Introduction
Renal cell carcinoma (RCC) is the most common form
of kidney cancer. Its incidence has steadily risen over the past 10
years, and it accounts for 2–3% of all adult malignancies (1). RCC cells show high multidrug
resistance, which makes chemotherapy ineffective, and radiotherapy
can only relieve the symptoms (2).
RCC cells have a lower rate of cleavage than other cancer cells,
which is speculated to be a cause of resistance to chemotherapy and
radiotherapy (3,4). Due to the poor response of RCC to
radiotherapy and chemotherapy, the 5-year survival rate for
metastatic RCC is only 10% (5).
Previously, treatment of RCC was rare and was occasionally managed
by the patient's own immune system (2). For this reason, immunotherapy using
IFN-γ and IL-2 has been experimentally used for the treatment of
RCC (6–8). However, several clinical applications
of IFN-γ and IL-2 immunotherapy have shown that the overall
prognosis of patients remains poor, with only 20% of patients
responding for short periods of time; the therapy also induces
severe toxicity (9). Due to these
disappointing outcomes, the development of novel therapies for RCC
is urgent, and a number of studies are now being conducted
worldwide.
4-1BB (CD137) is expressed on activated T cells and
delivers co-stimulatory signals for T cell activation when it binds
to 4-1BB ligand or in ligation with agonistic anti-4-1BB monoclonal
antibodies (mAbs) (10,11). The systemic administration of
agonistic anti-4-1BB mAbs to tumor-bearing mice with P815
mastocytoma, AG104A sarcoma (12),
B10.2 fibrosarcoma (13) and CT26
colon carcinoma (14) caused tumor
regression, but they had no effect on weakly or poorly immunogenic
tumors, such as B16 melanoma, C3 tumors and TC-1 lung carcinoma
(15). Researchers have tested the
efficacy of combinations of anti-4-1BB mAbs and various reagents to
enhance the anticancer effects. Previously, using a murine RCC
tumor model, we showed that a combination of subtoxic doses of
fluorouracil (5-FU) and anti-4-1BB mAb eradicated established
tumors, while either 5-FU or anti-4-1BB mAb monotherapy did not
(16). In addition, immune
checkpoint blockade in combination with anti-VEGF or
anti-programmed cell death protein 1 (PD-1) have been found to
augment T cell-mediated antitumor immunity in metastatic RCC
(17,18). The expression of PD-1 on 4-1BB T
cells may be novel therapeutic targets for immunotherapy of
metastatic RCC (19,20).
Actinobacillus actinomycetemcomitans (A.
actinomycetemcomitans) is associated with several human
diseases, including endocarditis, meningitis, osteomyelitis,
subcutaneous abscesses and periodontal disease (21–25).
The immunostimulatory factor (ISTF; 13 kDa) isolated from A.
actinomycetemcomitans, has potent mitogenic activity on mouse B
cells and human peripheral blood mononuclear cells (26). ISTF has been reported to stimulate
macrophages and dendritic cells in the spleens of BALB/c mice and
also has the ability to induce the direct activation of mouse
macrophages to induce IL-6, TNF-α, nitric oxide and major
histocompatibility complex (MHC) class II expression (27). In addition, ISTF is a proteinaceous
material that directly induces the proliferation of B lymphocytes,
but does not affect the proliferation of T lymphocytes, even in the
presence of antigen-presenting cells (APCs) (26).
To fully activate T cells, both antigen recognition
(peptide and MHC complex) and co-stimulatory signals provided by
APCs are required (28). In the
absence of co-stimulatory signals, antigen presentation induces T
cell anergy, while co-stimulatory signals activate non-responding
tumor-specific T cells (29). ISTF
is expected to be highly active against APCs, and anticancer
activity can be expected to be increased by the amplification of
the overall immune response (27).
Therefore, in the present study it was hypothesized that a
combination of ISTF, promoting antigen presentation, and 4-1BB
mAbs, stimulating T cell co-stimulation signals, could increase T
cell activity to eradicate RCC. The aim of this study was to
evaluate the efficacy of a combination of ISTF and agonistic
anti-4-1BB mAbs in the RCC model, which is associated with
multidrug resistance and does not respond well to anticancer
therapy.
Materials and methods
Animals and reagents
Female Balb/c mice (7 weeks of age, 156 mice) were
purchased from Orient Bio, Inc. The mice were housed under specific
pathogen-free conditions at 18–24°C and 40–70% humidity in a 12 h
light-dark cycle, with ad libitum access to food and water.
Animal studies were approved by the University of Ulsan Animal Care
and Use Committee (approval no. HTC-14-030; Nam-gu, Republic of
Korea). All mice were subjected to anesthesia by tribromoethanol
[intraperitoneally (i.p.) injection, 250 mg/kg, 30 min] or
euthanasia by using a flow rate of 3–7 liters per min with
CO2 for a 10-liter volume. Hybridomas (Clone 3E1)
producing agonistic anti-4-1BB mAb were a gift from Dr Robert
Mittler (Emory University, Atlanta, GA, USA). Anti-CD3 monoclonal
antibody (cat. no. 557306), FITC-CD3 (cat. no. 553061), PE-CD8
(cat. no. 553032), FITC-CD8 (cat. no. 553030), PerCP-Cy™ 5.5-CD8
(cat. no. 551162), PE-B220 (cat. no. 553089), PerCP-Cy™5.5-CD4
(cat. no. 550954), PE-CD4 (cat. no. 557308), PE-Foxp3 (cat. no.
560408), PE-F4/80 (cat. no. 565410), PE-DX5 (cat. no. 553858),
PE-CD11b (cat. no. 557397), FITC-Gr1 (cat. no. 553126), PE-CD11c
(cat. no. 557401), FITC-CD11c (cat. no. 557400), PE-IFN-γ (cat. no.
562020) were purchased from BD Pharmingen (BD Biosciences). The
gene encoding an ISTF from Actinobacillus
actinomycetemcomitans was cloned into the pET-32a(+)
DNA-Novagen expression vector (Sigma-Aldrich; Merck KGaA). The
recombinant vector containing a full-length ISTF gene fused with a
C-terminal His6 tag was transformed into E. coli BL21 (DE3;
Stratagene; Agilent Technologies, Inc.). The expression of the
recombinant ISTF was induced by incubation with 1 mM isopropyl
β-D-1-thiogalactopyranoside (IPTG; Sigma-Aldrich; Merck KGaA) at
20°C for 4 h, and the recombinant ISTF was purified using a
Protino® Ni-TED column (Machery-Nagel GmbH) according to
the manufacturer's protocols.
Tumor cells and animal
experiments
Renca cells were purchased from the Korean Cell Line
Bank (Korean Cell Line Research Foundation) and cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.).
For the animal experiments, log-phase cells were washed and
resuspended in PBS immediately before injection into mice. Mice
weighing 20±0.5 g were subcutaneously (s.c.) injected with Renca
tumor cells into the site over the right flank
(1×106/mouse, 100 µl). ISTF was injected i.p. on days 3,
7 and 12, and anti-4-1BB mAb (3E1) was injected i.p. on days 7 and
12, while a control group received PBS and rat IgG (16,27).
The tumor diameter was measured every 2–3 days, and the tumor
volume (in mm3) was calculated using a caliper. The
tumor size was expressed as the tumor volume based on the following
formula: Tumor volume (mm3) × (major axis) × (minor
axis) × (height) ×0.52. The animals were sacrificed when the
longest dimensions of the tumors were >20 mm. Mice were
considered tumor free when the tumor dimensions were <1 mm; they
were kept under observation for at least 60 days. At least 6–10
mice/treatment group were examined throughout the day, and each
reported experiment was representative of at least three similarly
performed experiments.
Isolation of splenocytes
The murine spleen was placed in a petri dish with 5
ml Hanks' balanced salt solution buffer, and the spleen was cut
into small pieces (~0.2 cm2) with a scalpel blade. The
small pieces were crushed using the plunger end of a syringe and
then the cell suspensions were passed through a cell strainer.
After centrifugation (220 × g, 5 min, 4°C) the cell pellet was
suspended in 2–5 ml cold 1X RBC Lysis buffer (eBioscience; Thermo
Fisher Scientific, Inc.). After incubating the suspension for 5 min
on ice, cells were washed with 10–20 ml cold PBS. Cells
(1×106 cells/ml) were suspended in 1% BSA (Roche
Diagnostics) in PBS for fluorescence activated cell sorting (FACS)
analysis or in RPMI-1640 medium for cytokine analysis.
Isolation of tumor-infiltrating
lymphocytes (TILs) from tumor tissue
TILs were isolated from tumor tissues as described
previously (30) with minor
modifications. Briefly, tumors were excised (2–3 mm in width), and
the fragments were incubated in RPMI-1640 medium containing 10%
FBS, collagenase type I (300 U/ml; Gibco; Thermo Fisher Scientific,
Inc.) and DNase I (50 U/ml; Sigma-Aldrich; Merck KGaA) at 37°C for
90 min. Thereafter, the digested fragments were passed through
steel mesh, layered over superimposed layers of 54 and 63% Percoll
and centrifuged at 400 × g for 45 min at room temperature. TILs
were recovered at the interface between 54 and 63% of the Percoll
layers.
