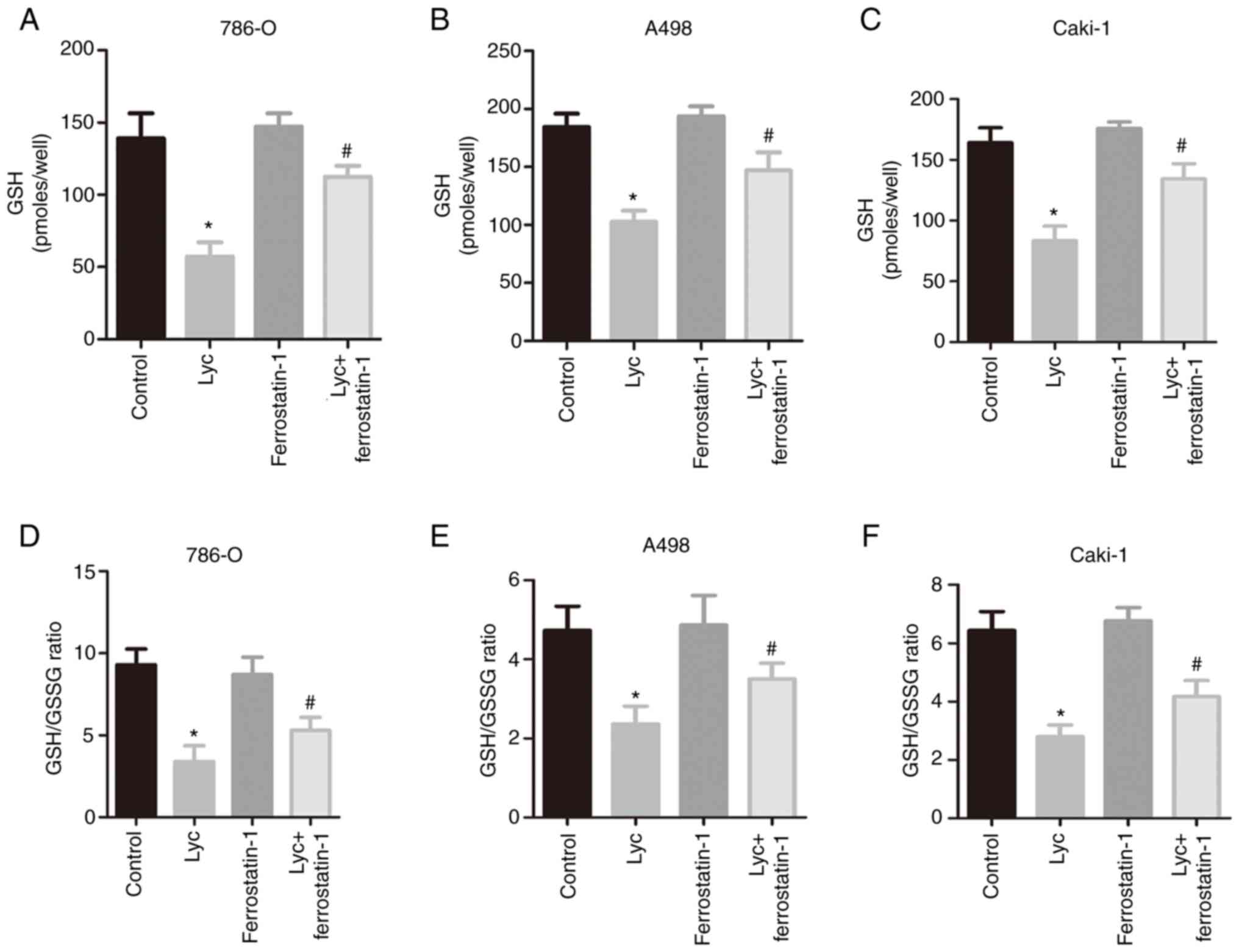
Furthermore, ferrostatin-1 abrogated the inhibitory
effect on GSH induced by Lycorine (Fig.
