Introduction
Cervical cancer ranks second of all gynecological malignancies with high morbidity and mortality worldwide (1–3). In China, there are >150,000 newly diagnosed cases and >20,000 deaths annually (4). Cervical squamous cell carcinoma accounts for 80–85% of all cervical cancers (5). Molecular and epidemiological studies suggest that multiple genes are involved in the transformation from normal cervical epithelial cells to cervical intraepithelial neoplasia (CIN), and ultimate progression to invasive cervical squamous cell carcinoma (6,7). Therefore, it is very important to clarify the molecular mechanisms to determine prevention and treatment strategies, early diagnostic biomarkers, and therapeutic targets of cervical squamous cell carcinoma.
The relationship between chronic inflammation and cancer has always been a concern. Chronic inflammation is the major cause of prostate cancer (8), cervical cancer (9), and ovarian cancer (10). Some studies have shown that infections trigger a range of inflammatory responses, and inflammation promotes cancer progression (11). The effects of inflammation on tumorigenesis include regulation of the tumor microenvironment through the production of inflammatory cytokines, accumulation of reactive oxygen species (ROS), and activation of transcription factors (12). Lipopolysaccharide (LPS) is a gram-negative bacterial antigen that causes significant systemic inflammation and participates in the progression and metastasis of a variety of tumors (13). It is well known that LPS can promote the progression and metastasis of cervical cancer by stimulating the release of interleukin-1β (IL-1β); thus, promoting downstream cytokines and pyroptosis (14). IL-1β, a generally accepted product of pyroptotic cell death and a crucial cytokine in tumor occurrence and development, has been chosen as a target for cancer immunotherapy (15). However, the underlying mechanism of LPS in cervical carcinoma is not clear.
Forkhead box O1 (FOXO1), a member of the Forkhead box O (FoxO) family, was found to be down-regulated in cervical cancer, and it was considered as a tumor suppressor and negatively modulator of the PI3K/Akt signaling pathway (16). Activation of FOXO1 results in cell proliferation, pyroptosis, autophagy, and DNA repair (17). Therefore this study explored whether FOXO1 expression was altered after LPS treatment in cervical squamous carcinoma SiHa cells. The role of FOXO1 in mitochondrial function in SiHa cells was checked. Finally, it was tested whether overexpression of FOXO1 could reverse the effects of LPS in SiHa cells. Overall, we confirmed the possibility that FOXO1 could serve as a target for the prevention and treatment of human cervical squamous cell carcinoma.
Materials and methods
Chemicals and reagents
Cervical squamous carcinoma SiHa cells were obtained from the American Type Culture Collection (ATCC). Dulbecco's modified Eagle's medium (DMEM) powder, fetal bovine serum (FBS), and L-glutamine were purchased from Gibco; Thermo Fisher Scientific, Inc. The Lipofectamine® 2000 transfection reagent was obtained from Thermo Fisher Scientific, Inc. Dimethyl sulfoxide (DMSO) and LPS were obtained from Sigma-Aldrich; Merck KGaA. FOXO1 small interfering RNA (siRNA) was designed and synthesized by Wuhan Genesil Biotechnology Co., Ltd. FOXO1 and actin gene primers were designed and synthesized by Shanghai Boya Biotechnology Co., Ltd.
Cell culture and LPS treatment
SiHa cells were maintained in DMEM containing 10% FBS, 100 U/ml penicillin, 100 µg/ml streptomycin, and 2 mM L-glutamine in a 5% CO2 incubator at 37°C. The cells were cultured to 80–90% confluence and then treated with 40 µg/ml LPS for 24 h or other reagents at the indicated times and concentrations. The choice of LPS concentrations was based on previous reports (18).
Cloning and transfection of the FOXO1 vector plasmids
The pcDNA3.1-FOXO1 plasmid was synthesized by Hangzhou Hibio Biotech Co., Ltd. (Hangzhou, Zhejiang, China). FOXO1: Primer-F: 5′-GCGGGCTGGAAGAATTCAAT-3′ and Primer-R: 5′-TCCAGTTCCTTCATTCTGCA-3′. The resulting pcDNA3.1-FOXO1 plasmids or pcDNA3.1 plasmids were then transfected into SiHa cells using Lipofectamine 2000. Briefly, the SiHa cells were added with serum-free DMEM for 24 h; then 2.5 µg pcDNA3.1-FOXO1 plasmid and 8 µl of Lipofectamine 2000 were diluted in 750 µl of OptiMEM (Life Technologies), and incubated for 45 min and then overlaid onto SiHa cells for 6 h at 37°C. Finally, cell medium containing 20% fetal bovine serum was added for further culture for 48 h.
FoxO1 siRNA-expressing plasmid construction
The FOXO1 siRNA plasmid was constructed using the primers 5′-CCCAAGGCTTTGGTCCTATC-3′ (forward) and 5′-GCCGGATTCACTGTATTCTTG-3′ (reverse). Negative siRNAs were sequenced (an unrelated gene) as follows: 5′-GUACCGCACGUCAUUCGUAUC-3′ (forward) and 5′-UACGAAUGACGUGCGGUACGU-3′ (reverse). The reconstituted FOXO1 siRNA or negative siRNA expression plasmid was transfected into SiHa cells, along with or without LPS (40 µg/ml) treatment. Mitochondrial function and biological function of SiHa cells were measured.
Reverse transcription-quantitative PCR (qPCR)
Extraction of total RNA from SiHa cells was performed using RNAiso Plus (Takara). Then, 1 µg of RNA was reverse transcribed into complementary DNA (cDNA) by a RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Inc.). Specific primers used in this experiment were as follows: FOXO1 sense, 5′-GCGGGCTGGAAGAATTCAAT-3′ and antisense, 5′-TCCAGTTCCTTCATTCTGCA-3′; β-actin sense, 5′-CGAGCGGGAAATCGTGCGTGACAT-3′; and antisense, 5′-CGTCATACTCCTGCTTGCTGATCCACATCT-3′. Real-time qPCR was performed on a StepOne real-time PCR System (Applied Biosystems). The relative level of the FOXO1 mRNA was calculated using the threshold cycle (2−ΔΔCT) method (19).