Surgical removal of tumor-draining
lymph nodes (TDLNs)
At day 14 after injection with ISTF and anti-4-BB
mAb, tumor-bearing mice were subjected to surgical removal of the
adjacent inguinal lymph nodes (3).
Hepatocellular damage assay
To detect serum alanine aminotransferase (ALT) and
aspartate aminotransferase (AST), serum was collected by
centrifugation at 585 × g for 15 min at 4°C from peripheral blood
obtained through the retro-orbital bleeding procedure. ALT and AST
activity, indicators of hepatocellular injury, were measured using
the Enzy-Chrom™ Alanine Transaminase Assay Kit (cat. no. EALT-100;
BioAssay Systems) and Enzy-Chrom™ Aspartate Transaminase Assay Kit
(cat. no. EASTR-100; BioAssay Systems).
FACS analysis
For FACS analysis, splenocytes and TILs were blocked
with the Fc receptor-blocking mAb 2.4G2 for 20 min at 4°C and
stained with 4G2, FITC-CD3, PE-CD4, PE-CD8, PE-DX5, PE-CD11b and
FITC-CD11c mAbs for 30 min at 4°C. After washing, they were
analyzed with a FACSCalibur flow cytometer (Becton, Dickinson and
Company). Data were analyzed by using FlowJo v10 software (FlowJo
LLC).
Determination of intracellular
cytokines
To measure the expression of IFN-γ, spleens were
isolated from Renca-bearing mice on day 14 and cultured with PMA
(50 ng/ml; Sigma-Aldrich; Merck KGaA) and ionomycin (500 ng/ml;
Sigma-Aldrich; Merck KGaA), and cytokine release was prevented by
treatment with Golgi-stop (BD Pharmingen; BD Biosciences).
Following surface staining for Cy-CD8 and FITC-C11c, the cells were
fixed in Cytofix/Cytoperm solution (BD Pharmingen; BD Biosciences)
for 30 min at 4°C and stained with PE-conjugated anti-mouse IFN-γ
for 30 min at 4°C (cat. no. 562333; BD Biosciences). Finally, they
were analyzed on a FACSCanto™ II (BD Biosciences) with FlowJo v10
software.
Cytokine analysis
Splenocytes (2×106) were stimulated with
CD3 mAb (0.1 µg/ml), anti-4-1BB mAb (5 µg/ml) and ISTF (10 µg/ml).
After 48 h of incubation, culture supernatants were collected. The
cytokines in the culture supernatants were quantified using a
cytometric bead array kit (BD Biosciences), according to the
manufacturer's protocol. They were analyzed on a FACSCanto™ II (BD
Biosciences) with FlowJo v10 software.
Statistical analysis
All experiments were conducted at least three times,
and the data are presented as the mean ± standard error of the
mean. All the data were analyzed using GraphPad Prism 9 (GraphPad
Software, Inc.). Statistical analyses were performed using one-way
ANOVA with Tukey's post hoc test, and Shapiro-Wilk test was
performed as a test of normality. Survival analysis was performed
using the Kaplan-Meier method; the log-rank test was used to
determine statistical significance. P<0.05 was considered to
indicate a statistically significant difference.
Results
Combined therapy with ISTF and
anti-4-1BB mAb eradicates Renca tumors
To observe whether combined therapy with ISTF and
anti-4-1BB mAb could eradicate Renca tumors, mice were treated with
100 µg ISTF and 100 µg anti-4-1BB mAb. Mice inoculated with Renca
tumors were injected with ISTF on days 3, 7 and 12 with anti-4-1BB
mAb on days 7 and 12, while the control group received PBS and rat
IgG. The tumors in the control mice grew, and these mice died from
tumor overgrowth by day 37. The tumor growth of the
4-1BB-mono-treated mice was slower than that of the control mice,
but the ISFF-mono-treated group was similar to the control group;
i.e., ISTF mono-treatment was ineffective. The combined treatment
of mice with ISTF and agonistic 4-1BB antibody had a greater
antitumor effect than ISTF or anti-4-1BB mAb alone, and 50% of mice
showed partial tumor regression between days 12 and 17 (Fig. 1A and B). In addition, ISTF alone or
4-1BB alone, or 4-1BB and ISTF co-administration did not induce
liver toxicity, as indicated by the levels of ALT and AST (Fig. 1C). The control and ISTF monotherapy
mice all died between days 31 and 37 after tumor cell inoculation.
Monotherapy with anti-4-1BB mAb showed modest antitumor effects.
The anti-4-1BB mAb monotherapy group had slower tumor growth and
survived longer than the control group, but eventually, these mice
died between 37 and 43 days. ISTF and anti-4-1BB mAb co-treatment
notably inhibited tumor growth (Fig.
1A) and increased the mouse survival time compared with the
other experimental groups. A total of 50% of the ISTF and
anti-4-1BB mAb co-treated mice survived until the end of the
experiment (Fig. 1D).
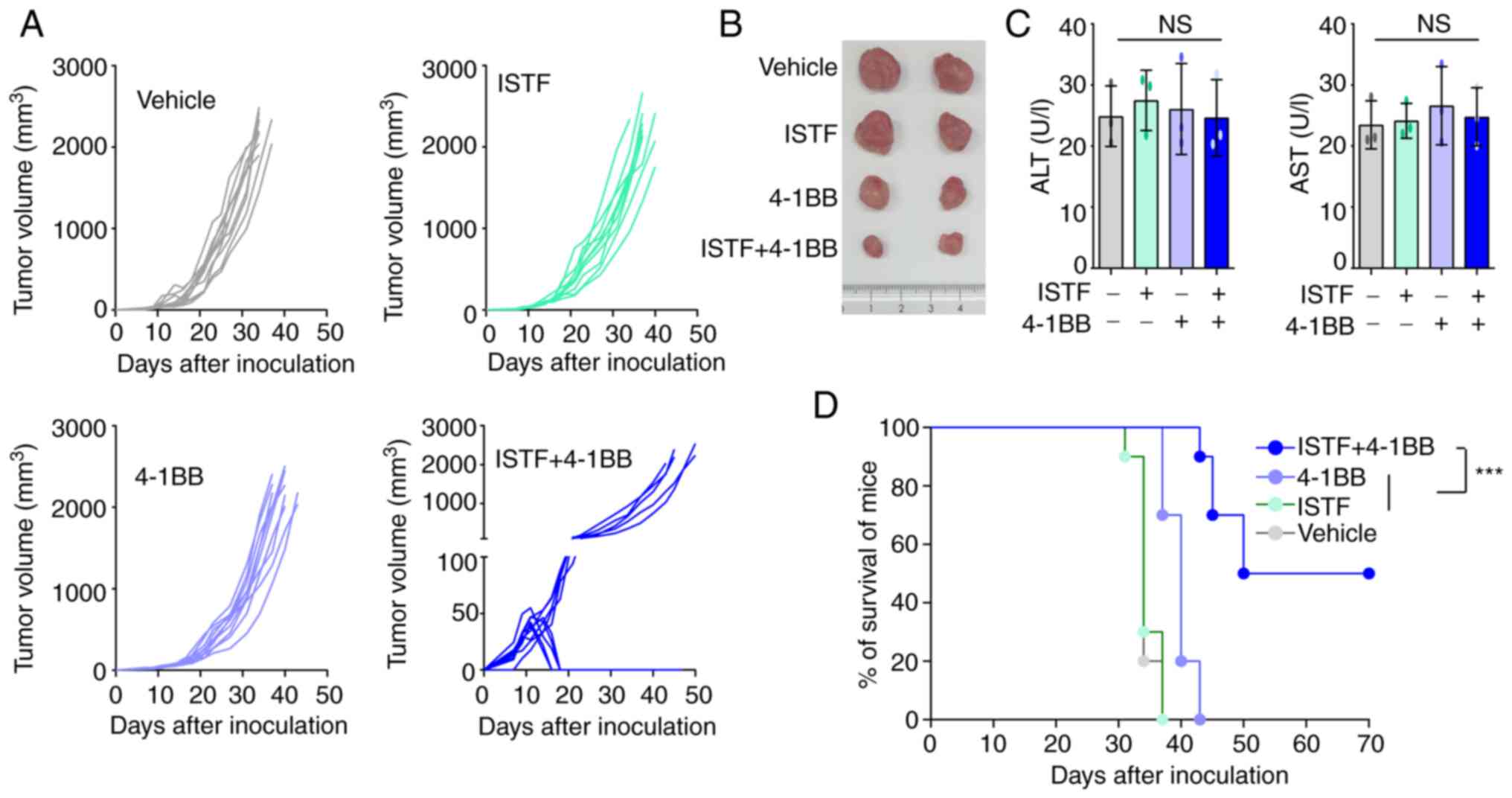 | Figure 1.Combined therapy with ISTF and
anti-4-1BB mAb has antitumor effects in mice inoculated with Renca
cells. Mice were inoculated with Renca tumor cells on day 0.
