Introduction
Approximately one in three individuals are, or have
been previously, infected with Toxoplasma gondii (T.
gondii) at varying degrees (1,2).
Once the body's immune function is impaired, T. gondii may
opportunistically cause diseases (3). Furthermore, T. gondii
infection may lead to alterations in the expression of certain
microRNAs (miRNAs/miRs) in their hosts. Thus, differentially
expressed miRNAs from two genetically distinct strains of T.
gondii were able to be developed as diagnostic biomarkers for
toxoplasmosis (4). T.
gondii infection was reported to specifically increase the
levels of key host miRNAs (5).
Comparison of splenocyte miRNA expression in pigs during acute and
chronic infections indicated that differentially expressed miRNAs
have important roles in the host's immune response to T.
gondii infection by modulating the expression levels of
cellular immunity-related cytokines and immune-related C-type
lectins (6). The miRNA data from
porcine alveolar macrophages infected with T. gondii first
demonstrated the association between miRNAs and macrophages of
swine origin (7). Differential
brain miRNA expression in mice infected with T. gondii
oocysts indicated that T. gondii infection may alter the
abundance of miRNAs in the mouse brain, particularly at the chronic
stage (8).
Previous studies have indicated that T.
gondii infection is able to improve immunity against cancer in
the host (9–13). A previous study by our group
revealed changes in tumor-related factors after T. gondii
infection (14). As infection with
T. gondii may change the levels of miRNAs in hosts, it is
necessary to analyze the expression of the targets of the
cancer-related miRNAs in the host pre- and post-infection with
T. gondii.
Materials and methods
Animal experiment
Female BALB/c mice (age, 8 weeks; mean weight,
20.25±0.93 g) were obtained from the Shandong University Laboratory
Animal Center. A total of 48 mice were randomly divided into two
groups (control group, n=24; infected group, n=24). They were
reared in groups of six mice per cage under specific pathogen-free
conditions under a 12-h light/dark cycle at 25°C and had ad
libitum access to tap water and self-produced mouse feed
(8). All animal experiments were
approved by the Ethics Committee on Animal Experiments of the
Medical School of Shandong University (Jinan, China). Each BALB/c
mouse (infected group) was challenged with 10 T. gondii
cysts by gavage and the spleens of experimental mice were collected
for RNA extraction one month after the challenge. All efforts were
made to minimize suffering and humane endpoints were used. Mice
with unkempt fur and diarrhea were euthanized. For euthanasia, mice
were placed in a chamber and CO2 was administered at a
concentration of 60–70% over a 5-min exposure time, after which the
cervical dislocation method was at times used to ensure that
effective euthanasia had occurred.
Parasite
T. gondii (low virulent Prugniaud strain,
obtained from the Department of Pathogen Biology, Anhui Medical
University, Hefei, China) was maintained in our laboratory using
the passage of cysts in eight-week-old Kunming mice obtained from
Shandong University Laboratory Animal Center (mean weight,
42.31±5.31 g).
RNA extraction and high-throughput
sequencing
RNA was obtained from different spleen samples (one
mouse from the control group and one mouse from the infected group)
using TRIzol reagent (Takara Bio, Inc.) according to the
manufacturer's instructions. RNA was isolated using the improved
cetyltrimethylammonium bromide (CTAB) method, applying isopropanol
instead of lithium chloride for RNA precipitation. In brief, one
gram of spleen sample was added to liquid nitrogen and ground into
a fine powder, which was evenly mixed in 5 ml preheated (65°C)
extraction buffer (2% CTAB, 2% polyvinylpyrrolidone, 0.1 M
Tris-HCl, 2.0 M NaCl, 25 mM EDTA, 2% beta-mercaptoethanol; pH 8.0).
The mixture was incubated for 5 min at 65°C and shaken three times
during the incubation period. After a short cooling period,
isopropanol (2.5 ml) was added to the mixture, after which the
mixture was vortexed for 1 min and centrifuged at 10,900 × g for 15
min at 4°C. DNase was then used to treat the extract and RNA was
precipitated at 25°C for 10 min using the same volume of
isopropanol. The extracted RNA was first resuspended in an equal
volume of phenol/chloroform/isopropanol mixture (25:24:1) and then
in an equal volume of chloroform/isopropanol (24:1). Both a 0.1
volume of 3M NaOAC (pH 5.2) and 2.5 volumes of cold ethanol were
added to the mixture to precipitate the RNA overnight at −20°C. The
quality of the prepared RNA from each sample was analyzed using an
Agilent 2100 Bioanalyzer (Agilent Technologies, Inc.). The
fragmentation buffer was added to cut the mRNA into short fragments
(200–700 nucleotides). First-strand cDNA was synthesized using the
templates of the short fragments. DNA polymerase I (New England
Biolabs, Inc.), dNTPs, RNase H (Invitrogen; Thermo Fisher
Scientific, Inc.) and buffer were used to synthesize the
second-strand cDNA. The fragments were purified using the QiaQuick
PCR kit and washed with EB buffer (consists of sodium chloride,
magnesium chloride, HEPES and sucrose) for end repair prior to
adding polyA tails and adaptors. Fragments of suitable size were
detected by agarose gel electrophoresis and amplified using PCR
(8), after which the products were
sequenced using Illumina HiSeq™ 2000 (Illumina, Inc.).
Bioinformatics analysis and
identification of miRNA targets
A total of two rat miRNA transcriptome libraries
were sequenced using the Illumina HiSeq 2000 platform. Details of
the raw data are listed in Table
I. High-quality 20±21 nucleotide-long reads were processed with
the CleaveL pipeline for small RNA target identification, as
previously described (15).
Sequences of ribosomal RNAs, transfer RNAs, small nucleolar RNAs
and small nuclear RNAs were retrieved from the RNA families
database (xfam.org). The control (S01) and infected (S02) sample
data were analyzed separately. P≤0.01 and fold change ≥2 were used
to identify significant differentially expressed genes. The miRDB
software (16) was used to predict
the target genes of differentially expressed miRNAs.
 | Table I.Raw data before filtering of
sequencing results. |
Table I.
Raw data before filtering of
sequencing results.
| Group | Raw data (bp) | Raw reads | Q30 (%) | Clean reads |
|---|
| S01 | 1407676653 | 27601503 | 98.17 | 27295019 |
| S02 | 1398651642 | 27424542 | 98.15 | 27008080 |
Gene ontology (GO) functional
enrichment and kyoto encyclopedia of genes and genomes (KEGG)
pathway analysis
The selected sequence was constructed using BLASTX
and the National Center for Biotechnology Information (NCBI)
database (https://www.ncbi.nlm.nih.gov/guide/sequence-analysis/)
to better observe the function of miRNA targets and the metabolic
regulatory networks related to mouse miRNAs. BLASTX searches using
the InterPro and KEGG databases were used to collect the predicted
target proteins with an E-value of 1×1030. Target gene
function and metabolic pathways of miRNAs were verified using the
best hits. Finally, the terms in the categories molecular function,
biological process and cellular component of target genes were
obtained using the GO and InterPro databases (https://ngdc.cncb.ac.cn/databasecommons/). The GO
terms and KEGG pathways with P≤0.01 and fold change ≥2 were
considered significant.
miRNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
miRNAs were extracted from spleen samples using the
miRNeasy Serum/Plasma Kit (Qiagen GmbH) according to the
manufacturer's protocol. Subsequently, miRNA was reverse
transcribed into cDNA using the One Step PrimeScript & Reg
miRNA cDNA Synthesis Kit (Takara Bio, Inc.). PCR was performed with
the Prime-Script™ RT reagent kit (Takara Bio, Inc.) according to
the manufacturer's protocol and in an ABI 7000 real-time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) using the
following thermocycling conditions: 95°C for 5 sec and 58°C for 20
sec for 35 cycles, and 72°C for 30 sec. The experiments were
performed once and set up in triplicate. The mRNA levels of large
tumor suppressor kinase (Lats)2, Lats1 and TNF receptor superfamily
member 11b (Tnfrsf11b) were detected by qPCR with the
2−∆∆Cq quantification method (17) and the primers are listed in
Table II.
| Table II.Primers used for quantitative PCR
(5′-3′). |
Table II.
