Introduction
Oral squamous cell carcinoma (OSCC) is one of the
most common malignancies of the head and neck, accounting for
>90% of all oral cancer cases, and is the sixth most common type
of cancer globally (1). Currently,
the management of patients with OSCC is based on histological
parameters, such as TNM stage and tumor grade (2). If there is no cervical lymph node
metastasis in patients with early-stage oral cancer, surgery or
radiotherapy alone has been observed to exert a beneficial effect,
whereas surgery combined with radiotherapy is a more suitable
treatment for intermediate- or advanced-stage oral cancer (3). Long-term addiction to alcohol and
tobacco, poor oral hygiene, long-term irritation by foreign bodies
and malnutrition, particularly insufficient vitamin A levels, have
been indicated to cause OSCC (4).
The complex occurrence and development of oral cancer is regulated
at the genetic level, and protein expression is regulated at the
transcriptional and translational levels (5,6). The
analysis and use of valuable biomarkers that are primarily
associated with the pathogenesis of OSCC can provide a more
comprehensive and detailed understanding of the disease, which may
lead to the development of targeted, personalized, practical and
effective oral cancer treatment options (7). Therefore, to improve the survival
rate and quality of life of patients with OSCC, effective
tumor-related factors, novel therapeutic methods and antitumor
drugs must be urgently identified.
Protein kinase, membrane-associated
tyrosine/threonine 1 (PKMYT1) is located at 16p13.3 on human
chromosome 16, and encodes an important protein belonging to the
WEE kinase family (8,9). PKMYT1 is responsible for encoding a
member of the serine/threonine protein kinase family, and the
encoded protein negatively regulates the G2/M transition
of the cell cycle via phosphorylation and inactivation of
cyclin-dependent kinase 1 (10).
An increasing number of studies have indicated that PKMYT1
overexpression promotes cell proliferation, migration, invasion,
colony-forming ability and epithelial-mesenchymal transition (EMT)
in multiple tumor types (11). For
example, PKMYT1 has previously been revealed to be upregulated in
breast cancer, and was also indicated to regulate cell
proliferation by maintaining the cell cycle and genomic stability
(12). It has also been revealed
that PKMYT1 may promote cell proliferation, invasion and migration
in ovarian cancer by targeting sirtuin 3 (13). Additionally, PKMYT1 has been
indicated to promote cell proliferation and apoptosis resistance in
gastric cancer cells by activating the MAPK signaling pathway
(8). Although PKMYT1 has been
studied in various cancer types, the role of PKMYT1 in OSCC remains
to be investigated.
The hypothesis that PKMYT1 also serves a role in
oral cancer has been previously proposed. According to the Gene
Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/), PKMYT1 is highly
expressed in OSCC tissues (GSE37991). Therefore, the present study
aimed to explore the role of PKMYT1 in OSCC, and its target was
predicted using bioinformatics analysis. The results of the current
study may provide useful information on the mechanisms of cell
proliferation, migration and the EMT process in OSCC, and may
facilitate the identification of a novel target that may be used in
the treatment of this disease.
Materials and methods
Cell culture and transfection
Human oral keratinocytes (HOK-16B), human tongue
squamous cell carcinoma (CAL-27, HSC-4 and SCC-9) cells were
purchased from BeNa Culture Collection (Beijing Beina Chunglian
Institute of Biotechnology) and were cultured in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C in a humidified atmosphere
with 5% CO2.
The pcDNA3.1-cyclin A2 (CCNA2) and empty pcDNA3.1
vectors, short hairpin RNA (shRNA) targeting PKMYT1
(shRNA-PKMYT1-1, target sequence: 5′-CTATGCGGTAAAGCGTTCCAT−3′ and
shRNA-PKMYT1-2, target sequence: 5′-GCTGCGTTCTGTCCTTGTCAT−3′) and a
nonspecific sequence used as a negative control (NC; target
sequence: 5′-CAACAAGATGAAGAGCACCAA−3′) were purchased from Shanghai
GenePharma Co., Ltd. SCC-9 cells (5×105 cells/well) were
seeded into 6-well plates and transfected with 10 nM pcDNA plasmid
or shRNA using Lipofectamine® 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Following 6 h of transfection, the transfection reagent in the
wells were replaced with fresh DMEM supplemented with 10% FBS and
cells were cultured for 2 days (all from Gibco; Thermo Fisher
Scientific, Inc.).
Cell Counting Kit-8 (CCK-8) assay
SCC-9 cells (1×103 cells/well) were
seeded into 96-well plates and incubated for 24 h. Following 24, 48
and 72 h of incubation, 10 µl CCK-8 solution (Sigma-Aldrich; Merck
KGaA) was added to each well, and the cells were incubated at 37°C
for an additional 1 h. Finally, the optical density was determined
at 450 nm using a microplate reader (Thermo Fisher Scientific,
Inc.).
Colony formation assay
SCC-9 cells (3×103 cells/well) were
seeded into 6-well plates and incubated for 14 days at 37°C. The
colonies were then fixed with 4% methanol at room temperature for
15 min, and next stained using 0.1% crystal violet at room
temperature for 15 min. Visible colonies of >50 cells were
observed using an inverted microscope (magnification, ×100; Olympus
Corporation) and analyzed using ImageJ software (v1.8; National
Institutes of Health).
Wound healing assay
SCC-9 cells (5×105 cells/well) were
inoculated in 6-well plates and incubated until the reaching ~90%
confluence. A 200-µl sterile pipette tip was applied to create a
wound in the cell monolayer. Following washing with PBS, the cells
were cultured in serum-free DMEM (Gibco; Thermo Fisher Scientific,
Inc.) at 37°C for 24 h. Images were captured at 0 and 48 h under an
inverted microscope (magnification, ×100; Olympus Corporation).
