Introduction
Cervical carcinoma (CC) ranks among the top four
most common cancer types in women worldwide (1), and is characterized by high incidence
and fatality rates (2). At
present, the primary treatments for CC are limited to radiotherapy,
chemotherapy and surgery (3–5).
Although they are considered relatively effective, these modalities
inevitably give rise to serious side effects, and are not
sufficient to prevent cancer metastasis (6). With the development of modern
molecular biology and genomics, immunotherapy has become a hot spot
in the treatment of advanced and recurrent CC; this type of
treatment is in a stage of rapid development and is considered to
bring new hope to patients with advanced CC (7). In addition, prophylactic vaccines
against numerous strains of high-risk human papillomavirus (HPV)
have been developed, but are not a completely effective treatment
option for CC (8). The HPV vaccine
does not protect against all types of HPV infection, can only be
administered within a certain age range, and is unavailable in
numerous developing countries (9).
Therefore, it is necessary to investigate other effective means of
preventing and treating CC.
Tetramethylpyrazine (TMP) is a natural compound
derived from Ligusticum wallichii (L. Wallichii),
which is also known as Rhizoma Chuanxiong (Chuanxiong) (10). Chuanxiong is believed to possess
numerous beneficial properties, which are recorded in ancient
Chinese medical works, including Annotation of Materia
Medica and Compendium of Materia Medica (11). Chuanxiong has also been used in
Traditional Chinese Medicine for thousands of years (12). Along with ferulic acid, Chuanxiong
contains TMP and is reported to be able to prevent and attenuate
the progression of numerous diseases, such as cardiovascular
diseases, ischemic stroke and diabetes (13–16).
Furthermore, over the last 10 years, several studies have confirmed
the inhibitory effects and mechanisms of TMP on numerous types of
cancer, including prostate (17),
lung (18) and bladder cancer
(19). For example, a study
reported that TMP treatment reduces the viability and increases the
apoptosis of prostate cancer (PCa) cells in a dose-dependent manner
(20), indicating that TMP may be
a promising therapeutic agent for PCa. TMP has also been
demonstrated to significantly decrease the viability, migration and
invasiveness of breast adenocarcinoma MDA-MB-231 cells, and to
increase their apoptosis in a dose-dependent manner (21). Therefore, we hypothesized that TMP
may also exert an inhibitory effect on CC cells.
Hedgehog (Hh) signaling molecule is a local protein
ligand secreted by signaling cells, and its signaling pathway
controls cell fate, proliferation and differentiation (22). The abnormal activation of the Hh
signaling pathway has been demonstrated to be involved in tumor
occurrence and development (23).
For example, activation of the Hh signaling pathway is associated
with tissue invasion and possible metastasis in gastric cancer
(22). The regulation of the Hh
signaling pathway by TMP has also been investigated in numerous
studies. For example, TMP was demonstrated to effectively inhibit
the Hh signaling pathway in hepatic fibrosis (24), has also been reported to be
activated in cervical carcinoma cell lines (25). However, the role of TMP in CC, and
the associated underlying mechanisms, remain unclear. Therefore,
the aim of the present study was to investigate the role of TMP in
inhibiting the development of CC by blocking the Hh signaling
pathway, which may provide the foundations of novel therapeutics
for the prevention and treatment of CC.
Materials and methods
Cell culture and treatments
The human cervical epithelial Ect1 cell line and the
CC C33A cell line were purchased from the American Type Culture
Collection. Cells were cultured in DMEM supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C in an atmosphere of
