Introduction
Lung cancer is the leading cause of
cancer-associated death among men and the second leading cause
among women worldwide (1).
Non-small cell lung cancer (NSCLC), including adenocarcinoma,
squamous cell carcinoma and large cell carcinoma, represents ~85%
of all lung cancer cases, with an overall 5-year survival rate of
<15% (2,3). Most patients with NSCLC are diagnosed
with advanced stage tumors owing to the inadequate screening
methods and the late onset of clinical symptoms; consequently, most
patients have a poor prognosis (4). Therefore, new screening regimens and
technologies, as well as new tumor markers to maximize the
detection of early NSCLC, are key goals for solving the high
mortality rate of NSCLC.
Tumorigenesis and progression are highly related to
gene regulation. Recently, some studies have found that microRNA
(miRNA/miR) levels can be used for the early diagnosis of NSCLC
(5). miRNA is a family of
endogenous, single-stranded, non-coding small RNA molecules, which
are ~20-24 nucleotides (6). They
bind to the 3′-untranslated region (3′-UTR) of the target mRNA and
participate in the fine-tuning of various biological processes as a
key regulator of gene expression (6,7). An
increasing number of studies have showed that miRNAs serve a vital
role in the progression of NSCLC (6,7). A
previous study reported that hsa-miR-338 may have a tumor
inhibitory effect in the progression of NSCLC (8). miR-126 targeting PI3K inhibits cell
proliferation, migration and invasion in the A549 cell line by
regulating the PTEN/PI3K/AKT pathway (9). Nevertheless, the mechanism of miRNA
in NSCLC is still unclear.
The inhibitors of programmed cell death protein 1
(PD-1) and its ligand programmed death ligand-1 (PD-L1) have been
the focus of tumor immunotherapy in recent years (10). The protein expression level of
PD-L1 is upregulated in numerous types of tumor cells (11). PD-L1 binds to PD-1 on T cells and
subsequently inhibits the proliferation, and activation of these
cells. The T cell response is a prominent part of the antitumor
immune response via the direct killing of target tumor cells or via
indirect inhibition by cytokines (12). PD-1/PD-L1 immune checkpoints play
an important role in immune regulation by delivering inhibitory
signals to maintain the balance in T cell activation, tolerance and
immune-mediated tissue damage (13).
In the present study, it was shown that the
expression level of miR-20a in NSCLC cell lines was upregulated.
miR-20a promoted the proliferation of human NSCLC cells by directly
targeting PTEN to promote the expression level of PD-L1 and
activating the Wnt/β-cantenin pathway. These results demonstrated
that miR-20a may be a potential therapeutic target in NSCLC.
Materials and methods
Cell culture, transfection and
proliferation assay
The cell lines (BEAS-2B, A549 and H1299) were
purchased from American Type Culture Collection. The cells were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml streptomycin
at 37°C, and 5% CO2. The cells were detected for
mycoplasma contamination using routine PCR.
The miR-20a mimics, miR-Con mimics, anti-miR-20a
mimics, anti-miR-Con mimics, PTEN-short hairpin RNA (shRNA) and
knockdown (KD)-PTEN-shRNA were obtained from Shanghai GenePharma
Co., Ltd. The shRNA sequence for the PTEN gene was as follows:
sh-PTEN:
5′-CCGGCCACAAATGAAGGGATATAAACTCGAGTTTATATCCCTTCATTTGTGGTTTTTG-3′.
Flag-PD-L1/pcDNA3.0, Flag-PTEN/pcDNA3.0 and Flag/pcDNA3.0 plasmids
were from our laboratory. Flag plasmids and miRNA mimics
(concentration of 50 nM) were transfected into A549 cells and H1299
cells using SuperFectin II (Shanghai Pufei Biotechnology Co., Ltd.)
and 24 h after transfection, the cells were processed as described
for each experiment. A549 and H1299 cells were transduced with the
lentivirus at multiplicity of infection of MOI 20 for 48 h, and
then selected with puromycin (2 mg/ml). Independent stable clones
were evaluated by Western blotting.
Cell proliferation was determined using the Cell
Counting Kit (CCK)-8 (GlpBio Technology) at 37°C for 72 h. The
absorbance was detected by enzyme labeling instrument (450 nm)
every 24 h; the XAV-939 (cat. no. HY-15147; 10 µM) was purchased
from MedChemExpress.
Reverse transcription-quantitative
(RT-qPCR)
Total RNA was isolated from A549 cells and H1299
cells using TRIzol® (Thermo Fisher Scientific, Inc.).
RT-qPCR analysis of miRNA or mRNA was performed as previously
reported (14). The following
primer sequences were used: PD-L1 forward,
5′-CCTACTGGCATTTGCTGAACGCAT-3′ and reverse,
5′-ACCATAGCTGATCATGCAGCGGTA-3′; PTEN forward,
5′-GATGAGGCATTATCCTGTACACA-3′ and reverse,
5′-CTCTTCAGATACTCTTGTGCTGT-3′ and β-actin forward,
5′-ACCATTGGCAATGAGCGGT-3′ and reverse,
5′-GTCTTTGCGGATGTCCACGT-3′.
Western blotting
Total protein extracts of A549 cells and H1299 cells
were prepared using Keygen Protein Extraction Reagent (cat. no.
KGP250; Nanjing KeyGen Biotech Co., Ltd.) according to the
manufacturer's instructions. The proteins were quantified by BCA
Protein Assay kit (Beyotime Institute of Biotechnology) and the
mass of protein loaded per lane was 25 µg. The protein was
fractionated using 10% SDS-PAGE for 2 h at 110 V and transferred
onto the PVDF membranes. Membranes were blocked in 5% skim milk
powder diluted with Tri-buffered saline Tween-20 (20 mM Tris-HCl,
150 mM NaCl and 0.1% Tween-20) at room temperature for 1 h, and
immunostained with the following primary antibodies at 4°C
overnight: PD-L1 (1:1,000; cat. no. 28076-1-AP), PTEN (1:5,000;
cat. no. 22034-1-AP), β-catenin (1:1,000; cat. no. 51067-2-AP),
Cyclin D1 (1:1,000; cat. no. 26939-1-AP), β-actin (1:2,000; cat.
no. 20536-1-AP) and GAPDH (1:10,000; cat. no. 10494-1-AP), and were
all purchased from ProteinTech Group, Inc. GAPDH and β-actin were
analyzed to show equal protein loading. The secondary antibodies
used were goat-rabbit IgG (1:10,000; cat. no. sc-2004) and goat
anti-mouse IgG (1:10,000; cat. no. sc-2005) (both purchased from
Santa Cruz Biotechnology, Inc.). The blots were detected with an
enhanced chemiluminescence detection kit (Thermo Fisher Scientific,
Inc.) and exposed in a Molecular Imager® ChemiDoc XRS
system (Bio-Rad Laboratories, Inc.).