Western blot analysis
SiHa cells were thawed using lysis buffer containing 0.5% deoxycholate, 50 mM Tris-HCl (pH 7.0), 0.1% Triton X-100, 150 mM sodium chloride (NaCl), 0.1% sodium dodecyl sulfate (SDS), and 2 mM ethylene diamine tetraacetic acid (EDTA). Total proteins were electrophoresed using a 10–15% gradient SDS-polyacrylamide gel and subsequently transferred onto polyvinylidene fluoride (PVDF) membranes. After blocking in 5% nonfat milk in Tris-buffered saline for 2 h, the PVDF membranes were probed with primary antibodies specific to FOXO1 (1:1,000 dilution; Abcam: ab52857), Caspase-1 (1:500 dilution; Santa Cruz Biotechnology, Inc.), and β-actin (1:2,000; ab8227; Abcam). The membranes were washed and incubated with the horseradish peroxidase (HRP)-conjugated secondary antibody (1:4,000) for 1 h at room temperature. The proteins were visualized by the ECL Chemiluminescence Western Detection System (Cell Signaling Technology).
Immunohistochemistry
SiHa cells were digested by trypsin and centrifuged at 1,000 × g for 10 min at 4°C. The cell masses were cut into 5 µm slices and then treated with 4% H2O2 for 30 min. Antigen retrieval was performed in 0.01 M citrate buffer for 25 min. The slides were incubated with FOXO1 antibodies (dilution, 1:100; Santa Cruz Biotechnology, Inc.) for 30 min at 37°C and a 3,3′-diaminobenzidine Sigma-D8001 staining kit (Sigma-Aldrich; Merck KGaA). Positive (brown) staining indicated the presence of the FOXO1 protein, as detected by light microscopy (magnification, ×200).
Enzyme-linked immunosorbent assay (ELISA) analysis
The production of IL-1β was detected by an ELISA kit (R&D System). Two hundred microliters of the supernatant were added and incubated for 45 min at 37°C. After incubation, the plates were washed 4 times and 200 µl of conjugates were added for 35 min at 37°C. After washing 4 times, 200 µl of the substrate solution was added. The absorbance was read using an ELISA reader at 450 nm.
Electron microscope
Cell masses were fixed with 2% paraformaldehyde and placed in 2.5% glutaraldehyde in a 0.1 M cacodylate buffer (pH 7.4). Then, the pellets were washed twice and dehydrated in a gradient series of ethanol solutions (25, 50, 70, 80, 90, and twice at 100% for 15 min). The samples were treated with acetone and flat-embedded in Durcupan (Fluka Chemic AG) and then sectioned to 60–70 nM thickness on 300 mesh copper slot grids. The ultrastructures of cells, including mitochondrial morphology, were examined (magnification, ×3,700, ×12,500), and the images were viewed under a Zeiss 109 electron microscope (Carl Zeiss).
Assay of intracellular ROS
The production of intracellular ROS was measured by a H2DCFDA-based ROS assay kit (Beyotime). Briefly, SiHa cells were subjected to different treatments, harvested, and then incubated with a final concentration of H2DCFDA (10 µM) for 30 min in the dark at 37°C. The fluorescence intensity of dichlorofluorescein (DCF) was observed by a fluorescence microscope. Flow cytometry (BD FACSCalibur) was used to detect the intracellular ROS generation.
Measurement of the mitochondrial membrane potential (ΔΨm)
Loss of the mitochondrial membrane potential (ΔΨm) was examined in SiHa cells using the fluorescent cationic dye JC-1 (Molecular Probes; Thermo Fisher Scientific, Inc.). SiHa cells were subjected to different treatments and then stained with 10 µM of JC-1 for 20 min at room temperature. The fluorescence levels were measured at two different wavelengths, 488 and 530 nm, using flow cytometry (BD FACSCalibur).
SiHa cell migration analysis
SiHa cell migration was detected using the Transwell (Corning Incorporated) assay. The upper chamber was filled with 100 µl cell suspension, and 600 µl culture medium containing 20% of FBS was added to the lower chamber. Migrated SiHa cells were fixed with 95% ethanol and then stained with 0.1% crystal violet staining. Migrated cells were counted under a microscope (×400).
SiHa cell proliferation assay
3H-thymidine was incorporated into SiHa cells to indicate DNA synthesis. The cultured SiHa cells were trypsinized and then harvested onto a glass fiber filter paper. A scintillation solution was added to detect the radioactivity, which was counted by a TopCount NxT scintillation counter (LKB Instruments).
Statistical analysis
Data are shown as mean ± standard deviation (SD). Statistical analyses were performed using IBM SPSS Statistics for Windows, version 22.0 (IBM Corp.). For the comparison of two groups, unpaired two-tailed Student's t-test was used. For the comparison of multiple groups, one-way ANOVA with Tukey's post hoc test was used. P<0.05 was considered statistically significant. All experiments were repeated 3 times and performed in triplicate.
Results
Localization and expression of FOXO1 in LPS-stimulated cervical squamous carcinoma SiHa cells
To detect the effects of LPS on FOXO1 expression, we carried out RT-qPCR, western blot, and immunohistochemical staining in cervical squamous carcinoma SiHa cells. According to this paper (18), LPS at 40 µg/ml leads to the most expression of TLR4. Therefore, we use this concentration of LPS to detect the expression changes of FOXO1 in our study. The RT-qPCR results indicated that LPS stimulation significantly inhibited the mRNA level of FOXO1 (Fig. 1A). Western blot data showed that the protein expression of FOXO1 was decreased in the LPS group compared with the mock group (Fig. 1B). In addition, immunohistochemical staining results showed that FOXO1 expression in the cytoplasm of SiHa cells was significantly decreased in the LPS-treated group (Fig. 1C).