Transwell assay
The cell invasion assay was performed using
Transwell chambers (8 µm pore size; Corning, Inc.). SCC-9 cells
(3×104 cells) were suspended in 1 ml serum-free DMEM.
The upper chamber was pre-coated with 50 µl Matrigel
(Sigma-Aldrich; Merck KGaA) with serum-free DMEM (1:8 diluted) to
form a gel at 37°C for 30 min, and 0.1 ml cell suspension was added
to each well of the upper chamber. The lower chamber was filled
with DMEM supplemented with 20% FBS. Following 24 h of incubation
at 37°C, the cells in the lower chamber were collected, fixed using
4% paraformaldehyde at room temperature for 10 min and stained
using 0.5% crystal violet solution at room temperature for 10 min
(Sigma-Aldrich; Merck KGaA). Finally, the stained cells were
counted using an inverted microscope (magnification, ×100; Olympus
Corporation).
Co-immunoprecipitation (co-IP)
assay
Co-IP assay was performed to verify the binding
between PKMYT1 and CCNA2. The isolation and quantification of
proteins from SCC-9 cells was conducted using RIPA lysis buffer
(Beijing Solarbio Science & Technology Co., Ltd.) and BCA kits
(Beyotime Institute of Biotechnology), respectively. For IP, 2 µg
antibodies against PKMYT1 (1:100; cat. no. H00009088-D01; Abnova)
or CCNA2 (1:100; cat. no. 67955; Cell Signaling Technology, Inc.)
were combined with 500 µg proteins and incubated overnight at 4°C.
Subsequently, 40 µl protein G/A agarose beads (Invitrogen; Thermo
Fisher Scientific, Inc.) were added to the solution and incubated
with the cell lysates for 2 h. Subsequently, the precipitated
proteins were re-suspended in 2X SDS-PAGE loading buffer (Beyotime
Institute of Biotechnology), boiled for 5 min and eluted from the
beads. Finally, the protein complexes were determined using western
blot analysis as described below.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was collected from SCC-9 cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.) and
placed on ice for 15 min. PrimeScript™ Reverse Transcription
Reagent Kit (Takara Bio, Inc.) was used to reverse transcribe RNA
into cDNA according to the manufacturer's protocol. Subsequently,
QuantiNova SYBR-Green PCR Kit (Qiagen AB) was employed according to
the manufacturer's protocol for qPCR in conjunction with an ABI
7500 System (Thermo Fisher Scientific, Inc.). The 2−ΔΔCq
method (14) was used for analysis
of the results (14). GAPDH was
used as a control for normalization. The primers used were as
follows: PKMYT1 forward, 5′-CATGGCTCCTACGGAGAGGT-3′ and reverse,
5′-ACATGGAACGCTTTACCGCAT−3′; and GAPDH forward,
5′-GGGTGTGAACCATGAGAAGT-3′ and reverse,
5′-GGCATGGACTGTGGTCATGA-3′.
Western blot analysis
Total proteins from SCC-9 cells were extracted using
RIPA lysis buffer (Beijing Solarbio Science & Technology Co.,
Ltd.) and quantified using a BCA kit (Beyotime Institute of
Biotechnology). After protein denaturation by boiling at 100°C for
8 min, 12% SDS-PAGE was performed. Upon transfer onto PVDF
membranes, the membranes were blocked in 5% fat-free milk for 2 h
at room temperature, washed with TBS twice at room temperature (5
sec per wash), and incubated with primary antibodies (all from
Abcam) at 4°C overnight. The primary antibodies used were as
follows: Anti-PKMYT1 (1:500; cat. no. ab200387), anti-MMP2
(1:1,000; cat. no. ab92536), anti-MMP9 (1:1,000; cat. no.
ab283575), anti-E-cadherin (1:10,000; cat. no. ab40772),
anti-N-cadherin (1:5,000; cat. no. ab76011), anti-Snail (1:1,000;
cat. no. ab216347), anti-zinc finger E-box binding homeobox 1
(ZEB1; 1:500; cat. no. ab203829), anti-CCNA2 (1:2,000; cat. no.
ab181591) and GAPDH (1:10,000; cat. no. ab181602). The next day,
the membranes were washed with TBS-0.05% Tween-20 thrice at room
temperature (5 min per wash), and then incubated with an
IgG-horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (1:10,000; cat. no. ab6721; Abcam) for 1 h at room
temperature. Finally, protein bands were detected using an ECL kit
(Beyotime Institute of Biotechnology), and the results were
normalized to GAPDH and analyzed using ImageJ software (v1.8;
National Institutes of Health).
Bioinformatics and statistical
analysis
The Search Tool for the Retrieval of Interacting
Genes/Proteins (STRING; http://www.string-db.org/) (15) and GeneMANIA (http://genemania.org/) (16) online databases were used to search
associations between known proteins. All experiments were repeated
at least three times independently, and the results were expressed
as the mean ± standard deviation. Statistical analysis was
performed using SPSS 19.0 software (IBM Corp.). One-way ANOVA
followed by Tukey's post hoc test was used to evaluate the
statistical significance of the results. P<0.05 was considered
to indicate a statistically significant difference.
