Introduction
Eukaryotic gene expression primarily includes the
processes of transcription, splicing and translation. First, DNA is
transcribed into an mRNA precursor (pre-mRNA) by RNA polymerases.
Then, the pre-mRNA is spliced into mature mRNA by spliceosomes,
after which the mature mRNA is translated into protein. Since
multiple splicing sites are usually present in pre-mRNA, several
splicing isoforms may be generated in different types of cells or
biological processes by alternative splicing. These isoforms have a
synergistic or antagonistic biological function, and >90% of
human genes are spliced after transcription (1,2).
Normal gene splicing is critical for the functions of tissues and
organs (3). However, aberrant
splicing of key genes can lead to various diseases, particularly
the occurrence and development of tumors (4–7).
Spliceosomes usually consist of small nuclear
ribonucleoproteins (snRNPs), core proteins and cofactors (8). Core proteins include two highly
conserved families: i) Serine/arginine-rich splicing factors
(SRSFs); and ii) heterogeneous nuclear ribonucleoproteins (hnRNPs).
During splicing, the serine/arginine-rich domain (RS) of the SRSF
binds to the exonic splicing enhancer (ESE) of the splicing
regulatory element, facilitating the binding of snRNP to the 5′-end
and the 3′-end splicing sites, which cause phosphorylation or
dephosphorylation of target proteins (9,10).
At present, 12 members of the SRSF family have been
identified, among which SRSF3 is the smallest; the N-terminal of
SRSF3 contains an RNA recognition motif that specifically
recognizes the splicing sites, and the C-terminal of SRSF3 contains
a highly phosphorylated serine/arginine-rich domain (RS) region
(11). SRSF3 binds to
cytosine-rich motifs and promotes the inclusion of exons,
especially genes encoding neutral amino acids (such as serine and
threonine), which can be phosphorylated (12). SRSF3 interacts with RNA cis-acting
elements in a concentration- or cell differentiation-dependent
manner. Moreover, SRSF3 promotes the inclusion of exons in target
genes by recruiting Yes1 associated transcriptional regulator
(YTHDC1). YTHDC1 is closely associated with the nuclear
localization, RNA binding ability and related splicing events of
SRSF3 (13,14). SRSF3 has been discovered to be
widely expressed in tumor tissues and involved in the splicing of
numerous hub genes, such as CD44, HER2, MDM4, Rac family small
GTPase 1 (RAC1) and tumor protein p53 (TP53) (Table I; Figs. 1 and 2) (15).
Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis
has demonstrated that SRSF3 is highly associated with in the
PI3K-AKT signaling pathway. Consequently, SRSF3 functions as an
oncogene with critical regulatory roles in tumorigenesis,
proliferation, metastasis, anti-apoptosis, anti-senescence and
anti-autophagy (Xiong et al, unpublished data). In addition,
SRSF3 plays important roles in various biological processes, such
as protein translation (16),
termination of transcription (17), insulin signaling (18), mRNA transport (19) and embryogenesis (20).
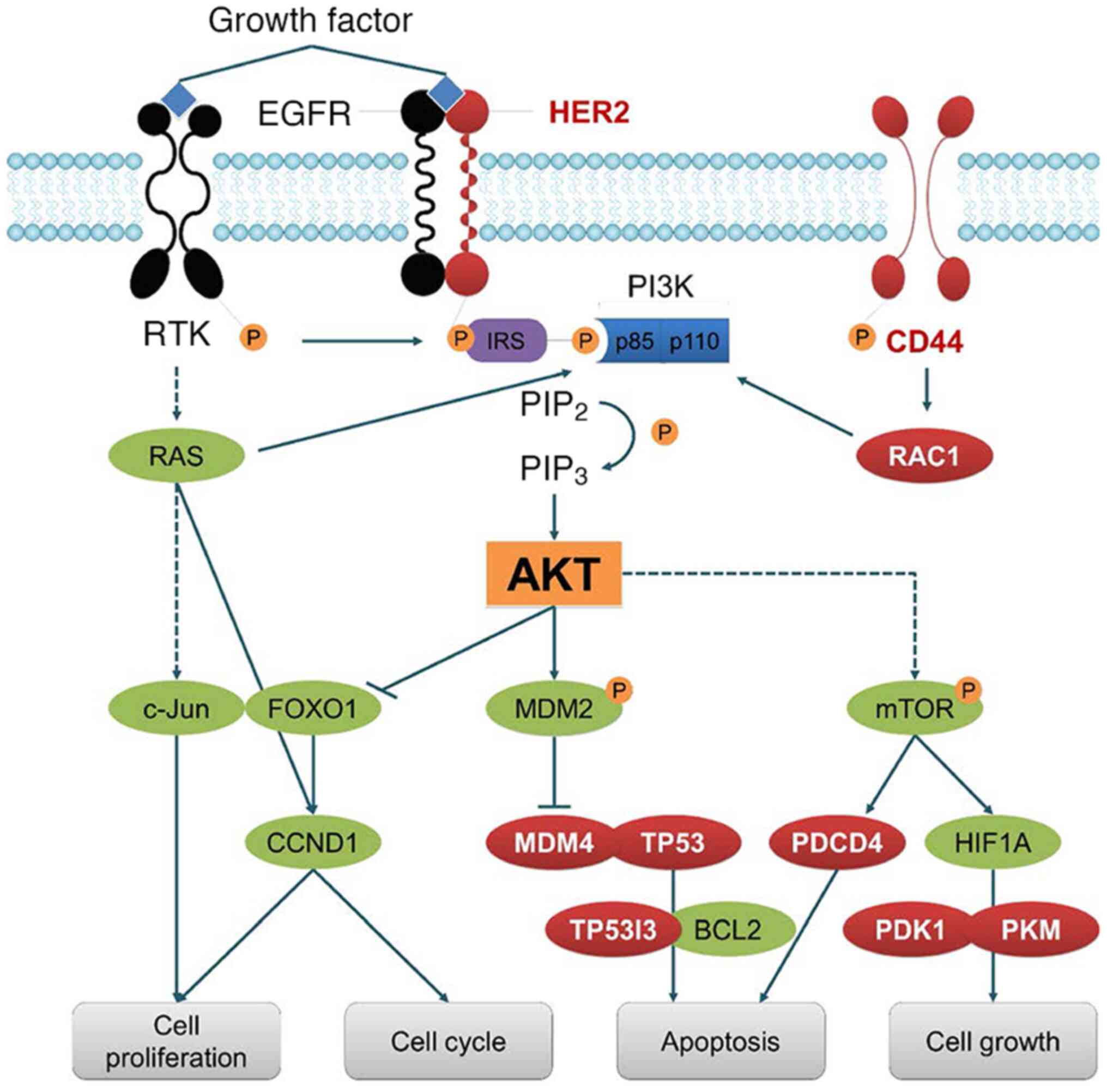 | Figure 1.Targets of SRSF3. SRSF3 is involved
in the splicing of numerous hub genes, including CD44, HER2, MDM4,
RAC1 and TP53. Therefore, SRSF3 plays important roles in the
regulation of cell proliferation, cell cycle, apoptosis and
proliferation. SRSF3, serine/arginine rich splicing factor 3; MDM,
mouse double minute; RAC1, Rac family small GTPase 1; TP53, tumor
protein p53; RTK, receptor tyrosine kinases; IRS, insulin receptor
substrate; PIP3, phosphatidylinositol (3,4,5)-trisphosphate; PIP2,
phosphatidylinositol 4,5-bisphosphate; CCND1, cyclin D1; PDCD4,
programmed cell death 4; PDK1, pyruvate dehydrogenasekKinase 1;
PKM, pyruvate kinase M; HIF1A, hypoxia inducible factor 1 subunit
α; P, phosphate. |
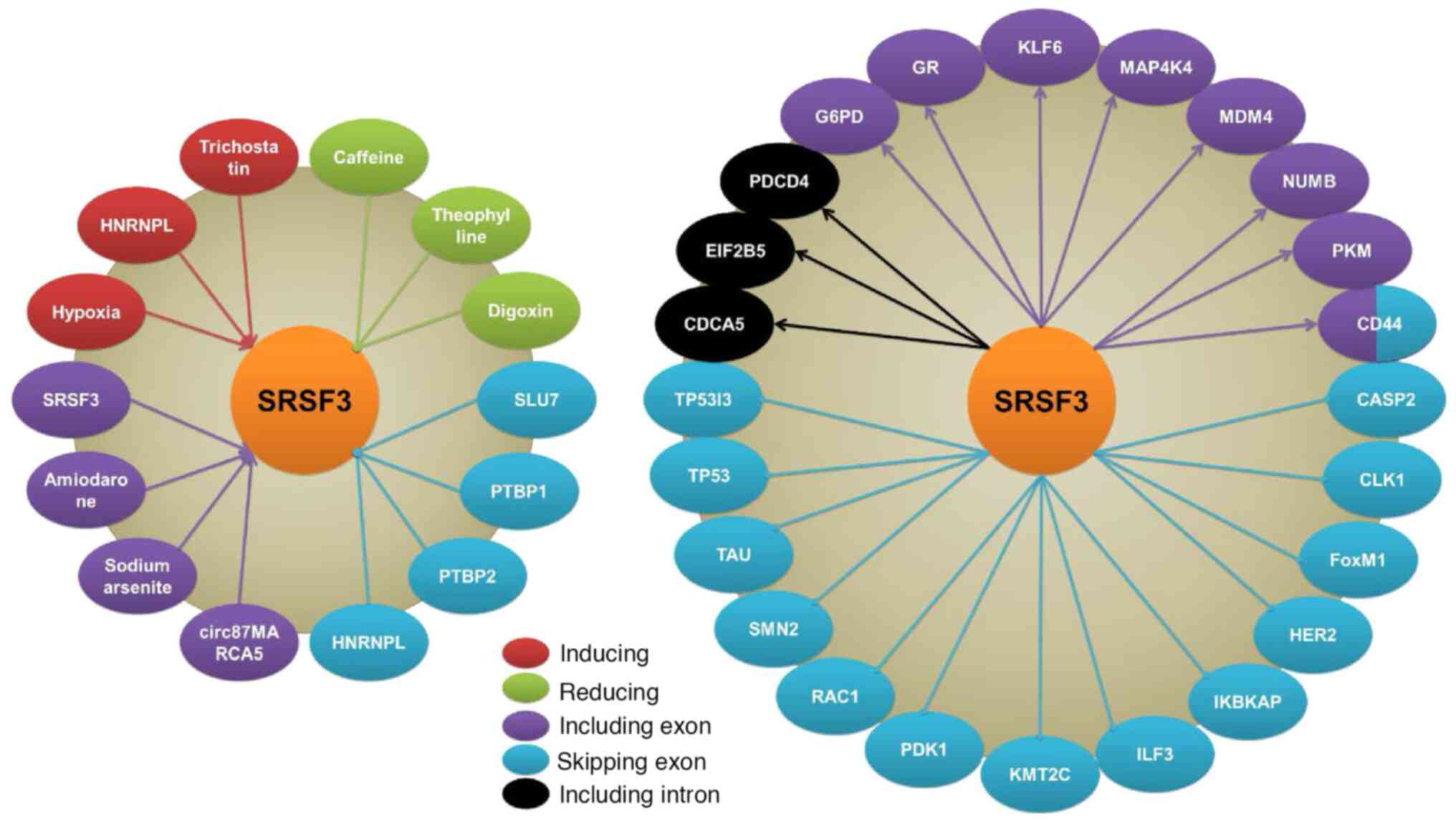 | Figure 2.Regulators, targets and splicing
events of SRSF3. Hypoxia, HNRNPL and trichostatin induce SRSF3
expression, while caffeine, theophylline and digoxin inhibit SRSF3
expression. The splicing of SRSF3 is regulated by SRSF3 itself,
amiodarone, sodium arsenite and cir87MARCA5 by including exon, and
also regulated by HNRNPL, PTBP1, PTBP2 and SLU7 through exon
skipping. SRSF3 regulates splicing of multiple genes in different
ways such as including exon, skipping exon and including intron.