Tumor-bearing mice were divided into four groups and treated with
the following reagents: i) PBS and control mAb (rat IgG); ii) ISTF
monotherapy; iii) 4-1BB mAb monotherapy; and iv) ISTF and 4-1BB mAb
combined. Mice were subcutaneously injected with Renca tumor cells
(1×106/mouse). ISTF (100 µg/mouse) was injected i.p. on
days 3, 7 and 12, and anti-4-1BB mAb (100 µg/mouse) was injected
i.p. on days 7 and 12, while a control group received PBS and/or
rat IgG, respectively. Tumor diameter was measured every 2–3 days,
and tumor volume (in mm3) was calculated using a
caliper. Tumor size was expressed as tumor volume based on the
following formula: Tumor volume (mm3)=(major axis) ×
(minor axis) × (height) ×0.52. (A) Effect of combined therapy with
ISTF and anti-4-1BB mAb on Renca tumor volume. Each line indicates
the tumor volume of the individual animal. Results shown are
representative of three independent experiments. n, number of mice
per experiment. (B) Representative images of tumors on days 21 in
the same condition as (A). (C) Serum ALT and AST were measured. (D)
Survival rate of mice was determined. The animals were sacrificed
when the longest dimensions of the tumors were >20 mm.
Comparison of survival curves with log-rank test yielded a
statistical significance of ***P<0.001. NS, not significant;
ISTF, immunostimulatory factor; mAb, monoclonal antibody; i.p.,
intraperitoneally; ALT, alanine aminotransferase; AST, aspartate
aminotransferase. |
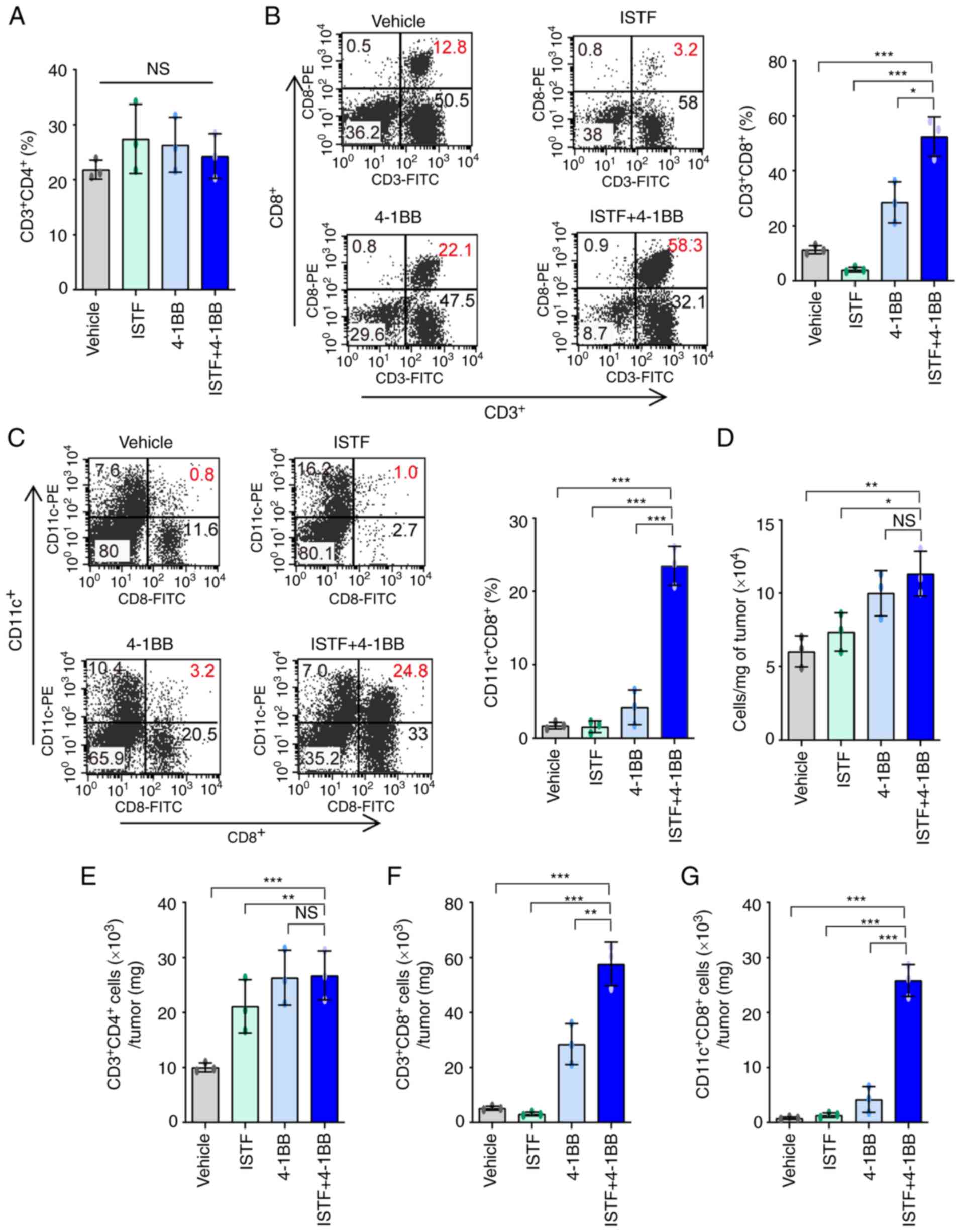
ISTF and anti-4-1BB mAb co-treatment
induces marked expansion of CD11c+CD8+ cells
in splenocytes of tumor-bearing mice
To analyze the relationship between tumor regression
and immune cell populations, the cell populations in the spleens of
mice bearing the Renca tumors were analyzed. As shown in Fig. 1A, ISTF and 4-1BB co-treatment
almost removed the tumor ~day 17. Thus, on day 14 after Renca tumor
cell inoculation, spleens were harvested by sacrificing the mice,
and the percentage of CD3+CD4+ T cells,
CD3+CD8+ T cells, B220+ B cells,
CD3+DX5+ natural killer (NK) cells,
CD11b+F480+ macrophages,
CD4+Fox3+ Treg cells and CD11c+
CD8+ T cells were analyzed by FACS. The proportion of
CD3+CD8+ T cells in the ISTF + 4-1BB group
was significantly higher than that in the control mice, but
comparable to that in the 4-1BB-monotherapy mice (Figs. 2A and S1A). The percentage of
CD11c+CD8+ T cells in the ISTF + 4-1BB group
showed the most prominent increase compared with that for either
treatment alone. These cells constituted 18% of the total spleen
cells of the ISTF and anti-4-1BB mAb co-treated mice, but only 1.1%
of those of the control IgG and PBS-treated mice. The percentage of
CD11c+CD8+ cells in the anti-4-1BB mAb
monotherapy group was 4.9%, higher than that in the control group,
but lower than that in the ISTF + 4-1BB group. Even with 4-1BB
treatment alone, the percentage of CD11c+CD8+
cells increased 4.5-fold compared with the control group, but the
increase was an 16-fold increase in the ISTF + 4-1BB group compared
with the control group (Figs. 2B
and S1B). The percentages of
CD4+Fox3+ Treg cells (Figs. 2C and S1C), CD11b+F480+
macrophages (Figs. 2D and S2D), CD3+CD4+ T
cells (Figs. 2E and S1E), CD3+DX5+ NK
cells (Figs. 2F and S1F) and CD11b+Gr1inT cells
(Figs. 2G and S1G) showed no significant differences
between groups. However, the percentage of B220+ B cells
(Figs. 2H and S1H) was decreased in the ISTF + 4-1BB
group compared with the control group. These results showed that
combination therapy with ISTF and anti-4-1BB mAb had a synergistic
effect on the increase in CD8+CD11c+ T cells
in tumor-bearing mice.