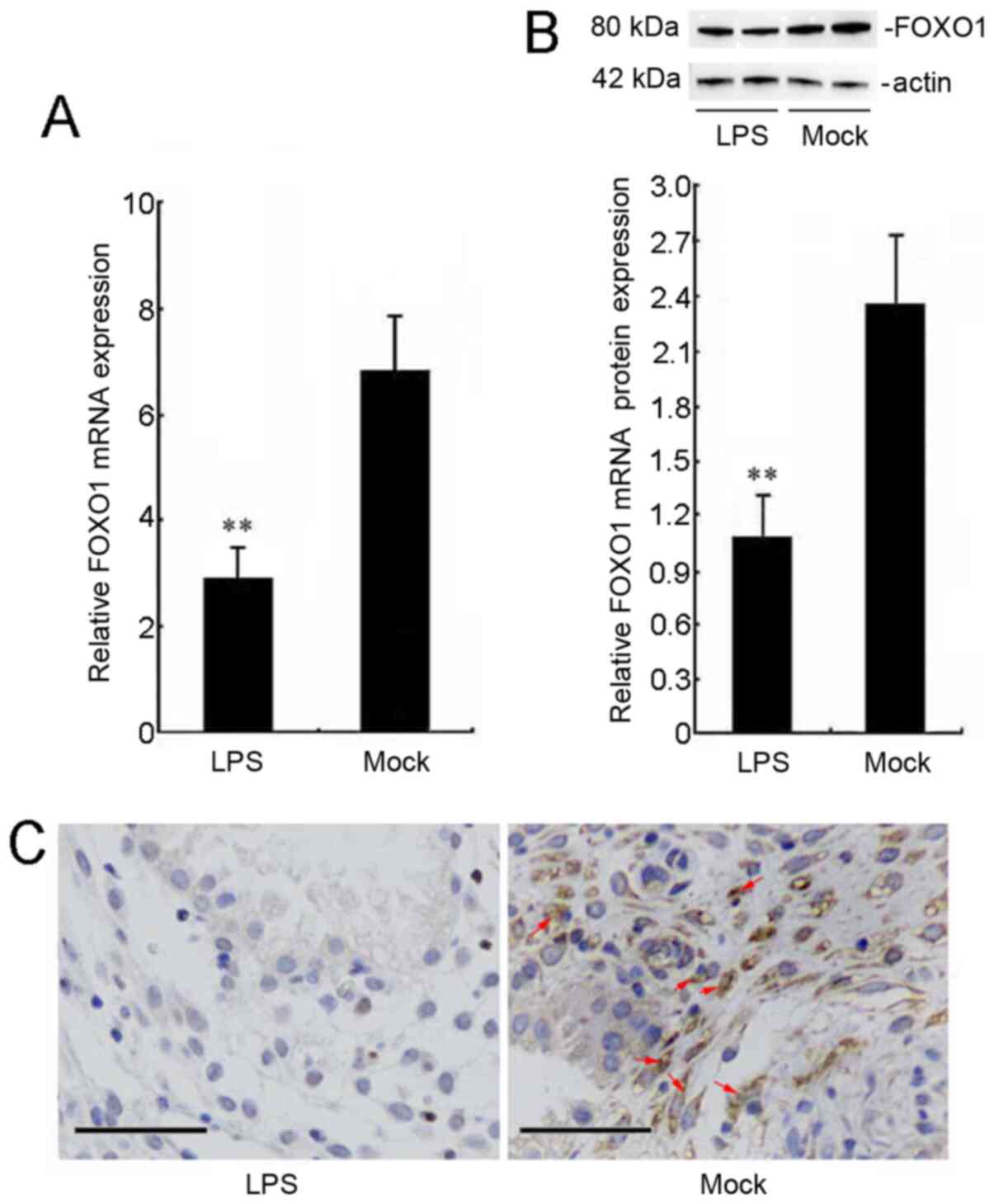 |
Figure 1.
Localization and expression of FOXO1 in LPS-stimulated cervical squamous carcinoma SiHa cells. SiHa cells were stimulated with LPS (40 µg/ml) for 24 h. (A) mRNA level of FOXO1 was examined using reverse transcription-quantitative PCR analysis. (B) Protein level of FOXO1 in lysates of SiHa cells was detected using western blot assay. The bar chart shows the ratio of FOXO1 to β-actin in each group. (C) Expression levels and localization of FOXO1 were detected using immunohistochemical staining analysis. SiHa cells that stained with brown cytoplasm were positive for FOXO1 (red arrows). Scale bars, 50 µm. **P<0.01 versus the mock group. FOXO1, Forkhead box O1; LPS, lipopolysaccharide.
|
Silencing FOXO1 leads to mitochondrial dysfunction in cervical squamous carcinoma SiHa cells
To study the effects of FOXO1 on mitochondrial function, we transfected FOXO1 siRNA into SiHa cells. The transfection efficiency of FOXO1 siRNA was good as shown in Fig. 2A and B. The mRNA expression of FOXO1 was reduced to 25% compared to the mock group (Fig. 2A). And the protein expression of FOXO1 was reduced to 35% compared to the mock group (Fig. 2B). Then the ROS production and mitochondrial membrane potential in SiHa cells were checked. These data indicated that ROS accumulation was enhanced by ~2.85-fold in the FOXO1 siRNA group compared with the mock treatment group (Fig. 2C). In addition, silencing FOXO1 decreased the mitochondrial membrane potential by ~63.4% compared with that in the mock group (Fig. 2D). However, in the negative siRNA group, the ROS accumulation and mitochondrial membrane potential showed no obvious changes compared with the mock group. Transmission electron microscopy images indicated that silencing FOXO1 caused changes in the mitochondrial morphology, including damage or disappearance of the mitochondrial cristae, and mitochondrial swelling (Fig. 2E).
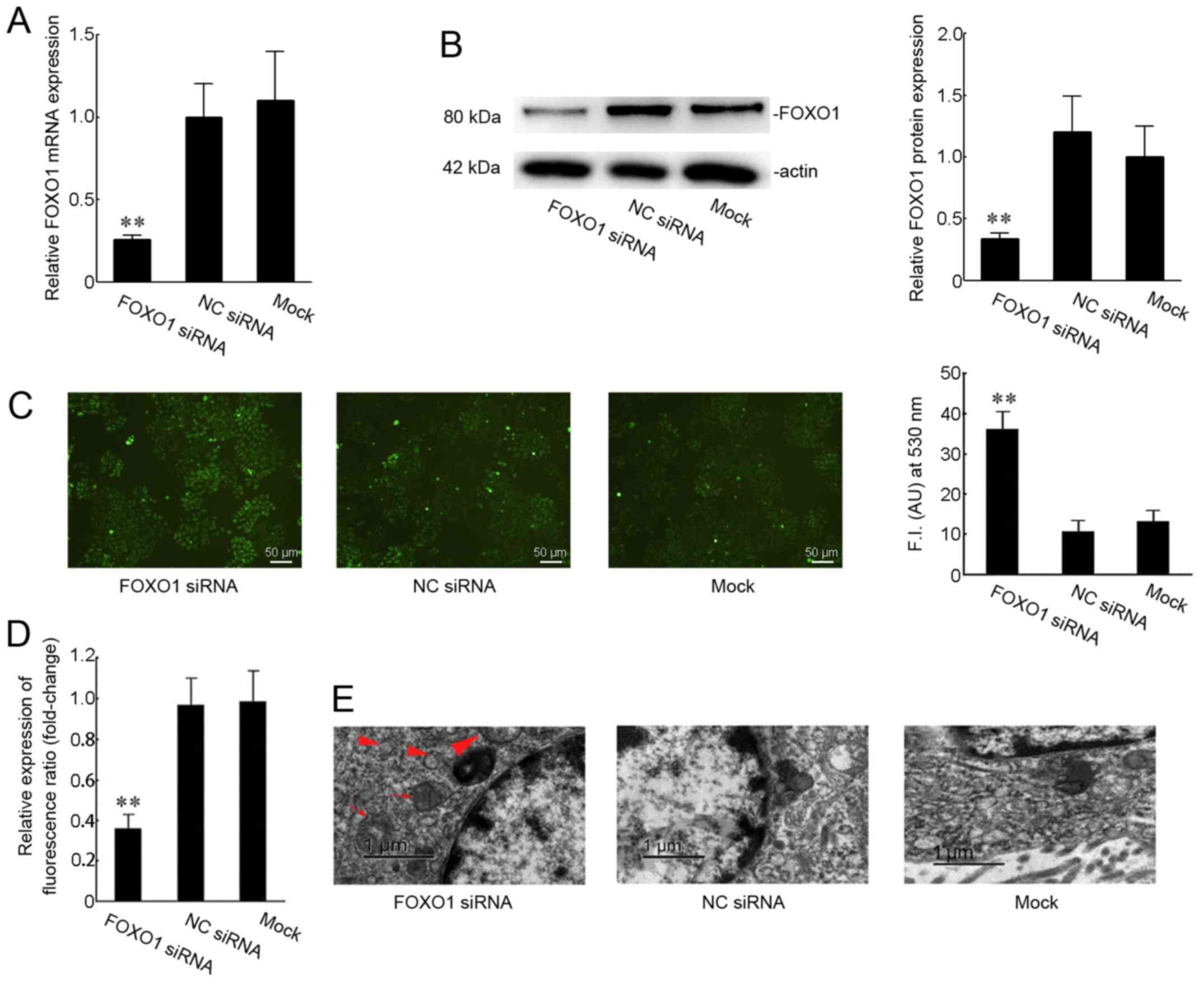 |
Figure 2.