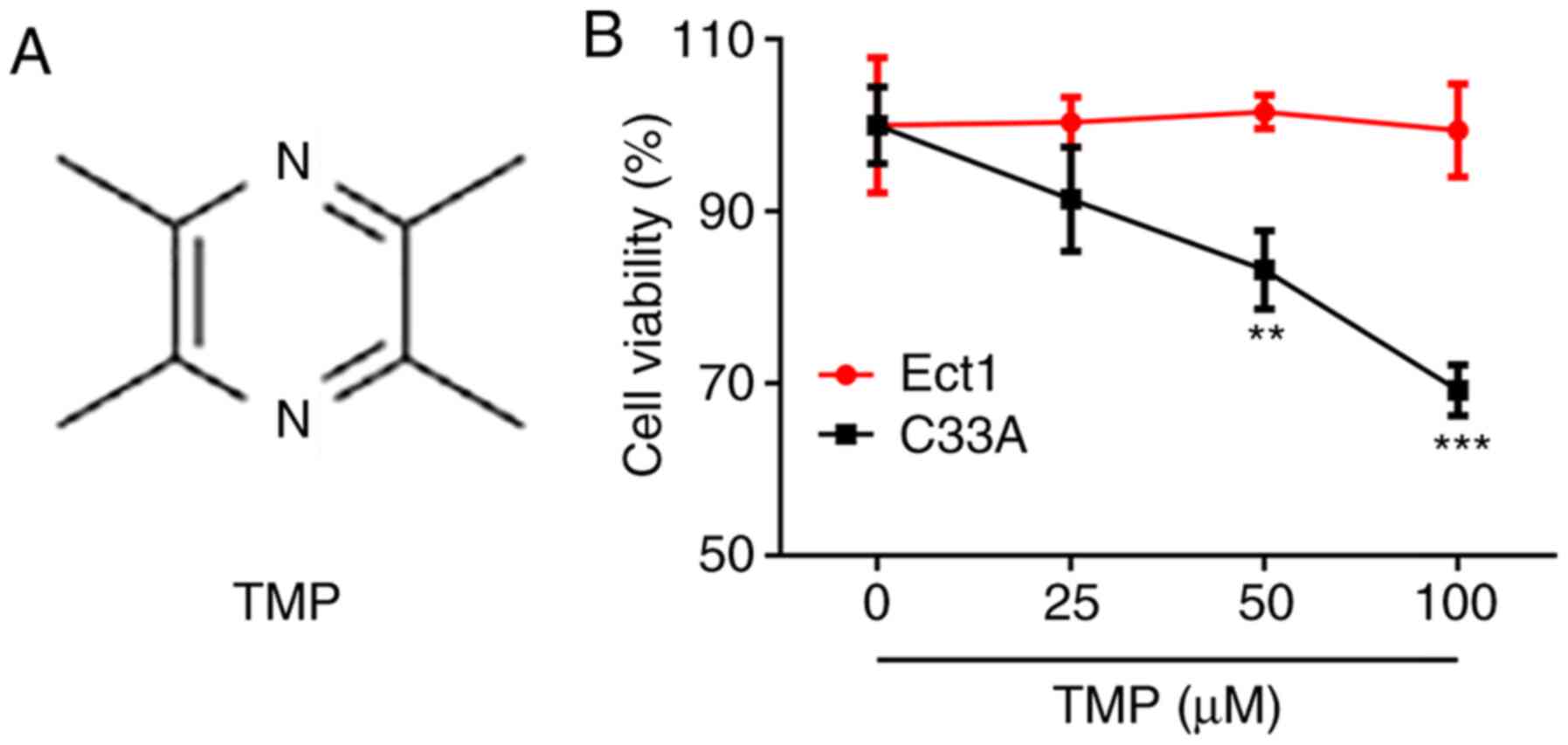
95% air and 5% CO2. TMP (purity ≥99%; Fig. 1A) was purchased from Beijing
Solarbio Science & Technology Co., Ltd., dissolved in DMSO
solution and used to treat C33A cells at doses of 25, 50 and 100 µM
(26). Hh and smoothened agonist
(SAG) was purchased from Sigma-Aldrich (Merck KGaA) and diluted to
100 nM (27).
MTT assay
To determine the effect of TMP on cell viability,
Ect1 and C33A cells were separately collected in the logarithmic
phase and 100 µl cell suspension was added to each well of a
96-well plate. The density of cells was adjusted to
2×103 cells/well. The cells were then incubated in 5%
CO2 at 37°C until a monolayer covered the bottom of the
well. TMP (25, 50 and 100 µM) or TMP (100 µM) + SAG was
subsequently added, and the cells were incubated at 37°C for 24 h.
Then 0.5% MTT solution (Beyotime Institute of Biotechnology) was
added and the cells were incubated for a further 4 h. Then, the
medium was removed and 150 µl DMSO was added to each well. The
plate was oscillated at low speed on a shaker for 10 min to fully
dissolve the formazan crystals, and the absorbance value of each
well was analyzed at 570 nm.
To determine the effect of TMP on cellular
proliferation, C33A cells were treated with TMP (25, 50 and 100 µM)
or TMP (100 µM) + SAG, followed by culture for 24, 48 and 72 h.
Samples were then treated with MTT as aforementioned.
Colony formation assay
C33A cells in the logarithmic growth phase were
digested with 0.25% trypsin. The cell density was adjusted to
1×105 cells/ml in DMEM containing 20% FBS and TMP (25,
50 and 100 µM) or TMP (100 µM) + SAG. Low melting point agar
solutions of 1.2 and 0.7% were prepared using distilled water, and
were maintained at 40°C to prevent solidification. A 3-ml mixture
of 1.2% agarose and 2X DMEM (containing 2X antibiotics and 20% FBS)
at a 1:1 ratio was cooled, solidified and placed in a
CO2 incubator for use as the bottom agar layer.
Subsequently, 0.7% agarose and 2X DMEM were mixed in a sterile test
tube at a 1:1 ratio, and 0.2 ml cell suspension was added, mixed
and injected into the 1.2% agarose layer to form a double agar
layer. Once solidified, the plates were placed in an incubator at
37°C (5% CO2) for 14 days. The plates were then placed
under an inverted microscope (magnification, ×100; Olympus
Corporation) to observe the colonies. Subsequently, the cells were
stained with 0.1% crystal violet (Thermo Fisher Scientific, Inc.)
at room temperature for 2 min and imaged. The number of colonies
(>50 cells) was quantified using ImageJ v1.8 software (National
Institutes of Health).
Western blotting
C33A cells were washed three times with 3 ml PBS and
precooled at 4°C to remove the medium. The PBS was discarded and
the flask containing the cells was placed on ice. To each flask,
400 µl PMSF-containing Cell Lysis Buffer (Cell signaling
Technology, Inc.) was added at 4°C for 20 min, followed by
centrifugation at 12,000 × g at 4°C for 5 min. The supernatant was
stored at −20°C. Protein concentration was quantified using a BCA
kit (Beyotime Institute of Biotechnology). 10% SDS-PAGE was then
performed for 5 h at 40 V to separate total protein (30 µg/lane).
The proteins were then transferred to a PVDF membrane and blocked
with 5% non-fat milk for 2 h at room temperature. The membranes
were incubated with primary antibodies at 4°C overnight, followed
by secondary antibodies for 2 h at room temperature; all antibody
details are presented in Table I.
The membranes were washed with TBS-0.1% Tween for chemiluminescence
detection with an ECL kit (Beyotime Institute of Biotechnology).
The molecular weight and net optical density of the target bands
were analyzed using ImageJ v1.8 software (National Institutes of
Health).
 | Table I.Antibodies used for western
blotting. |
Table I.