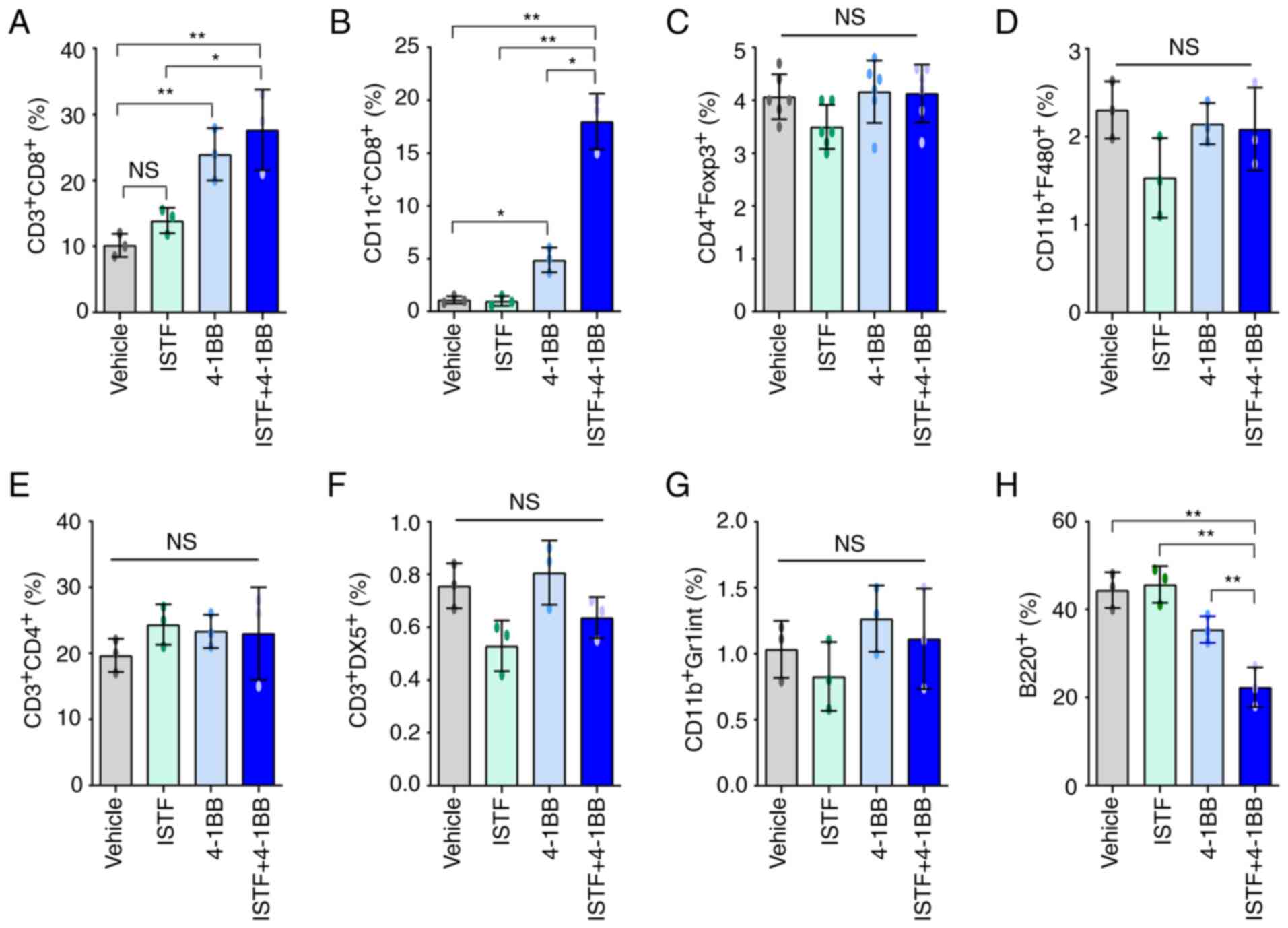 | Figure 2.ISTF and anti-4-1BB mAb co-treatment
induces marked expansion of CD11c+CD8+ T
cells in splenocytes of tumor-bearing mice. Renca tumors were
established and four groups of mice were treated with ISTF and/or
anti-4-1BB mAb as described in Fig.
1. On day 14, splenocytes were excised; double stained with
PE-, Cy- and FITC-conjugated antibodies and analyzed by
fluorescence activated cell sorting. Bar charts indicating the
proportions of (A) CD3+CD8+ T cells, (B)
CD11c+CD8+ T cells, (C)
CD4+Foxp3+ Treg cells, (D)
CD11b+F480+ macrophages, (E)
CD3+CD4+ T cells, (F)
CD3+DX5+ natural killer cells, (G)
CD11b+Gr1int+ myeloid cells and (H)
B220+ B cells. Data are presented as the mean ± SD
(n=3). *P<0.05 and **P<0.01. NS, not significant; ISTF,
immunostimulatory factor; mAb, monoclonal antibody. |
Combined therapy with ISTF and
anti-4-1BB mAb induces a greater increase in the number of
tumor-reactive effector cells than suppressor cells
As shown in the image in Fig. 3A, the sizes of the spleens and
TDLNs of the mice in the ISTF + 4-1BB group were larger than those
in the other experimental groups. Consistent with this, the number
of splenocytes in the ISTF + 4-1BB group was also significantly
higher than in the other experimental groups. On the other hand,
the number of splenocytes in the 4-1BB-treated mice was higher than
that in the control, but the number of cells in the ISTF-treated
group was similar to that in the control (Fig. 3B). Subsequent detailed analysis of
the immune cell subpopulations revealed that the number of
CD11c+ CD8+ T cells in the ISTF + 4-1BB group
was significantly increased compared with the other groups
(Fig. 3C). The numbers of
CD3+CD4+ T cells (Fig. 3D) and
CD3+CD8+ T cells (Fig. 3E) increased in the ISTF + 4-1BB
group compared with the control and ISTF groups, but were similarly
increased in the 4-1BB group. The numbers of
CD3+DX5+ NK cells (Fig. 3F),
CD11b+F480+ macrophages (Fig. 3G), and B220+ B cells
(Fig. 3H) were similar between the
groups. The numbers of CD11b+Gr1inT cells (Fig. 3I) and
CD4+Foxp3+ cells (Fig. 3J) increased in the ISTF + 4-1BB
group compared with the control and ISTF groups, however, there
were no significant differences compared with the 4-1BB
administration group. The ratio of tumor-reactive immune cells to
suppressor cells has been shown to be more important than the
density or total number of each subtype for antitumor immune
responses in human and animal models (31–33).
Therefore, the present study next investigated the ratios of
CD4+Foxp3+ cells and CD11b+Gr1inT
cells, which are considered regulatory T cells and myeloid-derived
suppressor cells, respectively (31,34,35),
to CD8+ T cells and CD11c+CD8+ T
cells. The CD3+CD8+
cell/CD4+Foxp3+ cell,
CD3+CD8+ cell/CD11b+Gr1inT cell,
CD11c+CD8+
cell/CD4+Foxp3+ cell, and
CD11c+CD8+ cell/CD11b+Gr1inT cell
ratios were increased 2.8-, 2.6-, 13.6- and 11.3-fold,
respectively, in the ISTF + 4-1BB group, compared with the control
group (Fig. 3K-N). This indicated
that combined therapy with ISTF and anti-4-1BB mAb led to a greater
increase in the number of tumor-reactive effector cells than
suppressor cells.
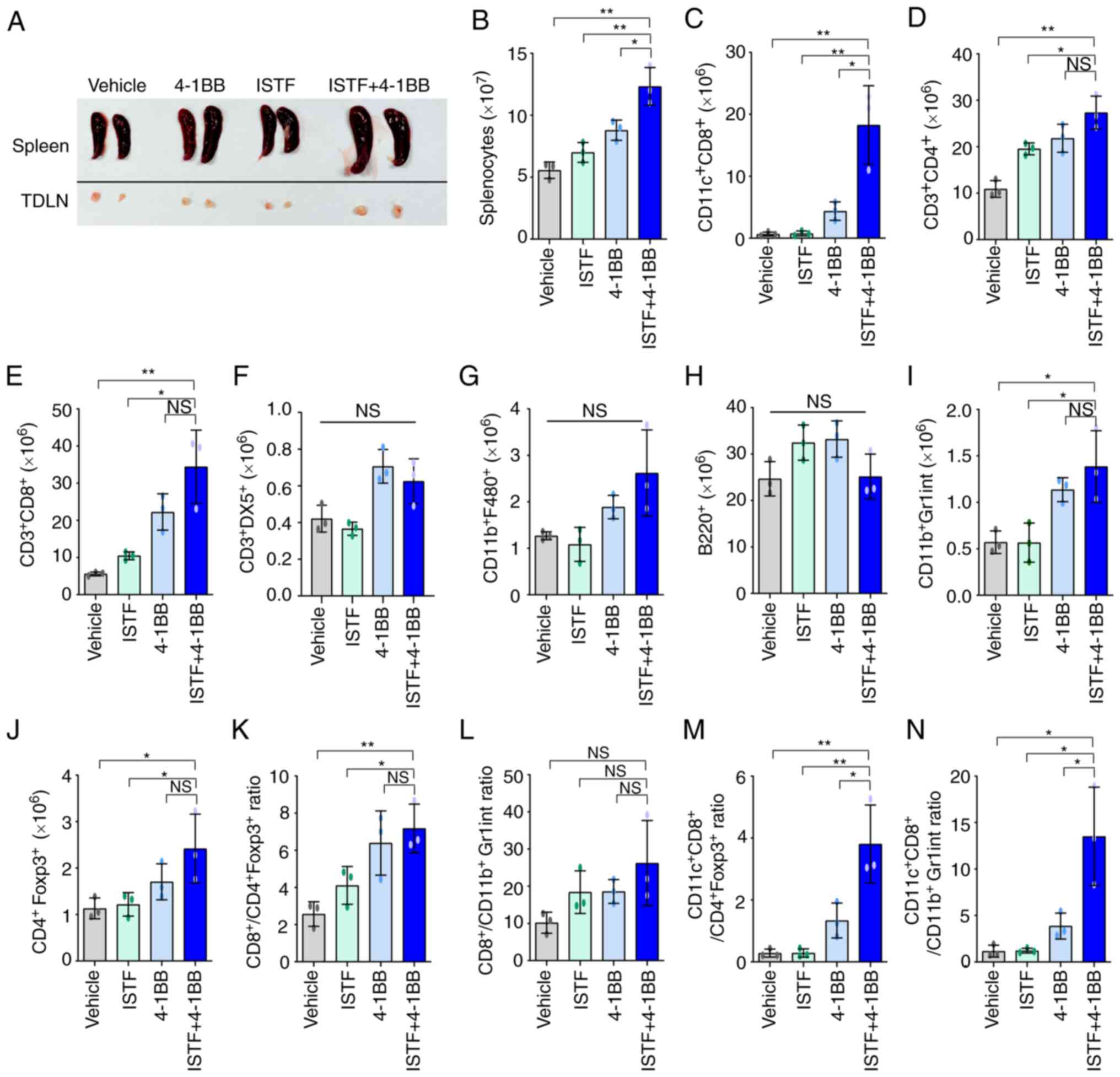 | Figure 3.Combined therapy with ISTF and
anti-4-1BB mAb leads to a greater increase in the number of
tumor-reactive effector cells than suppressor cells. Renca tumors
were established and four groups of mice were treated with ISTF
and/or anti-4-1BB mAb as described in Fig. 1. On day 14, splenocytes were
excised; double stained with PE-, cy and FITC-conjugated antibodies
and analyzed by fluorescence activated cell sorting. (A)
Representative images of spleen and inguinal TDLNs in each group on
day 14. (B) The total numbers of splenocytes were determined on day
14. The numbers of (C) CD11c+CD8+T cells, (D)
CD3+CD4+ T cells, (E)
CD3+CD8+ T cells, (F)
CD3+DX5+ natural killer cells, (G) CD11b+
F480+ cells, (H) B220+ B cells, (I) CD11b+Gr1inT cells
and (J) CD4+Foxp3+ Treg cells. (K) The
CD3+CD8+
cell/CD4+Foxp3+ cell, (L)
CD3+CD8+ cell/CD11b+Gr1inT cell, (M)
CD11c+CD8+
cell/CD4+Foxp3+ cell, and (N)
CD11c+CD8+ cell/CD11b+Gr1inT cell
ratios were analyzed. Data are presented as the mean ± SD (n=3).