Effect of FOXO1 on the mitochondrial function in SiHa cells. (A) Transfection success was confirmed using reverse transcription-quantitative PCR. Expression of FOXO1 was measured when SiHa cells were transfected with FOXO1 siRNA or NC siRNA for 48 h. (B) Protein expression levels of FOXO1 were checked in SiHa cells transfected with FOXO1 siRNA or NC siRNA for 48 h. (C) Production of intracellular reactive oxygen species was measured using H2DCFDA fluorescence (green). Scale bars, 50 µm. (D) Change in the mitochondrial membrane potential was analyzed by staining for JC-1 fluorescence. (E) Representative images of mitochondrial morphology visualized by transmission electron microscopy. The mitochondrial cristae were broken or disappeared (red triangles) and mitochondrial swelling was noted (red arrows). Scale bar, 1 µm. **P<0.01 versus the mock group. NC, negative control; si, short interfering; FOXO1, Forkhead box O1.
|
Effects of FOXO1 on mitochondrial function in LPS-stimulated SiHa cells
Since LPS treatment caused downregulation of FOXO1, and silencing FOXO1 effectively induced mitochondrial dysfunction, including ROS accumulation and mitochondrial membrane disruption, we hypothesized that overexpression of FOXO1 could reverse the effects of LPS on mitochondrial function in SiHa cells. To test this hypothesis, we transfected FOXO1 overexpression plasmids into SiHa cells and checked the transfection efficiency. The mRNA expression of FOXO1 was increased to 35-fold compared to the mock group (Fig. 3A). And the protein expression of FOXO1 was increased to 7 fold compared to the mock group (Fig. 3B). Then we performed H2DCFDA-based ROS assay and JC-1 staining, as well as transmission electron microscopy assay. The results revealed that mitochondrial ROS were maintained at a high level under 40 µg/ml LPS stimulation, and the ROS levels in the LPS (+) and FOXO1 vector (+) groups were decreased compared with those in the LPS alone-treated group (P<0.01; Fig. 3C). Treatment with LPS (+) and empty vector (+) caused no changes in ROS accumulation when compared with that in the LPS alone-treated group (Fig. 3C). Metformin was previously reported to protect the mitochondrial function and promote mitochondrial metabolism (20,21). Our results revealed that treatment with metformin attenuated LPS-induced ROS production (P<0.01; Fig. 3C). The mitochondrial membrane potentials were significantly increased in the LPS (+) and FOXO1 vector (+) group or the metformin treatment group compared with the LPS (+) and empty vector (+) group or the LPS alone-treated group (Fig. 3D). As shown in Fig. 3E, transmission electron microscopy images indicated changes in the mitochondrial morphology, including mitochondrial swelling and vesicular formation in the LPS alone-treated group or the LPS (+) and empty vector (+) group. However, enhancing FOXO1 expression or administration of metformin maintained the integrity of mitochondrial morphology (Fig. 3E).
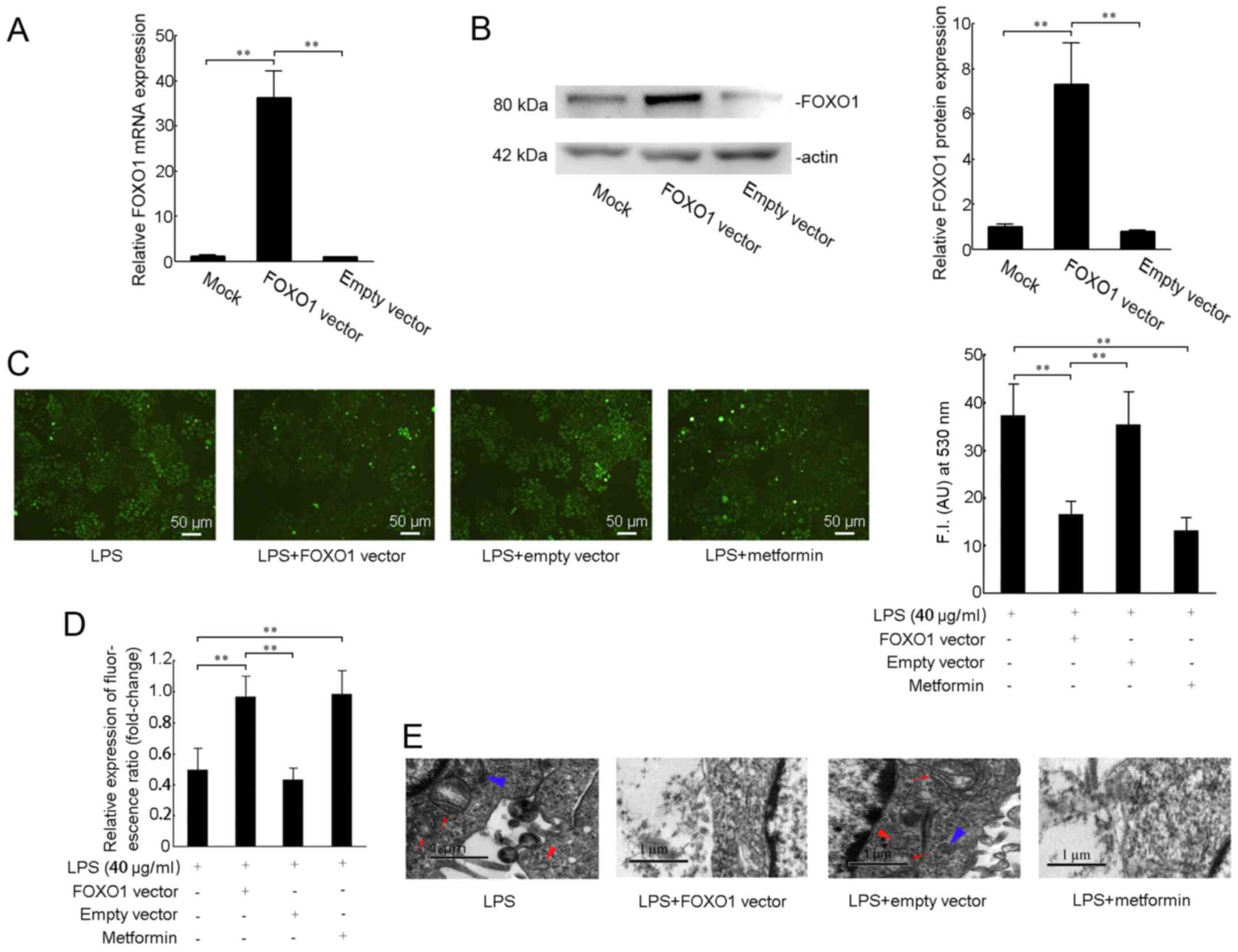 |
Figure 3.