Results
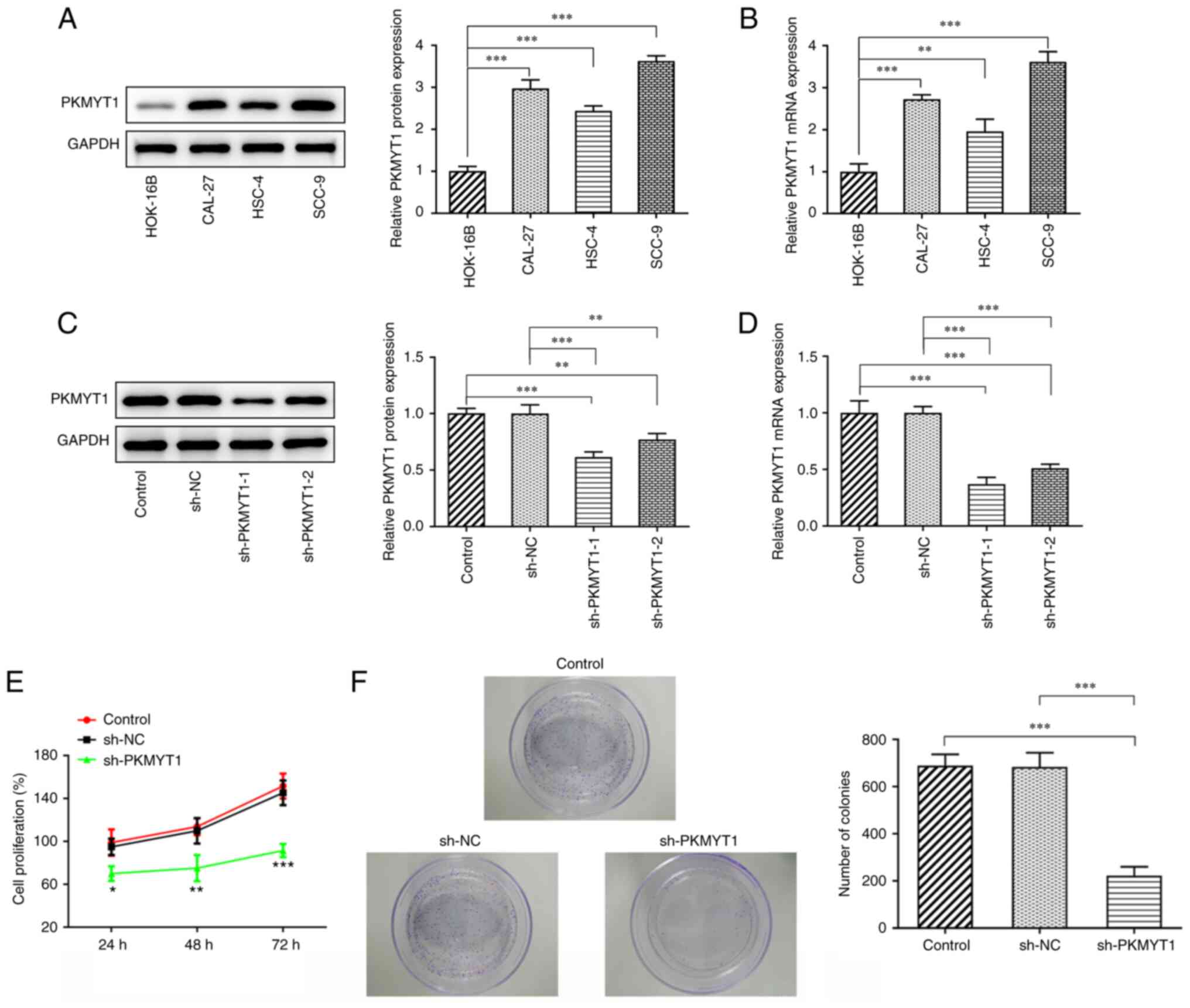
PKMYT1 is upregulated in OSCC cells,
and PKMYT1 knockdown inhibits the proliferation and colony
formation of SCC-9 cells
The relative mRNA and protein expression of PKMYT1
in normal human HOK-16B and OSCC cells, including CAL-27, HSC-4 and
SCC-9 cells, was detected using RT-qPCR and western blot analyses.
The results revealed that the relative mRNA and protein expression
of PKMYT1 in OSCC cells was significantly upregulated compared with
that of the HOK-16B group, particularly in SCC-9 cells (Fig. 1A and B). Therefore, SCC-9 cells
were selected for the subsequent experiments.
To investigate the effects of PKMYT1 on OSCC cells,
SCC-9 cells were transfected with sh-PKMYT1. As presented in
Fig. 1C and D, PKMYT1 mRNA and
protein expression was decreased in the sh-PKMYT1 group compared
with that of the shRNA (NC) group, particularly in the sh-PKMYT1-1
group. Therefore, SCC-9 cells transfected with sh-PKMYT1-1 were
used for subsequent experiments.
Cell proliferation was determined using a CCK-8
assay. As presented in Fig. 1E,
the proliferation of SCC-9 cells was decreased following
transfection with shRNA targeting PKMYT1. It was also revealed that
PKMYT1 knockdown could significantly inhibit SCC-9 cell
proliferation at the time points of 24, 48 and 72 h. Furthermore,
the colony-forming ability of SCC-9 cells was also found to be
suppressed by PKMYT1 knockdown. These results revealed that PKMYT1
knockdown could inhibit the proliferation and colony-forming
ability of SCC-9 cells.