Antibodies used for western
blotting.
| Name | Cat. no. | Dilution | Species | Company |
|---|
| Ki67 | orb389335 | 1:500 | Rabbit | Biorbyt |
| PCNA | orb48485 | 1:1,000 | Rabbit | Biorbyt |
| MMP2 | orb13578 | 1:1,000 | Rabbit | Biorbyt |
| MMP9 | orb13583 | 1:250 | Rabbit | Biorbyt |
| PTCH1 | orb389650 | 1:500 | Rabbit | Biorbyt |
| SMO | orb46519 | 1:1,000 | Rabbit | Biorbyt |
| GLI1 | ab134906 | 1:1,000 | Rabbit | Abcam |
| Shh | ab53281 | 1:1,000 | Rabbit | Abcam |
| GAPDH | orb555879 | 1:10,000 | Rabbit | Biorbyt |
| IgG H&L
(HRP) | ab6721 | 1:3,000 | Goat
anti-rabbit | Abcam |
Wound-healing assay
C33A cells (5×105 cells/well) were seeded
into a 6-well plate and cultured in DMEM containing 10% FBS at 37°C
until 80% confluent, and subsequently serum-starved overnight
(28). A 200-µl pipette tip was
used to create a wound in the cell monolayer, and the cells were
then cultured in serum-free DMEM with TMP (25, 50 and 100 µM) or
TMP (100 µM) + SAG. Following incubation at 37°C for 48 h, the
cells were washed with PBS, and the wounds were imaged using an
inverted light microscope (magnification, ×100; Olympus
Corporation). The results were analyzed using ImageJ v1.8 software
(National Institutes of Health), and migration rate was calculated
as follows: Cell migration rate=wound area difference between 0 and
48 h/wound area at 0 h ×100%.
Transwell assay
Matrigel (Corning, Inc.) was added into the upper
chamber of the Transwell plate at 37°C for 30 min, and C33A cells
(5×104 cells) were pre-incubated at 37°C in DMEM with
TMP (25, 50 and 100 µM) or TMP (100 µM) + SAG for 48 h. The cells
were then resuspended in DMEM, seeded into the upper chamber and
incubated at 37°C for 48 h. The cells remaining in the upper
chamber were removed with a cotton swab, and those in the lower
chamber were fixed with 10% formaldehyde at room temperature for 30
min. The cells were then stained with crystal violet at room
temperature for 20 min, and subsequently observed and counted under
an inverted light microscope (magnification, ×100; Olympus