Effect of FOXO1 on mitochondrial function in LPS-stimulated cervical squamous carcinoma SiHa cells. (A) mRNA and (B) protein expression levels of FOXO1 were measured after SiHa cells were transfected with pcDNA3.1-FOXO1 vector (2 µg/ml) or pcDNA3.1 empty vector for 48 h. (C) Cells were stimulated with LPS (40 µg/ml) for 24 h after transfection with the pcDNA3.1-FOXO1 vector (2 µg/ml), pcDNA3.1 empty vector or treatment with metformin for 48 h. Intracellular reactive oxygen species levels were measured using H2DCFDA fluorescence (green). Scale bars, 50 µm. (D) Mitochondrial membrane potential was detected by the change in JC-1 fluorescence. (E) Representative transmission electron microscopy images. The image shows that the mitochondria were swollen (red arrows), vesicles were formed (red triangle) or the length was significantly shortened (blue triangle). Scale bar, 1 µm. **P<0.01. FOXO1, Forkhead box O1; LPS, lipopolysaccharide.
|
Effects of FOXO1 on biological function in LPS-stimulated SiHa cells
A previous study has shown that LPS promoted the progression and metastasis of cervical cancer by stimulating the release of IL-1β; thus, promoting downstream cytokines and pyroptosis (14). LPS markedly induced pyroptosis by increasing the protein levels of caspase-1 (22). Metformin inhibits cancer cell proliferation, migration and invasion (23). In this study, ELISA analysis showed that 40 µg/ml LPS stimulation significantly increased the extracellular levels of IL-1β in culture media. Pre-transfection with the FOXO1 vector or pre-treatment with metformin attenuated LPS-induced IL-1β generation (Fig. 4A). Activation of caspase-1 is required during IL-1β production. As shown in Fig. 4B, 40 µg/ml LPS stimulation increased the expression of caspase-1, while overexpression of FOXO1 or treatment with metformin decreased the LPS-induced expression of caspase-1 (Fig. 4B). To further evaluate the role of FOXO1 in LPS-induced SiHa cell migration and proliferation ability, Transwell experiments and 3H-TdR incorporation assays showed that the migration and proliferation of cells were reduced in the LPS (+) and FOXO1 vector (+)-treated group or LPS (+) and metformin (+) group compared with the LPS alone group (P<0.01) (Figs. 4C and D). However, treatment with LPS (+) and empty vector (+) caused no difference compared with LPS alone-treated group (Fig. 4A-D).
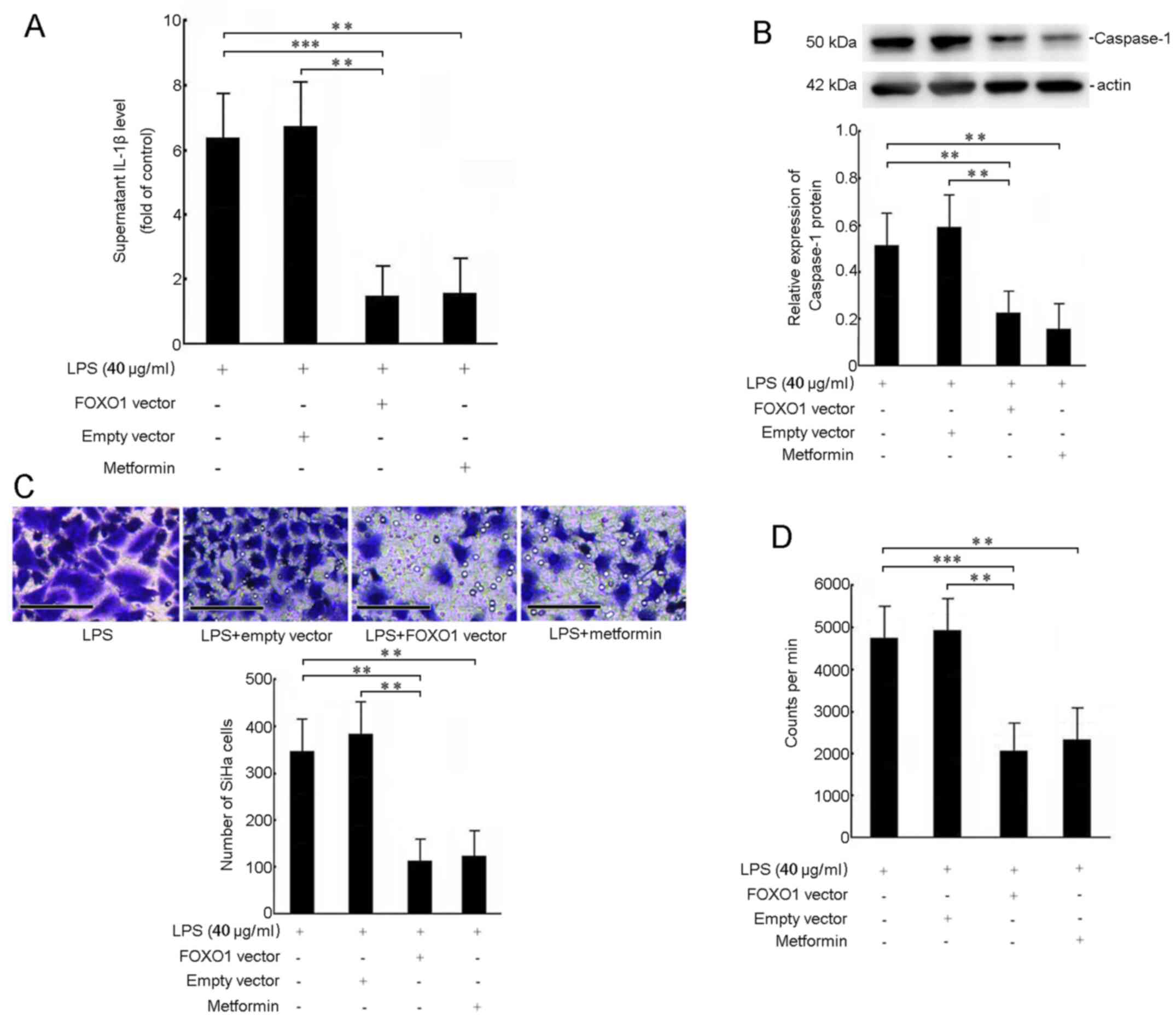 |
Figure 4.