SRSF3, serine/arginine rich splicing factor 3; HNRNPL,
heterogeneous nuclear ribonucleoprotein L; PTBP, polypyrimidine
tract binding protein; SLU7, SLU7 homolog. |
 | Table I.Regulators, targets and splicing
events of SRSF3. |
Table I.
Regulators, targets and splicing
events of SRSF3.
| Regulator | Target | Splicing event | Isoform | Cell line | (Refs.) |
|---|
| HNRNPL | SRSF3 | Skipping of exon
4 | SRSF3-FL | CAL 27 | (23) |
| PTBP1 and
PTBP2 |
|
|
| OSCC | (46) |
| SLU7 |
|
|
| PLC, H358, and
HeLa | (47) |
| SRSF3 | SRSF3 | Inclusion of exon
4 | SRSF3-TR | K46 | (43–45) |
| Amiodarone |
|
|
| HeLa | (49) |
| Sodium
arsenite |
|
|
| HCT116 | (50) |
| circ87MARCA5 |
|
|
| U87MG | (51) |
| SRSF3 | CDCA5 | Retention of intron
1 and2 | NA | PLC | (47) |
|
| EIF2B5 | Retention of intron
12 | EIF2B5-TR | SQ20B | (16) |
|
| PDCD4 | Retention of intron
2 | Isoform2 | SW480 | (29) |
|
| CD44 | Splicing of exons
8, 9, and 10 | CD44E | HEK293 | (32,59) |
|
| G6PD | Inclusion of exon
12 | G6PD-FL | HepG2 | (54) |
|
| GR | Inclusion of exon
9a | GRα | MDA-MB-231 | (74,75) |
|
| KLF6 | Inclusion of exon
3 | KLF6-FL | Hep3B, Huh7, and
HepG2 | (22) |
|
| MAP4K4 | Inclusion of exon
16 | Isoform2 and 5 | HCT-8 | (73) |
|
| MDM4 | Inclusion of exon
6 | MDM4-FL | A375 | (67) |
|
| NUMB | Inclusion of exon
12 | NUMB-FL | MDA-MB-231 | (76) |
|
| PKM | Inclusion of exon
10 | PKM2 | DLD-1 and WiDr | (61) |
|
| CASP2 | Skipping of exon
9 | Casp-2L | HeLa and
MDA-MB-231 | (107) |
|
| CLK1 | Skipping of exon
4 | CLK1-TR | WI-38 | (40) |
|
| FoxM1 | Skipping of exon 9
and/or 6 | Isoform b and
c | U2OS and HeLa | (24) |
|
| HER2 | Skipping of exon
16 | Δ16HER2 | SKBR3 | (108) |
|
| ELP1 | Skipping of exon
20 | ELP1-TR | HEK293 | (40) |
|
| ILF3 | Skipping of exon 18
or 18b | Isoform1 and2 | WI-38 | (66) |
|
| KMT2C | Skipping of exon
46a | KMT2C-S | A2780 | (68) |
|
| PDK1 | Skipping of exon
3a | PDK1-TR | HEK293 | (40) |
|
| RAC1 | Skipping of exon
3b | RAC1b | HT29 | (33) |
|
| SMN2 | Skipping of exon
7 | SMN2-TR | HeLa | (83) |
|
| MAPT | Skipping of exon
10 | MAPT-TR | SKN-MC and
C33A | (84) |
|
| TP53 | Skipping of exon
9i | p53β | MRC-5 | (25) |
|
| TP53I3 | Skipping of exon
4 | TP53I3-TR | HEK293 | (40) |
The present review outlines the regulation of SRSF3
expression and its recently identified functions, as well as its
implications in cancer. Other biological functions of SRSF3, and
its regulatory mechanisms, are also summarized herein.
Regulatory mechanisms of SRSF3
expression
Expression of SRSF3 in cancers
SRSF3 was discovered to be aberrantly upregulated in
various tumor types, including colorectal cancer (CRC) (21), gastric cancer (GC), hepatocellular
carcinoma (HCC) (22), non-small
cell lung cancer (NSCLC), breast cancer (BC) and oral squamous cell
carcinoma (OSCC) (23). High
expression of SRSF3 has also been correlated with the occurrence
and development of cancer types such as CRC, bladder cancer and BC
(21,24–28).
Cells with high expression levels of SRSF3 demonstrate typical
characteristics of tumor cells, such as accelerated cell cycle
progression (21,24), enhanced anti-apoptotic ability,
pro-proliferation, increased tumorigenicity (24), aerobic glycolysis (29) and anti-senescence (25). Park et al (30) detected the expression of splicing
factors in GC and CRC tissues, and found that the expression rates
of SRSF3 (74%) and heterogeneous nuclear ribonucleoprotein A1 (88%)
were the highest in CRC tissues.
Regulation of SRSF3 expression
Mechanistic studies have revealed that hypoxia is an
activator of SRSF3 in PC3 prostate cancer cells (31). Hypoxia leads to retained intron
(RI)12 status of eukaryotic translation initiation factor 2B
subunit ε (EIF2B5) by promoting the expression of SRSF3 and
enhancing the specific binding of the RI to SRSF3 and Ser2 RNA
polymerase II (16). The RI of
EIF2B5 is a major regulator of translational initiation, which
contains a premature termination codon. This leads to premature
termination of protein translation and generates truncated EIF2B5
(EIF2B5-TR). EIF2B5-TR expression results in an increase in the
survival of head and neck cancer cells during hypoxia, facilitating
adaptation to the hypoxic state (16). In CRC, SRSF3 is upregulated via
signaling pathways involving WNT and catenin β1
(CTNNB1)/transcription factor 4 (32–34).
Allele T of single nucleotide polymorphism rs1122608 in the SMARCA4
gene was found to be associated with increased expression of SRSF3,
which reduces the expression and secretion of interleukin (IL)-1β,
thereby reducing the risk of atherosclerosis and stroke (35). In addition, SRSF3 expression is
significantly inhibited in M. tuberculosis H37Rv-infected
THP-1 macrophages (36). However,
the regulatory mechanisms of the rs1122608-T allele and H37Rv in
SRSF3 expression are remain unknown.
Xenobiotic regulation of SRSF3
Caffeine inhibits SRSF3 expression in HeLa cells and
converts p53α to p53β to promote cell cycle arrest and apoptosis
(37). Similarly, theophylline
downregulates SRSF3 expression in HeLa and MCF-7 cells, and
facilitates p53α-to-p53β conversion (38). Moreover, the study indicated that
theophylline induces apoptosis and senescence, and reduces cell
colony formation. Notably, theophylline inhibited cellular
proliferation, whereas caffeine enhanced proliferation rates;
neither theophylline nor caffeine had an effect on the
proliferation of normal breast cells (MCF-10A). These findings
suggest that theophylline and caffeine may act as antitumor
compounds by targeting SRSF3 and its target genes. In addition,
digoxin inhibits SRSF3 expression in HeLa cells and converts p53α
to p53β isoforms, thereafter, affecting the cell cycle and DNA
damage repair (39). In cells
derived from patients with familial dysautonomia, digoxin exerts a
therapeutic effect by inhibiting SRSF3 expression and increasing
the expression of wild-type elongator complex protein 1 (ELP1)
containing exon 20 (40). In
addition, digoxin exerts antiviral effects by inhibiting SRSF3
expression in HIV-1, altering viral RNA splicing sites, and leading
to loss of the essential viral factor Rev and the synthesis of
viral structural protein (41).