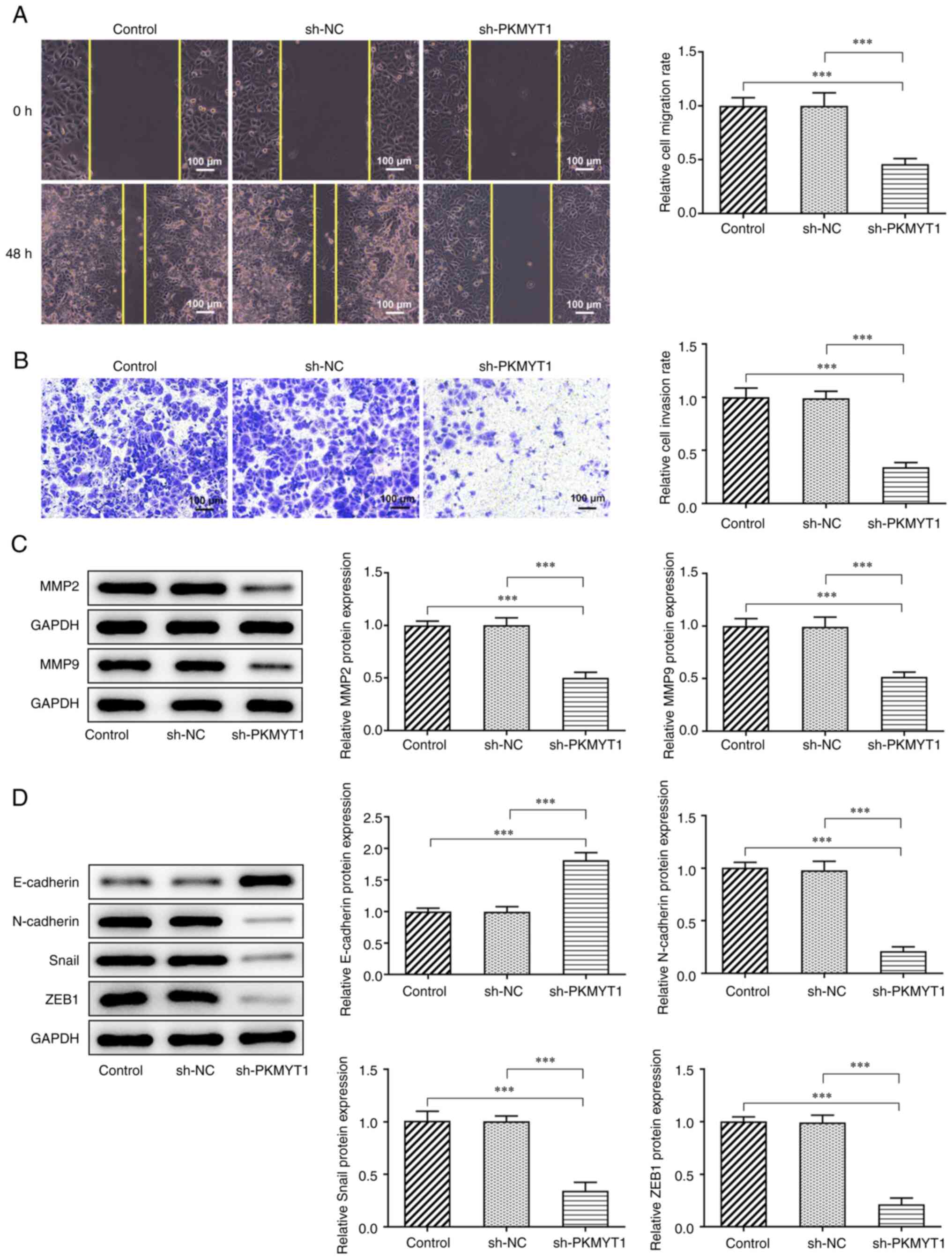
PKMYT1 knockdown inhibits the
migration, invasion and EMT of SCC-9 cells
The relative migration rate and invasion ability
were assessed using wound healing and Transwell assays,
respectively. The results presented in Fig. 2A showed that PKMYT1 knockdown
suppressed the migration rate of SCC-9 cells compared with that of
the shRNA NC group. The results also demonstrated that PKMYT1
knockdown exerted the same inhibitory effects on SCC-9 cell
invasiveness, which was indicated by the decrease in the relative
cell invasion rate (Fig. 2B).
Furthermore, the expression of MMP2 and MMP9, which
was determined using western blot analysis, was markedly decreased
by PKMYT1 knockdown (Fig. 2C).
Additionally, the expression of EMT-related proteins was detected
using western blot analysis. As presented in Fig. 2D, PKMYT1 knockdown significantly
promoted E-cadherin expression and downregulated the expression of
N-cadherin, Snail and ZEB1.
PKMYT1 knockdown inhibits the
expression of CCNA2 in OSCC cells
The STRING and GeneMANIA databases suggested an
association between PKMYT1 and CCNA2 (Fig. 3A). The mRNA and protein expression
of CCNA2 in normal human HOK-16B cells and in OSCC cells, including
CAL-27, HSC-4 and SCC-9 cells, was determined using RT-qPCR and
western blot analyses, respectively. The results presented in
Fig. 3B and C revealed that there
was higher expression of CCNA2 in OSCC cells, particularly in SCC-9
cells, than in HOK-16B cells.