Bioinformatics prediction
TargetScan 7.1 software (http://www.targetscan.org/vert_71/) was used to
predict the potential target genes of microRNA-20a. The search
terms ‘Human’ and ‘microRNA-20a’ were used.
Luciferase activity assay
The interaction between miR-20a and PD-L1 was
determined using a luciferase activity assay. The PD-L1 3′-UTR
containing miR-20a binding site was subcloned into the luciferase
reporter plasmid vector (Promega Corporation). The 3′-UTR
luciferase vector was co-transfected with miR-20a mimics or mimics
control (miR-Con) or anti-miR-20a mimics into the A549 or H1299
cells using SuperFectin™ II (Shanghai Pufei
Biotechnology Co., Ltd.) and Renilla luciferase reporters
were used as an internal control. A luciferase activity assay was
performed after 48 h using the Dual-Luciferase Reporter Assay
System (Promega Corporation) according to the manufacturer's
protocol.
Statistical analysis
All statistical analyses were performed using SPSS
v22.0 (IBM Corp.) statistical package for windows. All results were
expressed as the mean ± SD and the experiments were performed at
least 3 times. The statistical differences between categorical data
were evaluated using a Fisher's exact test. Comparisons between
continuous variables were analyzed using non-parametric
Mann-Whitney U test (2 groups) or one-way Analysis of Variance test
(>2 groups). All tests were two-sided and P≤0.05 was considered
to indicate a statistically significant difference.
Results
PD-L1 is expressed in NSCLC cell lines
and promotes NSCLC cell proliferation
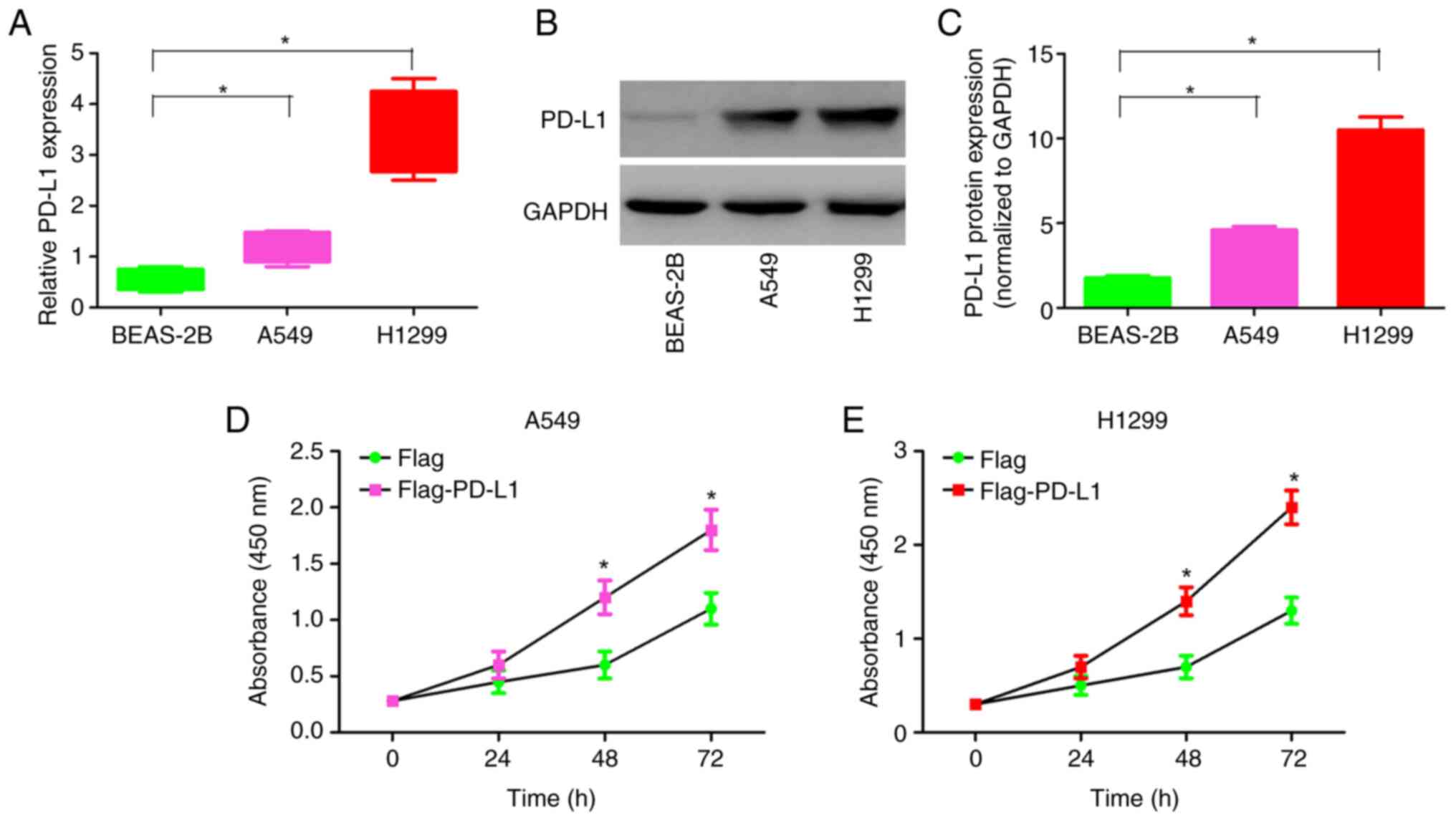
In order to gain insight into the function of PD-L1,
RT-qPCR was performed to verify the transcriptional level of PD-L1
in the NSCLC cell lines (A549 and H1299). Compared with that in the
human normal lung epithelial cell line (BEAS-2B), the mRNA
expression levels of PD-L1 in the A549 and H1299 cell lines were
upregulated (Fig. 1A). Consistent
with the mRNA expression levels, the PD-L1 protein expression
levels in the A549 and H1299 cell lines were significantly higher
compared with that in the BEAS-2B cell line (Fig. 1B and C).