*P<0.05 and **P<0.01. NS, not significant; ISTF,
immunostimulatory factor; mAb, monoclonal antibody; TDLN,
tumor-draining lymph node. |
Combined therapy with ISTF and
anti-4-1BB mAb induces marked expansion of
CD11c+CD8+ cells in TILs
We previously reported that
CD11c+CD8+ T cells play a role in antitumor
immunity in melanoma mouse models (36). The administration of anti-4-1BB mAb
to B16F10-melanoma-bearing mice can induce the marked expansion of
CD11c+CD8+ T cells in parallel with the
suppression of pulmonary tumors (37). As CD11c+CD8+
T cells express high levels of CD107a, a marker of activated
cytotoxic T lymphocytes (CTLs), they have been suggested to be
cells with a role in the antitumor immunity induced by anti-4-1BB
mAb (36). In addition,
CD8+ T cells in TILs are considered critical immune
effector cells for antitumor immune responses (3). This prompted the present study to
analyze CD8+ T cells and
CD11c+CD8+ T cells in TILs. TILs from the
tumors of each type of mouse were purified and characterized. The
TILs mainly consisted of CD4+ T cells, CD8+ T
cells, and CD11c+CD8+ T cells. The
percentages of CD3+CD4+ T cells were similar
between the groups (Figs. 4A and
S2). The percentages and numbers
of CD3+CD8+ T cells (Fig. 4B) and
CD11c+CD8+ T cells (Fig. 4C) showed greater increases in the
ISTF + 4-1BB group than in the other experimental groups. The in
the ISTF + 4-1BB group had the highest numbers of TIL cells per mg
of tumor compared with the other groups (Fig. 4D). As the difference in tumor size
between the ISTF + 4-1BB group and control group may have affected
the number and frequency of TIL subtypes in tumors, the number of
TIL subtypes in the same amounts of tumor tissue were compared. The
numbers of CD3+CD4+ T cells,
CD3+CD8+ T cells and
CD11c+CD8+ T cells per mg of tumor tissue
were 2.7-, 11.1-, 32.3-fold higher, respectively, in the ISTF +
4-1BB group than in the control group (Fig. 4E-G).
CD11c+CD8+ T
cells increased by combination therapy with ISTF and anti-4-1BB mAb
have high IFN-g production ability
The next goal was to determine whether the
significant increases in CD8+ T cells and
CD11c+CD8+ T cells induced by anti-4-1BB mAb
and ISTF co-administration resulted in antitumor activity. IFN-γ
has been considered to be a key cytokine in tumor immunity
(38). To further clarify the
mechanisms underlying the ISTF- and 4-1BB-mediated antitumor
immunity in our model, IFN-γ production in CD8+ T cells
and CD11c+CD8+ T cells was investigated by
intracellular cytokine staining. As expected, the IFN-γ-producing
CD8+ T cells and CD11c+CD8+ T
cells were much higher in the ISTF + 4-1BB group than those in the
control mice. IFN-γ-producing CD8+ T cells constituted
31.9% of the total splenocytes of mice in the ISTF + 4-1BB group,
and 5.2 and 18.9% of those in the ISTF or 4-1BB monotherapy groups,
respectively (Fig. 5A). Among
CD8+ T cells, the proportion and number of
CD11c+IFN-γ+ cells were 10.8-fold or 4.1-fold
and 36.4-fold or 6.6-fold higher in the mice in the ISTF + 4-1BB
group than the ISTF or 4-1BB monotherapy groups, respectively
(Fig. 5B). In the mice in the ISTF
+ 4-1BB group, 66.7% of the CD8+CD11c+ cells
secreted IFN-γ (Fig. 5C).
Therefore, a significantly increased number of
CD11c+CD8+ T cells, due to anti-4-1BB mAb and
ISTF co-administration, had high IFN-γ production activity and
represent antitumor effector cells.
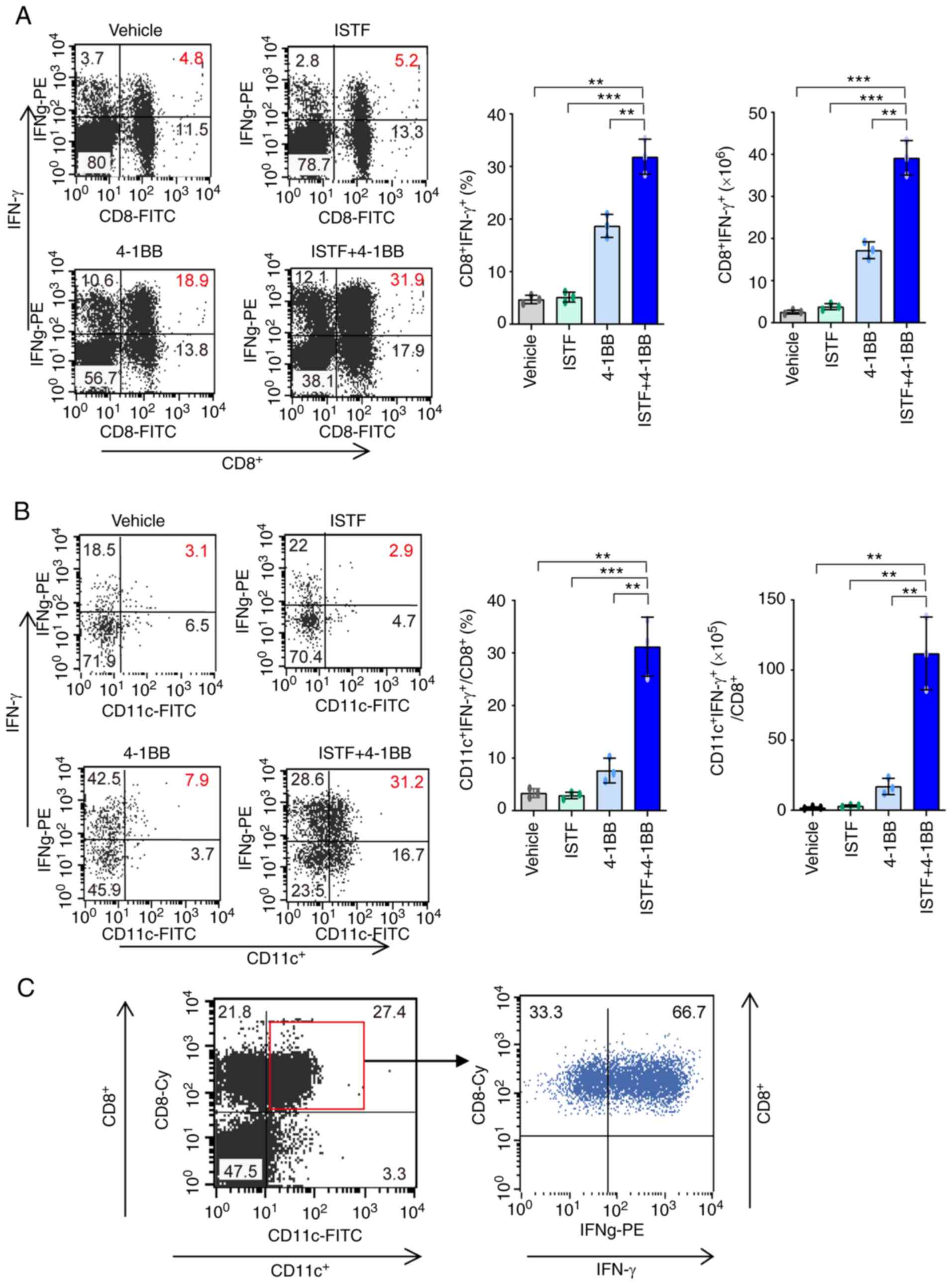 | Figure 5.CD11+CD8+ T
cells increased by combination therapy with ISTF and anti-4-1BB mAb
have high IFN-γ production ability. Renca tumors were established
and four groups of mice were treated with ISTF and/or anti-4-1BB
mAb as described in Fig. 1. To
measure the expression of IFN-γ, spleens were isolated from
Renca-bearing mice on day 14 and cultured with PMA (50 ng/ml) and
ionomycin (500 ng/ml); cytokine release was prevented by treatment
with Golgi-stop. Following surface staining for FITC-CD8 or Cy-CD8
and FITC-CD11c, the cells were fixed, permeabilized and
intracellularly stained with PE-conjugated anti-IFN-γ. (A) Dot
plots show CD8+IFN-γ+ cells (left panel), and