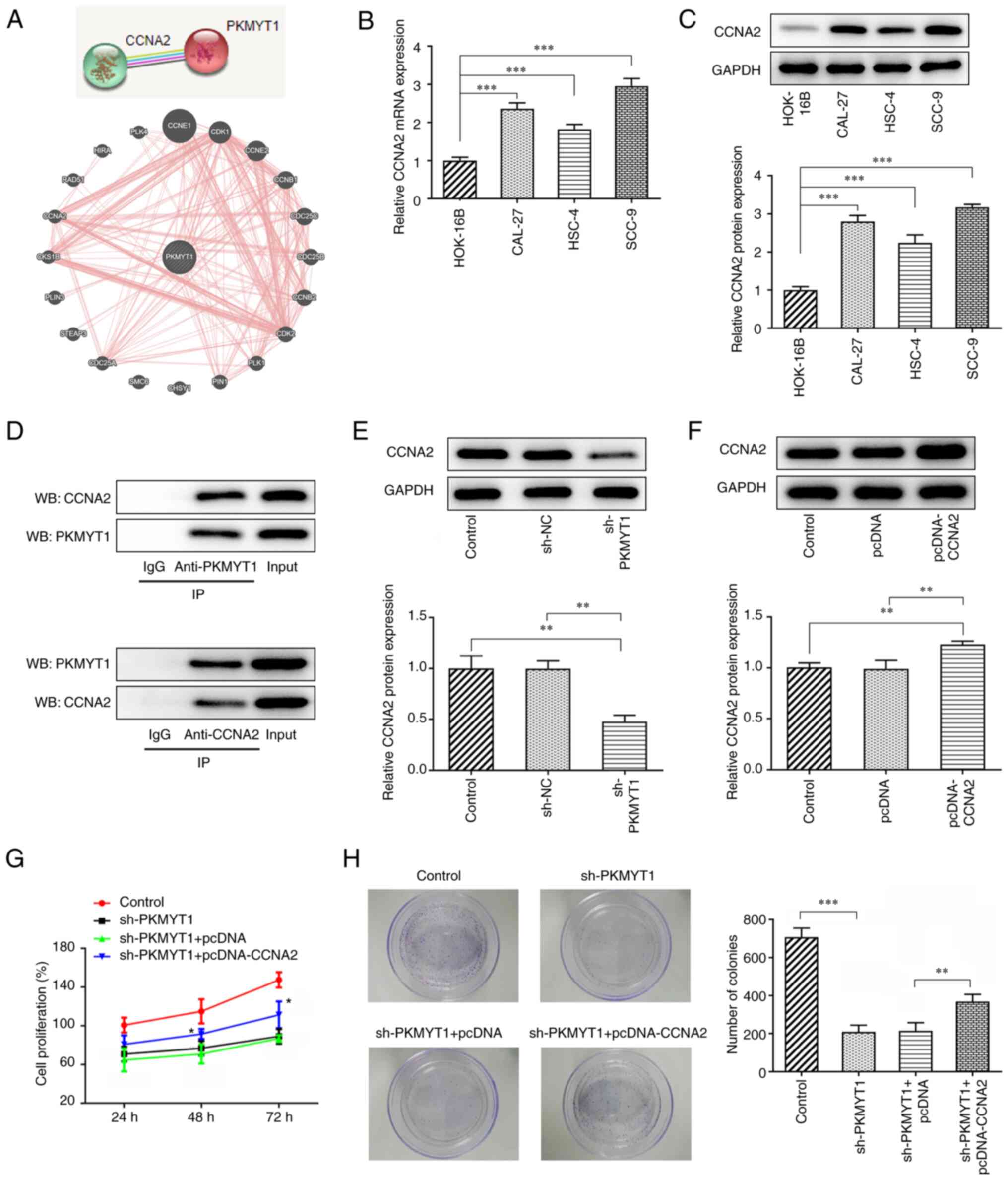 | Figure 3.PKMYT1 knockdown inhibits the
proliferation of SCC-9 cells by regulating CCNA2 expression. (A)
PKMYT1 was found to be able to bind CCNA2 according to the STRING
and GeneMANIA databases. (B and C) Relative mRNA and protein
expression levels were detected using (B) RT-qPCR and (C) western
blot analysis, respectively. (D) Co-IP assay was used for
verification of the binding between PKMYT1 and CCNA2. (E) CCNA2
expression in the sh-PKMYT1 group was detected using western blot
analysis. (F) CCNA2 expression in the pcDNA-CCNA2 group was
detected using western blot analysis. (G) A Cell Counting Kit-8
assay was used for the determination of cell proliferation.
*P<0.05 vs. sh-PKMYT1 + pcDNA. (H) A colony formation assay was
used for the detection of cell colony formation. **P<0.01,
***P<0.001. PKMYT1, protein kinase, membrane-associated
tyrosine/threonine 1; CCNA2, cyclin A2; RT-qPCR, reverse
transcription-quantitative PCR; co-IP, Co-immunoprecipitation; sh,
short hairpin; sh-NC, negative control short hairpin RNA; WB,
western blotting. |
Considering the positive association between PKMYT1
and CCNA2, a co-IP assay was used to further analyze the binding of
PKMYT1 to CCNA2. As presented in Fig.
3D, PKMYT1 was expressed in the anti-CCNA2 group and CCNA2 was
expressed in the anti-PKMYT1 group, while IgG expression was not
detected, revealing that PKMYT1 may combine with CCNA2. Compared
with that of the shRNA NC group, the expression of CCNA2 was
decreased in the sh-PKMYT1 group (Fig.
3E).
PKMYT1 knockdown inhibits the
proliferation and colony formation of OSCC cells by downregulating
CCNA2 expression
SCC-9 cells were transfected with pcDNA-CCNA2, and
the expression of CCNA2 was subsequently determined using western
blot analysis. The results demonstrated that the relative protein
expression of CCNA2 in SCC-9 cells was upregulated in comparison
with that of the pcDNA group (Fig.
3F). As is shown Fig. 3G and
H, the decreased cell proliferation and colony formation caused
by PKMYT1 knockdown was partially reversed by CCNA2 overexpression,
suggesting that CCNA2 overexpression may partly abolish the
inhibitory effects of PKMYT1 knockdown on SCC-9 cell proliferation
and colony formation.