Primers used for quantitative PCR
(5′-3′).
| Gene | Forward primer | Reverse primer |
|---|
| Lats1 |
GTGCAACATTCAATTAACCG |
TCCAGACAGAGGTCTTCCTA |
| Lats2 |
TGAGCAGATTGTGCGAGTCA |
GCGGCGGGGCCCTCGTAGTT |
| Tnfrsf11b |
TGTGCTGCGCACTCCTGGTGC |
TGCAGTGCTGTTTTAGGTAGG |
| β-actin |
TAGGCACCAGGGTGTGATGG |
GTGCCAGATCTTCTCCATGTC |
Luciferase activity assay
Selected sequences from wild-type (WT) 3′-UTRs were
cloned into the pMir-reporter vector (Ambion; Thermo Fisher
Scientific, Inc.) and the mutant 3′-UTRs were generated by altering
the predicted miR-31-5p, miR-135a-5p, miR-145a-5p, miR-204-5p,
miR-211-5p and miR-429-3p 3′-UTR binding sites via a two-step PCR
approach (the template was obtained from mouse miRNAs) (18). The miRNA mimics are listed in
Table III. 293-T cells were
purchased from FuXiang Biotechnology Co. 293-T cells were
cultivated in Dulbecco's modified Eagles medium (DMEM) with
antibiotics and Fetal Bovine Serum at 37°C in a humidified 5%
CO2 atmosphere. Cells were co-transfected with either a
WT or mutant 3′-UTR reporter vector and miR-31-5p, miR-135a-5p,
miR-145a-5p, miR-204-5p, miR-211-5p and miR-429-3p mimics or
negative control constructs and were cultured for 24 h. The cells
were then assessed to ascertain their luciferase activity using a
dual-luciferase reporter assay system (Promega Corporation). The
experiments were performed once and set up in triplicate.
| Table III.Sequences for miRNA mimics. |
Table III.
Sequences for miRNA mimics.
| miRNA | Mimic |
|---|
| miR-31-5p |
AGGCAAGAUGCUGGCAUAGCUG |
| miR-135a-5p |
UAUGGCUUUUUAUUCCUAUGUGA |
| miR-145a-5p |
GUCCAGUUUUCCCAGGAAUCCCU |
| miR-204-5p |
UUCCCUUUGUCAUCCUAUGCCU |
| miR-211-5p |
UUCCCUUUGUCAUCCUUUGCCU |
| miR-429-3p |
UAAUACUGUCUGGUAAUGCCGU |
Western blot analysis
In brief, the samples were treated with RIPA lysis
buffer (50 mM Tris, pH 7.4, 150 mM NaCl, 1% Triton X-100, 1% sodium
deoxycholate, 0.1% SDS) containing 1 mM protease inhibitor
phenylmethanesulfonyl fluoride and centrifuged at 12,100 × g at 4°C
for 10 min. The supernatant was then separated and mixed with 50 µl
of SDS-PAGE sample buffer and boiled for 5 min, after which 5 µg
(the protein concentration was determined by bicinchoninic acid
protein assay kit) per lane was loaded onto the polyacrylamide gel
and SDS-PAGE was performed with a 10% gel. Proteins were
transferred onto polyvinylidene fluoride membranes (Beyotime
Institute of Biotechnology) via electrophoresis, performed at 80 V
for 3 h, using a Bio-Rad transfer system (Bio-Rad Laboratories,
Inc.). The membranes were blocked for 2 h with skimmed milk
(Beyotime Institute of Biotechnology) at room temperature and
probed with the corresponding antibody (rabbit) diluted in blocking
buffer at 1:10,000 (anti-LATS1, cat. no. ab70561; anti-LATS2, cat.
no. ab111054; Anti-Tnfrsf11b, cat. no. ab183910; all from Abcam) at
4°C for 12 h. The membrane was then incubated for 2 h at room
temperature with horseradish peroxidase-labeled goat anti-rabbit
IgG antibody (cat. no. 18772; MilliporeSigma), diluted in blocking
buffer at 1:20,000, and signals were detected with a
super-sensitive signal enhanced chemiluminescence system (Beyotime
Institute of Biotechnology). The levels of Lats2, Lats1 and
Tnfrsf11b proteins were measured in infected and control mice in
this experiment. The experiments were performed once and set up in
triplicate.
Statistical analysis
Values are expressed as the mean ± standard
deviation. Statistically significant differences between two groups
were determined using Student's t-test. ANOVA and least-significant
difference test were used for comparison among multiple groups. The
results of the western blot were analyzed using ImageJ software
(v1.51; National Institutes of Health). P<0.05 was considered to
indicate a statistically significant difference.
Results
Identification of miRNAs in the
presence or absence of T. gondii
A total of 1,899 miRNAs were identified after a
series of filtering. Of these, 327 miRNAs were identified as known
miRNAs, whereas 477 were identified as homologs in miRbase.
Specifically, the expression of 1,000 miRNAs was detected in the
control group, among which 320 miRNAs were known miRNAs and 362
were novel miRNAs. Furthermore, the expression of 996 miRNAs was
detected in the infected group, of which 304 were known miRNAs and
202 were novel miRNAs. Of the 1,899 miRNAs identified, 114 were
specifically expressed in the control group, while 110 were
specifically expressed in the infected group. Furthermore, it was
observed that most of the known miRNAs, such as miR-99a-5p,
miR-143-5p and miR-27b-5p, were highly expressed in the control and
infected groups, which was not unexpected. However, various novel
miRNAs exhibited low expression or were expressed in only one group
(Table SI).