Trichostatin, a histone deacetylase inhibitor, acts on dendritic
cells by upregulating SRSF3, hypoxia inducible factor 1 subunit
α-dependent glycolytic genes and pyruvate kinase M2 (PKM2)
(42). These studies offer
insights into the pharmacodynamic roles of SRSF3 in cancer
therapy.
Splicing of SRSF3
In the late 1990s, Jumaa and Nielsen (43–45)
first demonstrated that SRSF3 regulated its own expression by
promoting the inclusion of exon 4, which contains a stop codon, and
consequently expresses SRSF3-TR. In OSCC cells, polypyrimidine
tract binding protein (PTBP) 1 and PTBP2 bind to exonic splicing
suppressors of SRSF3 exon 4 to inhibit its inclusion, which results
in the upregulation of full-length SRSF3 (SRSF3-FL) (46). Heterogeneous nuclear
ribonucleoprotein L has also been observed to promote the
transcription and skipping of exon 4 in SRSF3 (23). A splicing regulator, SLU7, promotes
exon 4 skipping in SRSF3 (47) and
plays an important role in hepatocellular differentiation (48). SLU7 is also a key gene in
hepatocyte homeostasis, the expression of which is inhibited in
liver cancer. SLU7-knockdown in human hepatocytes results in
impaired glucose and lipid metabolism, generation of key metabolic
hormone tolerance, and reversal of fetal-like gene expression
patterns by increasing the expression of SRSF3-TR (47,48).
Amiodarone has been reported to promote the inclusion of the SRSF3
stop codon and consequent SRSF3-TR expression and apoptosis of
tumor cells (49). Treatment of
human colon cancer cells with sodium arsenite also increases the
incidence of the SRSF3 termination codon and consequently increases
SRSF3-TR expression by inhibiting nonsense-mediated mRNA decay
(50). Knockout of SRSF3-TR
significantly decreases the arsenite-induced JUN protein and the
binding and promotive activity of the transcription factor JUN in
the IL-8 promoter, thereby significantly decreasing IL-8 expression
(50). Moreover, the inclusion of
SRSF3 exon 4 is also induced by the overexpression of circ87MARCA5
in U87MG cells, which consequently inhibits the proliferation and
migration of U87MG cells (51).
Phosphorylation of SRSF3
The localization and activity of SRSF3 are
stimulated by phosphorylation of its C-terminal RS domain. The
conserved SR Protein Kinase (SRPK)-specific substrate docking
groove in SRPK2 affects the phosphorylation and localization of
SRSF3. SRPK-mediated SRSF3 phosphorylation sites extend throughout
the RS domain. In addition, hypothalamic gonadotropin-releasing
hormone (GnRH) and AKT have also been identified as inducers of
SRSF3 phosphorylation (52,53).
Thus, inhibition of SRSF3 can reverse the GnRH-elevated
Homer1a-to-Homer1b/c ratio and regulate the effect of GnRH on the
expression of follicle stimulating hormone β and luteinizing
hormone subunit β (52). In rat
primary hepatocytes, insulin also promotes the phosphorylation of
SRSF3, which in turn bonds to the ESE in exon 12 of
glucose-6-phosphate dehydrogenase (G6PD), resulting in increased
splicing and expression of G6PD (54). Overexpression of protein
phosphatase, Mg2+/Mn2+ dependent 1G (PPM1G)
promotes the dephosphorylation of SRSF3 and changes the alternative
splicing patterns of genes related to the cell cycle and
transcriptional regulation in HCC cells (55). SRSF3 is phosphorylated at AKT
consensus sites downstream of PI3K-mediated platelet-derived growth
factor receptor α signaling in mouse palatal mesenchymal cells,
resulting in its nuclear translocation (56). However, SRSF3 appears to be
hypo-phosphorylated and stable in cells (57), which may be a way to maintain cell
homeostasis.
Oncogenic roles of SRSF3
Promotion of tumorigenesis
Studies have reported that SRSF3-mediated aberrant
splicing is closely associated with hepatocarcinogenesis (47,58).
SLU7 is a key factor in genomic stability; SLU7-knockdown results
in transcription-related RNA-DNA hybrids (R-loops), DNA damage,
cell cycle arrest and severe mitotic disorders, as well as loss of
sister chromatid cohesion, through the production of SRSF3-TR and
abnormal splicing, and expression of SRSF1 and the key sister
chromatid cohesion protein cell division cycle associated 5
(47). This unique function of
SLU7 has been observed in cancer cells and the livers of normal
mice, demonstrating the conserved and essential role of SLU7 in
preserving genomic integrity. Therefore, low expression of SLU7 in
the cirrhotic liver may be involved in the development of
hepatocarcinogenesis (47). RNA
sequencing revealed that 50% of HCC tissues had aberrant splicing
of the coiled-coil domain of coiled-coil domain containing 50
(CCDC50). Overexpression of truncated CCDC50 (CCDC50-TR) promotes
HCC growth and metastasis by activating the RAS/FOXO4 signaling
pathway; inhibition of MAP2K7/MAPK1 phosphorylation or
overexpression of FOXO4 significantly attenuates the
CCDC50-TR-mediated phenotype. SRSF3 directly binds to CCDC50-TR
mRNA to maintain its stability in the cytoplasm (58). The retention of SRSF3 in the
cytoplasm is mediated by the interaction between HBV-encoded X
protein (HBx) and tyrosine 3-monooxygenase/tryptophan
5-monooxygenase activation protein β (YWHAB). HBx upregulation
induces the expression of SRSF3 and CCDC50-TR in the cytoplasm.
Therefore, the HBx/SRSF3/YWHAB complex upregulates CCDC50-TR and
promotes the progression of HCC via the RAS/FOXO4 signaling pathway
(58).
The cell adhesion molecule CD44 is also a target
gene of SRSF3. SRSF3 alternatively splices exons 8, 9 and 10 of
CD44 to generate isoform CD44E, which promotes tumorigenesis in BC
and GC (32,59). protein phosphatase 1 regulatory
inhibitor subunit 1B (PPP1R1B) regulates the stability of SRSF3
protein and forms a complex with SRSF3 to regulate the expression
of CD44E (60). Knockdown of
PPP1R1B significantly downregulates CD44E expression in GC cells,
and decreases the expression of SRSF3 and CD44E in transplanted
tumors. In addition, SRSF3 is one of the major regulators of
induced pluripotent stem cell reprogramming and plays an important
role in dedifferentiation of tumorigenesis (61). SRSF5, a novel oncogenic splicing
factor, is overexpressed in OSCC tissues and cells and is critical
for the proliferation and tumorigenesis of OSCC cells (62). Overexpression of SRSF5 transforms
immortal rodent fibroblasts to form tumors in nude mice, while
downregulation of SRSF5 in OSCC cells impedes proliferation, cell
cycle progression, and tumor growth. SRSF3 is also upregulated in
OSCC tissues, especially in patients with lymphatic metastasis
(62). SRSF3 induces poison
cassette usage and/or intron 5 retention (63) or inhibits the usage of the proximal
3′-splice site in exon 6 of SRSF5, which disrupts SRSF5-mediated
self-regulation and results in the upregulation of full-length
SRSF5 in cancer cells (62).
Knockdown of SRSF3 inhibits snail family transcriptional repressor
1 and cadherin 2 (CDH2) expression, while the expression of SRSF3
is induced in primary cultured oral epithelial cells by the
carcinogen dimethylol butanoic acid, suggesting that SRSF3 is
involved in the occurrence of oral cancer (64).
Promotion of cellular
proliferation
The oncogene HER2 (erb-b2 receptor tyrosine kinase
2) is overexpressed in 20–30% of invasive BC and is associated with
poor patient prognosis. SRSF3 specifically binds to HER2 exon 15
and generates the oncoprotein Δ16HER2 (with exclusion of exon 16).
Downregulation of SRSF3 results in decreased expression of Δ16HER2
and increased expression of subtypes with inclusion of exon 15a or
intron 15, thereby inhibiting the proliferation of BC cells
(65). SRSF3 also promotes the
proliferation of cancer cells by splicing interleukin enhancer
binding factor 3 (ILF3), of which isoform-1 and −2 induce
proliferation and transformation, whereas isoform-5 and −7 inhibit
proliferation and induce apoptosis in tumor cells. SRSF3 directly
binds to the RNA motif of ILF3, resulting in exclusion/inclusion
of, or 3-terminal alternative splicing of, exon 18. Tumor cells
with low expression of SRSF3 generate ILF3 isoform-5 and −7,
thereby inhibiting the proliferation of tumor cells (66). In a variety of human cancers, SRSF3
promotes the inclusion of MDM4 exon 6 and results in increased
expression of full-length MDM4, an inhibitor of p65 (67). Thus, antisense
oligonucleotide-mediated MDM4 exon 6 skipping significantly
inhibits MDM4 expression and exerts good antineoplastic activity in
a variety of human melanoma cells and patient-derived melanoma
xenografts, as well as enhancing the sensitivity of tumors to
MAPK-targeting therapeutics (67).