the bar chart indicates % of CD8+IFN-γ+cells
(middle panel) and the number of CD8+IFN-γ+
cells (right panel). (B) Dot plots show
CD11c+IFN-γ+ cells among CD8+
cells (left panel). The bar chart indicates % of
CD11c+IFN-γ+ cells among CD8+ T
cells (middle panel), and the number of
CD11c+IFN-γ+ cells among CD8+ T
cells (right panel). The cells were first gated on CD8+
T cells, and the gated cells were analyzed by FACS for
CD11c+IFN-γ+ cells. (C) Dot plots show
IFN-γ+ cells among CD11c+CD8+ T
cells. The cells were first gated on
CD11c+CD8+ T cells, and the gated cells were
analyzed by FACS for IFN-γ+ cells. Data are presented as
the mean ± SD (n=3). **P<0.01 and ***P<0.001. ISTF,
immunostimulatory factor; mAb, monoclonal antibody; FACS,
fluorescence activated cell sorting. |
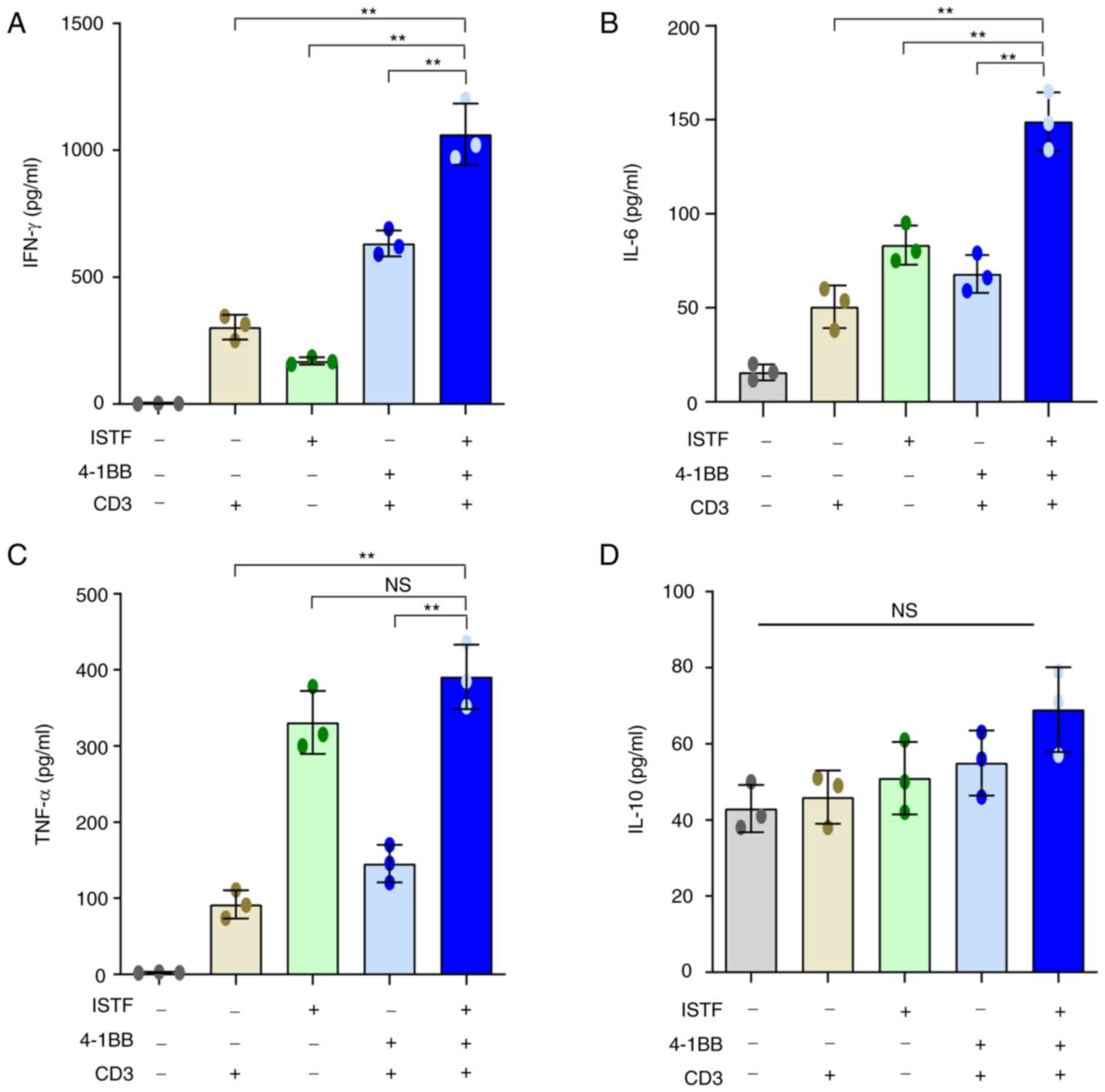
ISTF and anti-4-1BB mAb co-treatment
increases the production of inflammatory cytokines
Studies have shown that the production of
pro-inflammatory cytokines can contribute to cancer immunotherapy,
acting on every phase of the cancer immune cycle, including
improving antigen priming, increasing the number of effector immune
cells in the tumor microenvironment and enhancing their cytolytic
activity (39,40). IFN-γ is known to play an important
role in the anticancer activity of CD8+ T cells
(38,41).
Next, the ability of ISTF and anti-4-1BB mAb
combined therapy to directly induce cytokine production was
analyzed in vitro. Pro-inflammatory cytokines, including
IFN-γ, were measured in the supernatants of splenocytes from naïve
mice stimulated with various combinations of anti-CD3 mAb (0.1
µg/ml), anti-4-1BB mAb (5 µg/ml) and ISTF (10 µg/ml). ISTF,
anti-4-1BB mAb and anti-CD3 mAb co-treatment increased the
production of IFN-γ (Fig. 6A),
IL-6 (Fig. 6B) and TNF-α (Fig. 6C). In particular, IFN-γ showed a
significant synergistic effect compared with the mono-treatment
groups (Fig. 6A). The ISTF
mono-treatment without stimulation with 4-1BB and CD3 also resulted
in increased production of IL-6, IFN-γ and TNF-α compared with the
vehicle-treated group, but the levels of IFN-γ and IL-6 were
significantly lower than that of the co-treatment group (Fig. 6A-C). IL-10 is considered an
immunosuppressive cytokine because it can decrease the
antigen-presenting activity of dendritic cells (DCs) and inhibit
the cytotoxic and cytokine-release functions performed by T and NK
lymphocytes (42,43). The level of IL-10 production in the
ISTF and 4-1BB co-stimulation group was similar to that in the ISTF
and 4-1BB mono-stimulated groups, and there was no significant
difference (Fig. 6D).
Discussion
The present study reported a marked increase in
CD11c+CD8+ T cells in response to anti-4-1BB
mAb and ISTF co-treatment in a tumor model, together with the
suppression of tumor growth. ISTF is a component of the bacterial
outer membrane, a proteinaceous material distinct from LPS, that
can exist in a soluble form and is released by growing and/or lysed
bacteria (26). Influenced by the
fact that ISTF stimulates APC cells such as macrophages and DCs,
the present study investigated whether ISTF could increase the
immunotherapeutic efficiency in the treatment of tumors.
Although cytokine therapy with IL-2 and IFN-γ has
been used for advanced RCC, it has only proved efficacious in a
limited proportion of patients (16,44).
RCC is an immunogenic tumor, based on its response to immunotherapy
and the increase in tumor T cell infiltration (45). Agonistic mAbs targeting 4-1BB have
been developed to harness 4-1BB signaling for cancer immunotherapy
(46). 4-1BB, an inducible
costimulatory receptor, is transiently expressed after T cell
receptor engagement on T cells (3). In the tumor micro-environment (TME),
4-1BB activation can enhance the activity of tumor-specific CTLs
(47,48). In the present study, anti-4-1BB mAb
monotherapy caused slower tumor growth and longer survival compared
with the control group. Further studies should be performed to
address the mechanism underlying this effect, but we suggest that
it was due to the increase of myeloid-derived suppressor cells,
regulatory T cells and TGF-α in TME (49,50).