Effect of FOXO1 on biological function in LPS-stimulated SiHa cells. Cells were stimulated with LPS (40 µg/ml) for 24 h after transfection with the pcDNA3.1-FOXO1 vector (2 µg/ml), pcDNA3.1 empty vector (2 µg/ml) or treatment with metformin for 48 h. (A) Extracellular levels of IL-1β were determined using ELISA. (B) Protein levels of caspase-1 in lysates of SiHa cells were measured using western blotting. (C) SiHa cell migration was measured using a Transwell migration assay (n=3). Scale bar, 50 µm. (D) 3H-TdR incorporation was executed to assess the proliferation of SiHa cells. The absolute value of CPM indicated the proliferation of cells (cpm/106 cells). **P<0.01, ***P<0.001. FOXO1, Forkhead box O1; LPS, lipopolysaccharide; CPM, counts per min.
|
Discussion
The development of cervical squamous cell carcinoma is strongly associated with persistent infection and chronic inflammation. LPS is a common inducer of inflammation, and various signaling pathways, including Janus kinase/signal transducers and activators of transcription (JAK/STAT), mitogen-activated protein kinases (MAPKs), and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) are activated under stimulation with LPS (24). LPS exposure is positively related to the survival, invasion, proliferation, and metastasis of tumor cells, by stimulating the release of tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and IL-1β (25–27). In this work, we first tested the influence of LPS on FOXO1 expression. The results showed that LPS could downregulate FOXO1 expression in cervical squamous carcinoma SiHa cells. The mechanisms may be strongly associated with signaling pathways, including JAK/STAT, MAPKs, and NF-κB. Largely consistent with our results, exposure to LPS was found to cause downregulation of FOXO1 in renal tubular epithelial cells (28).
Mitochondria are the major physiological source of ROS, and they cause an increase in oxidative stress when they become dysfunctional (29,30). Mitochondrial dysfunction is characterized by excessive ROS production in defective mitochondria, loss of ATP, and disruption of the integrity of the mitochondrial membrane. Previously published studies have shown that the transcription factor FOXO1 regulates cell cycle progression, oxidative stress, apoptosis, autophagy, tumor suppression, cell differentiation, and energy metabolism (31,32). Particularly, FOXO1 is a critical regulator of the mitochondrial metabolism, which is involved in the occurrence and development of tumors. In this study, we found that silencing FOXO1 increases ROS production, and leads to ΔΨm loss and disruptions of mitochondrial morphology in cervical squamous carcinoma SiHa cells. These observations imply the roles of FOXO1 in mitochondrial dysfunction in SiHa cells.
Recent studies have proved that LPS stimulation induced mitochondrial damage-mediated ROS production in HepG2 cells (24), increased the levels of inflammatory cytokines in the brain, and deregulated the mitochondrial function in mice (33). ROS accumulation induced by LPS leads to the state of oxidative stress, which also promotes inflammatory progression (34). In this work, we found that LPS induced ROS production, inhibited the mitochondrial membrane potential, and disrupted the mitochondrial morphology in cervical squamous carcinoma SiHa cells. These data uncovered that LPS-induced mitochondrial dysfunction may promote inflammatory progression in cervical squamous cell carcinoma.
Previous studies have revealed that overexpression of FOXO1 improved LPS-induced renal injury and mitochondrial dysfunction partly through PGC1-α signaling (28). Other studies have reported that cobalt protoporphyrin upregulated FOXO1 expression, further increased the expression of the heme oxygenase-1 (HO-1) gene and other oxidative stress-responsive genes, and reduced the production of mitochondria-derived ROS (35). These findings proposed the roles of FOXO1 in improving mitochondrial function during LPS exposure. Also, decreased expression of FOXO1 mediated the role of LPS in mitochondrial dysfunction in SiHa cells. The mechanism may occur partly through PGC1-α signaling or upregulation of the HO-1 gene. However, further studies are needed to explore the detailed mechanisms.
As previously described, mitochondrial dysfunction contributes to the loss of cell function, apoptosis, and pyroptosis (36,37). Pyroptosis is a form of programmed cell death dependent on the activation of caspase-1. Caspase-1 was a marker of cell pyroptosis (38). NLRP3 inflammasome (a complex of NLRP3, apoptosis-associated speck-like (ASC), and procaspase-1) cleaves procaspase-1 as an activated form of caspase-1, leading to gasdermin D (GSDMD) cleavage. Then the cleaved GSDMD forms a pore in the cellular membrane and induces pyroptosis. Additionally, cleaved caspase-1, enhances the maturation of proinflammatory cytokines, such as IL-1β (38). LPS stimulation activated the TLR4-NF-κB signaling pathway and inflammatory mediators, such as IL-1β, in cervical squamous carcinoma cells (18). LPS markedly induced pyroptosis by increasing the protein levels of caspase-1 (22). Cell migration and proliferation were also promoted by LPS in cervical cancer SiHa cells (39). FOXO1 is an important transcription factor with a regulatory role in cell proliferation and differentiation (40). The present study revealed that overexpression of FOXO1 could reverse LPS-induced IL-1β secretion, caspase-1 expression, cell migration, and proliferation of SiHa cells. These results suggested that decreased expression of FOXO1 mediated the role of LPS in SiHa cell pyroptosis, migration, and proliferation. However, further studies are needed to explore the detailed mechanisms. For the pyroptosis, although we determined caspase-1 and IL-1β expression in this study, the mRNA and protein levels of NLRP3, ASC, GSDMD should be checked in the future.