To examine the contribution of endogenous PD-L1 in
NSCLC cell proliferation, overexpression of PD-L1 was performed in
the A549 and H1299 cell line (Fig.
S1). Overexpression of PD-L1 enhanced the proliferation of the
A549 and H1299 cell lines (Fig. 1D and
E). These results indicate that PD-L1 may play the role of
oncoprotein in NSCLC and induce the proliferation of NSCLC
cells.
miR-20a regulates PD-L1
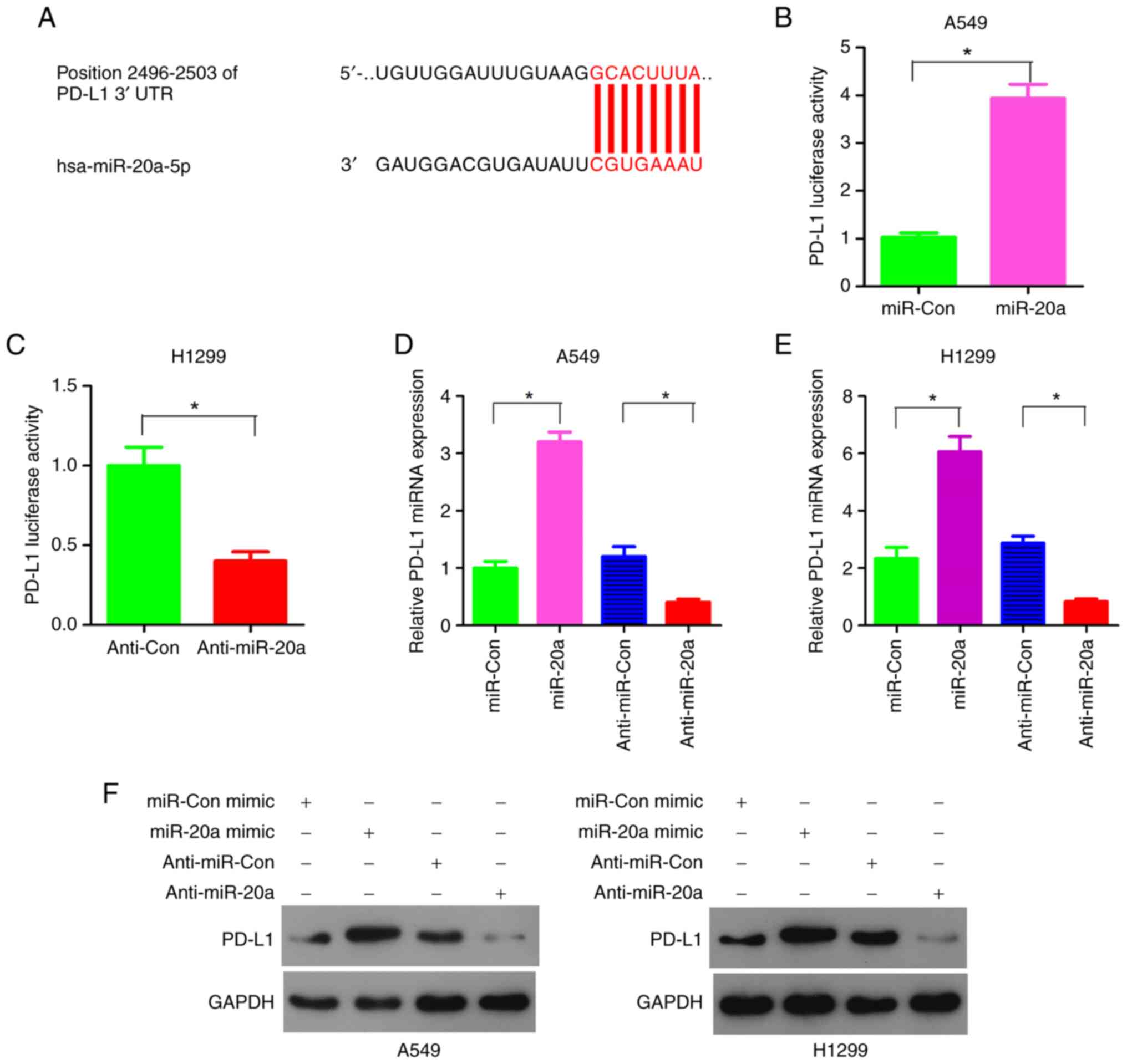
miRNAs regulate gene expression after transcription
by binding to the 3′-UTR of mRNAs. miRNAs play an important role in
the immune response (6,7). Bioinformatics analysis showed that
PD-L1 was a potential target gene of miR-20a (Fig. 2A). Luciferase reporter plasmid,
with PD-L1 3′-UTR and miR-20a mimics or inhibitor were
co-transfected into the A549 and H1299 cell lines. The results
confirmed that miR-20a mimics significantly enhanced the activity
of PD-L1 in the A549 cell line (Fig.
2B). Consistent with the results from the A549 cells, the
luciferase activity of the PD-L1 reporter gene was inhibited by
anti-miR-20a inhibitors in the H1299 cell line (Fig. 2C).