Knockdown of SRSF3 in ovarian cancer cells affects homologous
recombination-mediated DNA repair activity and increases the
expression of the DNA double-strand break marker H2A.X variant
histone. Moreover, the splicing pattern of lysine methyltransferase
2C, an H3K4-specific histone methyltransferase, is altered,
resulting in decreased expression of mono-methylated and
tri-methylated H3K4 (68). SRSF3
also increases the expression of PKM2 by promoting inclusion of
exon 10 in PKM2. However, SRSF3-knockout increases PKM1 expression,
decreases the generation of lactic acid, and consequently inhibits
cellular proliferation, suggesting that SRSF3 also promotes
proliferation by regulating cancer cell metabolism (61). Various studies have indicated that
microRNAs (miRNAs/miRs) are involved in the SRSF3-mediated
promotion of cancer cell proliferation. For instance, knockdown of
SRSF3 results in decreased RE1 silencing transcription factor and
increased miR-132-3p and miR-212-3p expression, which directly
inhibit Yes1 associated transcriptional regulator and subsequent
cyclin D1 (CCND1), thereby suppressing the proliferation of cancer
cells (69). Knockdown of SRSF3
also decreases miR-1908-5p expression by inhibiting the
transactivation of NF-κB. miR-1908-5p directly inhibits NFKB
inhibitor interacting Raslike 2 (NKIRAS2) expression, a negative
regulator of the NF-κB pathway. Silencing NKIRAS2 inhibits NFKB
inhibitor β and promotes NF-κB translocation into the nucleus,
consequently activating miR-1908-5p and promoting tumor cell
proliferation (70).
Promotion of the cell cycle
SRSF3 is associated with the formation of chromatin
before and after mitosis, and is involved in regulation of the
structure and function of chromatin, demonstrating the important
roles of SRSF3 in the cell cycle (71). It has been reported that SRSF3
promotes cell cycle progression from the G2 to the M
phase by regulating transcription factor forkhead box M1 and its
downstream target genes, polo like kinase 1 and cell division cycle
25B (24). Therefore, inhibition
of SRSF3 in tumor cells leads to cell cycle arrest (21,24).
RAC1 is a small GTPase of the Rho family, which is involved in
tumor cell proliferation and invasiveness by regulating
actin-dependent cell motility and gene transcription. In CRC cells,
SRSF3 alternatively splices exon 3b of RAC1 to generate the isoform
RAC1b, so as to maintain tumor cell viability and promote tumor
progression (33). Knockdown of
SRSF3 in CRC cells decreases the expression of CCND1, CCND3, CCNE1,
E2F1 and E2F7, and consequently prevents cell progression from
G1 to S phase. Ajiro et al (59) investigated RNA splicing and gene
expression regulated by SRSF3 in tumor cells at the genome-wide
level, and revealed that SRSF3 regulated the expression of ≥224
genes, most of which are associated with the cell cycle or
proliferation. Spliced exons are rich in two binding motifs of
SRSF3, CCAGC(G)C and A(G)CAGCA. In addition, SRSF3 significantly
regulates the expression of at ≥20 miRNAs, including a subset of
oncogenic or tumor-suppressive miRNAs, indicating that SRSF3
maintains cellular homeostasis by affecting overall changes in gene
expression.
Promotion of tumor metastasis
Studies have supported that SRSF3 facilitates the
migration and invasiveness of CRC cells. Torres et al
(72) detected the protein
expression profile of two pairs of cell lines, KM12SM/KM12C and
SW620/SW480, indicating that SRSF3 was highly expressed in
metastatic KM12SM and SW620 cells. Knockout of SRSF3 significantly
inhibited the adhesion, survival, proliferation, migration and
liver homing abilities of KM12 cells. The observation that SRSF3
expression in CRC stem cells is significantly higher than in
non-cancer stem cells further suggests the involvement of SRSF3 in
metastasis (34). Lin et al
(73) discovered that SRSF3
induces CRC cell metastasis through a RBM4-SRSF3-MAP4K4 axis and
consequent MAPK8 signaling pathway. SRSF3 promotes the inclusion of
MAP4K4 exon 16 in CRC cells, which activates the phosphorylation of
MAPK8 and subsequent expression of CDH2 and vimentin. Moreover, the
authors found that expression deficiency of SRSF3 was significantly
associated with poor overall and disease-free-survival in patients.
Previous studies have also shown that high expression of GR (NR3C1)
in triple-negative breast cancer (TNBC) is associated with
chemoresistance and increased mortality rates. receptor for
activated C kinase 1 (RACK1) is a scaffold protein with a
glucocorticoid response element site on its promoter, which
participates in the migration and invasiveness of BC cells. SRSF3
alternatively splices GR to produce GRα, which regulates RACK1
expression at the transcriptional level and is involved in cell
migration (74,75). In addition, SRSF3-mediated splicing
of GR is positively regulated by cortisol (74,75).
Clinical data indicate that TAR DNA binding protein (TARDBP) is
highly expressed in TNBC and is associated with poor patient
prognosis. Knockout of TARDBP inhibits the proliferation and
metastasis of tumor cells, while its overexpression promotes
proliferation and malignancy in mammary epithelial cells. As an
important splicing regulator, TARDBP is responsible for the unique
splicing profile in TNBC (76).
TARDBP regulates gene splicing by interacting with SRSF3. For
instance, knockdown of TARDBP or SRSF3 prevents the inclusion of
NUMB endocytic adaptor protein exon 12 and thereby inhibits the
metastasis and proliferation of TNBC cells (76). The intensity and prevalence of
SRSF3 expression in invasive epithelial ovarian cancer cells are
significantly higher than those in tumor lesions, suggesting that
SRSF3 is associated with tumor cell metastasis. This was confirmed
by in vitro experiments that silence SRSF3, significantly
inhibiting the proliferation and metastasis abilities of U2OS and
HeLa cells (77).
Inhibition of apoptosis via programmed
cell death 4 (PDCD4)
It has been reported that SRSF3 promotes tumor
progression by inhibiting the expression of the pro-apoptotic
protein PDCD4 (60). SRSF3
inhibits alternative splicing and nuclear export of the minor
isoform of PDCD4 mRNA, as well as the translation of the major
isoform. Thus, the absence of PDCD4 isoforms in various cancers may
be associated with the high expression of SRSF3 (78). Moreover, silencing of SRSF3 leads
to a significant increase in PDCD4 mRNA in translated polysome
fractions, whereas overexpression of SRSF3 induces the
transformation of PDCD4 mRNA to non-translational ribosomal
components (78). Another study
demonstrated that SRSF3 co-localizes with PDCD4 mRNA in the
cytoplasmic processing corpuscle of live cells. SRSF3 directly
binds to the 5′ untranslated region (UTR) of PDCD4 and inhibits its
expression (29). Moreover, the
expression of anti-apoptotic proteins BCL2 and homeodomain
interacting protein kinase 2 are also inhibited by SRSF3 silencing,
thereby promoting the apoptosis of CRC cells (33).
Inhibition of cell senescence via
p53
Senescence is the ability of cells to enter an
irreversible proliferative stagnation state, and is an important
mechanism for preventing cancers. Through direct binding, SRSF3
alternatively splices TP53 to produce p53β, thereby preventing
p53-mediated cellular senescence (25). Down-regulation of SRSF3 in human
cells promotes cellular senescence through its alternative
splicing-dependent function, which is an important biological
process to prevent cancer and promote individual senescence
(25). Moreover, downregulation of
SRSF3 induces cellular senescence through alternative
polyadenylation (poly(A)) (79).
Knockout of SRSF3 results in preferential use of the proximal
polyadenylation site, leading to an overall shortening of the
3′-UTR of the senescence-associated mRNA. For instance, silencing
of SRSF3 induces the expression of PTEN, resulting in decreased AKT
phosphorylation and senescence-associated phenotypes (79). Astrocytes express both Δ133p53 and
p53β isoforms, which respectively show toxicity to, and protection
of, neurons. In vitro aging is caused by successive batches
of primary human astrocytes. Thus, SRSF3 can be expected to be a
therapeutic target for neurodegenerative diseases by reducing the
expression of Δ133p53 and increasing the expression of p53β
(80).
Inhibition of cell autophagy via p65
and FoxO1
Autophagy is an evolutionarily conserved cell
catabolic process. Dysfunction in the autophagy pathway has been
shown to be associated with numerous human disorders, including
cancer (81). SRSF3 is
significantly downregulated during hypoxia-induced autophagy in
OSCC cells. SRSF3-knockdown induces autophagy whereas SRSF3
overexpression inhibits hypoxia-induced autophagy. Mechanistic
investigation demonstrates that SRSF3 inhibits cell autophagy by
suppressing the expression of p65 and FoxO1, as well as their
downstream target gene beclin 1, a key regulator of autophagy
(82).