Differentially expressed miRNAs
following exposure to T. gondii
The differentially expressed miRNAs identified are
listed in Table SII. The
expression levels of a total of 398 miRNAs were significantly
changed (P<0.05; cut-off criterion: Fold change ≥2) after T.
gondii challenge. Compared with the control group, 111 miRNAs
were upregulated and 287 miRNAs were downregulated in the infected
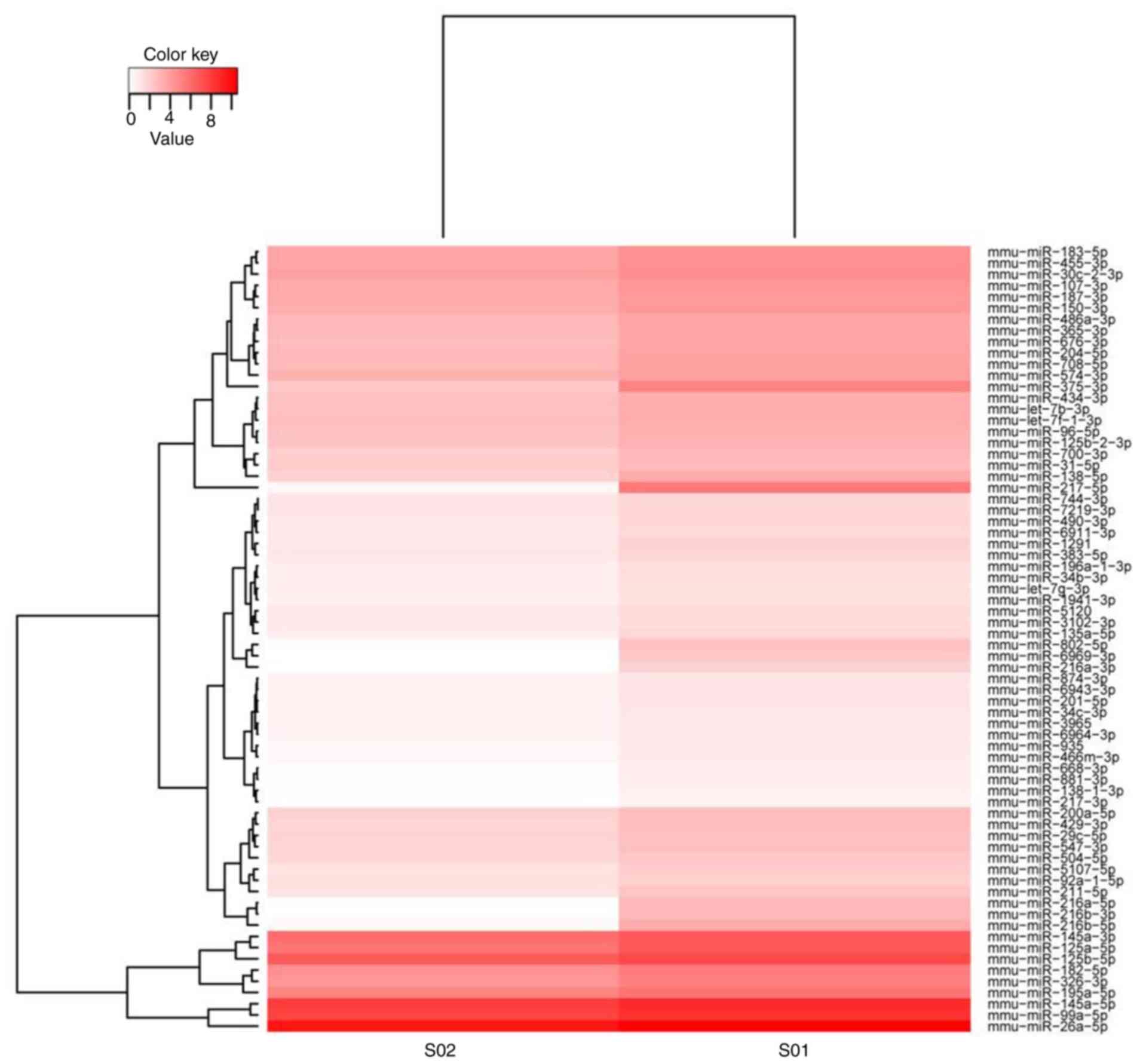
group. The heatmap indicated that 72 miRNAs were markedly
differentially expressed (P<0.01; cut-off criterion: Fold change
≥3) between the infected and control groups. Among these
differentially expressed miRNAs, 18 were upregulated and 54 were
downregulated (Fig. 1).
GO and KEGG enrichment analyses of
cancer-related target genes
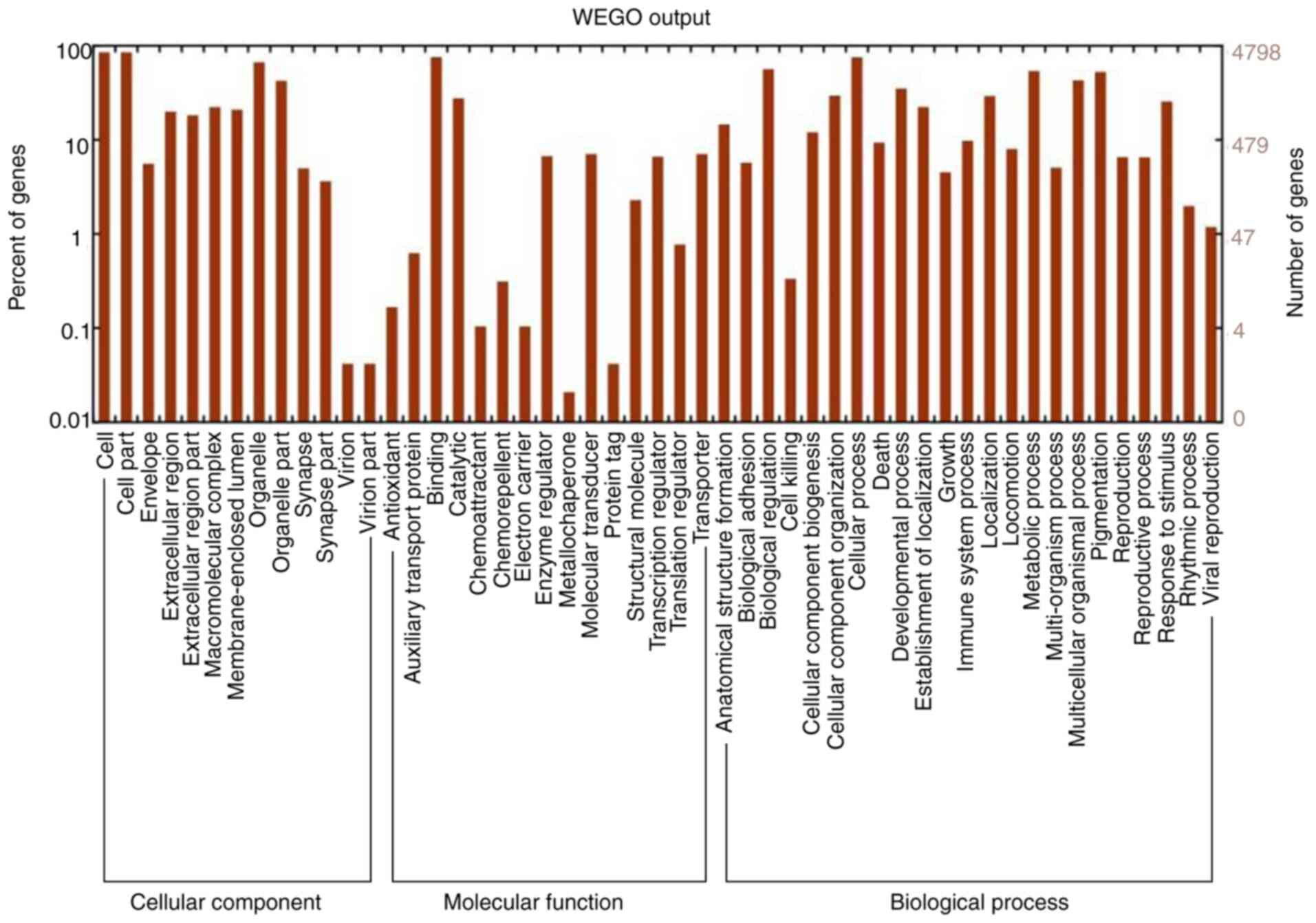
As presented in Fig.