In order to verify that the PD-L1 expression levels
were regulated by miR-20a, the NSCLC cell lines were transiently
transfected with miR-20a mimics or anti-miR-20a inhibitors. The
RT-qPCR results showed that miR-20a mimics increased the mRNA
expression levels of PD-L1 in the A549 and H1299 cell lines
(Fig. 2D and E). At the same time,
anti-miR-20a inhibitors inhibited the mRNA expression levels of
PD-L1 in A549 and H1299 cells (Fig. 2D
and E). Consistent with the results at the transcriptional
level, miR-20a promoted the expression level of PD-L1 protein in
both the A549 and H1299 cell lines. When miR-20a inhibitors were
used, the PD-L1 protein expression levels were downregulated in
both the A549 and H1299 cells (Fig.
2F). Taken together, these results indicate that miR-20a may
regulate the expression level of PD-L1 by binding to the 3′-UTR of
PD-L1.
miR-20a enhances NSCLC cell
proliferation
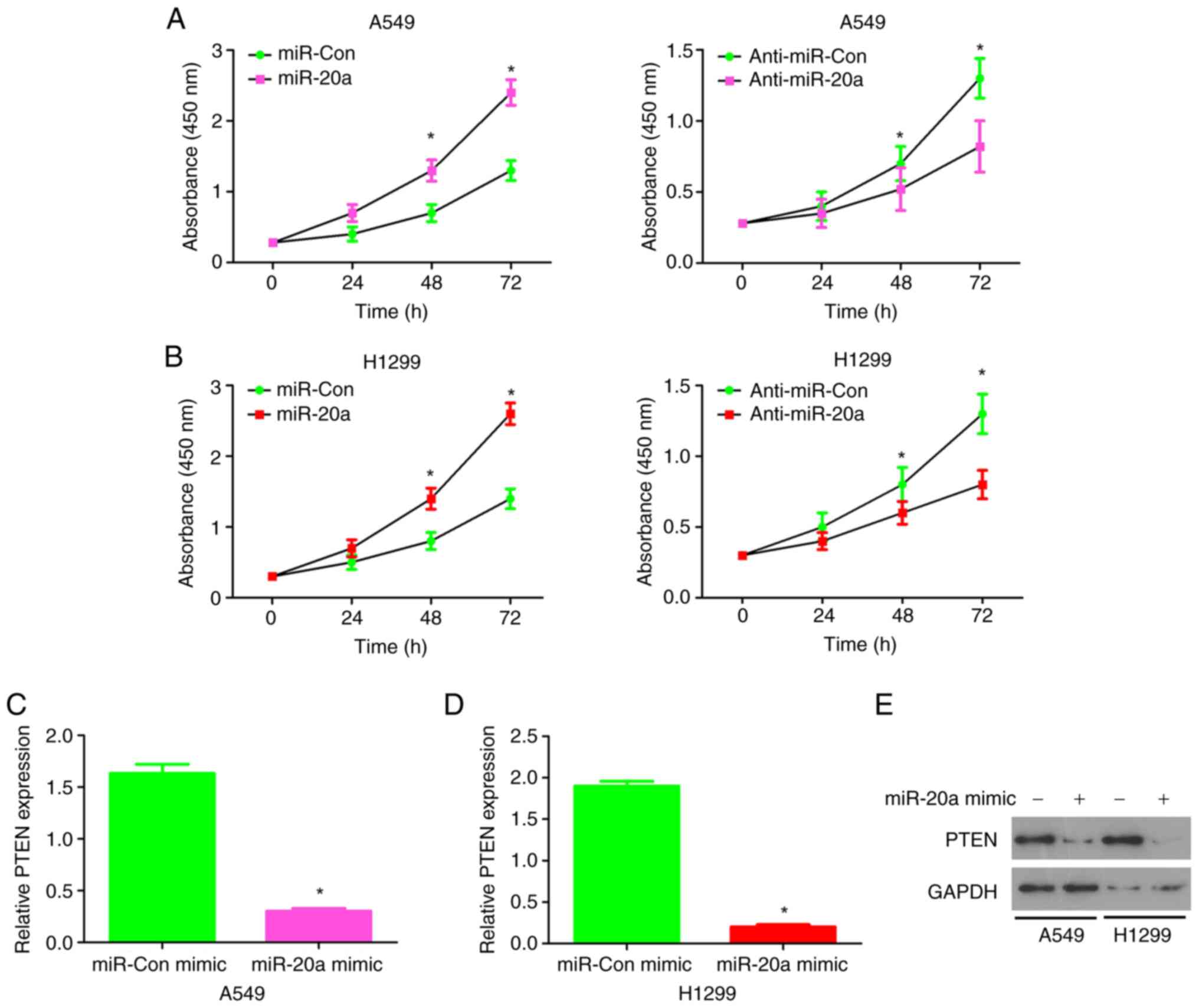
To investigate the biological role of miR-20a in
NSCLC cells, a CCK-8 assay was performed and the effect of
miR-20aon the proliferation of NSCLC cells was evaluated. miR-20a
mimic, miR-Con mimic, anti-miR-20a mimic and anti-miR-Con mimic
were transfected into A549 cells (Fig. S2) and H1299 cells (Fig. S3), respectively, and the
expression level of miR-20a was detected. miR-20a mimics increased
cell proliferation, while miR-20a inhibitor decreased cell
proliferation in the A549 cell line (Fig. 3A). Similar results were found in
the H1299 cell line (Fig. 3B). In
summary, these results suggest that miR-20a enhanced NSCLC cell
proliferation.
miR-20a inhibits the transcription and
protein expression level of PTEN in NSCLC cells
When miR-20a mimics were highly expressed in A549
cells and H1299 cells, the expressions of PTEN were downregulated
(Fig. 3C and D). At the protein
expression level, miR-20a mimics significantly inhibited PTEN in
both the A549 and H1299 cell lines (Fig. 3E). These results suggest that
miR-20a may directly target the tumor suppressor, PTEN and inhibit
PTEN transcription and protein expression levels in the A549 and
H1299 cell lines.