In Figs. 3 and 4, ‘no LPS’ was not used. Our study focus on the roles of rescue effects of FOXO1 in LPS-induced situation. And the effects of LPS on mitochondrial function in Fig. 3 could be compared to the mock group in Fig. 2. In Fig. 4, various study demonstrated that LPS could increase IL-1β expression, caspase-1 expression, cell migration and proliferation compared to the no LPS control (18,22,39,40). Thus, ‘no LPS’ was not used.
Prior studies have implicated that the functions of FOXO1 in cancer progression were controversial. It has been reported that FOXO1 silencing in OVCA429 and OVCA433 cells resulted in decreased cell proliferation. In addition, knockout of FOXO1 had a significant negative effect on the epithelial ovarian cancer cell migration (41). However, recent studies have indicated that FOXO1 played an antitumor role by regulating proapoptotic genes, such as Bcl-2 interacting mediator of cell death (BIM), fas ligand protein (FasL), and TNF related apoptosis inducing ligand (TRAIL), which in turn inhibited the growth of various tumors, such as glioblastoma and cervical squamous cell carcinoma (42). In this study, we examined the roles of FOXO1 in human cervical squamous carcinoma SiHa cell growth (proliferation) and metastasis (migration) in LPS-induced functional studies. Our results showed that overexpression of FOXO1 attenuated LPS-induced migration and proliferation of cervical squamous carcinoma SiHa cells. Another study has implicated that metformin inhibited estrogen-dependent endometrial cancer cell growth by activating the AMPK-FOXO1 signal pathway (43). The present study demonstrated that metformin reversed the roles of LPS in mitochondrial function, cell pyroptosis, proliferation, and migration. The mechanism may be that metformin upregulated the FOXO1 activity by AMPK activation, and then reversed the roles of LPS. However, further mechanism studies are required.
Taken together, our study uncovered that decreased expression of FOXO1 may mediate the functions of LPS in cervical squamous carcinoma SiHa cell mitochondrial function, pyroptosis, proliferation, and migration. Also, FOXO1 may be a potential therapeutic target for prevention and treatment of human cervical squamous cell carcinoma from the mitochondrial perspective. However, further studies are required to investigate the therapeutic potential of FOXO1.
Acknowledgements
Not applicable.
Funding
The present work was supported by the Basic Scientific Research Operating Expense Project of Heilongjiang Provincial Department of Education (grant no. 2019-KYYWF-0982).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
SL made substantial contributions to the conception and design of the study. HW and SL confirm the authenticity of all the raw data. HW and SL designed the experiments and wrote and edited the manuscript. ZM, FG, WJ and YL performed the experiments, analyzed and interpreted the data and performed the statistical analysis. All authors have read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Authors' information
Shuping Li ORCID: https://orcid.org/0000-0001-7545-4750.
References
|
1
|
Arbyn M, Weiderpass E, Bruni L, de Sanjosé S, Saraiya M, Ferlay J and Bray F: Estimates of incidence and mortality of cervical cancer in 2018: A worldwide analysis. Lancet Glob Health. 8:e191–e203. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A and Bray F: Global Cancer Statistics 2020: GLOBOCAN Estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Di J, Rutherford S and Chu C: Review of the cervical cancer burden and population-based cervical cancer screening in China. Asian Pac J Cancer Prev. 16:7401–7407. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wen X, Liu S and Cui M: Effect of BRCA1 on the concurrent chemoradiotherapy resistance of cervical squamous cell carcinoma based on transcriptome sequencing analysis. BioMed Res Int. 2020:35984172020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fang J, Zhang H and Jin S: Epigenetics and cervical cancer: From pathogenesis to therapy. Tumour Biol. 35:5083–5093. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takekuma M, Kasamatsu Y, Kado N, Kuji S, Tanaka A, Takahashi N, Abe M and Hirashima Y: The issues regarding postoperative adjuvant therapy and prognostic risk factors for patients with stage I–II cervical cancer: A review. J Obstet Gynaecol Res. 43:617–626. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tewari AK, Stockert JA, Yadav SS, Yadav KK and Khan I: Inflammation and prostate cancer. Adv Exp Med Biol. 1095:41–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sadri Nahand J, Moghoofei M, Salmaninejad A, Bahmanpour Z, Karimzadeh M, Nasiri M, Mirzaei HR, Pourhanifeh MH, Bokharaei-Salim F, Mirzaei H, et al: Pathogenic role of exosomes and microRNAs in HPV-mediated inflammation and cervical cancer: A review. Int J Cancer. 146:305–320. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xie X, Yang M, Ding Y and Chen J: Microbial infection, inflammation and epithelial ovarian cancer. Oncol Lett. 14:1911–1919. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Katoh M: Multi layered prevention and treatment of chronic inflammation, organ fibrosis and cancer associated with canonical WNT/β catenin signaling activation (Review). Int J Mol Med. 42:713–725. 2018.PubMed/NCBI
|
|
12
|
Monkkonen T and Debnath J: Inflammatory signaling cascades and autophagy in cancer. Autophagy. 14:190–198. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu X, Zhao W, Wang W, Lin S and Yang L: Puerarin suppresses LPS-induced breast cancer cell migration, invasion and adhesion by blockage NF-kappaB and Erk pathway. Biomed Pharmacother. 92:429–436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
You L, Cui H, Zhao F, Sun H, Zhong H, Zhou G and Chen X: Inhibition of HMGB1/RAGE axis suppressed the lipopolysaccharide (LPS)-induced vicious transformation of cervical epithelial cells. Bioengineered. 12:4995–5003. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Winograd R, Simeone DM and Bar-Sagi D: A novel target for combination immunotherapy in pancreatic cancer: IL-1β mediates immunosuppression in the tumour microenvironment. Br J Cancer. 124:1754–1756. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang B, Gui LS, Zhao XL, Zhu LL and Li QW: FOXO1 is a tumor suppressor in cervical cancer. Genet Mol Res. 14:6605–6616. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goto T and Takano M: Transcriptional role of FOXO1 in drug resistance through antioxidant defense systems. Adv Exp Med Biol. 665:171–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He A, Ji R, Shao J, He C, Jin M and Xu Y: TLR4-MyD88-TRAF6-TAK1 complex-mediated NF-κB activation contribute to the anti-inflammatory effect of V8 in LPS-induced human cervical cancer SiHa cells. Inflammation. 39:172–181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao LJ, Gu PQ, Fan WM, Liu Z, Qiu F, Peng YZ and Guo XR: The role of gC1qR in regulating survival of human papillomavirus 16 oncogene-transfected cervical cancer cells. Int J Oncol. 39:1265–1272. 2011.PubMed/NCBI