2, the top enriched GO terms for cancer-related genes were
determined. Furthermore, KEGG enrichment analysis was performed
using the target mRNAs of the differentially expressed miRNAs. The
functional terms and numbers of genes in the categories cellular
component, molecular function and biological process are listed in
Fig. 2. In the category cellular
component, genes were associated with cell part, envelope and
organelle. In the category molecular function, terms associated
with binding, catalytic activity, enzyme regulation and molecular
transduction were significant. In the category biological process,
significantly enriched terms were associated with biological
regulation, cellular process, developmental process, death,
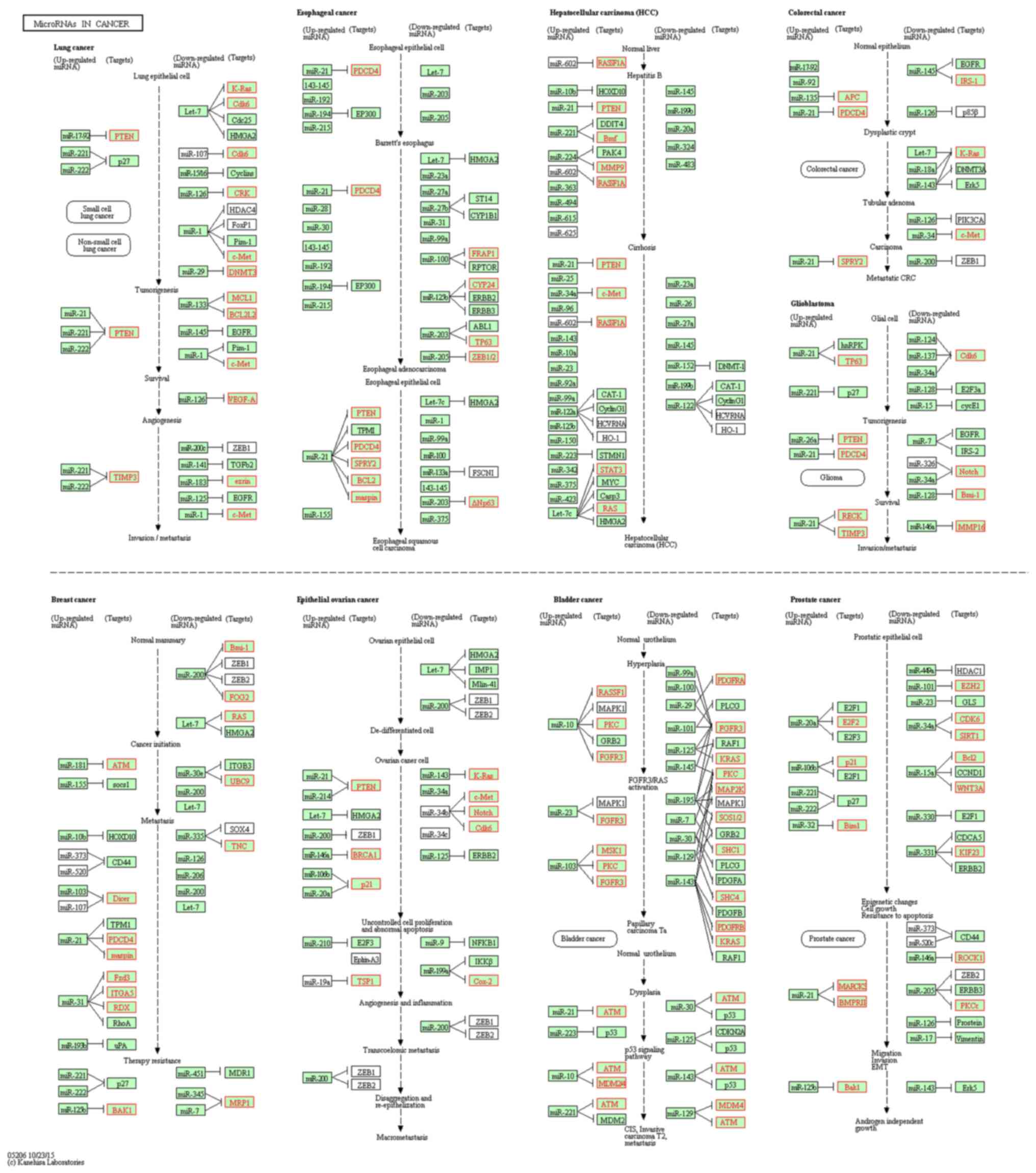
metabolic process and pigmentation. In Fig. 3, KEGG pathways associated with
cancer signaling are presented. Various miRNAs, including miR-145,
miR-31, miR-21, miR-107, miR-125, miR-135 and miR-96, were
analyzed. Compared with the control group, the genes PDCD4, PKC,
ITGA5 and ATM were upregulated, while genes such as CYP24, Fzd3,
p21 and RDX were downregulated in the infected group (Table SIII). Certain genes did not change
(e.g. ERBB2, RAF1 and ERBB3) in the infected group.
Prediction of cancer-related genes
targeted by differentially expressed miRNAs
Through the degradation of target gene mRNAs or the
inhibition of the translation of target transcripts, miRNAs have
important roles in cell proliferation and differentiation,
apoptosis and a variety of diseases. A total of 8,116 target genes
were analyzed for 45 differentially expressed cancer-related genes
using miRDB (Table SIV). On the
one hand, miRNAs are able to target a variety of cancer-related
genes. For instance, miR-211-5p targets Tnfaip8, C1qtnf3, Lats2,
Lats1 and St7; miR-145a-5p targets Tnfrsf11b, Etaa1 and C1qtnf9;
and miR-429-3p targets Lats2, Lrp1b, Mtus1 and Tnfrsf11b. On the
other hand, a single cancer-related gene may be targeted by
multiple miRNAs. For instance, the genes Tnfaip8, C1qtnf3, Lats2,
Lats1 and St7 may all be targeted by both miR-211-5p and
miR-204-5p. Furthermore, miR-211-5p, miR-31-5p, miR-135a-5p,
miR-204-5p and miR-429-3p are all able to target the Lats2 gene.
The Lats1 gene is able to be targeted by four miRNAs (miR-201-5p,
miR-211-5p, miR-204-5p and miR-107-3p), whereas Lzts3 is only
targeted by miR-138-5p.