4A, C and E) and also reversed the changes in GSH/GSSG ratio
(Fig. 4B, D and F). In addition, the
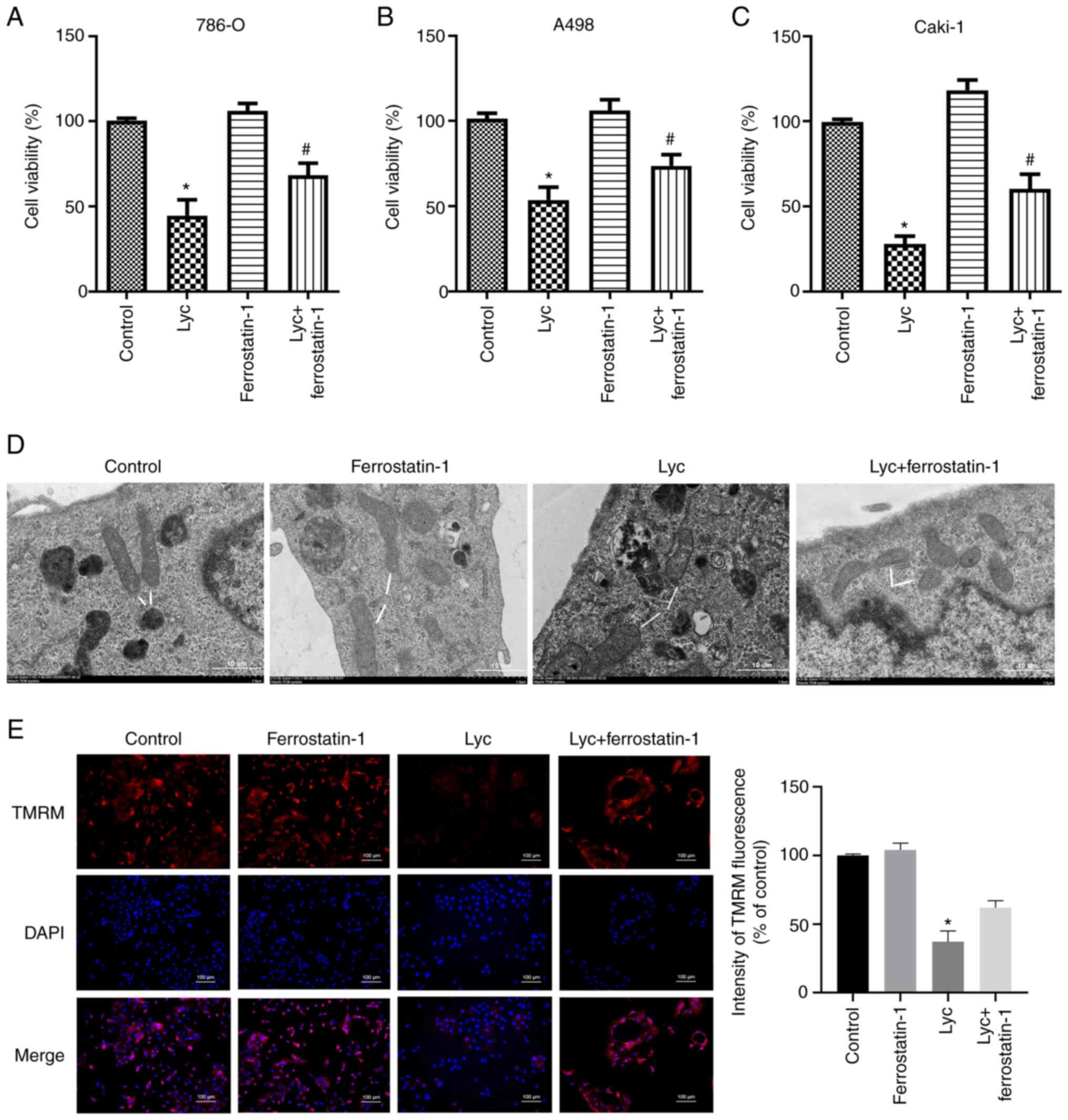
MTS assay was used to detect the effect of ferrostatin-1 on the
viability of human RCC cells. The results showed that ferrostatin-1
reversed the inhibitory effect of lycorine on the viability of
renal cancer cells (Fig. 5A-C).
Moreover, TEM was used to observe the morphological features
associated with ferroptosis. Clear mitochondrial shrinkage was
observed, together with increased membrane density in the lycorine
group compared with the control group. The morphological changes of
mitochondria were also ameliorated in the lycorine + ferrostatin-1
group (Fig. 5D). TMRM staining was
performed to evaluate the mitochondrial membrane potential
following lycorine treatment. TMRM levels in the lycorine group was
significantly reduced compared with the control group. However,
ferrostatin-1 abrogated this suppressive effect (Fig. 5E). Thus, ferrostatin-1 abrogated the
inhibitory effect of Lycorine on human RCC cells, suggesting that
its anti-tumor effect on RCC cells can be attributed to its
ferroptosis-inducing effect.