miR-20a enhances PD-L1 expression by
repressing PTEN
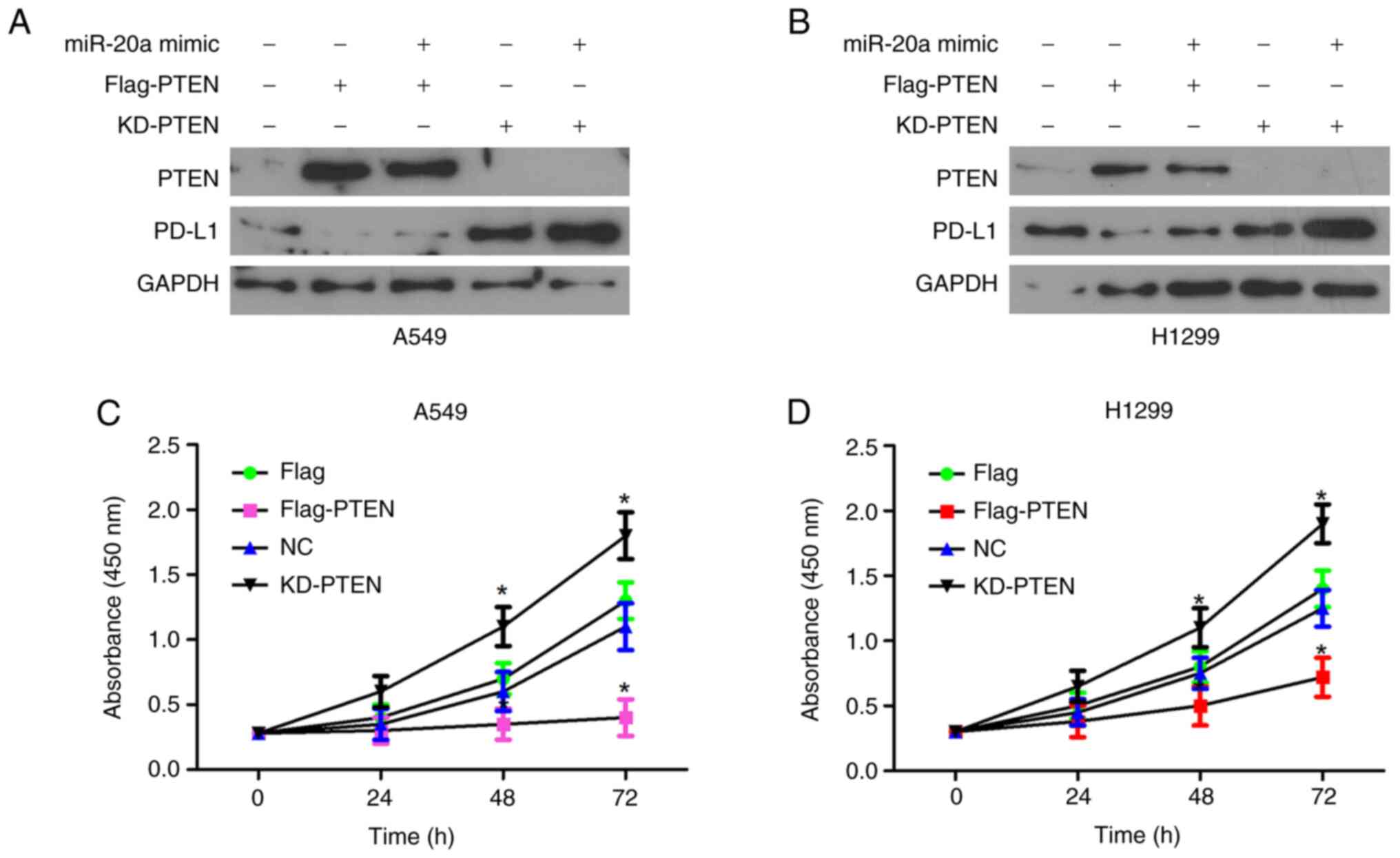
PD-L1 protein expression level was decreased in the
PTEN overexpressing A549 cell line. In addition, in cells
transfected with miR-20a mimics and PTEN overexpression vector, the
protein expression level of PD-L1 was lower compared with that in
the control cells (Figs. 4A and B,
and S4). Furthermore, the
expression level of PD-L1 was increased following knockdown of PTEN
expression, while the protein expression level of PD-L1was further
increased following transfection with miR-20a mimics and knockdown
of PD-L1 (Figs. 4A and B, and
S4) The proliferation rate of the
A549 cell line, transfected with PTEN overexpression vector was the
slower compared with that in control A549 cell line, while
knockdown of PTEN in the A549 cell line was fastest compared with
that in the other groups (Fig.
4C). Similar results were observed with the H1299 cell line
(Fig. 4D). These findings
indicated that PTEN may be an inhibitor of PD-L1, by affecting the
proliferation of NSCLC cells.