RT-qPCR confirmation of differentially
expressed miRNAs
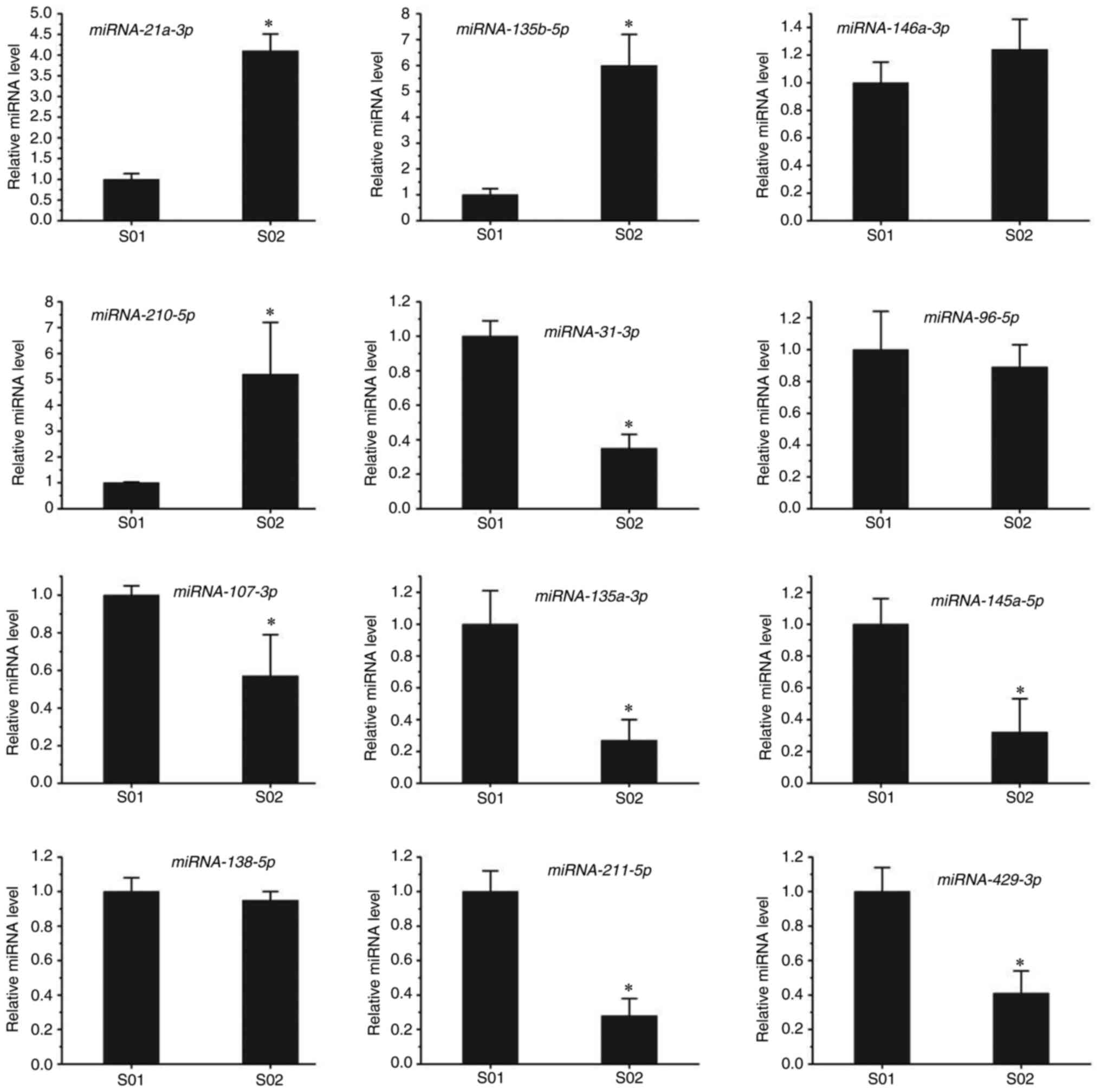
To further confirm the miRNA-seq results, RT-qPCR
was performed in the present study. Candidate miRNAs were selected
from cancer metabolic signaling pathways. Compared with the control
group, miR-210-5p, miR-135b-5p, miR-21a-3p and miR-146a-3p were
upregulated, while miR-135a-5p, miR-125a-5p, miR-145a-5p,
miR-107-3p, miR-211-5p, miR-429-3p, miR-96-5p and miR-31-3p were
downregulated in the infected group (Fig. 4). The RT-qPCR results suggested
that the levels of miR-210-5p, miR-135b-5p and miR-21a-3p were
significantly upregulated after T. gondii infection, whereas
no difference in miR-146a-3p levels was observed in the infected
group. The levels of miR-210-5p and miR-135b-5p in the infected
group were five times higher than those in the control group. In
addition, the levels of miR-135a-5p, miR-125a-5p, miR-145a-5p and
miR-31-3p were markedly downregulated after infection with T.
gondii, while no difference in the levels of miR-96-5p or
miR-138-5p was observed between the infected and control groups.
The levels of miR-135a-5p and miR-145a-5p in the infected group
were only one-quarter of those in the control group. These results
were in line with those of the RNA-seq.
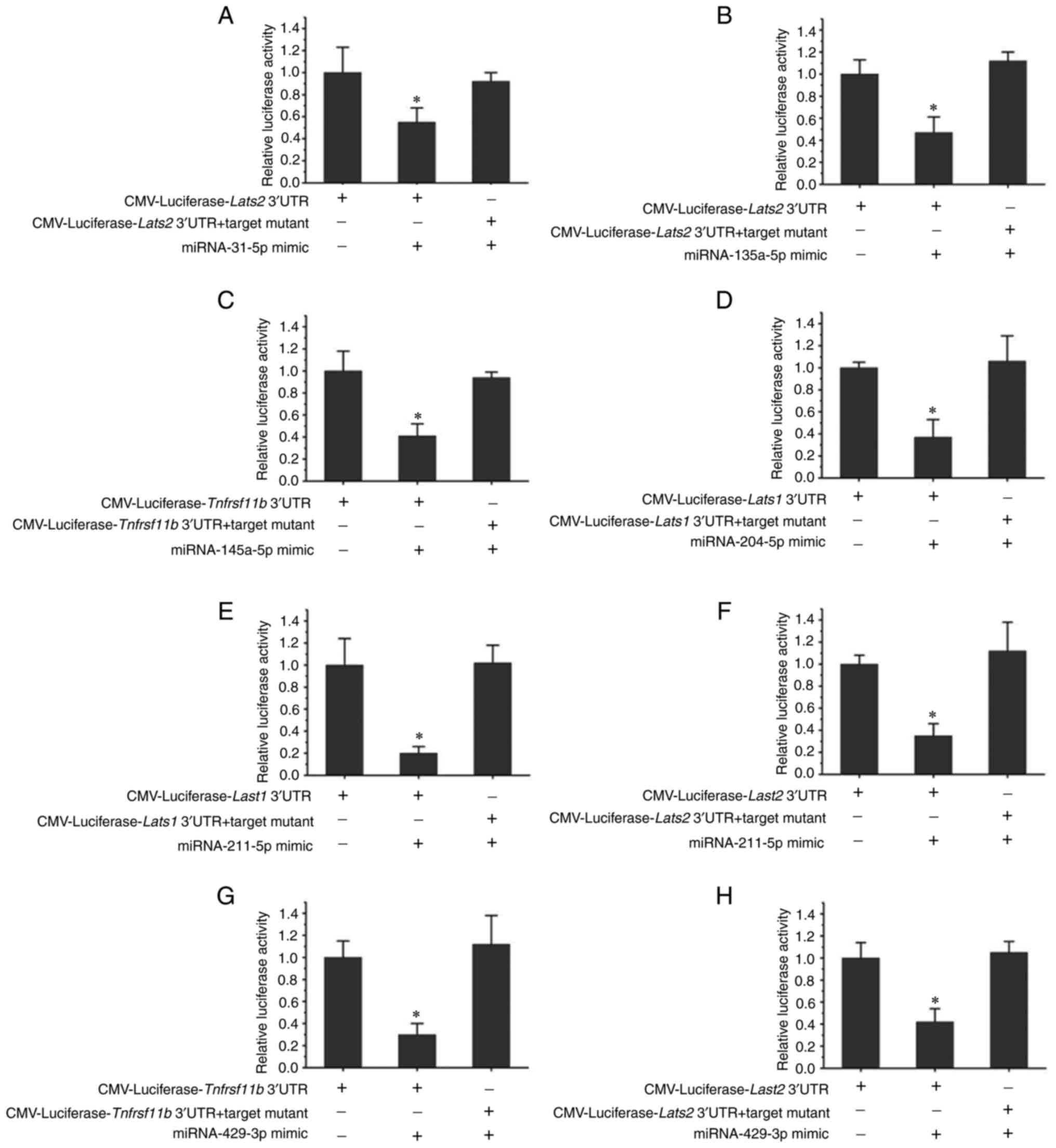
Luciferase activity assay
Targets of miR-31-5p, miR-135a-5p, miR-145a-5p,
miR-204-5p, miR-211-5p and miR-429-3p were verified by luciferase
activity assays in 293T cells (Fig.