Discussion
Men with RCC are prone to peripheral invasion and
distant metastasis, leading to death (31). Although great progress has been made
in the treatment of RCC, treatment failure remains frequent
(32). The main findings of the
present study indicated that the anti-tumor activity of lycorine
against RCC was moderate at physiologically achievable
concentrations. These findings also demonstrated that RCC cells
were prone to ferroptosis. Moreover, administration of lycorine
inhibited the proliferation of RCC cells.
In recent years, Chinese herbal medicine have
gradually attracted researchers' attention. Lycorine is one of the
most commonly used Chinese herbal medicine. Lycorine has strong
antiviral activity, including against severe acute respiratory
syndrome associated coronavirus, poliovirus, enterovirus and
hepatitis C virus (33–35). The anti-tumor activity of lycorine
was first reported in 1976, when Jimenez et al (25) found that it inhibited protein
synthesis in eukaryotic cells. Subsequently, an increasing number
of studies identified lycorine as a potent tumor suppressor. For
example, Wang et al (26)
found that lycorine could target multiple myeloma stem cell-like
cells through inhibition of the Wnt/β-catenin pathway. Liu et
al (27) have investigated the
proapoptotic effect of lycorine in hepatoblastoma HepG2 cells.
These studies suggested that lycorine has an inhibitory effect on
different cancer types. However, few studies have focused on the
effect of lycorine on RCC. Therefore, the present study explored
the mechanism through which lycorine could inhibit the
proliferation and promote the apoptosis of human RCC cells. In the
present study, the proliferation of three RCC cell lines (786-O,
A498 and Caki-1) was evaluated following treatment with lycorine.
The results of MTS and colony formation assays suggested that
lycorine had a significant inhibitory effect on the proliferation
of human RCC cells. Its inhibitory effect was time- and
dose-dependent.
Ferroptosis is an iron-dependent cell death
progress. Recent studies have shown that Ferroptosis plays an
important role in tumorigenesis (36). A large number of studies have shown
that a variety of drugs can induce tumor cell ferroptosis at the
protein and gene levels and trigger changes in GPX4 and ACSL4
expression (37,38). Therefore, regulation of ferroptosis
in tumor cells holds great potential for tumor treatment. In
addition, the anti-tumor effect of lycorine is associated with
ferroptosis, as evidenced by the inhibition of GPX4 expression and
upregulation of ACSL4. The present study indicated that the
expression of GPX4 in RCC cells was significantly reduced, while
that of ACSL4 was increased following lycorine treatment. However,
these effects were reversed following treatment with the
ferroptosis inhibitor ferrostatin-1. These results suggest that the
anti-tumor effect of lycorine on RCC may be related to the
induction of ferroptosis. However, the present study lacks in
vivo experiments.
In conclusion, the present study confirmed that
lycorine could inhibit the proliferation in human RCC cells. The
anti-tumor effect of lycorine was associated with the induction of
ferroptosis. These findings indicated that lycorine could be used
as a potential candidate anti-tumor drug, which will shed new light
on the treatment against RCC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YD was responsible conception and design. HeCZ and
YJ contributed to the analysis of data and revised the manuscript.
HeCZ and YJ agreed to be accountable for all aspects of the work in
ensuring that questions related to the accuracy or integrity of any
part of the work are appropriately investigated and resolved. HeCZ
and HoCZ drafted and revised the manuscript, and confirm the
authenticity of all the raw data. HoCZ and LW carried out the
statistical analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
RCC
|
renal cell carcinoma
|
|
HETE
|
hydroxyeicosatetraenoic acid
|
|
GSH
|
reduced glutathione
|
|
GSSG
|
oxidized glutathione
|
|
MDA
|
malondialdehyde
|
|
GPX4
|
glutathione peroxidase 4
|
|
ACSL4
|
acyl-CoA synthetase long chain family
member 4
|
References
|
1
|
Marques-Magalhães Â, Graça I,
Miranda-Gonçalves V, Henrique R, Lopez M, Arimondo PB and Jerónimo
C: Anti-neoplastic and demethylating activity of a newly
synthetized flavanone-derived compound in renal cell carcinoma cell
lines. Biomed Pharmacother. 141:1116812021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Medina-Rico M, Ramos HL, Lobo M, Romo J
and Prada JG: Epidemiology of renal cancer in developing countries:
Review of the literature. Can Urol Assoc J. 12:E154–E162. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hsieh JJ, Purdue MP, Signoretti S, Swanton
C, Albiges L, Schmidinger M, Heng DY, Larkin J and Ficarra V: Renal
cell carcinoma. Nat Rev Dis Primers. 3:170092017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sathianathen NJ, Krishna S, Anderson JK,
Weight CJ, Gupta S, Konety BR and Griffith TS: The current status
of immunobased therapies for metastatic renal-cell carcinoma.