Corporation). The result was analyzed using ImageJ v1.8 software
(National Institutes of Health), and the invasion rate was
calculated as follows: Cell invasion rate=the number of invasive
cells/number of inoculated cells ×100%.
Statistical analysis
All experiments were performed in triplicate; the
data were analyzed using SPSS 20.0 (IBM Corp.) and are presented as
the mean ± SD. One-way ANOVA followed by a Tukey's post hoc test
was used to analyze statistical differences, and P<0.05 was
considered to indicate a statistically significant difference.
Results
TMP inhibits the survival of C33A
cells
To investigate how TMP affects the survival of CC
cells, an MTT assay was performed. The results demonstrated that
TMP had no significant effect on Ect1 cell viability, whereas TMP
decreased C33A cell survival in a dose-dependent manner (Fig. 1B). These results indicated that TMP
impeded the survival of CC cells.
TMP inhibits C33A cell
proliferation
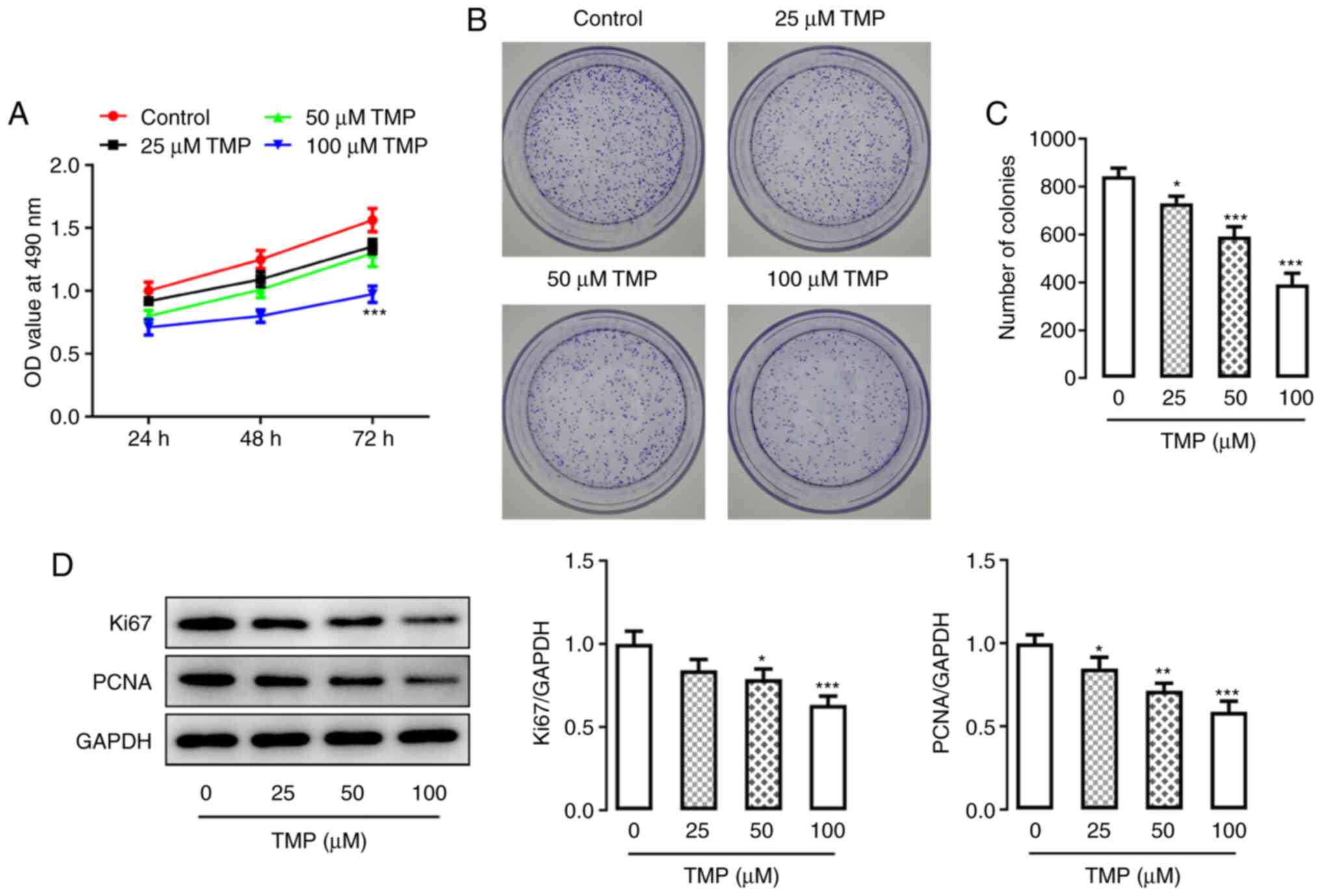
A series of experiments were performed to determine
whether TMP affected the proliferation of CC cells. C33A cell
proliferation was quantified at 24, 48 and 72 h, demonstrating a
general ascending trend over time. However, in response to an
increase in TMP dose, the results demonstrated significantly
decreased C33A cell proliferation at 72 h (100 µM; Fig. 2A). Furthermore, the colony
formation assay demonstrated that TMP treatment inhibited the
clonogenicity of C33A cells in a dose-dependent manner (Fig. 2B and C). The expression of
proliferation-related proteins, Ki67 and proliferating cell nuclear
antigen (PCNA), was detected via western blotting and both
exhibited reduced expression levels following TMP, which occurred
in a dose-dependent manner (Fig.
2D). These results demonstrated the possible inhibitory effect
of TMP treatment on CC cell proliferation.
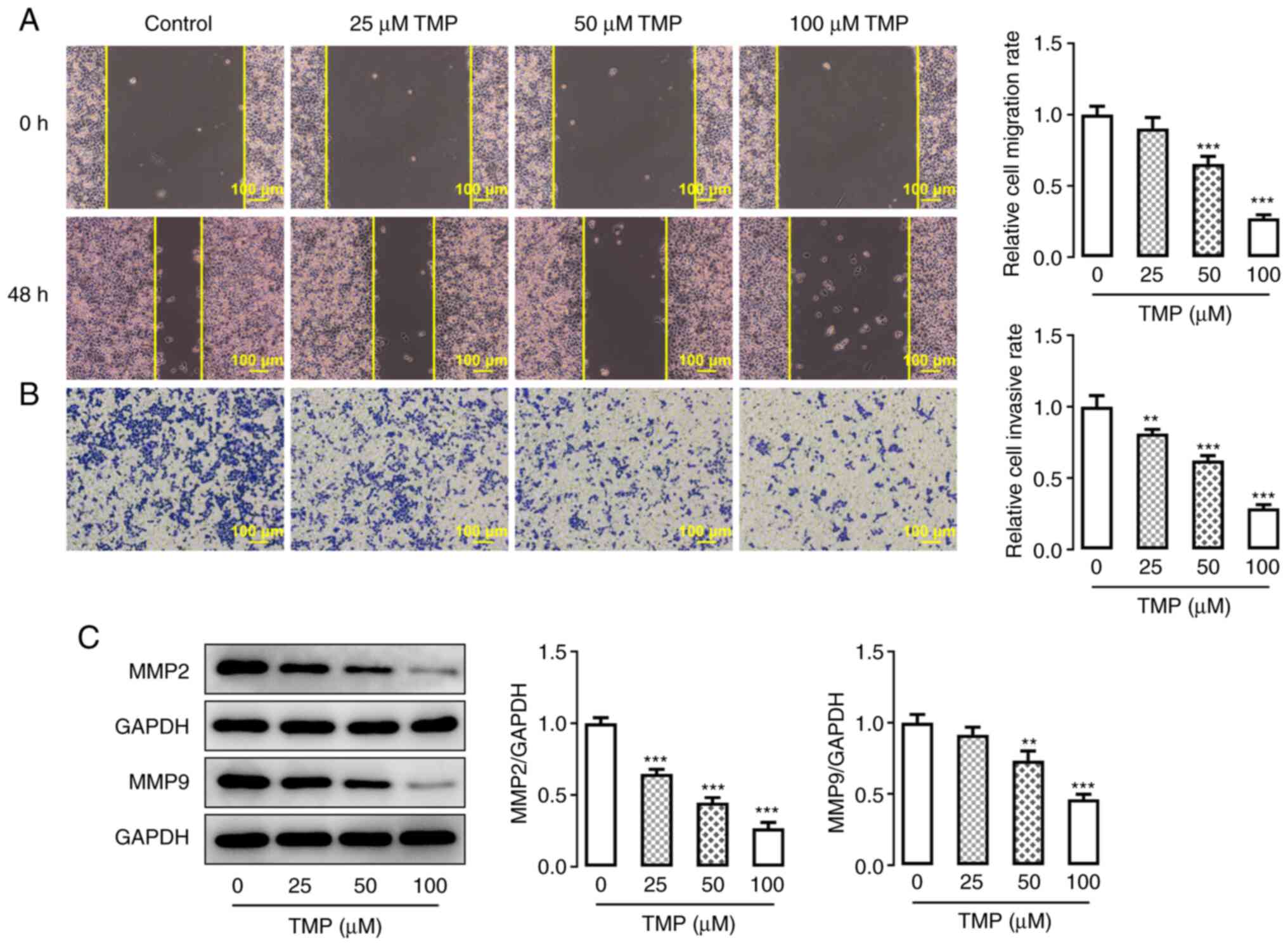
TMP inhibits the invasion and
migration abilities of C33A cells
The invasiveness and migration of C33A cells were
detected using Transwell and wound-healing assays, respectively.