5). While miR-31-5p, miR-135a-5p, miR-211-5p and miR-429-3p
were confirmed to be able to target the Lats2 gene, miR-204-5p and
miR-211-5p target the Lats1 gene. In addition, miR-145a-5p and
miR-429-3p were indicated to be targets of Tnfrsf11b. Furthermore,
miR-211-5p was able to target the Lats1 and Lats2 genes, and
miR-429-3p was confirmed to target the Tnfrsf11b and Lats2
genes.
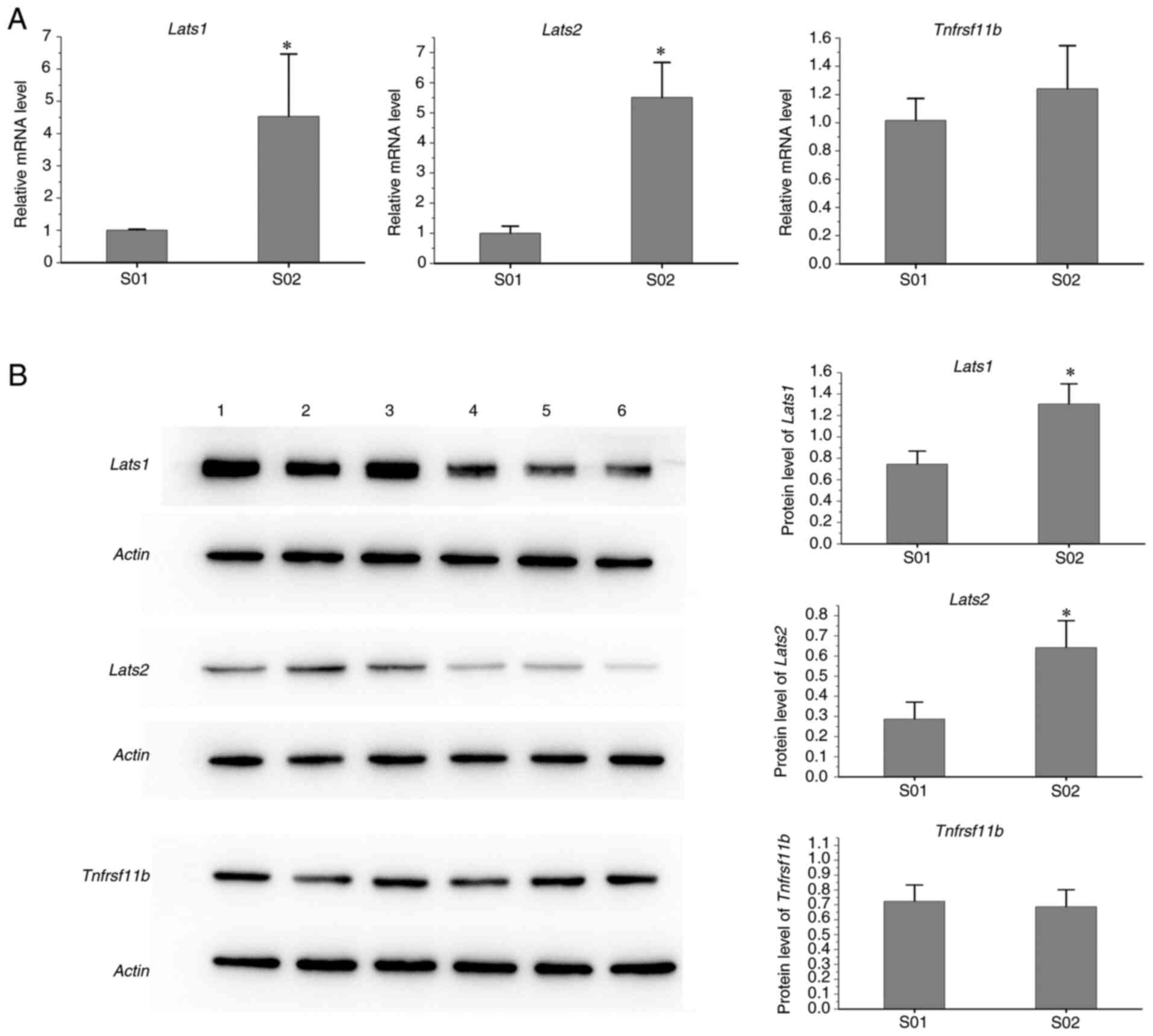
Protein changes
As presented in Fig.
6, the protein levels of Lats1 and Lats2 in spleen samples of
infected mice were higher than those in the control group, while
the levels of Tnfrsf11b protein from the infected group were
similar to those of the control mice.
Discussion
T. gondii is most likely to manipulate
pathways involved in host innate immunity, biosynthesis and
transferase activity (7).