Immunotargets Ther. 6:83–93. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Argiris A, Harrington KJ, Tahara M,
Schulten J, Chomette P, Ferreira Castro A and Licitra L:
Evidence-based treatment options in recurrent and/or metastatic
squamous cell carcinoma of the head and neck. Front Oncol.
7:722017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lamoral-Theys D, Andolfi A, Van
Goietsenoven G, Cimmino A, Le Calvé B, Wauthoz N, Mégalizzi V, Gras
T, Bruyère C, Dubois J, et al: Lycorine, the main phenanthridine
Amaryllidaceae alkaloid, exhibits significant antitumor activity in
cancer cells that display resistance to proapoptotic stimuli: An
investigation of structure-activity relationship and mechanistic
insight. J Med Chem. 52:6244–6256. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hwang YC, Chu JJ, Yang PL, Chen W and
Yates MV: Rapid identification of inhibitors that interfere with
poliovirus replication using a cell-based assay. Antiviral Res.
77:232–236. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bendaif H, Melhaoui A, Ramdani M,
Elmsellem H, Douez C and El Ouadi Y: Antibacterial activity and
virtual screening by molecular docking of lycorine from Pancratium
foetidum Pom (Moroccan endemic Amaryllidaceae). Microb Pathog.
115:138–145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fürst R: Narciclasine-an Amaryllidaceae
alkaloid with potent antitumor and anti-inflammatory properties.
Planta Med. 82:1389–1394. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cole ER, de Andrade JP, Filho JFA, Schmitt
EFP, Alves-Araújo A, Bastida J, Endringer DC, de S Borges W and
Lacerda V: Cytotoxic and genotoxic activities of alkaloids from the
bulbs of griffinia gardneriana and habranthus itaobinus
(Amaryllidaceae). Anticancer Agents Med Chem. 19:707–717. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao L, Feng Y, Ge C, Xu X, Wang S, Li X,
Zhang K, Wang C, Dai F and Xie S: Identification of molecular
anti-metastasis mechanisms of lycorine in colorectal cancer by
RNA-seq analysis. Phytomedicine. 85:1535302021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu J, Hu WX, He LF, Ye M and Li Y:
Effects of lycorine on HL-60 cells via arresting cell cycle and
inducing apoptosis. FEBS Lett. 578:245–250. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li L, Dai HJ, Ye M, Wang SL, Xiao XJ,
Zheng J, Chen HY, Luo YH and Liu J: Lycorine induces cell-cycle
arrest in the G0/G1 phase in K562 cells via HDAC inhibition. Cancer
Cell Int. 12:492012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Roy M, Liang L, Xiao X, Peng Y, Luo Y,
Zhou W, Zhang J, Qiu L, Zhang S, Liu F, et al: Lycorine
downregulates HMGB1 to inhibit autophagy and enhances bortezomib
activity in multiple myeloma. Theranostics. 6:2209–2224. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu M, Peng S, He Y, Qin M, Cong X, Xing Y,
Liu M and Yi Z: Lycorine is a novel inhibitor of the growth and
metastasis of hormone-refractory prostate cancer. Oncotarget.
6:15348–15361. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu M, Yu Z, Mei P, Li J, Luo D, Zhang H,
Zhou M, Liang F and Chen R: Lycorine induces autophagy-associated
apoptosis by targeting MEK2 and enhances vemurafenib activity in
colorectal cancer. Aging (Albany NY). 12:138–155. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng J, Fan YQ, Liu BH, Zhou H, Wang JM
and Chen QX: ACSL4 suppresses glioma cells proliferation via
activating ferroptosis. Oncol Rep. 43:147–158. 2020.PubMed/NCBI
|
|
19
|
Ooko E, Saeed ME, Kadioglu O, Sarvi S,
Colak M, Elmasaoudi K, Janah R, Greten HJ and Efferth T:
Artemisinin derivatives induce iron-dependent cell death
(ferroptosis) in tumor cells. Phytomedicine. 22:1045–1054. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Houessinon A, François C, Sauzay C,
Louandre C, Mongelard G, Godin C, Bodeau S, Takahashi S, Saidak Z,
Gutierrez L, et al: Metallothionein-1 as a biomarker of altered
redox metabolism in hepatocellular carcinoma cells exposed to
sorafenib. Mol Cancer. 15:382016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qin Y, Qiao Y, Wang D, Tang C and Yan G:
Ferritinophagy and ferroptosis in cardiovascular disease:
Mechanisms and potential applications. Biomed Pharmacother.