miR-20a promotes NSCLC cell
proliferation by targeting PTEN to activate the Wnt/β-catenin
pathway
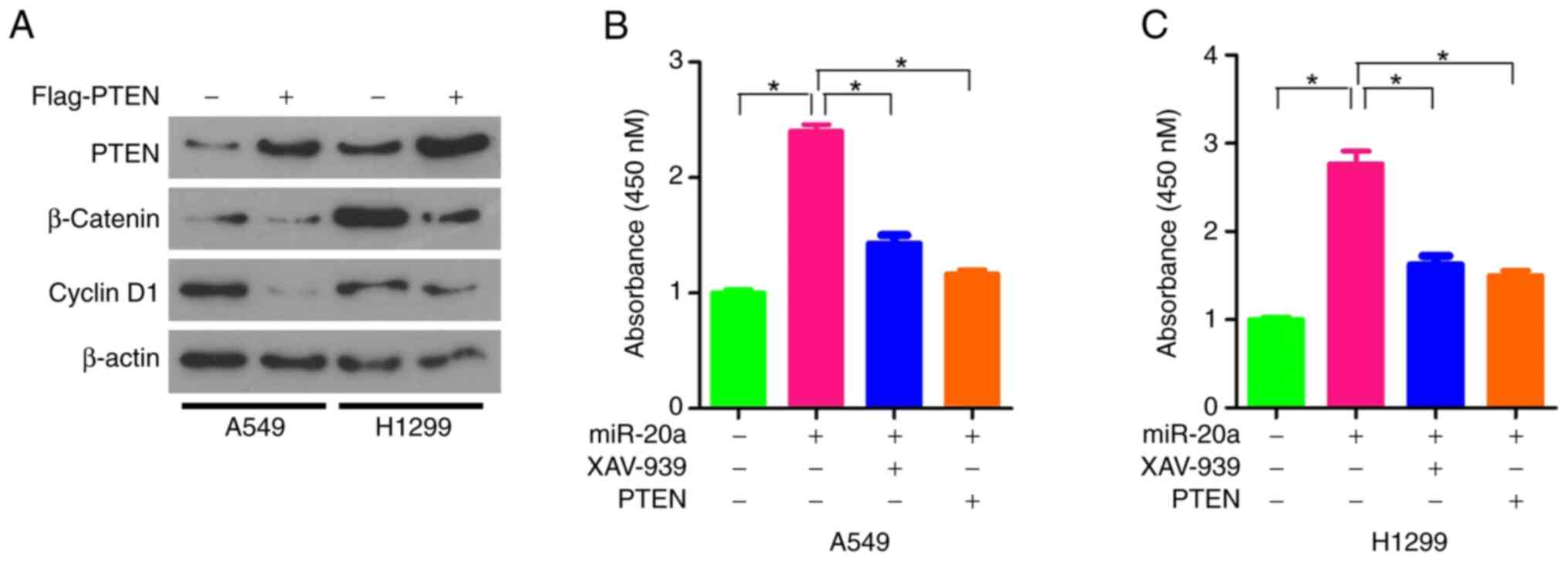
A previous study has shown that there was a
synergistic effect between the knockdown of PTEN expression and the
activation of Wnt/β-catenin pathway (15). Thus, we hypothesized that miR-20a
may enhance the proliferation of NSCLC cells by regulating the
PTEN/Wnt/β-catenin pathway. Therefore the protein expression levels
of β-catenin and Cyclin D1 were analyzed. The results revealed that
they were both inhibited by the overexpression of PTEN in the A549
and H1299 cell lines (Fig. 5A).
XAV-939 is a potent tankyrase inhibitor that targets the
Wnt/β-catenin signaling pathway. XAV-939 stabilizes Axin by
inhibiting tankyrase 1 and tankyrase 2, thereby stimulating
β-catenin degradation (16). The
miR-20a mimics were transfected into A549 and H1299 cells
respectively and the proliferation ability of A549 and H1299 cells
was analyzed with or without XAV-939 treatment. Compared with the
cells treated with miR-20 alone, the proliferation ability of A549
or H1299 cells treated with miR-20a combined with XAV-939 or PTEN
was significantly lower, but still higher than that of
untransfected and untreated cells (Fig. 5B and C). Additionally, it was found
that the inhibitory effect of PTEN was similar to that of XAV-939,
as the proliferation rate was similar in cells transfected with
miR-20a mimics and PTEN overexpression vector (Fig. 5B and C). In conclusion, these
results indicated that miR-20a may enhance the proliferation of
NSCLC cells by targeting PTEN and the activating Wnt/β-catenin
pathway.
Discussion
NSCLC is one of the most common malignant tumors
(17). However, the high mortality
rate of NSCLC has not decreased, mainly due to the lack of early
diagnosis, leading to the loss of surgical opportunity, as patients
with NSCLC are found at an advanced stage (18). Therefore, early diagnosis is the
key to reduce mortality. A number of studies have reported that
some important functional miRNAs could play a role in NSCLC cells.
For example, miR-196b-5p-mediated downregulation of TSPAN12 and
GATA6 promotes tumor progression in NSCLC (19). miR-7-5p suppresses tumor metastasis
of NSCLC by targeting NOVA2 (20).
miR-20a belongs to the miR-17-92 cluster and is
located on chromosome 13q31.1 (21). Previous studies have shown that
miR-20a is upregulated in liver cancer and breast cancer,
indicating that miR-20a could play a key role in tumorigenesis and
progression (21,22). However, miR-20a could act as a
tumor suppressor in other types of tumor, including endometrial and
liver cancer (21,23). These findings indicated that the
function of miR-20a may vary between different cell types. In a
previous study, using samples from patients with adenocarcinoma
from China, miR-20a-induced WTX deficiency promoted gastric cancer
progression by regulating the PI3K/AKT signaling pathway (24). A previous study has demonstrated
that the high expression level of plasma miR-20a was associated
with shorter disease free survival (DFS) and overall survival (OS)
in patients with NSCLC, which was an independent risk factor
(25). Consistent with this
finding, the present study revealed that miR-20a promoted the
proliferation of NSCLC cells.
To further determine how miR-20a acted as an
oncogene, the present study confirmed that PD-L1 was a potential
target for miR-20a and miR-20a promoted the expression level of
PD-L1. The PD-1/PD-L1 axis is responsible for cancer immune escape
and has a marked effect on cancer therapy (26). Blocking inducible PD-L1 expression,
upon tumor-antigen specific T cell infiltration, was the key event
leading to the response to anti-PD-1 or anti-PD-L1 antibody therapy
in patients with NSCLC (27). The
suppression of PD1/PD-L1 immune checkpoint provided a promising new
method for the treatment of NSCLC. A number of studies showed that
PD-L1 was highly expressed in NSCLC cells (27,28).
The use of drugs to block PD1/PD-L1 immune checkpoint (such as
atezolizumab) could prolong the survival time in patients with
advanced NSCLC (29). The present
study found that miR-20a promoted the expression level of PD-L1 by
inhibiting the expression level of PTEN, which in turn promoted the
proliferation of NSCLC cells.
PTEN is a key tumor suppressor gene and one of the
most frequently mutated genes in human tumors. The expression level
of PTEN was downregulated in numerous tumor types (30–32).
A previous study described the association between the expression
of PD-L1 and PTEN (31). In other
cases, knockdown or inhibition of PTEN resulted in increased PD-L1
expression level in breast and prostate cancer (31,32).
The expression level of PD-L1 in prostate, breast and lung cancer
was dependent on PI3K and regulated by PTEN (27,29).