Both invasion and migration ability were significantly decreased in
response to an increase in TMP concentration (Fig. 3A and B). Furthermore, the protein
expression levels of migration-related proteins were further
evaluated using western blotting. Similar to the results of the
wound-healing and Transwell assays, it was demonstrated that the
protein expression levels of MMP2 and MMP9 were negatively
associated with an increase in TMP dose (Fig. 3C). These results indicated that TMP
had an inhibitory effect on the invasion and migration capacities
of C33A cells.
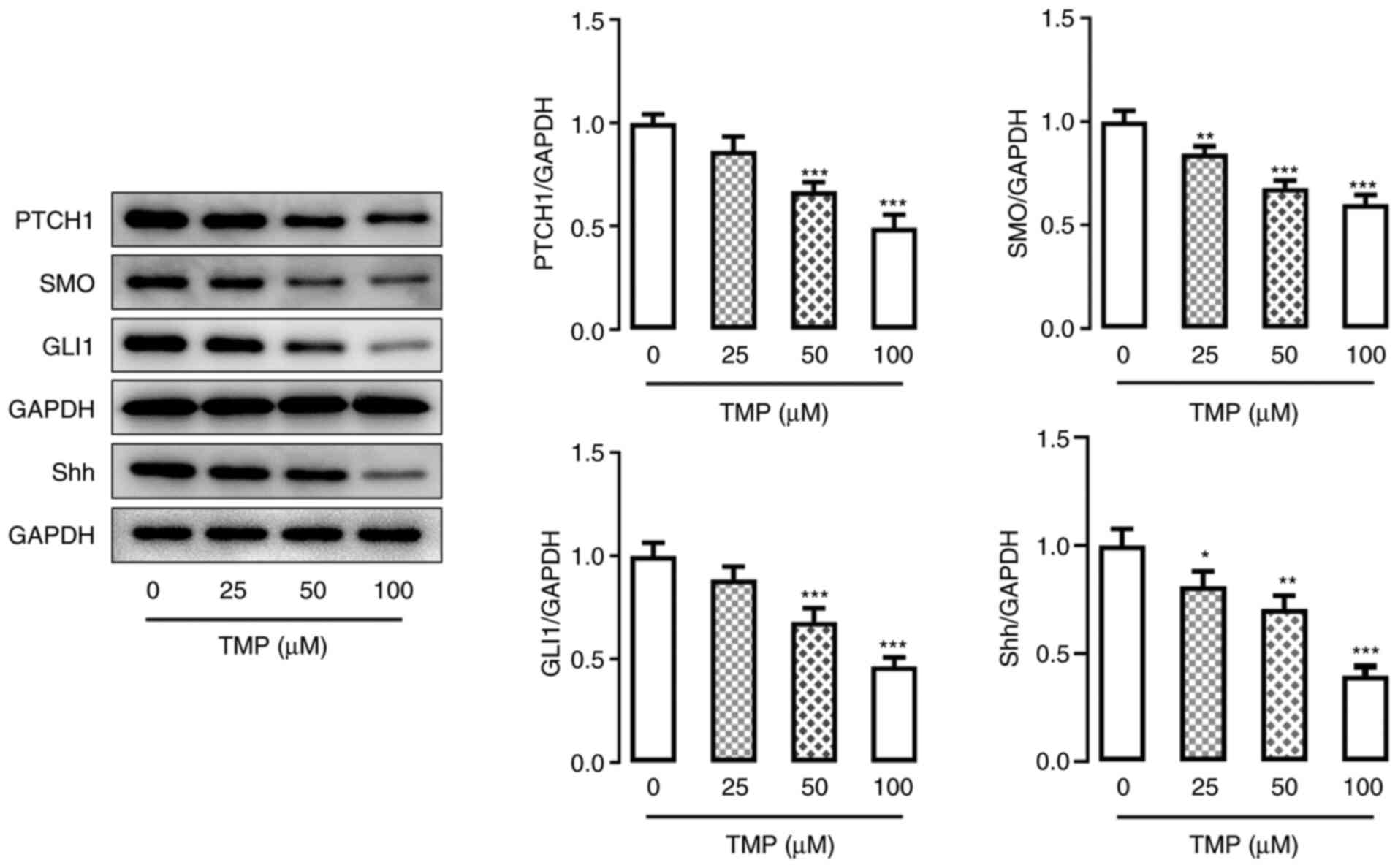
TMP treatment inhibits the Hh
signaling pathway in C33A cells
To understand the mechanism of TMP, its potential
regulatory effects on the Hh signaling pathway were investigated
using CC cells. The protein expression levels of Hh
signaling-related proteins patched 1, smoothened homolog precursor,
GLI family zinc finger 1 and sonic hedgehog, were assessed by
western blotting. The results demonstrated that these proteins were
all markedly reduced in C33A cells following TMP treatment
(Fig. 4), and in a dose-dependent
manner. This result revealed that TMP may block the Hh signaling
pathway in CC cells.