Other functions of SRSF3
SRSF3 in non-cancerous diseases
Spinal muscular atrophy (SMA) is a major genetic
factor in pediatric death and is caused by dysregulation of
survival of motor neuron (SMN) proteins, of which SMN2 with
exclusion of exon 7 is regulated by SRSF3 and encodes a
function-deficient protein. Knockout of SRSF3 increases the
inclusion of SMN2 exon 7 and the expression level of SMN protein,
providing a possible therapeutic approach for SMA (83). The p53 isoforms Δ133p53 and p53β
are expressed in astrocytes and regulate the toxicity and
protection of neurons. SRSF3 decreases Δ133p53 and increases p53β,
which significantly increases the number of senescent astrocytes
and senescence-like astrocytes in the brain tissues of patients
with Alzheimer's disease and amyotrophic lateral sclerosis
(80). Moreover, SRSF3 promotes
the development of microtubule associated protein τ (MAPT)
proteinosis by inducing skipping of the MAPT exon 10 (84). SRSF3 can also cause the
muscle-specific symptoms of hereditary myopathy with lactic
acidosis via erroneous splicing of mutant iron-sulfur cluster
assembly enzyme (85). Knockout of
the Srsf3 gene in mouse hepatocytes results in destruction of the
liver structure, growth retardation before and after birth,
impaired hepatocyte maturation, and changes in glucose and lipid
homeostasis that are manifested as decreased glycogen storage,
fasting hypoglycemia, increased insulin sensitivity, and decreased
cholesterol synthesis. Multiple aberrant splicing events occur in
the Srsf3-knockout mouse liver, especially the aberrant splicing of
key regulators of glucose and lipid metabolism, such as Hnf1α,
Ern1, Hmgcs1, Dhcr7 and Scap genes (86). In addition, Srsf3 is critical to
maintaining the transcriptome integrity of mouse oocytes.
Conditional deletion of Srsf3 in mature germinal vesicle oocytes
significantly impairs germinal vesicle breakdown, and thus oocytes
are unable to enter into meiosis (87).
Interestingly, SRSF3 also regulates the
translational efficiency of viral RNA by interacting with the RNA
binding protein poly(rC) binding protein 2 (88). Viral mRNAs generated from
intronless genes are particularly unstable owing to degradation by
nuclear RNA exosomes. The viral RNA binding protein microtubule
associated protein RP/EB family member 2 (MAPRE2) stabilizes viral
mRNAs in the nucleus, and stimulates their export to the cytoplasm
and subsequent translation into proteins. In the absence of MAPRE2,
SRSF3 destabilizes viral mRNAs by interacting with RNA exosomes and
their adaptor complex NEXT (89).
SRSF3 also participates in the splicing of the human papillomavirus
(HPV) 18 pre-mRNAs (90). If
differentiated keratinocytes are infected by HPV16, E2 protein
stimulates SRSF3, L1 mRNA and capsid protein expression, and thus
controls late events in the viral life cycle (91). Among Kaposi's sarcoma-associated
herpesviruses, SRSF3 is a cellular cofactor involved in open
reading frame (ORF)57-mediated K8β splicing. In the absence of
ORF57, SRSF3 binds to the K8β intron and inhibits K8β splicing,
whereas knockdown of SRSF3 promotes K8β splicing. The N-terminal
half of ORF57 binds to the RNA recognition motif of SRSF3,
preventing the binding of SRSF3 to the K8β intron, thereby
attenuating the inhibitory effect of SRSF3 on K8β splicing
(92).
SRSF3 also plays a role in cardiac diseases. SRSF3
is vital for heart development and homeostasis.
Cardiomyocyte-specific loss of SRSF3 expression results in
decapping of critical mRNAs involved in cardiac contraction
(93). Moreover, a
cardiomyocyte-specific Srsf3 conditional knockout mouse model
showed changes in expression levels and alternative splicing of
several transcripts related to mitochondrial integrity and
oxidative phosphorylation. Thus, SRSF3 also regulates mitochondrial
integrity and function (94).
Involvement in miRNA maturation
The RNA-induced silencing complex (RISC), consisting
of drosha ribonuclease III (DROSHA) and DGCR8 microprocessor
complex subunit (DGCR8) dimer, is responsible for the splicing of
primary miRNA transcripts (pri-miRNA). DROSHA and DGCR8 localize
RISC by interacting with the basal UG motifs and the apical UGU
motifs of the pri-miRNA, respectively. SRSF3 binds to CNNC motifs,
which are located ~17 nt from the RISC binding sites, recruiting
DROSHA to the basal junction, thereby enhancing RISC activity and
facilitating pri-miRNA splicing (95). Fernandez et al (96) found that the genetic variation
G>A in pri-mir-30c-1 results in secondary RNA structure
rearrangement, which enhances the binding of SRSF3 to pri-mir-30c-1
and consequently, the level of miR-30c. The involvement of SRSF3 in
miRNA processes is also supported by the finding that knockdown of
SRSF3 results in increased miR-132-3p and miR-212-3p (69) and decreased miR-1908-5p (70). Moreover, Ajiro et al
(59) reported that at least 20
miRNAs were dysregulated by SRSF3 in tumor cells. However, the
mechanisms underlying the involvement of SRSF3 in the processing of
miRNA are still unclear.
Contribution to RNA transport
Apart from splicing activities, SRSF3 is also
involved in nuclear RNA export factor 1 (NXF1)-dependent RNA
transport from the nucleus to the cytosol (97–99).
For instance, SRSF3 directly binds to the transcript of the histone
H2A gene, promoting its transport from nucleus to cytoplasm
(100). YTHDC1, a recognition
protein of N6-methyladenosine, is also required in the process of
precursor mRNA transcripts in the oocyte nucleus through
interaction with SRSF3 (101). In
HeLa cells, YTHDC1 interacts with SRSF3 and promotes the binding of
RNA to SRSF3 and NXF1, thereby driving export of methylated mRNA
from the nucleus to the cytoplasm (102). In mice, SRSF3 binds to the core
pluripotent transcription factor nanog homeobox (NANOG) mRNA and
facilitates its transport from the nucleus to the cytoplasm. In the
absence of SRSF3, NANOG mRNA is sequestered in the nucleus and its
protein expression level is markedly reduced. In addition, SRSF3
also regulates alternative splicing of the translocator Nxf1 and
RNA regulators, as well as the steady-state level of mRNA that
encodes chromatin modifications (19). SRSF3 promotes the binding of NXF1
to mRNA and confers the sequence specificity of NXF1 to the last
exon of that RNA. Thus, the alternative splicing and poly(A) is
linked with NXF1-mediated mRNA transport through SRSF3, which
ensures the abundance of transcripts with different 3′-UTR lengths
in the cytoplasm (103).
Post-transcriptional regulation of
gene expression
SRSF3 is an important component of RNA granules,
such as stress granules and cytoplasmic processing corpuscles, and
plays an important role in post-transcriptional regulation in the
cytoplasm (104). Brugiolo et
al (17) determined the
binding sites of SRSF3 on target mRNAs and RNAs in chromatin, the
nucleoplasm and cytoplasmic subcellular fractions. The authors
found that SRSF3 bonded to RNA in both the nucleus and cytoplasm.
SRSF3 also participates in the poly(A) of mRNA, indicating its role
in recognizing terminal exons in mRNA precursor splicing (105). SRSF3 modulates the recognition of
poly(A) splicing sites in calcitonin-encoding gene calcitonin
related polypeptide α by disrupting the binding of cleavage
stimulation factor subunit 2 (105). In addition, Boutej et al
(106) found that SRSF3 inhibited
the translation of overexpressed mRNA by binding to its 3′-UTR.
Serum amyloid A3, and other genes that are regulated by
lipopolysaccharide, contain multiple binding sites for SRSF3 in
their 3′-UTRs. By inhibiting the translation of
lipopolysaccharide-stimulated genes, SRSF3 acts as a major
regulator of the innate immune responses in resident microglia.
Conclusions
SRSF3 is involved in the splicing of a variety of
key genes, and has versatile biological functions. SRSF3 promotes
the inclusion of exons of target mRNAs by binding to their ESE. The
activity of SRSF3 depends on the phosphorylation of its C-terminal
RS domain, which is regulated by SRPK, GnRH, PPM1G and insulin.
Abnormal expression of SRSF3 leads to aberrant gene splicing and
the subsequent occurrence and development of various diseases, such
as cancers, SMA, Alzheimer's disease, amyotrophic lateral
sclerosis, MAPT proteinosis and hereditary myopathy with lactic
acidosis. Studies have identified abnormally highly expressed SRSF3
in various tumor tissues, including CRC, GC, HCC, NSCLC, OSCC and
BC. SRSF3 expression is regulated by the WNT pathway, SLU7, and
hypoxia. The expression of SRSF3 is inhibited by xenobiotics, such
as caffeine, theophylline, digoxin and amiodarone. SRSF3
participates in the occurrence of tumors and promotes the
proliferation and metastasis capacity of tumor cells by inhibiting
apoptosis, senescence and autophagy, as well as promoting cellular
proliferation and cell cycle. Progression. Furthermore, SRSF3
promotes the invasiveness and metastasis of tumor cells. In
addition, SRSF3 is involved in biological processes such as RNA
transport, miRNA maturation, protein translation, heart development
and embryogenesis. Notably, several controversial functions of
SRSF3 complicate its pathological roles. For instance, contrary to
the anti-apoptosis role of SRSF3, it binds to exon 8 of caspase 2
(CASP2) pre-mRNA, promoting the skipping of exon 9 and upregulating
pro-apoptotic protein Casp-2L. Knockdown of SRSF3 induces the
inclusion of exon 9 and increases anti-apoptotic protein Casp-2S
(107). SRSF3 has also been
reported to inhibit fibrosis by regulating gene splicing, mitotic
splicing, and epithelial-mesenchymal transition (EMT) to prevent
the development of HCC (18).