Previously, anti-4-1BB mAb, urelumab (IgG4), showed hepatotoxicity
at high doses (≥1 mg/kg), but was demonstrated to be safe at 0.1
mg/kg every 3 weeks (51). In
addition, utomilumab (IgG2) has been found to activate 4-1BB
through Fc-mediated crosslinking and shows weaker agonist activity
than urelumab (52,53). The modest antitumor activity of
utomilumab is better tolerated by patients as monotherapy (51), and no synergic effects with PD-1
blockade in combination therapy (54). Thus, to reduce adverse effects and
improve the antitumor activity of 4-1BB, it has been identified
that bispecific antibody (MCLA-145) can activate 4-1BB without
crosslinking via engagement of PD-L1 to enhance tumor-specific T
cell response (55). Therefore, it
was hypothesized in the current study that a combination of ISTF,
promoting antigen presentation and anti-4-1BB mAb, stimulating T
cell co-stimulation signals, could increase T cell activity to
eradicate RCC. It was found that a combined treatment of mice with
ISTF and agonistic 4-1BB antibody increased the antitumor effects
and CD11c+CD8+ T cells than just ISTF or
anti-4-1BB mAb alone. Additionally, most of the
CD11c+CD8+ T cells were effector cells that
produced IFN-γ, which is very important for anticancer activity. It
was also confirmed in vitro that ISTF and 4-1BB stimulation
increased IFN-γ production in T cells. In addition, ISTF,
anti-4-1BB mAb and anti-CD3 mAb co-treatment showed increased
synergistic effects on IFN-γ compared with the effects in the
mono-treatment groups.
Consistent with our previous report (36), Takeda et al (56) and Choi et al (57) also demonstrated that
CD11c+CD8+ T cells are effectors of
anti-4-1BB-mediated tumor suppression through the induction of
Ag-specific CD8+ T cells and an increase in
IFN-γ-producing ability, which represents an active phenotype of
the effector CTLs. The increase in CD11c expression in
CD8+ T cells can be induced by various immunological
stimuli, such as microbial infections or Ag and agonistic
anti-4-1BB mAb (58). The level of
CD11c expression in CD8+ T cells can be a useful marker
for the evaluation of the degree of expansion and the quality of
tumor-specific CTLs as well as a marker for predicting the efficacy
of antitumor immunotherapies (56). As such,
CD11c+CD8+ T cells have been considered
effector cells that play a role in the antitumor immunity induced
upon stimulation with Ag and anti-4-1BB mAbs. The ultimate goal of
cancer immunotherapies is to establish large numbers of effector T
cells that have potent antitumor activity. Although ISTF alone did
not show anticancer activity in the present study, it showed
anticancer activity when co-administered with anti-4-1BB mAb. Thus,
it was suggested that it may be used as an adjuvant agent for
anticancer immunotherapy.
These results indicated that a combination of ISTF
and anti-4-1BB mAb eradicated established tumors by the marked
expansion of CD11c+CD8+ T cells with
anticancer activity in tumor-bearing mice; therefore, it could be a
useful strategy by which to target both antigen presentation and T
cell co-stimulatory signals in incurable RCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was supported by the Basic Science Research
Program through the National Research Foundation of Korea (NRF),
funded by the Ministry of Education (grant nos.
NRF-2017R1D1A1B03036287, NRF-2017R1D1A1B03032831,
NRF-2014R1A6A1030318 and NRF-2015R1D1A1A01059994).
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
SAJ, HTC, WGA and BSK were responsible for the
conceptualization of the present study. SAJ, SMP and YJ acquired,
analyzed and interpreted the data. SAJ, SMP and YJ confirm the
authenticity of all the raw data. SAJ, HTC, WGA and BSK provided
the resources. SAJ, YJ, HTC, WGA and BSK wrote, reviewed and edited
the manuscript, and gave the final approval of the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Animal studies were approved by the University of
Ulsan Animal Care and Use Committee (approval no. HTC-14-030;
Nam-gu, Republic of Korea).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vitale MG and Carteni G: Recent
developments in second and third line therapy of metastatic renal
cell carcinoma. Expert Rev Anticancer Ther. 16:469–471. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Makhov P, Joshi S, Ghatalia P, Kutikov A,
Uzzo RG and Kolenko VM: Resistance to systemic therapies in clear
cell renal cell carcinoma: Mechanisms and management strategies.
Mol Cancer Ther. 17:1355–1364. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartkowiak T and Curran MA: 4-1BB
Agonists: Multi-potent potentiators of tumor immunity. Front Oncol.
5:1172015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yagoda A, Abi-Rached B and Petrylak D:
Chemotherapy for advanced renal-cell carcinoma: 1983–1993. Semin
Oncol. 22:42–60. 1995.PubMed/NCBI
|
|
5
|
Reeves DJ and Liu CY: Treatment of
metastatic renal cell carcinoma. Cancer Chemother Pharmacol.
64:11–25. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hah YS and Koo KC: Immunology and
immunotherapeutic approaches for advanced renal cell carcinoma: A
comprehensive review. Int J Mol Sci. 22:44522021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brown LC, Desai K, Zhang T and Ornstein
MC: The immunotherapy landscape in renal cell carcinoma. BioDrugs.
34:733–748. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Considine B and Hurwitz ME: Current status
and future directions of immunotherapy in renal cell carcinoma.
Curr Oncol Rep. 21:342019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Negrier S, Escudier B, Lasset C, Douillard
JY, Savary J, Chevreau C, Ravaud A, Mercatello A, Peny J, Mousseau
M, et al: Recombinant human interleukin-2, recombinant human
interferon alfa-2a, or both in metastatic renal-cell carcinoma.
Groupe Francais d'Immunotherapie. N Engl J Med. 338:1272–1278.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sica G and Chen L: Modulation of the
immune response through 4-1BB. Adv Exp Med Biol. 465:355–362. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vinay DS and Kwon BS: Role of 4-1BB in
immune responses. Semin Immunol. 10:481–489. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Melero I, Shuford WW, Newby SA, Aruffo A,
Ledbetter JA, Hellström KE, Mittler RS and Chen L: Monoclonal
antibodies against the 4-1BB T-cell activation molecule eradicate
established tumors. Nat Med. 3:682–685. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Miller RE, Jones J, Le T, Whitmore J,
Boiani N, Gliniak B and Lynch DH: 4-1BB-specific monoclonal
antibody promotes the generation of tumor-specific immune responses
by direct activation of CD8 T cells in a CD40-dependent manner. J
Immunol. 169:1792–1800. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Taraban VY, Rowley TF, O'Brien L, Chan HT,
Haswell LE, Green MH, Tutt AL, Glennie MJ and Al-Shamkhani A:
Expression and costimulatory effects of the TNF receptor
superfamily members CD134 (OX40) and CD137 (4–1BB), and their role
in the generation of anti-tumor immune responses. Eur J Immunol.
32:3617–3627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wilcox RA, Flies DB, Zhu G, Johnson AJ,
Tamada K, Chapoval AI, Strome SE, Pease LR and Chen L: Provision of
antigen and CD137 signaling breaks immunological ignorance,
promoting regression of poorly immunogenic tumors. J Clin Invest.
109:651–659. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ju SA, Cheon SH, Park SM, Tam NQ, Kim YM,
An WG and Kim BS: Eradication of established renal cell carcinoma
by a combination of 5-fluorouracil and anti-4-1BB monoclonal
antibody in mice. Int J Cancer. 122:2784–2790. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wallin JJ, Bendell JC, Funke R, Sznol M,
Korski K, Jones S, Hernandez G, Mier J, He X, Hodi FS, et al:
Atezolizumab in combination with bevacizumab enhances
antigen-specific T-cell migration in metastatic renal cell
carcinoma. Nat Commun. 7:126242016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McDermott DF, Sosman JA, Sznol M, Massard
C, Gordon MS, Hamid O, Powderly JD, Infante JR, Fassò M, Wang YV,
et al: Atezolizumab, an Anti-programmed death-ligand 1 antibody, in
metastatic renal cell carcinoma: Long-term safety, clinical
activity, and immune correlates from a phase ia study. J Clin
Oncol. 34:833–842. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zizzari IG, Napoletano C, Di Filippo A,
Botticelli A, Gelibter A, Calabrò F, Rossi E, Schinzari G, Urbano
F, Pomati G, et al: Exploratory pilot study of circulating
biomarkers in metastatic renal cell carcinoma. Cancers (Basel).
12:26202020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Wang Z, Jiang W, Zeng H, Liu Z, Lin
Z, Qu Y, Xiong Y, Wang J, Chang Y, et al: Tumor-infiltrating
TNFRSF9 + CD8 + T cells define different subsets of clear cell
renal cell carcinoma with prognosis and immunotherapeutic response.