TMP inhibits the proliferation,
invasiveness and migration of C33A cells by retarding the Hh
signaling pathway
To further elucidate the mechanism underlying TMP
treatment, and to verify its effect on the Hh signaling pathway,
C33A cell proliferation, invasion and migration abilities were
investigated in the presence of a Hh signaling pathway agonist
(SAG) and 100 µM TMP. Western blotting verified the increased
activation of the Hh signaling pathway, which was evidenced by the
upregulated protein expression levels of Hh signaling
pathway-related proteins in TMP-treated C33A cells following SAG
inducement (Fig. 5A). Moreover,
the MTT assay demonstrated that the inhibited proliferation of C33A
cells following TMP treatment was reversed by SAG inducement,
compared with the control group (Fig.
5B). Furthermore, the colony formation assay demonstrated a
reduction in the number of C33A colonies formed when treated with
TMP (compared with the control), which was markedly reversed by SAG
treatment (Fig. 5C and D). Western
blotting also detected increased protein expression levels of the
proliferation-related proteins Ki67 and PCNA in TMP-treated C33A
cells induced by SAG, which were originally inhibited by TMP
(Fig. 5E). Moreover, wound-healing
and Transwell assays demonstrated that both the migration and
invasion abilities of TMP-treated C33A cells were increased
following SAG inducement (Fig. 5F and
G). The protein expression levels of migration-related proteins
MMP2 and MMP9 also exhibited a similar trend (Fig. 5H). These results collectively
indicated that TMP may block the Hh signaling pathway, and thereby
inhibit the proliferation, invasiveness and migration of CC
cells.
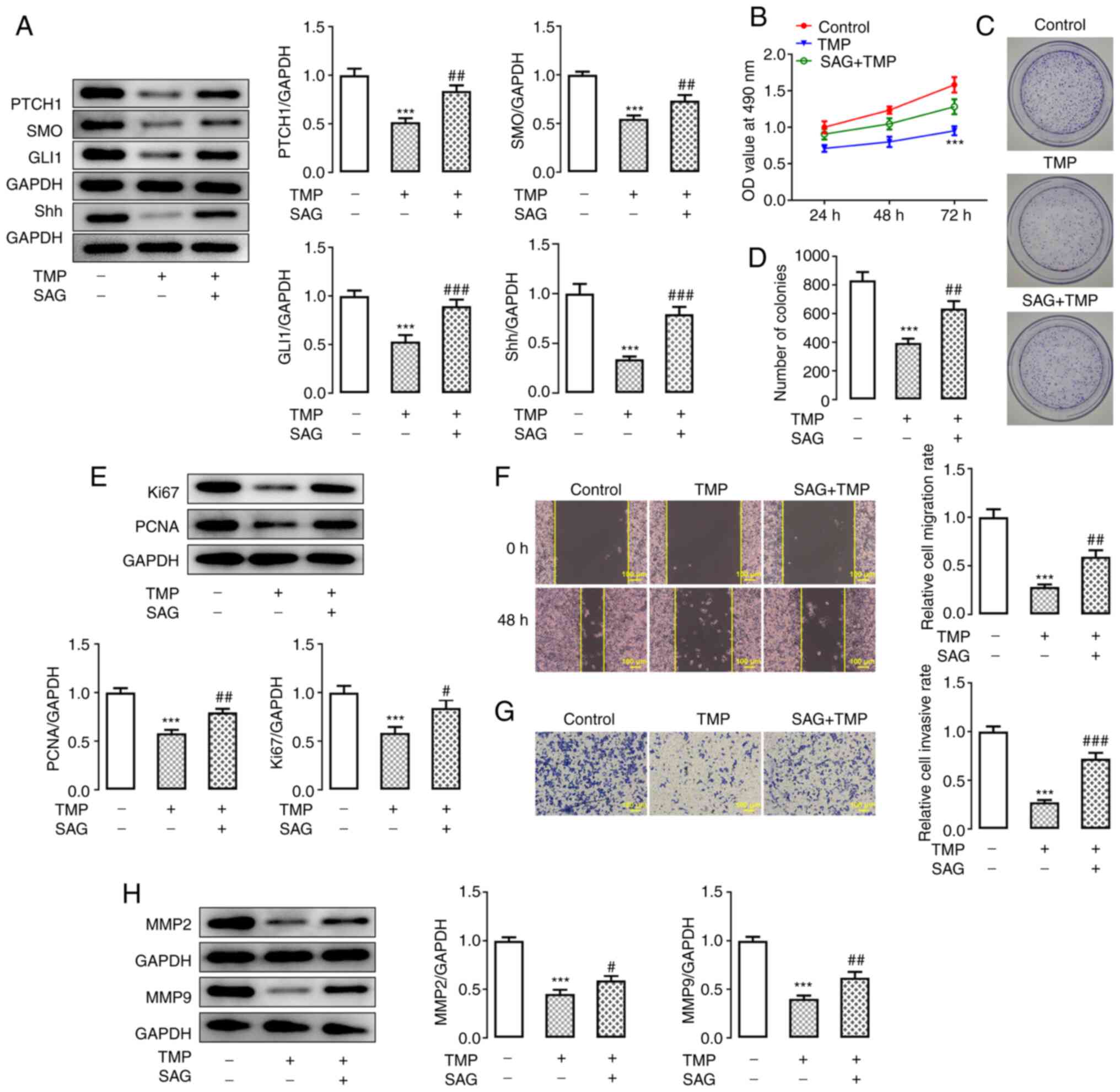 | Figure 5.TMP inhibits the proliferation,
invasion and migration abilities of C33A cells by retarding the Hh
signaling pathway. (A) Protein expression levels of Hh signaling
pathway-related proteins in TMP-treated C33A cells, before and
after SAG inducement, were detected via western blotting. (B)
Proliferation of TMP-treated C33A cells before and after SAG
inducement was detected using an MTT assay. (C and D) Colony
formation in TMP-treated C33A cells, before and after SAG
inducement, was detected using a colony formation assay. (E)
Protein expression levels of proliferation-related proteins in
TMP-treated C33A cells, before and after SAG inducement, were
detected via western blotting. (F and G) Migration and invasiveness
of TMP-treated C33A cells, before and after SAG inducement, were
detected using wound-healing and Transwell assays, respectively.