However, this association was context-dependent, as the regulation
of PD-L1 expression was controlled by a number of factors and
pathways (33). A previous study
has shown that the increased expression level of PD-L1 could
directly mediate the activation of the β-catenin/TCF/LEF
transcriptional complex (34). The
Wnt/β-catenin signal pathway played an important role in regulating
the growth and metastasis of glioblastoma cells (34). Another study also confirmed that
the Wnt/β-catenin signaling pathway could promote the growth and
progression of numerous cancers, including NSCLC (35). The Wnt/β-catenin signaling pathway
could be activated by frizzled and low-density lipoprotein
receptor-related protein 5/6. Dishevelled was recruited and
phosphorylated to induce the dissociation of glycogen synthase
kinase-3 beta (GSK-3β) from axon proteins (36). The inhibition of GSK-3β expression
led to the inactivation of the degradation complex (36). The phosphorylation and degradation
of β-catenin were inhibited and accumulated stably in the cytoplasm
(36). A previous study showed
that active Wnt/β-catenin signal transduction could lead to T cell
rejection and resistance to the treatment of anti-PD-L1/anti-CTLA-4
monoclonal antibodies in melanoma (37). XAV-939 is an effective inhibitor of
the Wnt/β-catenin signal pathway. The present study found that PTEN
inhibited the expression of β-catenin and cyclin D1. The inhibitory
effect of PTEN on cell proliferation was similar to that of
XAV-939.
In summary, the present study showed that miR-20a
promoted the proliferation of NSCLC cells by inhibiting the
expression level of PTEN and enhancing the expression level of
PD-L1. These findings suggest that miR-20a could be used as a
biomarker and therapeutic target in the treatment of NSCLC.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Professor Jun Ma
(Second Affiliated Hospital of Zhengzhou University, Zhengzhou,
Henan, China) for his technical assistance and Professor Weijuan
Cao (Zhejiang Pharmaceutical College, Ningbo, Zhejiang, China) for
her assistance in revising the language.
Funding
The present study was supported by Henan Science and Technology
Project (grant no. 182102311236) and Henan Health and Planning
Commission (grant nos. 2018020495 and LHGJ20200406).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JG and MC designed the research. JG, YS, FJ, YW and
LC performed the experiments. JG and YS wrote the manuscript. JS
and PS provided materials and performed data analysis. JG, YS and
MC confirm the authenticity of all raw data. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work were appropriately investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Second Affiliated Hospital of Zhengzhou
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Backman M, La Fleur L, Kurppa P,
Djureinovic D, Elfving H, Brunnstrom H, Mattsson JSM, Lindberg A,
Ponten V, Eltahir M, et al: Infiltration of NK and plasma cells is
associated with a distinct immune subset in non-small cell lung
cancer. J Pathol. 255:245–256. 2021. View Article : Google Scholar
|
|
2
|
Park K, Haura EB, Leighl NB, Mitchell P,
Shu CA, Girard N, Viteri S, Han JY, Kim SW, Lee CK, et al:
Amivantamab in EGFR exon 20 insertion-mutated non-small-cell lung
cancer progressing on platinum chemotherapy: Initial results from
the CHRYSALIS phase I study. J Clin Oncol. 39:3391–3402. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mielgo-Rubio X, Martín M, Remon J, Higuera
O, Calvo V, Jarabo JR, Conde E, Luna J, Provencio M, De Castro J,
et al: Targeted therapy moves to earlier stages of non-small-cell
lung cancer: Emerging evidence, controversies and future
challenges. Future Oncol. 17:4011–4025. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu Z, Sun D, Zhu Q and Liu X: The
screening of immune-related biomarkers for prognosis of lung
adenocarcinoma. Bioengineered. 12:1273–1285. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ye Q, Putila J, Raese R, Dong C, Qian Y,
Dowlati A and Guo NL: Identification of prognostic and
chemopredictive microRNAs for non-small-cell lung cancer by
integrating SEER-medicare data. Int J Mol Sci. 22:76582021.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu X, Kudo M, Huang X, Sui H, Tian H,
Croce CM and Cui R: Frontiers of microRNA signature in non-small
cell lung cancer. Front Cell Dev Biol. 9:6439422021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hou J, Meng F, Chan LW, Cho WC and Wong
SC: Circulating plasma microRNAs as diagnostic markers for NSCLC.
Front Genet. 7:1932016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Y, Chen P, Zu L, Liu B, Wang M and Zhou
Q: MicroRNA-338-3p suppresses metastasis of lung cancer cells by
targeting the EMT regulator Sox4. Am J Cancer Res. 6:127–140.
2016.PubMed/NCBI
|
|
9
|
Peng LP: Regarding: MicroRNA-126 targeting
PIK3R2 inhibits NSCLC A549 cell proliferation, migration, and
invasion by regulation of PTEN/PI3K/AKT pathway. Clin Lung Cancer.
22:e446–e450. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han Y, Liu D and Li L: PD-1/PD-L1 pathway:
Current researches in cancer. Am J Cancer Res. 10:727–742.
2020.PubMed/NCBI
|
|
11
|
Cha JH, Chan LC, Li CW, Hsu JL and Hung
MC: Mechanisms controlling PD-L1 expression in cancer. Mol Cell.
76:359–370. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng S, Wang R, Zhang X, Ma Y, Zhong L, Li
K, Nishiyama A, Arai S, Yano S and Wang W: EGFR-TKI resistance
promotes immune escape in lung cancer via increased PD-L1
expression. Mol Cancer. 18:1652019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang Z, Su W, Lu T, Wang Y, Dong Y, Qin
Y, Liu D, Sun L and Jiao W: First-line immune-checkpoint inhibitors
in non-small cell lung cancer: Current landscape and future
progress. Front Pharmacol. 11:5780912020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang B, Zhao H, Zhao L, Zhang Y, Wan Q,
Shen Y, Bu X, Wan M and Shen C: Up-regulation of OLR1 expression by
TBC1D3 through activation of TNFα/NF-κB pathway promotes the
migration of human breast cancer cells. Cancer Lett. 408:60–70.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Patel R, Brzezinska EA, Repiscak P, Ahmad
I, Mui E, Gao M, Blomme A, Harle V, Tan EH, Malviya G, et al:
Activation of β-catenin cooperates with loss of pten to drive
AR-independent castration-resistant prostate cancer. Cancer Res.