Furthermore, the levels of miRNAs in host cells were previously
reported to be acutely changed after T. gondii infection;
compared with uninfected DC2.4 cells, 3,434 differentially
expressed miRNAs were obtained from T. gondii-infected DC2.4
cells after high-throughput sequencing (19). Host miRNA expression is altered by
T. gondii, which is reflected in the differences in
regulation of important biological processes that are related to
host responses to both chronic and acute T. gondii infection
(6). miRNAs of porcine macrophages
were reported to be differentially expressed after T. gondii
infection (7); this knowledge may
aid in the investigation of T. gondii infections. Hu et
al (8) indicated that T.
gondii infection, particularly at the chronic stage, may change
the abundance of miRNAs in the mouse brain. Another previous study
suggested that miRNAs may be related to the communication between
hosts and T. gondii and several specific miRNAs have been
proposed and confirmed (20).
miRNA expression in the brain of mice may change with the invasion
of cyst-forming T. gondii (21). Chronic and acute infections may
cause differential expression of miRNAs in the host. In the present
study, compared with the control group, 207 miRNAs were upregulated
and 414 miRNAs were downregulated in the T. gondii-infected
group. The fact that certain miRNAs were differentially expressed
upon T. gondii infection was consistent with previous
studies.
Infection with T. gondii may stimulate
immunity against tumors (11,13,22).
Furthermore, the levels of tumor-related mRNAs were altered in
T. gondii-infected mice from a previous study by our group
(14). In the present study, the
levels of numerous miRNAs were changed in infected mice compared to
the control group. Specifically, the levels of cancer-related
miRNAs, such as miR-135a-5p, miR-135b-5p, miR-145a-5p, miR-146a-3p,
miR-21a-3p, miR-107-3p, miR-31-3p, miR-96-5p, miR-210-5p,
miR-211-5p and miR-429-3p, were altered in the infected mice.
miRNAs have important roles in cancer. miR-135a-5p, a significant
tumor regulator, may affect the development of diverse cancers by
impacting multiple genes in oncogenic pathways (23–25).
miR-135b-5p was able to enhance doxorubicin sensitivity in breast
cancer cells by targeting anterior gradient 2 protein (26). miR-96-5p was reported to promote
the migration and proliferation of ovarian cancer cells by
suppressing caveolae 1 (27). In
the present study, the RT-qPCR results indicated that the levels of
miR-210-5p, miR-135b-5p and miR-21a-3p were significantly
upregulated after infection with T. gondii, while the levels
of miR-135a-5p, miR-125a-5p, miR-145a-5p, miR-211-5p, miR-429-3p
and miR-31-3p were markedly downregulated after T. gondii
challenge. miR-135a-5p was previously reported to inhibit head and
neck squamous cell carcinoma (HNSCC) cell proliferation and promote
apoptosis by directly targeting HOXA10, suggesting the importance
of miR-135a-5p in HNSCC treatment (28). miR-125a-5p was downregulated in
colorectal cancer tissues and cell lines and inhibited colorectal
cancer cell proliferation, migration and invasion; it also reduced
the ability of human umbilical vein endothelial cells to form tubes
(29). Overexpression of miR-31-3p
inhibited malignant behaviors and epithelial-to-mesenchymal
transition of cervical cancer cells in vitro (30).
In the present study, cancer-related genes (Lats2,
Lats1 and Tnfrsf11b) were targeted by miR-429-3p, miR-211-5p,
miR-145a-5p, miR-31-3p, miR-204-5p and miR-135a-5p. In a previous
study, the expression of both Lats2 and Lats1 was significantly
downregulated in human breast cancer and loss of either one
accelerated mammary tumorigenesis in mice (31). Lats2 was able to inhibit oncogenic
Wnt/β-catenin-mediated transcription by disrupting β-catenin/BCL9
interactions, suggesting that it may be an important target for
anti-cancer therapies (32).
Nucleocytoplasmic translocation of Lats1 protein and upregulated
expression of Lats1 mRNA were observed during tumorigenesis of
HNSCC (33). In addition,
cancer-related miRNAs were altered in mice after T. gondii
infection. The altered miRNAs affected cancer-related genes,
suggesting that the challenge with T. gondii may alter the
host's resistance to tumors. In the present study, the levels of
miR-429-3p, miR-211-5p, miR-145a-5p, miR-31-3p and miR-135a-5p were
downregulated after infection, indicating that a smaller number of
antitumor genes (Lats2, Lats1 and Tnfrsf11b) may be targeted in
infected mice compared with others. Furthermore, the protein levels
of Lats2 and Lats1 were markedly increased in infected mice
compared with the control group, suggesting that the altered miRNAs
may enhance the expression of antitumor genes. Consequently, T.
gondii infection may increase host resistance to tumors, which
is consistent with the results of the previous study by our group
(14). Furthermore, previous
studies suggested that infection with T. gondii may improve
immunity against cancer in the host (9–12).
High expression of antitumor genes may have an important role in
immunity.
Numerous miRNAs have been indicated to be altered in
mice infected with T. gondii. Cancer-related miRNAs
(miR-429-3p, miR-211-5p, miR-145a-5p, miR-31-3p and miR-135a-5p)
were downregulated, suggesting that a smaller amount of antitumor
genes were targeted and more antitumor proteins were expressed. In
the present study, the findings were favorable for the suppression
of cancer and may provide novel ideas for the treatment of tumors.
The current study aimed to explore the tumor factors affected by
T. gondii. Although certain cancer-related proteins were
altered in mice after T. gondii infection, the exact
implication and type of cancer this may affect remain elusive and
require further exploration. In further studies, a variety of mouse
tumor models may be used as experimental models. Attenuated T.
gondii may be inoculated prior to and after tumorigenesis. The
tumor growth of mice may be observed and the changes of tumor
factors detected. The tumor model with the greatest impact may be
obtained by comparison. The upstream and downstream molecules of
the tumor factor may be detected to explore the possible pathways
through which T. gondii affects the growth of tumors in the
host.
In conclusion, cancer-associated miRNAs were altered
in mice after T. gondii infection. Antitumor genes
(Tnfrsf11b, Lats2 and Lats1) were changed accordingly. In addition,
a larger amount of Lats2 and Lats1 protein expression was detected
in T. gondii-infected mice than in the control group,
indicating that T. gondii may enhance host immunity against
cancer by enhancing the abundance of antitumor proteins. These
findings are favorable for the suppression of cancer and may
provide novel ideas for the treatment of tumors.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the National Foundation of Natural
Science of China (grant no. 81902079) and the Science Foundation of
Shandong Province (grant no. ZR2013HM033).
Availability of data and materials
The sequencing data generated in the present study
may be obtained from the Sequence Read Archive (SRA) of the
National Center for Biotechnology Information (NCBI; https://www.ncbi.nlm.nih.gov/sra; accession no.
PRJNA803770). The other datasets used and/or analyzed in the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
GL conceived and designed the study and critically
revised the manuscript. LW performed the experiments and drafted
the manuscript. NW and YHZ contributed to the analysis and
interpretation of data. GL and YHZ check and approve the
authenticity of the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Animal Care and Use Committee of Shandong University (no.
2011–0015). The animals were kept and the experiments were
performed in accordance with the committee's criteria for the care
and use of laboratory animals. This was also in accordance with the
NIH's Guide for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kochanowsky JA and Koshy AA: Toxoplasma
gondii. Curr Biol. 28:R770–R771. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Coutermarsh-Ott S: Toxoplasma
gondii as a model of in vivo host-parasite interactions.