Oncoimmunology. 9:18381412020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bromley GS and Solender M: Hand infection
caused by Actinobacillus actinomycetemcomitans. J Hand Surg
Am. 11:434–436. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hofstad T and Stallemo A: Subacute
bacterial endocarditis due to Actinobacillus
actinomycetemcomitans. Scand J Infect Dis. 13:78–79. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Patel PK and Seitchik MW:
Actinobacillus actinomycetemcomitans: A new cause for
granuloma of the parotid gland and buccal space. Plast Reconstr
Surg. 77:476–478. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Salman RA, Bonk SJ, Salman DG and Glickman
RS: Submandibular space abscess due to Actinobacillus
actinomycetemcomitans. J Oral Maxillofac Surg. 44:1002–1005.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Weir DM and Blackwell CC: Interaction of
bacteria with the immune system. J Clin Lab Immunol. 10:1–12.
1983.PubMed/NCBI
|
|
26
|
Jeong SJ, Yee ST, Jo WS, Yu SH, Lee SH,
Lim YJ, Yoo YH, Kim JM, Lee JD and Jeong MH: A novel factor
isolated from Actinobacillus actinomycetemcomitans
stimulates mouse B cells and human peripheral blood mononuclear
cells. Infect Immun. 68:5132–5138. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jo WS, Yee ST, Yoon S, Nam BH, Do E, Jung
BS, Jeong SJ, Hong SH, Yoo YH, Kang CD, et al: Immunostimulating
factor isolated from Actinobacillus actinomycetemcomitans
stimulates monocytes and inflammatory macrophages. Microbiol
Immunol. 50:535–542. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheuk AT, Mufti GJ and Guinn BA: Role of
4-1BB:4-1BB ligand in cancer immunotherapy. Cancer Gene Ther.
11:215–226. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schwartz RH: T cell anergy. Annu Rev
Immunol. 21:305–334. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ju SA, Lee SC, Kwon TH, Heo SK, Park SM,
Paek HN, Suh JH, Cho HR, Kwon B, Kwon BS and Kim BS: Immunity to
melanoma mediated by 4-1BB is associated with enhanced activity of
tumour-infiltrating lymphocytes. Immunol Cell Biol. 83:344–351.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shen Z, Zhou S, Wang Y, Li RL, Zhong C,
Liang C and Sun Y: Higher intratumoral infiltrated Foxp3+ Treg
numbers and Foxp3+/CD8+ ratio are associated with adverse prognosis
in resectable gastric cancer. J Cancer Res Clin Oncol.
136:1585–1595. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sato E, Olson SH, Ahn J, Bundy B,
Nishikawa H, Qian F, Jungbluth AA, Frosina D, Gnjatic S, Ambrosone
C, et al: Intraepithelial CD8+ tumor-infiltrating lymphocytes and a
high CD8+/regulatory T cell ratio are associated with favorable
prognosis in ovarian cancer. Proc Natl Acad Sci USA.
102:18538–18543. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ju SA, Park SM, Lee YS, Bae JH, Yu R, An
WG, Suh JH and Kim BS: Administration of 6-gingerol greatly
enhances the number of tumor-infiltrating lymphocytes in murine
tumors. Int J Cancer. 130:2618–2628. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sarkar R, Mathew A and Sehrawat S:
Myeloid-derived suppressor cells confer infectious tolerance to
dampen virus-induced tissue immunoinflammation. J Immunol.
203:1325–1337. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Obregon-Henao A, Henao-Tamayo M, Orme IM
and Ordway DJ: Gr1(int)CD11b+ myeloid-derived suppressor cells in
Mycobacterium tuberculosis infection. PLoS One. 8:e806692013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ju SA, Park SM, Lee SC, Kwon BS and Kim
BS: Marked expansion of CD11c+CD8+ T-cells in melanoma-bearing mice
induced by anti-4-1BB monoclonal antibody. Mol Cells. 24:132–138.
2007.PubMed/NCBI
|
|
37
|
Kim SH, Singh R, Han C, Cho E, Kim YI, Lee
DG, Kim YH, Kim SS, Shin DH, You HJ, et al: Chronic activation of
4-1BB signaling induces granuloma development in tumor-draining
lymph nodes that is detrimental to subsequent CD8+ T
cell responses. Cell Mol Immunol. 18:1956–1968. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Castro F, Cardoso AP, Goncalves RM, Serre
K and Oliveira MJ: Interferon-gamma at the crossroads of tumor
immune surveillance or evasion. Front Immunol. 9:8472018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen DS and Mellman I: Oncology meets
immunology: The cancer-immunity cycle. Immunity. 39:1–10. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen DS and Mellman I: Elements of cancer
immunity and the cancer-immune set point. Nature. 541:321–330.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bhat P, Leggatt G, Waterhouse N and Frazer
IH: Interferon-gamma derived from cytotoxic lymphocytes directly
enhances their motility and cytotoxicity. Cell Death Dis.
8:e28362017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Llopiz D, Ruiz M, Infante S, Villanueva L,
Silva L, Hervas-Stubbs S, Alignani D, Guruceaga E, Lasarte JJ and
Sarobe P: IL-10 expression defines an immunosuppressive dendritic
cell population induced by antitumor therapeutic vaccination.
Oncotarget. 8:2659–2671. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Moore KW, de Waal Malefyt R, Coffman RL
and O'Garra A: Interleukin-10 and the interleukin-10 receptor. Annu
Rev Immunol. 19:683–765. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Motzer RJ, Bander NH and Nanus DM:
Renal-cell carcinoma. N Engl J Med. 335:865–875. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Vuong L, Kotecha RR, Voss MH and Hakimi
AA: Tumor microenvironment dynamics in clear-cell renal cell
carcinoma. Cancer Discov. 9:1349–1357. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chester C, Sanmamed MF, Wang J and Melero
I: Immunotherapy targeting 4-1BB: Mechanistic rationale, clinical
results, and future strategies. Blood. 131:49–57. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gros A, Robbins PF, Yao X, Li YF, Turcotte
S, Tran E, Wunderlich JR, Mixon A, Farid S and Dudley ME: PD-1
identifies the patient-specific CD8+ tumor-reactive
repertoire infiltrating human tumors. J Clin Invest. 124:2246–2259.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ye Q, Song DG, Poussin M, Yamamoto T, Best
A, Li C, Coukos G and Powell DJ Jr: CD137 accurately identifies and
enriches for naturally occurring tumor-reactive T cells in tumor.
Clin Cancer Res. 20:44–55. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ochoa AC, Zea AH, Hernandez C and
Rodriguez PC: Arginase, prostaglandins, and myeloid-derived
suppressor cells in renal cell carcinoma. Clin Cancer Res. 13
(Suppl):721s–726s. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yu JW, Bhattacharya S, Yanamandra N,
Kilian D, Shi H, Yadavilli S, Katlinskaya Y, Kaczynski H, Conner M,
Benson W, et al: Tumor-immune profiling of murine syngeneic tumor
models as a framework to guide mechanistic studies and predict
therapy response in distinct tumor microenvironments. PLoS One.
13:e02062232018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Segal NH, Logan TF, Hodi FS, McDermott D,
Melero I, Hamid O, Schmidt H, Robert C, Chiarion-Sileni V, Ascierto
PA, et al: Results from an integrated safety analysis of urelumab,
an agonist Anti-CD137 monoclonal antibody. Clin Cancer Res.
23:1929–1936. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li Y, Tan S, Zhang C, Chai Y, He M, Zhang
CW, Wang Q, Tong Z, Liu K, Lei Y, et al: Limited cross-linking of
4-1BB by 4-1BB ligand and the agonist monoclonal antibody
utomilumab. Cell Rep. 25:909–920.e4. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chin SM, Kimberlin CR, Roe-Zurz Z, Zhang
P, Xu A, Liao-Chan S, Sen D, Nager AR, Oakdale NS, Brown C, et al:
Structure of the 4-1BB/4-1BBL complex and distinct binding and
functional properties of utomilumab and urelumab. Nat Commun.
9:46792018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tolcher AW, Sznol M, Hu-Lieskovan S,
Papadopoulos KP, Patnaik A, Rasco DW, Di Gravio D, Huang B,
Gambhire D, Chen Y, et al: Phase Ib study of utomilumab
(PF-05082566), a 4-1BB/CD137 agonist, in combination with
pembrolizumab (MK-3475) in patients with advanced solid tumors.
Clin Cancer Res. 23:5349–5357. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Geuijen C, Tacken P, Wang LC, Klooster R,
van Loo PF, Zhou J, Mondal A, Liu YB, Kramer A, Condamine T, et al:
A human CD137×PD-L1 bispecific antibody promotes anti-tumor
immunity via context-dependent T cell costimulation and checkpoint
blockade. Nat Commun. 12:44452021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Takeda Y, Azuma M, Matsumoto M and Seya T:
Tumoricidal efficacy coincides with CD11c up-regulation in
antigen-specific CD8(+) T cells during vaccine immunotherapy. J Exp
Clin Cancer Res. 35:1432016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Choi BK, Kim YH, Kang WJ, Lee SK, Kim KH,
Shin SM, Yokoyama WM, Kim TY and Kwon BS: Mechanisms involved in
synergistic anticancer immunity of anti-4-1BB and anti-CD4 therapy.
Cancer Res. 67:8891–8899. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Vinay DS and Kwon BS: CD11c+CD8+ T cells:
Two-faced adaptive immune regulators. Cell Immunol. 264:18–22.
2010. View Article : Google Scholar : PubMed/NCBI
|