(H) Protein expression levels of migration-related proteins in
TMP-treated C33A cells, before and after SAG inducement, were
detected via western blotting. ***P<0.001 vs. control;
#P<0.05, ##P<0.01 and
###P<0.001 vs. TMP. N≥3. TMP, tetramethylpyrazine;
PTCH1, patched 1; SMO, smoothened homolog precursor; GLI1, GLI
family zinc finger 1; Shh, sonic hedgehog. |
Discussion
CC is a primary malignant tumor of the cervix, which
is mainly caused by persistent high-risk HPV infection (29). Regular CC screening, management of
cervical precancerous lesions and HPV vaccinations are currently
the primary preventative measures for CC (29). Screening and management of
precancerous lesions contribute to the prognosis of CC, whereas HPV
vaccinations prevent infection from certain strains of HPV; however
these preventative measures have limitations (30). For example, a considerable number
of CCs have been confirmed to be HPV negative (31). As aforementioned, TMP was found to
serve an inhibitory role in numerous types of cancer (17–19).
The present study demonstrated that TMP reduced the survival rate
of CC cells in a dose-dependent manner. TMP also effectively
inhibited CC cell proliferation and colony formation, and reduced
proliferation-related protein expression. Furthermore, the
dose-dependent attenuation of CC cell migration and invasiveness by
TMP was also demonstrated. These results indicated that TMP may
exert a protective effect against CC.
In order to further investigate the mechanism
underlying the effects of TMP in CC, its effects on the Hh
signaling pathway were determined. A previous study demonstrated
that TMP disrupted the Hh signaling pathway to reduce sinusoidal
tubule angiogenesis, and to inhibit the angiogenic properties of
liver sinusoidal endothelial cells (32). Another study reported that a
compound extracted from Artemisia plants reduced the proliferative
index of HeLa and Caski CC cells through the Hh signaling pathway,
and inhibited tumor growth in tumor-bearing mice (33). Furthermore, sonic Hh gene silencing
has been demonstrated to inhibit the epithelial-mesenchymal
transition, as well as the proliferation, invasiveness and
migration of CC cells by inhibiting the Hh signaling pathway
(34). In the present study, the
levels of Hh signaling pathway-related protein expression in CC
cells were negatively associated with TMP in a dose-dependent
manner. To further verify the roles of the Hh signaling pathway and
TMP, CC cells were treated with the Hh agonist SAG (27). SAG induction of TMP-treated CC
cells reversed the anti-proliferative, anti-invasive and
anti-effects of TMP on CC cells.
In conclusion, the present study indicated that TMP
protected against CC, and inhibited the proliferation, invasiveness
and migration of CC cells by blocking the Hh signaling pathway.
These results highlighted the potential for a novel therapy for the
prevention or treatment of CC. In order to fully utilize TMP and
apply it in clinical practice for CC, more in-depth research is
required to determine its mechanism of action. future research will
include establishing animal cervical cancer models to verify the
effects of TMP in vivo.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
JR helped to draft the manuscript and performed the
experiments. JC performed the experiments and data analysis. CW
revised the manuscript and helped to design the experiments. All
authors read and approved the final manuscript. JR and CW confirm
the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vu M, Yu J, Awolude OA and Chuang L:
Cervical cancer worldwide. Curr Probl Cancer. 42:457–465. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ou R, Lv J, Zhang Q, Lin F, Zhu L, Huang
F, Li X, Li T, Zhao L, Ren Y and Xu Y: circAMOTL1 Motivates AMOTL1
expression to facilitate cervical cancer growth. Mol Ther Nucleic
Acids. 19:50–60. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ki EY, Lee KH, Park JS and Hur SY: A
clinicopathological review of pulmonary metastasis from uterine
cervical cancer. Cancer Res Treat. 48:266–272. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hwang JH, Yoo HJ, Lim MC, Seo SS, Kang S,
Kim JY and Park SY: Brain metastasis in patients with uterine
cervical cancer. J Obstet Gynaecol Res. 39:287–291. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hong JH, Tsai CS, Lai CH, Chang TC, Wang
CC, Chou HH, Lee SP and Hsueh S: Recurrent squamous cell carcinoma
of cervix after definitive radiotherapy. Int J Radiat Oncol Biol
Phys. 60:249–257. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li H, Wu X and Cheng X: Advances in
diagnosis and treatment of metastatic cervical cancer. J Gynecol
Oncol. 27:e432016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wendel Naumann R and Leath CA III:
Advances in immunotherapy for cervical cancer. Curr Opin Oncol.
32:481–487. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang A, Farmer E, Wu TC and Hung CF:
Perspectives for therapeutic HPV vaccine development. J Biomed Sci.
23:752016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lopez MS, Baker ES, Maza M, Fontes-Cintra
G, Lopez A, Carvajal JM, Nozar F, Fiol V and Schmeler KM: Cervical
cancer prevention and treatment in Latin America. J Surg Oncol.