Methods Mol Biol. 1960:237–247. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lima TS and Lodoen MB: Mechanisms of human
innate immune evasion by Toxoplasma gondii. Front Cell
Infect Microbiol. 9:1032019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Menard KL, Haskins BE and Denkers EY:
Impact of Toxoplasma gondii infection on host non-coding RNA
responses. Front Cell Infect Microbiol. 9:1322019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Medina L, Castillo C, Liempi A,
Guerrero-Muñoz J, Rojas-Pirela M, Maya JD, Prieto H and Kemmerling
U: Trypanosoma cruzi and Toxoplasma gondii induce a
differential MicroRNA profile in human placental explants. Front
Immunol. 11:5952502020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hou Z, Liu D, Su S, Wang L, Zhao Z, Ma Y,
Li Q, Jia C, Xu J, Zhou Y and Tao J: Comparison of splenocyte
microRNA expression profiles of pigs during acute and chronic
toxoplasmosis. BMC Genomics. 20:972019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li SY, Yang J, Wang LY, Du F, Zhao JL and
Fang R: Expression profile of microRNAs in porcine alveolar
macrophages after Toxoplasma gondii infection. Parasit
Vectors. 12:652019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu RS, He JJ, Elsheikha HM, Zhang FK, Zou
Y, Zhao GH, Cong W and Zhu XQ: Differential brain MicroRNA
expression profiles after acute and chronic infection of mice with
Toxoplasma gondii oocysts. Front Microbiol. 9:23162018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sanders KL, Fox BA and Bzik DJ: Attenuated
Toxoplasma gondii therapy of disseminated pancreatic cancer
generates long-lasting immunity to pancreatic cancer.
Oncoimmunology. 5:e11044472015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sanders KL, Fox BA and Bzik DJ: Attenuated
Toxoplasma gondii stimulates immunity to pancreatic cancer
by manipulation of myeloid cell populations. Cancer Immunol Res.
3:891–901. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Y, Poppoe F, Chen J, Yu L, Deng F, Luo
Q, Xu Y, Cai Y and Shen J: Macrophages polarized by expression of
ToxoGRA15II inhibit growth of hepatic carcinoma. Front
Immunol. 8:1372017.PubMed/NCBI
|
|
12
|
Pyo KH, Lee YW, Lim SM and Shin EH: Immune
adjuvant effect of a Toxoplasma gondii profilin-like protein
in autologous whole-tumor-cell vaccination in mice. Oncotarget.
7:74107–74119. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fox BA, Sanders KL, Rommereim LM, Guevara
RB and Bzik DJ: Secretion of rhoptry and dense granule effector
proteins by nonreplicating Toxoplasma gondii uracil
auxotrophs controls the development of antitumor immunity. PLoS
Genet. 12:e10061892016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu G, Zhou J, Zhao YH, Li QL, Gao YY and
Wang L: Transcriptome sequencing investigated the tumor-related
factors changes after T. gondii infection. Front Microbiol.
10:1812019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang T, Pan H, Wang J, Yang W, Cheng T and
Zhang Q: Identification and profiling of novel and conserved
microRNAs during the flower opening process in Prunus mume via deep
sequencing. Mol Genet Genomics. 289:169–183. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen Y and Wang X: miRDB: An online
database for prediction of functional microRNA targets. Nucleic
Acids Res. 48:D127–D131. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pjevac P, Hausmann B, Schwarz J, Kohl G,
Herbold CW, Loy A and Berry D: An economical and flexible dual
barcoding, two-step PCR approach for highly multiplexed amplicon
sequencing. Front Microbiol. 12:6697762021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li DL, Zou WH, Deng SQ and Peng HJ:
Analysis of the differential exosomal miRNAs of DC2.4 dendritic
cells induced by Toxoplasma gondii infection. Int J Mol Sci.
20:55062019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Acar İE, Saçar Demirci MD, Groß U and
Allmer J: The expressed MicroRNA-mRNA interactions of Toxoplasma
gondii. Front Microbiol. 8:26302018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou CX, Ai K, Huang CQ, Guo JJ, Cong H,
He SY and Zhu XQ: miRNA and circRNA expression patterns in mouse
brain during toxoplasmosis development. BMC Genomics. 21:462020.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hatai H, Lepelley A, Zeng W, Hayden MS and
Ghosh S: Toll-like receptor 11 (TLR11) interacts with flagellin and
profilin through disparate mechanisms. PLoS One. 11:e01489872016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Jiang WL, Yang JY, Huang J, Kang
G, Hu HB and Xie S: Downregulation of lysyl oxidase-like 4 LOXL4 by
miR-135a-5p promotes lung cancer progression in vitro and in vivo.
J Cell Physiol. 234:18679–18687. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zheng Y, Zheng B, Meng X, Yan Y, He J and
Liu Y: LncRNA DANCR promotes the proliferation, migration, and
invasion of tongue squamous cell carcinoma cells through
miR-135a-5p/KLF8 axis. Cancer Cell Int. 19:3022019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei X, Yang X, Wang B, Yang Y, Fang Z, Yi
C, Shi L and Song D: LncRNA MBNL1-AS1 represses cell proliferation
and enhances cell apoptosis via targeting miR-135a-5p/PHLPP2/FOXO1
axis in bladder cancer. Cancer Med. 9:724–736. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang Y, Xia F, Zhang F, Cui Y, Wang Q,
Liu H and Wu Y: miR-135b-5p enhances doxorubicin-sensitivity of
breast cancer cells through targeting anterior gradient 2. J Exp
Clin Cancer Res. 38:262019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu B, Zhang J and Yang D: miR-96-5p
promotes the proliferation and migration of ovarian cancer cells by
suppressing caveolae1. J Ovarian Res. 12:572019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo LM, Ding GF, Xu WC, Ge H, Jiang Y,
Chen XJ and Lu YF: MiR-135a-5p represses proliferation of HNSCC by
targeting HOXA10. Cancer Biol Ther. 19:973–983. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang X, Qiu J, Kang H, Wang Y and Qian J:
miR-125a-5p suppresses colorectal cancer progression by targeting
VEGFA. Cancer Manag Res. 10:5839–5853. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jing L, Bo W, Yourong F, Tian W, Shixuan W
and Mingfu W: Sema4C mediates EMT inducing chemotherapeutic
resistance of miR-31-3p in cervical cancer cells. Sci Rep.
9:177272019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Furth N, Pateras IS, Rotkopf R, Vlachou V,
Rivkin I, Schmitt I, Bakaev D, Gershoni A, Ainbinder E, Leshkowitz
D, et al: LATS1 and LATS2 suppress breast cancer progression by
maintaining cell identity and metabolic state. Life Sci Alliance.
1:e2018001712018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Chen X, Ding X, Cheng Y, Zhao B, Lai
ZC, Al Hezaimi K, Hakem R, Guan KL and Wang CY: LATS2 suppresses
oncogenic Wnt signaling by disrupting β-catenin/BCL9 interaction.
Cell Rep. 31:1077922020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu J, Zhao Z, Zhang H, Kong F, Jiang H,
Huang K and Zheng H: LATS1 inhibits metastasis and
epithelial-mesenchymal transition in head and neck squamous cell
carcinoma. Int J Clin Exp Pathol. 11:2053–2063. 2018.PubMed/NCBI
|