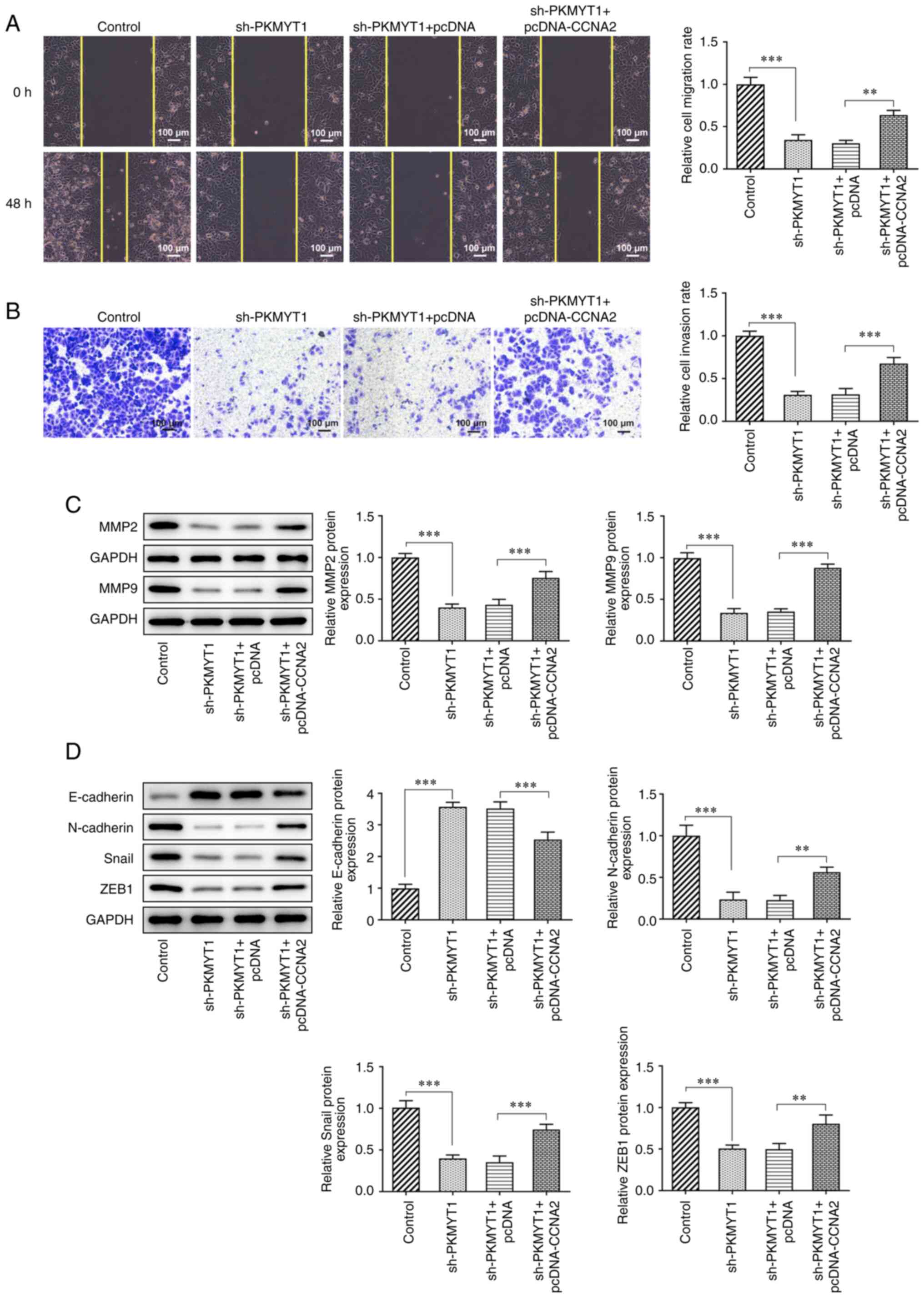
PKMYT1 knockdown inhibits the
migration, invasion and EMT of SCC-9 cells by downregulating CCNA2
expression
Compared with those of the sh-PKMYT1 + pcDNA group,
the decreased migration and invasion rates of SCC-9 cells were
increased following CCNA2 overexpression (Fig. 4A and B).
Furthermore, the expression of MMP2 and MMP9, which
was measured using western blot analysis, was partly increased in
the sh-PKMYT1 + pcDNA-CCNA2 compared with that in the sh-PKMYT1 +
pcDNA group (Fig. 4C).
Additionally, as presented in Fig.
4D, CCNA2 overexpression partly inhibited the expression of
E-cadherin, and increased the expression of N-cadherin, Snail and
ZEB1 in SCC-9 cells compared with the findings in the sh-PKMYT1 +
pcDNA group.
Discussion
OSCC is one of the most aggressive tumors worldwide,
and there is currently no optimal treatment for this disease
(17). PKMYT1 is closely
associated with tumor radiosensitivity, making it a candidate
target for the development of improved treatments for OSCC
(18). Previously, PKMYT1
inhibition has been reported to significantly suppress the
proliferation of prostate cancer cells, thus serving as a novel
therapeutic target for prostate cancer (19). Furthermore, Jeong et al
(20) have suggested that PKMYT1
serves a vital oncogenic role in colorectal cancer, which is
demonstrated by the increased proliferation, migration, invasion
and colony forming ability of colorectal cancer cell lines.
According to the GEO database, PKMYT1 was found to be upregulated
in patients with OSCC (21). The
results of the present study revealed that PKMYT1 was highly
expressed in three OSCC cell lines compared with its expression in
HOK-16B cells. The expression level of PKMYT1 in HSC-4 cells was
lower than that in other OSCC cell lines, which may be due to its
non-metastatic nature (22). In
addition, the expression levels of PKMYT1 in CAL-27 cells were
higher than those in HSC-4 cells in the present study. This may be
due to the relatively high level of keratin in CAL-27 cells, which
is associated with cell proliferation and differentiation (23). The SCC-9 cell line has been used in
the study of metastatic OSCC and displays strong migration and
invasiveness (24). The present
results also demonstrated that the proliferation and colony forming
ability of SCC-9 cells were decreased following the suppression of
PKMYT expression. Furthermore, PKMYT1 knockdown exerted inhibitory
effects on cell invasion, migration and expression of
migration-related proteins, including MMP2 and MMP9, as well as
EMT-related proteins, including N-cadherin, Snail and ZEB1.