Inhibition of SRSF3 increases the expression of insulin like growth
factor and INSR and aberrantly activates the mitotic signal and
promotes aberrant splicing and the expression of EMT genes.
Moreover, the WNT/CTNNB1 signal pathway is activated and MYC
expression is enhanced upon SRSF3 inhibition (18). Past studies suggest that the
regulatory roles of SRSF3 are cell-dependent and/or
disease-dependent. The function of SRSF3 in different tissue types
and diseases requires further elucidation. Antisense
oligonucleotide-mediated downregulation of SRSF3 makes OSCC and
breast cancer cells more sensitive to the chemotherapy drug
paclitaxel. Modulation of RNA splicing profiles in disease by
antisense oligonucleotides is another exciting therapeutic approach
that requires further investigation.
In conclusion, the present review provides an
overview of the expression, regulation and function of SRSF3, as
well as its mechanisms, especially the oncogenic roles. Since
numerous studies have confirmed the inhibitory role of
SRSF3-knockdown in tumor growth and metastasis, there is great
interest in developing SRSF3 inhibitors for cancer therapy.
Studying the molecular mechanisms of SRSF3-dependent signal
transduction is expected to reveal additional candidates and
therapeutic targets for cancer therapy.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 81773044), the Science and
Technology Special Project of Clinical Medicine in Jiangsu Province
(grant no. BL2014046) and the Social Development Project of Jiangsu
Province (grant no. BE2019657).
Availability of data and materials
Not applicable.
Authors' contributions
JX and YC collected the literature, wrote and
revised the manuscript. WW and JS reviewed and revised the
manuscript. All authors have read and approved the final
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang ET, Sandberg R, Luo S, Khrebtukova I,
Zhang L, Mayr C, Kingsmore SF, Schroth GP and Burge CB: Alternative
isoform regulation in human tissue transcriptomes. Nature.
456:470–476. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Baralle F and Giudice J: Alternative
splicing as a regulator of development and tissue identity. Nat Rev
Mol Cell Biol. 18:437–451. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnson JM, Castle J, Garrett-Engele P,
Kan Z, Loerch PM, Armour CD, Santos R, Schadt EE, Stoughton R and
Shoemaker DD: Genome-wide survey of human alternative pre-mRNA
splicing with exon junction microarrays. Science. 302:2141–2144.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scotti MM and Swanson MS: RNA mis-splicing
in disease. Nat Rev Genet. 17:19–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang J and Manley JL: Misregulation of
pre-mRNA alternative splicing in cancer. Cancer Discov.
3:1228–1237. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen J and Weiss WA: Alternative splicing
in cancer: Implications for biology and therapy. Oncogene. 34:1–14.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oltean S and Bates DO: Hallmarks of
alternative splicing in cancer. Oncogene. 33:5311–5318. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou Z, Licklider LJ, Gygi SP and Reed R:
Comprehensive proteomic analysis of the human spliceosome. Nature.
419:182–185. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Alzhanova-Ericsson AT, Sun X, Visa N,
Kiseleva E, Wurtz T and Daneholt B: A protein of the SR family of
splicing factors binds extensively to exonic Balbiani ring pre-mRNA
and accompanies the RNA from the gene to the nuclear pore. Genes
Dev. 10:2881–2893. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shepard PJ and Hertel KJ: The SR protein
family. Genome Biol. 10:2422009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
de la Mata M and Kornblihtt AR: RNA
polymerase II C-terminal domain mediates regulation of alternative
splicing by SRp20. Nat Struct Mol Biol. 13:973–980. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fontrodona N, Aubé F, Claude JB, Polvèche
H, Lemaire S, Tranchevent LC, Modolo L, Mortreux F, Bourgeois CF
and Auboeuf D: Interplay between coding and exonic splicing
regulatory sequences. Genome Res. 29:711–722. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiao W, Adhikari S, Dahal U, Chen YS, Hao
YJ, Sun BF, Sun HY, Li A, Ping XL, Lai WY, et al: Nuclear m(6)A
reader YTHDC1 regulates mRNA splicing. Mol Cell. 61:507–519. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ye F, Chen ER and Nilsen TW: Kaposi's
sarcoma-associated herpesvirus utilizes and manipulates RNA N
6-adenosine methylation to promote lytic replication. J
Virol. 91:e00466–e00417. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Corbo C, Orru S and Salvatore F: SRp20: An
overview of its role in human diseases. Biochem Biophys Res Commun.
436:1–5. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brady LK, Wang H, Radens CM, Bi Y,
Radovich M, Maity A, CIvan C, Ivan M, Barash Y and Koumenis C:
Transcriptome analysis of hypoxic cancer cells uncovers intron
retention in EIF2B5 as a mechanism to inhibit translation. PLoS
Biol. 15:e20026232017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brugiolo M, Botti V, Liu N,
Müller-McNicoll M and Neugebauer KM: Fractionation iCLIP detects
persistent SR protein binding to conserved, retained introns in
chromatin, nucleoplasm and cytoplasm. Nucleic Acids Res.
45:10452–10465. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sen S, Langiewicz M, Jumaa H and Webster
NJ: Deletion of serine/arginine-rich splicing factor 3 in
hepatocytes predisposes to hepatocellular carcinoma in mice.
Hepatology. 61:171–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ratnadiwakara M, Archer SK, Dent CI, Mozos
IR, Beilharz TH, Knaupp AS, Nefzger CM, Polo JM and Anko ML: SRSF3
promotes pluripotency through Nanog mRNA export and coordination of
the pluripotency gene expression program. Elife. 7:e374192018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jumaa H, Wei G and Nielsen PJ: Blastocyst
formation is blocked in mouse embryos lacking the splicing factor
SRp20. Curr Biol. 9:899–902. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kurokawa K, Akaike Y, Masuda K, Kuwano Y,
Nishida K, Yamagishi N, Kajita K, Tanahashi T and Rokutan K:
Downregulation of serine/arginine-rich splicing factor 3 induces G1
cell cycle arrest and apoptosis in colon cancer cells. Oncogene.
33:1407–1417. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yea S, Narla G, Zhao X, Garg R, Tal-Kremer
S, Hod E, Villanueva A, Loke J, Tarocchi M, Akita K, et al: Ras
promotes growth by alternative splicing-mediated inactivation of
the KLF6 tumor suppressor in hepatocellular carcinoma.
Gastroenterology. 134:1521–1531. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia R, Zhang S, Liu M, Zhang Y, Liu Y, Fan
M and Guo J: HnRNP L is important for the expression of oncogene
SRSF3 and oncogenic potential of oral squamous cell carcinoma
cells. Sci Rep. 6:359762016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jia R, Li C, McCoy JP, Deng CX and Zheng
ZM: SRp20 is a proto-oncogene critical for cell proliferation and
tumor induction and maintenance. Int J Biol Sci. 6:806–826. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang Y, Horikawa I, Ajiro M, Robles AI,
Fujita K, Mondal AM, Stauffer JK, Zheng ZM and Harris CC:
Downregulation of splicing factor SRSF3 induces p53beta, an
alternatively spliced isoform of p53 that promotes cellular
senescence. Oncogene. 32:2792–2798. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karthikeyan C, Amawi H, Viana AG, Sanglard
L, Hussein N, Saddler M, Ashby CR Jr, Moorthy NS, Trivedi P and
Tiwari AK: lH-Pyrazolo[3,4-b]quinolin-3-amine derivatives inhibit
growth of colon cancer cells via apoptosis and sub G1 cell cycle
arrest. Bioorg Med Chem Lett. 28:2244–2249. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim WJ, Lee SJ, Choi YD and Moon SK:
Decursin inhibits growth of human bladder and colon cancer cells
via apoptosis, G1-phase cell cycle arrest and extracellular
signal-regulated kinase activation. Int J Mol Med. 25:635–641.
2010.PubMed/NCBI
|
|
28
|
Morley KL, Ferguson PJ and Koropatnick J:
Tangeretin and nobiletin induce G1 cell cycle arrest but not
apoptosis in human breast and colon cancer cells. Cancer Lett.
251:168–178. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim J, Park RY, Chen JK, Kim J, Jeong S
and Ohn T: Splicing factor SRSF3 represses the translation of
programmed cell death 4 mRNA by associating with the 5′-UTR region.
Cell Death Differ. 21:481–490. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park WC, Kim HR, Kang DB, Ryu JS, Choi KH,
Lee GO, Yun KJ, Kim KY, Park R, Yoon KH, et al: Comparative
expression patterns and diagnostic efficacies of SR splicing
factors and HNRNPA1 in gastric and colorectal cancer. BMC Cancer.
16:3582016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bowler E, Porazinski S, Uzor S, Thibault
P, Durand M, Lapointe E, Rouschop KM, Hancock J, Wilson I and
Ladomery M: Hypoxia leads to significant changes in alternative
splicing and elevated expression of CLK splice factor kinases in
PC3 prostate cancer cells. BMC Cancer. 18:3552018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Goncalves V, Matos P and Jordan P: The
beta-catenin/TCF4 pathway modifies alternative splicing through
modulation of SRp20 expression. RNA. 14:2538–2549. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Goncalves V, Matos P and Jordan P:
Antagonistic SR proteins regulate alternative splicing of
tumor-related Rac1b downstream of the PI3-kinase and wnt pathways.