115:615–618. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao Y, Liu Y and Chen K: Mechanisms and
clinical application of tetramethylpyrazine (an interesting natural
compound isolated from Ligusticum Wallichii): Current status
and perspective. Oxid Med Cell Longev. 2016:21246382016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi J, Li R, Yang S, Phang Y, Zheng C and
Zhang H: The protective effects and potential mechanisms of
Ligusticum chuanxiong: Focus on anti-inflammatory,
antioxidant, and antiapoptotic activities. Evid Based Complement
Alternat Med. 2020:82059832020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan Z, Zhang J and Yang C: Ligusticum
wallichii extract inhibited the expression of IL-1β after AMI
in Rats. Evid Based Complement Alternat Med. 2014:6203592014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao Z and Moghadasian MH: Chemistry,
natural sources, dietary intake and pharmacokinetic properties of
ferulic acid: A review. Food Chem. 109:691–702. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu B, Liu M, Liu H, Li W, Tan S, Zhang S
and Fang Y: Meta-analysis of traditional Chinese patent medicine
for ischemic stroke. Stroke. 38:1973–1979. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang P, She G, Yang Y, Li Q, Zhang H, Liu
J, Cao Y, Xu X and Lei H: Synthesis and biological evaluation of
new ligustrazine derivatives as anti-tumor agents. Molecules.
17:4972–4985. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin LN, Wang WT and Xu ZJ: Clinical study
on ligustrazine in treating myocardial ischemia and reperfusion
injury. Zhongguo Zhong Xi Yi Jie He Za Zhi. 17:261–263. 1997.(In
Chinese). PubMed/NCBI
|
|
17
|
Zhou Y, Ji Z, Yan W, Zhou Z, Li H and Xiao
Y: Tetramethylpyrazine inhibits prostate cancer progression by
downregulation of forkhead box M1. Oncol Rep. 38:837–842. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang HH, Liu FB, Ruan Z, Zheng J, Su YJ
and Wang J: Tetramethylpyrazine (TMPZ) triggers S-phase arrest and
mitochondria-dependent apoptosis in lung cancer cells. Neoplasma.
65:367–375. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang S, Lei T and Zhang M: The reversal
effect and its mechanisms of tetramethylpyrazine on multidrug
resistance in human bladder cancer. PLoS One. 11:e01577592016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Y, Zhou Z, Ji Z, Yan W, Li H and Yu
X: Tetramethylpyrazine reduces prostate cancer malignancy through
inactivation of the DPP10AS1/CBP/FOXM1 signaling pathway. Int J
Oncol. 57:314–324. 2020.PubMed/NCBI
|
|
21
|
Shen J, Zeng L, Pan L, Yuan S, Wu M and
Kong X: Tetramethylpyrazine regulates breast cancer cell viability,
migration, invasion and apoptosis by affecting the activity of Akt
and caspase-3. Oncol Lett. 15:4557–4563. 2018.PubMed/NCBI
|
|
22
|
Skoda AM, Simovic D, Karin V, Kardum V,
Vranic S and Serman L: The role of the Hedgehog signaling pathway
in cancer: A comprehensive review. Bosn J Basic Med Sci. 18:8–20.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu F, Zhang Y, Sun B, McMahon AP and Wang
Y: Hedgehog signaling: From basic biology to cancer therapy. Cell
Chem Biol. 24:252–280. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu J, Cao G, Wu X, Cai H and Cai B:
Tetramethylpyrazine inhibits activation of hepatic stellate cells
through hedgehog signaling pathways in vitro. Biomed Res Int.
2015:6030672015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang C, Lu H, Li J, Xie X, Fan L, Wang D,
Tan W, Wang Y, Lin Z and Yao T: SOX2 regulates radioresistance in
cervical cancer via the hedgehog signaling pathway. Gynecol Oncol.
151:533–541. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luan Y, Liu J, Liu X, Xue X, Kong F, Sun
C, Wang J, Liu L and Jia H: Tetramethypyrazine inhibits renal cell
carcinoma cells through inhibition of NKG2D signaling pathways. Int
J Oncol. 49:1704–1712. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Feng J, Wang C, Liu T, Li J, Wu L, Yu Q,
Li S, Zhou Y, Zhang J, Chen J, et al: Procyanidin B2 inhibits the
activation of hepatic stellate cells and angiogenesis via the
Hedgehog pathway during liver fibrosis. J Cell Mol Med.
23:6479–6493. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shen X, Li L, He Y, Lv X and Ma J:
Raddeanin A inhibits proliferation, invasion, migration and
promotes apoptosis of cervical cancer cells via regulating
miR-224-3p/Slit2/Robo1 signaling pathway. Aging. 13:7166–7179.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
de Sanjosé S, Brotons M and Pavón MA: The
natural history of human papillomavirus infection. Best Pract Res
Clin Obstet Gynaecol. 47:2–13. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Menderes G, Black J, Schwab CL and Santin
AD: Immunotherapy and targeted therapy for cervical cancer: An
update. Expert Rev Anticancer Ther. 16:83–98. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kaliff M, Karlsson MG, Sorbe B, Bohr
Mordhorst L, Helenius G and Lillsunde-Larsson G: HPV-negative
tumors in a swedish cohort of cervical cancer. Int J Gynecol
Pathol. 39:279–288. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao S, Zhang Z, Yao Z, Shao J, Chen A,
Zhang F and Zheng S: Tetramethylpyrazine attenuates sinusoidal
angiogenesis via inhibition of hedgehog signaling in liver
fibrosis. IUBMB Life. 69:115–127. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu Z, Zou B, Zhang X and Peng X: Eupatilin
regulates proliferation and cell cycle of cervical cancer by
regulating hedgehog signalling pathway. Cell Biochem Funct.
38:428–435. 2020. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang F, Ren CC, Liu L, Chen YN, Yang L,
Zhang XA, Wang XM and Yu FJ: SHH gene silencing suppresses
epithelial-mesenchymal transition, proliferation, invasion, and
migration of cervical cancer cells by repressing the hedgehog
signaling pathway. J Cell Biochem. 119:3829–3842. 2018. View Article : Google Scholar : PubMed/NCBI
|