CCNA2 is a member of the highly conserved cyclin
family, which participates in the regulation of G1/S and
G2/M phases, and plays a role in DNA replication,
transcription and tumor progression. CCNA2 has been reported to be
highly expressed in a variety of cancer types (25). Yang et al (26) suggested that CCNA2 expression
promotes tumor growth in hepatocarcinoma xenotransplantation mouse
model. Furthermore, CCNA2 has also been previously revealed to
participate in the EMT and metastasis of colorectal cancer
(27). A previous study has
demonstrated the presence of CCNA2, B1, D1 and E1, within the CCND1
gene, in 67 cases of primary OSCC (28). CCNA2 downregulation has also been
revealed to regulate the migration and proliferation of
trophoblasts (29). Additionally,
CCND1 has been found to promote the invasion, migration and EMT of
non-small cell lung carcinoma (NSCLC) cells, and may serve as a
novel effective target for the treatment of NSCLC (30). CCNA2 regulates the EMT process
probably by activating the transcription of EMT-related genes (such
as Snail, Nanog and Myc) through the β-catenin (31) or Rho-associated coiled-coil
containing protein kinase signaling pathways (32). According to the STRING and
GeneMANIA databases, PKMYT1 may be associated with CCNA2. In the
present study, it was demonstrated that PKMYT1 could bind to CCNA2
in SCC-9 cells, and its knockdown may inhibit the expression of
CCNA2. Additionally, the decreased cell proliferation, migration,
invasion and EMT caused by PKMYT1 knockdown were demonstrated to be
reversed by CCNA2 overexpression, revealing that PKMYT1 may inhibit
the malignant progression of OSCC via targeting CCNA2. However, the
mechanism of CCNA2 downregulation remains to be further
investigated to clarify whether it occurs specifically through
regulating transcription, translation or promoting protein
degradation. Furthermore, the present study is only preliminary and
mainly based on one OSCC cell line. In future studies, clinical
samples should be collected, and the clinical significance of
PKMYT1 should be verified further.
In conclusion, the current study demonstrated that
the expression of PKMYT1 and CCNA2 was upregulated in OSCC cell
lines, and there was an association between the two proteins.
Furthermore, it was revealed that PKMYT1 knockdown exerted
inhibitory effects on cell proliferation, migration, invasion and
EMT, while these effects were reversed by CCNA2 overexpression,
revealing that PKMYT1 may serve a role in OSCC by targeting CCNA2.
The findings of the present study may have revealed a novel
biomarker or target that could be used in future treatment options
for patients with OSCC.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YC made substantial contributions to the conception
of the study and performed the experiments. WY was involved in
performing the experiments and writing the manuscript. YC and WY
confirm the authenticity of the raw data. Both authors have read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Panarese I, Aquino G, Ronchi A, Longo F,
Montella M, Cozzolino I, Roccuzzo G, Colella G, Caraglia M and
Franco R: Oral and Oropharyngeal squamous cell carcinoma:
Prognostic and predictive parameters in the etiopathogenetic route.
Expert Rev Anticancer Ther. 19:105–119. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cohen EEW, Bell RB, Bifulco CB, Burtness
B, Gillison ML, Harrington KJ, Le QT, Lee NY, Leidner R, Lewis RL,
et al: The Society for immunotherapy of Cancer consensus statement
on immunotherapy for the treatment of squamous cell carcinoma of
the head and neck (HNSCC). J Immunother Cancer. 7:1842019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Everts HB and Akuailou EN: Retinoids in
cutaneous squamous cell carcinoma. Nutrients. 13:1532021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jia T, Wang F, Qiao B, Ren Y, Xing L,
Zhang H and Li H: Knockdown of LncRNA PANDAR by CRISPR-dCas9
decreases proliferation and increases apoptosis in oral squamous
cell carcinoma. Front Mol Biosci. 8:6537872021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu Y, Zheng Z, Yuan Y, Pathak JL, Yang X,
Wang L, Ye Z, Cho WC, Zeng M and Wu L: The emerging role of
exosomes in oral squamous cell carcinoma. Front Cell Dev Biol.
9:6281032021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Glastonbury CM: Head and Neck Squamous
Cell Cancer: Approach to Staging and Surveillance. In: Diseases of
the Brain, Head and Neck, Spine 2020–2023: Diagnostic Imaging.
Hodler J. (Kubik-Huch RA and von Schulthess GK (eds). Springer,
Cham, CH). 2020. View Article : Google Scholar
|
|
8
|
Cai L, Zhang X, Hou M and Gao F: Natural
flavone tricetin suppresses oxidized LDL-induced endothelial
inflammation mediated by Egr-1. Int Immunopharmacol. 80:1062242020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Q, Zhao X, Zhang C, Wang W, Li F,
Liu D, Wu K, Zhu D, Liu S, Shen C, et al: Overexpressed PKMYT1
promotes tumor progression and associates with poor survival in
esophageal squamous cell carcinoma. Cancer Manag Res. 11:7813–7824.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ghelli Luserna di Rorà A, Cerchione C,
Martinelli G and Simonetti G: A WEE1 family business: Regulation of
mitosis, cancer progression, and therapeutic target. J Hematol
Oncol. 13:1262020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen P, Zhang Z and Chen X: Overexpression
of PKMYT1 facilitates tumor development and is correlated with poor
prognosis in clear cell renal cell carcinoma. Med Sci Monit.