Hum Mol Genet. 18:3696–3707. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Corbo C, Orrù S, Gemei M, Noto RD,
Mirabelli P, Imperlini E, Ruoppolo M, Vecchio LD and Salvatore F:
Protein cross-talk in CD133+ colon cancer cells indicates
activation of the Wnt pathway and upregulation of SRp20 that is
potentially involved in tumorigenicity. Proteomics. 12:2045–2059.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiong X, Xu C, Zhang Y, Li X, Wang B, Wang
F, Yang Q, Wang D, Wang X, Li S, et al: BRG1 variant rs1122608 on
chromosome 19p13.2 confers protection against stroke and regulates
expression of pre-mRNA-splicing factor SFRS3. Hum Genet.
133:499–508. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang W, Niu C, Fu RY and Peng ZY:
Mycobacterium tuberculosis H37Rv infection regulates alternative
splicing in macrophages. Bioengineered. 9:203–208. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lu GY, Huang SM, Liu ST, Liu PY, Chou WY
and Lin WS: Caffeine induces tumor cytotoxicity via the regulation
of alternative splicing in subsets of cancer-associated genes. Int
J Biochem Cell Biol. 47:83–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang YL, Hsu YJ, Chen Y, Wang YW and
Huang SM: Theophylline exhibits anti-cancer activity via
suppressing SRSF3 in cervical and breast cancer cell lines.
Oncotarget. 8:101461–101474. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu GY, Liu ST, Huang SM, Chang YL and Lin
WS: Multiple effects of digoxin on subsets of cancer-associated
genes through the alternative splicing pathway. Biochimie.
106:131–139. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu B, Anderson SL, Qiu J and Rubin BY:
Cardiac glycosides correct aberrant splicing of IKBKAP-encoded mRNA
in familial dysautonomia derived cells by suppressing expression of
SRSF3. FEBS J. 280:3632–3646. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wong RW, Balachandran A, Ostrowski MA and
Cochrane A: Digoxin suppresses HIV-1 replication by altering viral
RNA processing. PLoS Pathog. 9:e10032412013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jiang H, Zhang S, Song T, Guan X, Zhang R
and Chen X: Trichostatin a protects dendritic cells against
oxygen-glucose deprivation via the SRSF3/PKM2/glycolytic pathway.
Front Pharmacol. 9:6122018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jumaa H, Guenet JL and Nielsen PJ:
Regulated expression and RNA processing of transcripts from the
Srp20 splicing factor gene during the cell cycle. Mol Cell Biol.
17:3116–3124. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jumaa H and Nielsen PJ: Regulation of
SRp20 exon 4 splicing. Biochim Biophys Acta. 1494:137–143. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jumaa H and Nielsen PJ: The splicing
factor SRp20 modifies splicing of its own mRNA and ASF/SF2
antagonizes this regulation. EMBO J. 16:5077–5085. 1997. View Article : Google Scholar
|
|
46
|
Guo J, Jia J and Jia R: PTBP1 and PTBP2
impaired autoregulation of SRSF3 in cancer cells. Sci Rep.
5:145482015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jimenez M, Urtasun R, Elizalde M, Azkona
M, Latasa MU, Uriarte I, Arechederra M, Alignani D, Bárcena-Varela
M, Álvarez-Sola G, et al: Splicing events in the control of genome
integrity: Role of SLU7 and truncated SRSF3 proteins. Nucleic Acids
Res. 47:3450–3466. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Elizalde M, Urtasun R, Azkona M, Latasa
MU, Goñi S, García-Irigoyen O, Uriarte I, Segura V, Collantes M,
Scala MD, et al: Splicing regulator SLU7 is essential for
maintaining liver homeostasis. J Clin Invest. 124:2909–2920. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chang YL, Liu ST, Wang YW, Lin WS and
Huang SM: Amiodarone promotes cancer cell death through elevated
truncated SRSF3 and downregulation of miR-224. Oncotarget.
9:13390–13406. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kano S, Nishida K, Kurebe H, Nishiyama C,
Kita K, Akaike Y, Kajita K, Kurokawa K, Masuda K, Kuwano Y, et al:
Oxidative stress-inducible truncated serine/arginine-rich splicing
factor 3 regulates interleukin-8 production in human colon cancer
cells. Am J Physiol Cell Physiol. 306:C250–C262. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Barbagallo D, Caponnetto A, Cirnigliaro M,
Brex D, Barbagallo C, D'Angeli F, Morrone A, Caltabiano R,
Barbagallo GM, Ragusa M, et al: CircSMARCA5 inhibits migration of
glioblastoma multiforme cells by regulating a molecular axis
involving splicing factors SRSF1/SRSF3/PTB. Int J Mol Sci.
19:4802018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Q, Chikina MD, Pincas H and Sealfon
SC: Homer1 alternative splicing is regulated by
gonadotropin-releasing hormone and modulates gonadotropin gene
expression. Mol Cell Biol. 34:1747–1756. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Blaustein M, Pelisch F, Tanos T, Muñoz MJ,
Wengier D, Quadrana L, Sanford JR, Muschietti JP, Kornblihtt AR,
Cáceres JF, et al: Concerted regulation of nuclear and cytoplasmic
activities of SR proteins by AKT. Nat Struct Mol Biol.
12:1037–1044. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Walsh CM, Suchanek AL, Cyphert TJ, Kohan
AB, Szeszel-Fedorowicz W and Salati LM: Serine arginine splicing
factor 3 is involved in enhanced splicing of glucose-6-phosphate
dehydrogenase RNA in response to nutrients and hormones in liver. J
Biol Chem. 288:2816–2828. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen D, Zhao Z, Chen L, Li Q, Zou J and
Liu S: PPM1G promotes the progression of hepatocellular carcinoma
via phosphorylation regulation of alternative splicing protein
SRSF3. Cell Death Dis. 12:7222021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Dennison B, Larson ED, Fu R, Mo J and
Fantauzzo KA: Srsf3 mediates alternative RNA splicing downstream of
PDGFRα signaling in the facial mesenchyme. Development.
148:dev1994482021. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Long Y, Sou WH, Yung KW, Liu H, Wan SW, Li
Q, Zeng C, Law CO, Chan GH, Lau TC and Ngo JC: Distinct mechanisms
govern the phosphorylation of different SR protein splicing
factors. J Biol Chem. 294:1312–1327. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen H, Gao F, He M, Ding XF, Wong AM, Sze
SC, Yu AC, Sun T, Chan AWH, Wang X and Wong N: Long-read RNA
sequencing identifies alternative splice variants in hepatocellular
carcinoma and tumor-specific isoforms. Hepatology. 70:1011–1025.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ajiro M, Jia R, Yang Y, Zhu J and Zheng Z:
A genome landscape of SRSF3-regulated splicing events and gene
expression in human osteosarcoma U2OS cells. Nucleic Acids Res.
44:1854–1870. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhu S, Chen Z, Katsha A, Hong J, Belkhiri
A and El-Rifai W: Regulation of CD44E by DARPP-32-dependent
activation of SRp20 splicing factor in gastric tumorigenesis.
Oncogene. 35:1847–1856. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kuranaga Y, Sugito N, Shinohara H, Tsujino
T, Taniguchi K, Komura K, Ito Y, Soga T and Akao Y: SRSF3, a
splicer of the PKM gene, regulates cell growth and maintenance of
cancer-specific energy metabolism in colon cancer cells. Int J Mol
Sci. 19:30122018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yang S, Jia R and Bian Z: SRSF5 functions
as a novel oncogenic splicing factor and is upregulated by oncogene
SRSF3 in oral squamous cell carcinoma. Biochim Biophys Acta Mol
Cell Res. 1865:1161–1172. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Änkö ML, Müller-McNicoll M, Brandl H, Curk
T, Gorup C, Henry I, Ule J and Neugebauer KM: The RNA-binding
landscapes of two SR proteins reveal unique functions and binding
to diverse RNA classes. Genome Biol. 13:R172012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Peiqi L, Zhaozhong G, Yaotian Y, Jun J,
Jihua G and Rong J: Expression of SRSF3 is correlated with
carcinogenesis and progression of oral squamous cell carcinoma. Int
J Med Sci. 13:533–539. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gautrey H, Jackson C, Dittrich AL, Browell
D, Lennard T and Tyson-Capper A: SRSF3 and hnRNP H1 regulate a
splicing hotspot of HER2 in breast cancer cells. RNA Biol.
12:1139–1151. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Jia R, Ajiro M, Yu L, McCoy P Jr and Zheng
ZM: Oncogenic splicing factor SRSF3 regulates ILF3 alternative
splicing to promote cancer cell proliferation and transformation.
RNA. 25:630–644. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Dewaele M, Tabaglio T, Willekens K, Bezzi
M, Teo SX, Low DH, Koh CM, Rambow F, Fiers M, Rogiers A, et al:
Antisense oligonucleotide-mediated MDM4 exon 6 skipping impairs
tumor growth. J Clin Invest. 126:68–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
He X and Zhang P: Serine/arginine-rich
splicing factor 3 (SRSF3) regulates homologous
recombination-mediated DNA repair. Mol Cancer. 14:1582015.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kim HR, Hwang SJ, Shin CH, Choi KH, Ohn T
and Kim HH: SRSF3-regulated miR-132/212 controls cell migration and
invasion by targeting YAP1. Exp Cell Res. 358:161–170. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kim HR, Shin CH, Lee H, Choi KH, Nam DH,
Ohn T and Kim HH: MicroRNA-1908-5p contributes to the oncogenic
function of the splicing factor SRSF3. Oncotarget. 8:8342–8355.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Loomis RJ, Naoe Y, Parker JB, Savic V,
Bozovsky MR, Macfarlan T, Manley JL and Chakravarti D: Chromatin
binding of SRp20 and ASF/SF2 and dissociation from mitotic
chromosomes is modulated by histone H3 serine 10 phosphorylation.