80:576–590. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu J, Liu D, Sun X, Yang K, Yao J, Cheng
C, Wang C and Zheng J: CDX2 inhibits the proliferation and tumor
formation of colon cancer cells by suppressing Wnt/β-catenin
signaling via transactivation of GSK-3β and Axin2 expression. Cell
Death Dis. 10:262019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rotoli D, Santana-Viera L, Ibba ML,
Esposito CL and Catuogno S: Advances in oligonucleotide aptamers
for NSCLC targeting. Int J Mol Sci. 21:60752020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Horvath L, Thienpont B, Zhao L, Wolf D and
Pircher A: Overcoming immunotherapy resistance in non-small cell
lung cancer (NSCLC)-novel approaches and future outlook. Mol
Cancer. 19:1412020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liang G, Meng W, Huang X, Zhu W, Yin C,
Wang C, Fassan M, Yu Y, Kudo M, Xiao S, et al: miR-196b-5p-mediated
downregulation of TSPAN12 and GATA6 promotes tumor progression in
non-small cell lung cancer. Proc Natl Acad Sci USA. 117:4347–4357.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xiao H: MiR-7-5p suppresses tumor
metastasis of non-small cell lung cancer by targeting NOVA2. Cell
Mol Biol Lett. 24:602019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang BM, Zhao JR, Huo TT, Zhang ML and Wu
XH: MiR-20a lowers chemosensitivity of liver cancer Huh-7 cells via
regulating NF-кB expression. Eur Rev Med Pharmacol Sci.
24:11569–11577. 2020.PubMed/NCBI
|
|
22
|
Shi KY, Fan LY, Xu D, Ren LP, Wang LP,
Chen LY and Wang LJ: MiR-20a suppresses proliferation and
facilitates apoptosis of breast cancer cells via the MTOR signaling
pathway. Eur Rev Med Pharmacol Sci. 24:11650–11657. 2020.PubMed/NCBI
|
|
23
|
He Y, Ma H, Wang J, Kang Y and Xue Q:
miR-20a-5p inhibits endometrial cancer progression by targeting
janus kinase 1. Oncol Lett. 21:4272021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li J, Ye D, Shen P, Liu X, Zhou P, Zhu G,
Xu Y, Fu Y, Li X, Sun J, et al: Mir-20a-5p induced WTX deficiency
promotes gastric cancer progressions through regulating PI3K/AKT
signaling pathway. J Exp Clin Cancer Res. 39:2122020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chaniad P, Trakunran K, Geater SL,
Keeratichananont W, Thongsuksai P and Raungrut P: Serum miRNAs
associated with tumor-promoting cytokines in non-small cell lung
cancer. PLoS One. 15:e02415932020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yi M, Niu M, Xu L, Luo S and Wu K:
Regulation of PD-L1 expression in the tumor microenvironment. J
Hematol Oncol. 14:102021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hsu PC, Jablons DM, Yang CT and You L:
Epidermal growth factor receptor (EGFR) pathway, yes-associated
protein (YAP) and the regulation of programmed death-ligand 1
(PD-L1) in non-small cell lung cancer (NSCLC). Int J Mol Sci.
20:38212019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang H, Shan Q, Guo J, Han X, Zhao C, Li H
and Wang Z: PDL1 high expression without TP53, KEAP1 and EPHA5
mutations could better predict survival for patients with NSCLC
receiving atezolizumab. Lung Cancer. 151:76–83. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Brody R, Zhang Y, Ballas M, Siddiqui MK,
Gupta P, Barker C, Midha A and Walker J: PD-L1 expression in
advanced NSCLC: Insights into risk stratification and treatment
selection from a systematic literature review. Lung Cancer.
112:200–215. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xie P, Peng Z, Chen Y, Li H, Du M, Tan Y,
Zhang X, Lu Z, Cui CP, Liu CH, et al: Neddylation of PTEN regulates
its nuclear import and promotes tumor development. Cell Res.
31:291–311. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dastmalchi N, Hosseinpourfeizi MA,
Khojasteh SMB, Baradaran B and Safaralizadeh R: Tumor suppressive
activity of miR-424-5p in breast cancer cells through targeting
PD-L1 and modulating PTEN/PI3K/AKT/mTOR signaling pathway. Life
Sci. 259:1182392020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rennier K, Shin WJ, Krug E, Virdi G and
Pachynski RK: Chemerin reactivates PTEN and suppresses PD-L1 in
tumor cells via modulation of a novel CMKLR1-mediated signaling
cascade. Clin Cancer Res. 26:5019–5035. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lamberti G, Sisi M, Andrini E, Palladini
A, Giunchi F, Lollini PL, Ardizzoni A and Gelsomino F: The
mechanisms of PD-L1 regulation in non-small-cell lung cancer
(NSCLC): Which are the involved players? Cancers (Basel).
12:31292020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Du L, Lee JH, Jiang H, Wang C, Wang S,
Zheng Z, Shao F, Xu D, Xia Y, Li J, et al: β-Catenin induces
transcriptional expression of PD-L1 to promote glioblastoma immune
evasion. J Exp Med. 217:e201911152020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Song C, Xiong G, Yang S, Wei X, Ye X,
Huang W and Zhang R: PRDX1 stimulates non-small-cell lung carcinoma
to proliferate via the Wnt/β-catenin signaling. Panminerva Med. Sep
3–2020.(Epub ahead of print). View Article : Google Scholar
|
|
36
|
He S and Tang S: WNT/β-catenin signaling
in the development of liver cancers. Biomed Pharmacother.
132:1108512020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Trujillo JA, Luke JJ, Zha Y, Segal JP,
Ritterhouse LL, Spranger S, Matijevich K and Gajewski TF: Secondary
resistance to immunotherapy associated with β-catenin pathway
activation or PTEN loss in metastatic melanoma. J Immunother
Cancer. 7:2952019. View Article : Google Scholar : PubMed/NCBI
|