26:e9267552020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Y, Qi J, Dou Z, Hu J, Lu L, Dai H,
Wang H and Yang W: Systematic expression analysis of WEE family
kinases reveals the importance of PKMYT1 in breast carcinogenesis.
Cell Prolif. 53:e127412020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xuan ZH, Wang HP, Zhang XN, Chen ZX, Zhang
HY and Gu MM: PKMYT1 aggravates the progression of ovarian cancer
by targeting SIRT3. Eur Rev Med Pharmacol Sci. 24:5259–5266.
2020.PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
von Mering C, Huynen M, Jaeggi D, Schmidt
S, Bork P and Snel B: STRING: A database of predicted functional
associations between proteins. Nucleic Acids Res. 31:258–261. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Warde-Farley D, Donaldson SL, Comes O,
Zuberi K, Badrawi R, Chao P, Franz M, Grouios C, Kazi F, Lopes CT,
et al: The GeneMANIA prediction server: Biological network
integration for gene prioritization and predicting gene function.
Nucleic Acids Res. 38:W214–W220. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan X and Su H: YM155 Down-Regulates
Survivin and induces P53 Up-regulated modulator of apoptosis
(PUMA)-dependent in oral squamous cell carcinoma cells. Med Sci
Monit. 23:1963–1972. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Long HP, Liu JQ, Yu YY, Qiao Q and Li G:
PKMYT1 as a potential target to improve the radiosensitivity of
lung adenocarcinoma. Front Genet. 11:3762020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang J, Wang L, Chen S, Peng H, Xiao L, E
Du, Liu Y, Lin D, Wang Y, Xu Y and Yang K: PKMYT1 is associated
with prostate cancer malignancy and may serve as a therapeutic
target. Gene. 744:1446082020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jeong D, Kim H, Kim D, Ban S, Oh S, Ji S,
Kang D, Lee H, Ahn TS, Kim HJ, et al: Protein kinase,
membrane-associated tyrosine/threonine 1 is associated with the
progression of colorectal cancer. Oncol Rep. 39:2829–2836.
2018.PubMed/NCBI
|
|
21
|
Yadav M, Pradhan D and Singh RP:
Integrated analysis and identification of nine-gene signature
associated to oral squamous cell carcinoma pathogenesis. 3 Biotech.
11:2152021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sumita Y, Yamazaki M, Maruyama S, Abé T,
Cheng J, Takagi R and Tanuma JI: Cytoplasmic expression of SOX9 as
a poor prognostic factor for oral squamous cell carcinoma. Oncol
Rep. 40:2487–2496. 2018.PubMed/NCBI
|
|
23
|
Jung HM, Phillips BL, Patel RS, Cohen DM,
Jakymiw A, Kong WW, Cheng JQ and Chan EK: Keratinization-associated
miR-7 and miR-21 regulate tumor suppressor reversion-inducing
cysteine-rich protein with kazal motifs (RECK) in oral cancer. J
Biol Chem. 287:29261–29272. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chou SC, Azuma Y, Varia MA and Raleigh JA:
Evidence that involucrin, a marker for differentiation, is oxygen
regulated in human squamous cell carcinomas. Br J Cancer.
90:728–735. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gao T, Han Y, Yu L, Ao S, Li Z and Ji J:
CCNA2 is a prognostic biomarker for ER+ breast cancer and tamoxifen
resistance. PLoS One. 9:e917712014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang F, Gong J, Wang G, Chen P, Yang L and
Wang Z: Waltonitone inhibits proliferation of hepatoma cells and
tumorigenesis via FXR-miR-22-CCNA2 signaling pathway. Oncotarget.
7:75165–75175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bendris N, Arsic N, Lemmers B and
Blanchard JM: Cyclin A2, Rho GTPases and EMT. Small GTPases.
3:225–228. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Monteiro LS, Diniz-Freitas M,
Warnakulasuriya S, Garcia-Caballero T, Forteza-Vila J and Fraga M:
Prognostic significance of Cyclins A2, B1, D1, and E1 and CCND1
numerical aberrations in oral squamous cell carcinomas. Anal Cell
Pathol (Amst). 2018:72535102018.PubMed/NCBI
|
|
29
|
Li X, Ma XL, Tian FJ, Wu F, Zhang J, Zeng
WH, Lin Y and Zhang Y: Downregulation of CCNA2 disturbs trophoblast
migration, proliferation, and apoptosis during the pathogenesis of
recurrent miscarriage. Am J Reprod Immunol. 82:e131442019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ruan JS, Zhou H, Yang L, Wang L, Jiang ZS
and Wang SM: CCNA2 facilitates epithelial-to-mesenchymal transition
via the integrin αvβ3 signaling in NSCLC. Int J Clin Exp Pathol.
10:8324–8333. 2017.PubMed/NCBI
|
|
31
|
Cheung CT, Bendris N, Paul C, Hamieh A,
Anouar Y, Hahne M, Blanchard JM and Lemmers B: Cyclin A2 modulates
EMT via β-catenin and phospholipase C pathways. Carcinogenesis.
36:914–924. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Ying Y, Xie H, Jin K, Yan H, Wang S,
Xu M, Xu X, Wang X, Yang K, et al: Dual regulatory role of CCNA2 in
modulating CDK6 and MET-mediated cell-cycle pathway and EMT
progression is blocked by miR-381-3p in bladder cancer. FASEB.
33:1374–1388. 2019. View Article : Google Scholar : PubMed/NCBI
|