Mol Cell. 33:450–461. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Torres S, García-Palmero I, Marín-Vicente
C, Bartolomé RA, Calviño E, Fernández-Aceñero MJ and Casal JI:
Proteomic characterization of transcription and splicing factors
associated with a metastatic phenotype in colorectal cancer. J
Proteome Res. 17:252–264. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lin JC, Lee YC, Tan TH, Liang YC, Chuang
HC, Fann YC, Johnson KR and Lin YJ: RBM4-SRSF3-MAP4K4 splicing
cascade modulates the metastatic signature of colorectal cancer
cell. Biochim Biophys Acta Mol Cell Res. 1865:259–272. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Buoso E, Ronfani M, Galasso M, Ventura D,
Corsini E and Racchi M: Cortisol-induced SRSF3 expression promotes
GR splicing, RACK1 expression and breast cancer cells migration.
Pharmacol Res. 143:17–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Buoso E, Galasso M, Ronfani M, Serafini
MM, Lanni C, Corsini E and Racchi M: Role of spliceosome proteins
in the regulation of glucocorticoid receptor isoforms by cortisol
and dehydroepiandrosterone. Pharmacol Res. 120:180–187. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ke H, Zhao L, Zhang H, Feng X, Xu H, Hao
J, Wang S, Yang Q, Zou L, Su X, et al: Loss of TDP43 inhibits
progression of triple-negative breast cancer in coordination with
SRSF3. Proc Natl Acad Sci USA. 115:E3426–E3435. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
He X, Arslan AD, Pool MD, Ho TT, Darcy KM,
Coon JS and Beck WT: Knockdown of splicing factor SRp20 causes
apoptosis in ovarian cancer cells and its expression is associated
with malignancy of epithelial ovarian cancer. Oncogene. 30:356–365.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Park SK and Jeong S: SRSF3 represses the
expression of PDCD4 protein by coordinated regulation of
alternative splicing, export and translation. Biochem Biophys Res
Commun. 470:431–438. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Shen T, Li H, Song Y, Li L, Lin J, Wei G
and Ni T: Alternative polyadenylation dependent function of
splicing factor SRSF3 contributes to cellular senescence. Aging
(Albany NY). 11:1356–1388. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Turnquist C, Horikawa I, Foran E, Major
EO, Vojtesek B, Lane DP, Lu X, Harris BT and Harris CC: p53
isoforms regulate astrocyte-mediated neuroprotection and
neurodegeneration. Cell Death Differ. 23:1515–1528. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Levy J, Towers C and Thorburn A: Targeting
autophagy in cancer. Nat Rev Cancer. 17:528–542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhou L, Guo J and Jia R: Oncogene SRSF3
suppresses autophagy via inhibiting BECN1 expression. Biochem
Biophys Res Commun. 509:966–972. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Wee CD, Havens MA, Jodelka FM and Hastings
ML: Targeting SR proteins improves SMN expression in spinal
muscular atrophy cells. PLoS One. 9:e1152052014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Yu Q, Guo J and Zhou J: A minimal length
between tau exon 10 and 11 is required for correct splicing of exon
10. J Neurochem. 90:164–172. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Rawcliffe DF, Österman L, Lindsten H and
Holmberg M: The high level of aberrant splicing of ISCU in
slow-twitch muscle may involve the splicing factor SRSF3. PLoS One.
11:e01654532016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Sen S, Jumaa H and Webster NJ: Splicing
factor SRSF3 is crucial for hepatocyte differentiation and
metabolic function. Nat Commun. 4:13362013. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Do DV, Strauss B, Cukuroglu E, Macaulay I,
Wee KB, Hu TX, Igor RD, Lee C, Harrison A, Butler R, et al: SRSF3
maintains transcriptome integrity in oocytes by regulation of
alternative splicing and transposable elements. Cell Discov.
4:332018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Bedard KM, Daijogo S and Semler BL: A
nucleo-cytoplasmic SR protein functions in viral IRES-mediated
translation initiation. EMBO J. 26:459–467. 2007. View Article : Google Scholar
|
|
89
|
Mure F, Corbin A, Benbahouche NE, Bertrand
E, Manet E and Gruffat H: The splicing factor SRSF3 is functionally
connected to the nuclear RNA exosome for intronless mRNA decay. Sci
Rep. 8:129012018. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Ajiro M, Tang S, Doorbar J and Zheng ZM:
Serine/arginine-rich splicing factor 3 and heterogeneous nuclear
ribonucleoprotein A1 regulate alternative rna splicing and gene
expression of human papillomavirus 18 through two functionally
distinguishable cis elements. J Virol. 90:9138–9152. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Klymenko T, Hernandez-Lopez H, MacDonald
AI, Bodily JM and Graham SV: Human papillomavirus E2 regulates
SRSF3 (SRp20) to promote capsid protein expression in infected
differentiated keratinocytes. J Virol. 90:5047–5058. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Majerciak V, Lu M, Li X and Zheng ZM:
Attenuation of the suppressive activity of cellular splicing factor
SRSF3 by Kaposi sarcoma-associated herpesvirus ORF57 protein is
required for RNA splicing. RNA. 20:1747–1758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ortiz-Sánchez P, Villalba-Orero M,
López-Olañeta MM, Larrasa-Alonso J, Sánchez-Cabo F, Martí-Gómez C,
Camafeita E, Gómez-Salinero JM, Ramos-Hernández L, Nielsen PJ, et
al: Loss of SRSF3 in cardiomyocytes leads to decapping of
contraction-related mRNAs and severe systolic dysfunction. Circ
Res. 125:170–183. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Dumont A, Dumont L, Zhou D, Giguère H,
Pileggi C, Harper ME, Blondin DP, Scott MS and Auger-Messier M:
Cardiomyocyte-specific Srsf3 deletion reveals a mitochondrial
regulatory role. FASEB J. 35:e215442021. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kim K, Nguyen TD, Li S and Nguyen TA:
SRSF3 recruits DROSHA to the basal junction of primary microRNAs.
RNA. 24:892–898. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Fernandez N, Cordiner RA, Young RS, Hug N,
Macias S and Cáceres JF: Genetic variation and RNA structure
regulate microRNA biogenesis. Nat Commun. 8:151142017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Huang Y, Gattoni R, Stévenin J and Steitz
JA: SR splicing factors serve as adapter proteins for TAP-dependent
mRNA export. Mol Cell. 11:837–843. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hautbergue GM, Hung ML, Golovanov AP, Lian
LY and Wilson SA: Mutually exclusive interactions drive handover of
mRNA from export adaptors to TAP. Proc Natl Acad Sci USA.
105:5154–5159. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Escudero-Paunetto L, Li L, Hernandez FP
and Sandri-Goldin RM: SR proteins SRp20 and 9G8 contribute to
efficient export of herpes simplex virus 1 mRNAs. Virology.
401:155–164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Huang Y and Steitz JA: Splicing factors
SRp20 and 9G8 promote the nucleocytoplasmic export of mRNA. Mol
Cell. 7:899–905. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Kasowitz SD, Ma J, Anderson SJ, Leu NA, Xu
Y, Gregory BD, Schultz RM and Wang PJ: Nuclear m6A reader YTHDC1
regulates alternative polyadenylation and splicing during mouse
oocyte development. PLoS Genet. 14:e10074122018. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Roundtree IA, Luo GZ, Zhang Z, Wang X,
Zhou T, Cui Y, Sha J, Huang X, Guerrero L, Xie P, et al: YTHDC1
mediates nuclear export of N6-methyladenosine methylated
mRNAs. Elife. 6:e313112017. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Muller-McNicoll M, Botti V, de Jesus
Domingues AM, Brandl H, Schwich OD, Steiner MC, Curk T, Poser I,
Zarnack K and Neugebauer KM: SR proteins are NXF1 adaptors that
link alternative RNA processing to mRNA export. Genes Dev.
30:553–566. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Yoon SP, Kim HH, Kim J, Park RY and Ohn T:
Regulation of cellular RNA nano-particle assembly by splicing
factor SRp20. J Nanosci Nanotechnol. 13:184–187. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Lou H, Neugebauer KM, Gagel RF and Berget
SM: Regulation of alternative polyadenylation by U1 snRNPs and
SRp20. Mol Cell Biol. 18:4977–4985. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Boutej H, Rahimian R, Thammisetty SS,
Béland LC, Lalancette-Hébert M and Kriz J: Diverging mRNA and
protein networks in activated microglia reveal SRSF3 suppresses
translation of highly upregulated innate immune transcripts. Cell
Rep. 21:3220–3233. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Jang HN, Lee M, Loh TJ, Choi SW, Oh HK,
Moon H, Cho S, Hong SE, Kim DH, Sheng Z, et al: Exon 9 skipping of
apoptotic caspase-2 pre-mRNA is promoted by SRSF3 through
interaction with exon 8. Biochim Biophys Acta. 1839:25–32. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Stickeler E, Kittrell F, Medina D and
Berget SM: Stage-specific changes in SR splicing factors and
alternative splicing in mammary tumorigenesis. Oncogene.
18:3574–3578. 1999. View Article : Google Scholar : PubMed/NCBI
|