141:111872Jul 7–2021.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu X, Yang Y, Xiang L and Zhang C: The
fate of melanocyte: Mechanisms of cell death in vitiligo. Pigment
Cell Melanoma Res. 34:256–267. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu H, Guo P, Xie X, Wang Y and Chen G:
Ferroptosis, a new form of cell death, and its relationships with
tumourous diseases. J Cell Mol Med. 21:648–657. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan H, Li X, Zhang X, Kang R and Tang D:
Identification of ACSL4 as a biomarker and contributor of
ferroptosis. Biochem Biophys Res Commun. 478:1338–1343. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jimenez A, Santos A, Alonso G and Vazquez
D: Inhibitors of protein synthesis in eukarytic cells. Comparative
effects of some amaryllidaceae alkaloids. Biochim Biophys Acta.
425:342–348. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang H, Gong Y, Liang L, Xiao L, Yi H, Ye
M, Roy M, Xia J, Zhou W, Yang C, et al: Lycorine targets multiple
myeloma stem cell-like cells by inhibition of Wnt/beta-catenin
pathway. Br J Haematol. 189:1151–1164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu J, Kang R and Tang D: Signaling
pathways and defense mechanisms of ferroptosis. FEBS J. Jun
6–2021.(Epub ahead of print). doi: 10.1111/febs.16059. View Article : Google Scholar
|
|
28
|
Ni J, Chen K, Zhang J and Zhang X:
Inhibition of GPX4 or mTOR overcomes resistance to lapatinib via
promoting ferroptosis in NSCLC cells. Biochem Biophys Res Commun.
567:154–160. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang J, Ma S, Xu R, Wei Y, Zhang J, Zuo T,
Wang Z, Deng H, Yang N and Shen Q: Smart biomimetic metal organic
frameworks based on ROS-ferroptosis-glycolysis regulation for
enhanced tumor chemo-immunotherapy. J Control Release. 334:21–33.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wenzel SE, Tyurina YY, Zhao J, St Croix
CM, Dar HH, Mao G, Tyurin VA, Anthonymuthu TS, Kapralov AA,
Amoscato AA, et al: PEBP1 wardens ferroptosis by enabling
lipoxygenase generation of lipid death signals. Cell.
171:628–641.e26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chatzizacharias NA, Rosich-Medina A,
Dajani K, Harper S, Huguet E, Liau SS, Praseedom RK and Jah A:
Surgical management of hepato-pancreatic metastasis from renal cell
carcinoma. World J Gastrointest Oncol. 9:70–77. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ko JJ, Xie W, Kroeger N, Lee JL, Rini BI,
Knox JJ, Bjarnason GA, Srinivas S, Pal SK, Yuasa T, et al: The
international metastatic renal cell carcinoma database consortium
model as a prognostic tool in patients with metastatic renal cell
carcinoma previously treated with first-line targeted therapy: A
population-based study. Lancet Oncol. 16:293–300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Roy M, Liang L, Xiao X, Feng P, Ye M and
Liu J: Lycorine: A prospective natural lead for anticancer drug
discovery. Biomed Pharmacother. 107:615–624. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen D, Cai J, Yin J, Jiang J, Jing C, Zhu
Y, Cheng J, Di Y, Zhang Y, Cao M, et al: Lycorine-derived
phenanthridine downregulators of host Hsc70 as potential hepatitis
C virus inhibitors. Future Med Chem. 7:561–570. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li SY, Chen C, Zhang HQ, Guo HY, Wang H,
Wang L, Zhang X, Hua SN, Yu J, Xiao PG, et al: Identification of
natural compounds with antiviral activities against SARS-associated
coronavirus. Antiviral Res. 67:18–23. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guo J, Xu B, Han Q, Zhou H, Xia Y, Gong C,
Dai X, Li Z and Wu G: Ferroptosis: A novel anti-tumor action for
cisplatin. Cancer Res Treat. 50:445–460. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Belavgeni A, Bornstein SR and Linkermann
A: Prominin-2 suppresses ferroptosis sensitivity. Dev Cell.
51:548–549. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu N, Lin X and Huang C: Activation of
the reverse transsulfuration pathway through NRF2/CBS confers
erastin-induced ferroptosis resistance. Br J Cancer. 122:279–292.
2020. View Article : Google Scholar : PubMed/NCBI
|