|
|
21
|
Tyszka-Czochara M, Bukowska-Strakova K and Majka M: Metformin and caffeic acid regulate metabolic reprogramming in human cervical carcinoma SiHa/HTB-35 cells and augment anticancer activity of Cisplatin via cell cycle regulation. Food Chem Toxicol. 106:260–272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu XH, Wu LM, Wang JL, Dong XH, Zhang SC, Li XH, Xu H, Liu DB, Li ZH, Liu ZM, et al: Long non-coding RNA RP11-490M8.1 inhibits lipopolysaccharide-induced pyroptosis of human umbilical vein endothelial cells via the TLR4/NF-κB pathway. Immunobiology. 226:1521332021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Safe S, Nair V and Karki K: Metformin-induced anticancer activities: Recent insights. Biol Chem. 399:321–335. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nguyen TLL, Huynh DTN, Jin Y, Jeon H and Heo KS: Protective effects of ginsenoside-Rg2 and -Rh1 on liver function through inhibiting TAK1 and STAT3-mediated inflammatory activity and Nrf2/ARE-mediated antioxidant signaling pathway. Arch Pharm Res. 44:241–252. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu LM, Liang DY, Ye CG, Tu WJ and Zhu T: The UII/UT system mediates upregulation of proinflammatory cytokines through p38 MAPK and NF-κB pathways in LPS-stimulated Kupffer cells. PLoS One. 10:e01213832015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shen JQ, Zhang ZX, Shen CF and Liao JZ: Anticarcinogenic effect of Umbelliferone in human prostate carcinoma: An in vitro study. J BUON. 22:94–101. 2017.PubMed/NCBI
|
|
27
|
Veena VK, Popavath RN, Kennedy K and Sakthivel N: In vitro antiproliferative, pro-apoptotic, antimetastatic and anti-inflammatory potential of 2,4-diacetylphloroglucinol (DAPG) by Pseudomonas aeruginosa strain FP10. Apoptosis. 20:1281–1295. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang M, Dong W, Li Z, Xiao Z, Xie Z, Ye Z, Liu S, Li R, Chen Y, Zhang L, et al: Effect of forkhead box O1 in renal tubular epithelial cells on endotoxin-induced acute kidney injury. Am J Physiol Renal Physiol. 320:F262–F272. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moloney JN and Cotter TG: ROS signalling in the biology of cancer. Semin Cell Dev Biol. 80:50–64. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Poillet-Perez L, Despouy G, Delage-Mourroux R and Boyer-Guittaut M: Interplay between ROS and autophagy in cancer cells, from tumor initiation to cancer therapy. Redox Biol. 4:184–192. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang JB, Zhao ZB, Liu QZ, Hu TD, Long J, Yan K and Lian ZX: FoxO1 is a regulator of MHC-II expression and anti-tumor effect of tumor-associated macrophages. Oncogene. 37:1192–1204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Essers MA, de Vries-Smits LM, Barker N, Polderman PE, Burgering BM and Korswagen HC: Functional interaction between beta-catenin and FOXO in oxidative stress signaling. Science. 308:1181–1184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Haj-Mirzaian A, Ramezanzadeh K, Tafazolimoghadam A, Kazemi K, Nikbakhsh R, Nikbakhsh R, Amini-Khoei H, Afshari K, Haddadi NS, Shakiba S, et al: Protective effect of minocycline on LPS-induced mitochondrial dysfunction and decreased seizure threshold through nitric oxide pathway. Eur J Pharmacol. 858:1724462019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mihaly SR, Ninomiya-Tsuji J and Morioka S: TAK1 control of cell death. Cell Death Differ. 21:1667–1676. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu X, Cui Y, Li M, Xu H, Zuo J, Fang F and Chang Y: Cobalt protoporphyrin induces HO-1 expression mediated partially by FOXO1 and reduces mitochondria-derived reactive oxygen species production. PLoS One. 8:e805212013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li DX, Wang CN, Wang Y, Ye CL, Jiang L, Zhu XY and Liu YJ: NLRP3 inflammasome-dependent pyroptosis and apoptosis in hippocampus neurons mediates depressive-like behavior in diabetic mice. Behav Brain Res. 391:1126842020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shi C, Wang Q, Rao Z, Shi Y, Wei S, Wang H, Lu X, Wang P, Lu L, Zhou H, et al: Diabetes induces hepatocyte pyroptosis by promoting oxidative stress-mediated NLRP3 inflammasome activation during liver ischaemia and reperfusion injury. Ann Transl Med. 8:7392020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Oh S, Yang J, Park C, Son K and Byun K: Dieckol attenuated glucocorticoid-induced muscle atrophy by decreasing NLRP3 inflammasome and pyroptosis. Int J Mol Sci. 22:80572021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang H, Fang Z, Qu X, Zhang X and Wang Y: Procyanidin compound (PC) suppresses lipopolysaccharide-induced cervical cancer cell proliferation through blocking the TLR4/NF-κB pathway. Cancer Manag Res. 12:497–509. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Singh BK, Sinha RA, Zhou J, Tripathi M, Ohba K, Wang ME, Astapova I, Ghosh S, Hollenberg AN, Gauthier K, et al: Hepatic FOXO1 target genes are co-regulated by thyroid hormone via RICTOR protein deacetylation and MTORC2-AKT protein inhibition. J Biol Chem. 291:198–214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang N, Qiu J, Zheng T, Zhang X, Hua K and Zhang Y: Goserelin promotes the apoptosis of epithelial ovarian cancer cells by upregulating forkhead box O1 through the PI3K/AKT signaling pathway. Oncol Rep. 39:1034–1042. 2018.PubMed/NCBI
|
|
42
|
Prasad SB, Yadav SS, Das M, Govardhan HB, Pandey LK, Singh S, Pradhan S and Narayan G: Down regulation of FOXO1 promotes cell proliferation in cervical cancer. J Cancer. 5:655–662. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zou J, Hong L, Luo C, Li Z, Zhu Y, Huang T, Zhang Y, Yuan H, Hu Y, Wen T, et al: Metformin inhibits estrogen-dependent endometrial cancer cell growth by activating the AMPK-FOXO1 signal pathway. Cancer Sci. 107:1806–1817. 2016. View Article : Google Scholar : PubMed/NCBI
|














