In recent years, important advances have been made
in the exploration of the molecular pathogenesis of tumorigenesis
and progression, but this has not been applied to significantly
improve patient prognosis. It is thus essential to identify
biomarkers for diagnosis, as well as means of reducing drug
resistance and delivering treatments, including personalized
targeted therapies in the study of GBM. The role of Hedgehog (HH)
signaling in the pathophysiology of GBM is underscored by a growing
number of publications (5–7). The HH pathway is increasingly being
revealed to have an important role in the growth, progression,
prognosis and treatment of GBM (8–10).
The present review will discuss the contribution of
HH signaling in the development and treatment of GBM. Chemotherapy,
targeted therapy and radiotherapy in the HH pathway will also be
discussed and the issue of improving partial drug resistance
through this pathway will be addressed.
GBM originates from the glial stem or progenitor
cells and is characterized by molecular heterogeneity, with a mean
survival of only 15 months after diagnosis (11). Commonly mutated genes and core
pathways in sporadic GBM were identified based on molecular mapping
and three major GBM subpopulations were identified in combination
with other dimensions (gene expression, DNA methylation). The DNA
methylated α group amplified cyclin-dependent kinase (CDK)4 and
platelet-derived growth factor in the three ways (classical gene
expression; classical like; receptor tyrosine kinase II).
High-frequency amplification of EGFR and homozygous deletion of CDK
inhibitor 2A/B occurred in the DNA methylation group.
Mesenchymal/mesenchymal subtypes are abundant in tumors with loss
of neurofibromatosis type 1 and increased tumor macrophage
infiltration (12,13). The above three types are the most
common types of GBM and all involve mutations in telomerase reverse
transcriptase promoters (14,15).
In addition, characteristic epigenetic patterns are associated with
certain putative driving mutations that are important in GBM,
according to recent studies (16,17).
Examples include mutated isocitrate dehydrogenase (IDH)1 and IDH2,
H3.3 histone A or H3 clustered histone 2 mutations, particularly
H3K27M in diffuse midline glioma and H3G34R/H3G34V mutations in
young patients with GBM (16,17).
However, their clinical implication for these GBM subtypes has not
been proven. These studies indicate that different subtypes of GBM
are caused by different oncogenes, which paves the way for the
exploration of highly specific personalized targets.
GBM is characterized by continuous vascularization,
tissue invasion and metastasis, metabolic recombination or
alteration, immune regulation and promotion of the tumor
microenvironment. All of the above characteristics lead to high
resistance of GBM to radiotherapy and chemotherapy, which brings a
non-negligible challenge to the treatment of the disease (18).
GBM has different subtypes, but the current
international treatment methods mainly include chemotherapy [such
as temozolomide (TMZ)], radiotherapy (RT) and surgical treatment.
Monotherapy may be well tolerated in elderly patients (>65
years) with poor functional status. It has been reported that
low-grade RT (40 Gy/15 doses of 2.67 Gy over 3 weeks) was higher
than the standard 60 Gy for 6 weeks (19,20).
Relapsing patients may be treated with surgery (as palliative care
only) or other options include TMZ reactivation, nutrition and
bevacizumab. However, there is no specific clinical evidence of
prolonged survival (21,22). Several valuable clinical trials are
under development for the treatment of GBM, including targeted
molecular (precise) therapies (targeting gene mutations and
associated signaling pathways, DNA damage repair, tumor
metabolism), checkpoint inhibitors/immunomodulation agents and
viral therapies. Despite the GBM treatment options available,
metastatic disease remains a great concern. Therefore, it is of
marked importance to find novel therapeutic targets and new drugs
targeting the HH signaling pathway to regulate the occurrence,
development, treatment and chemotherapy resistance of GBM (23,24).
HH is a morphogenetic gene, which is highly
conserved from drosophila to humans. The HH signaling pathway has
an important role in embryonic development, cell proliferation,
differentiation and maintenance of tissue polarity (25). Inactivation of this pathway during
development may lead to congenital defects, while over-activation
in adults is related to tumorigenesis (26,27).
The HH protein family includes Sonic HH (SHH), Indian HH (IHH) and
desert HH (DHH) (28). In mammals,
the mechanism of HH signaling is complex and occurs in primary
cilia (PC) (29). In PC, HH
protein binds to 12 transmembranes (TM) receptors [Patch1 (PTCH1)
and PTCH2] to activate pathways, so that 7-TM protein smoothened
(SMO) is inhibited (30). The HH
signal is transmitted downstream of SMO through the complex
composed of kinesin 7 (KIF7), suppressor of fused homolog (SUFU)
and full-length glioma-associated oncogenes (GliFL), which promotes
the dissociation of SUFU from GLI protein and then releases
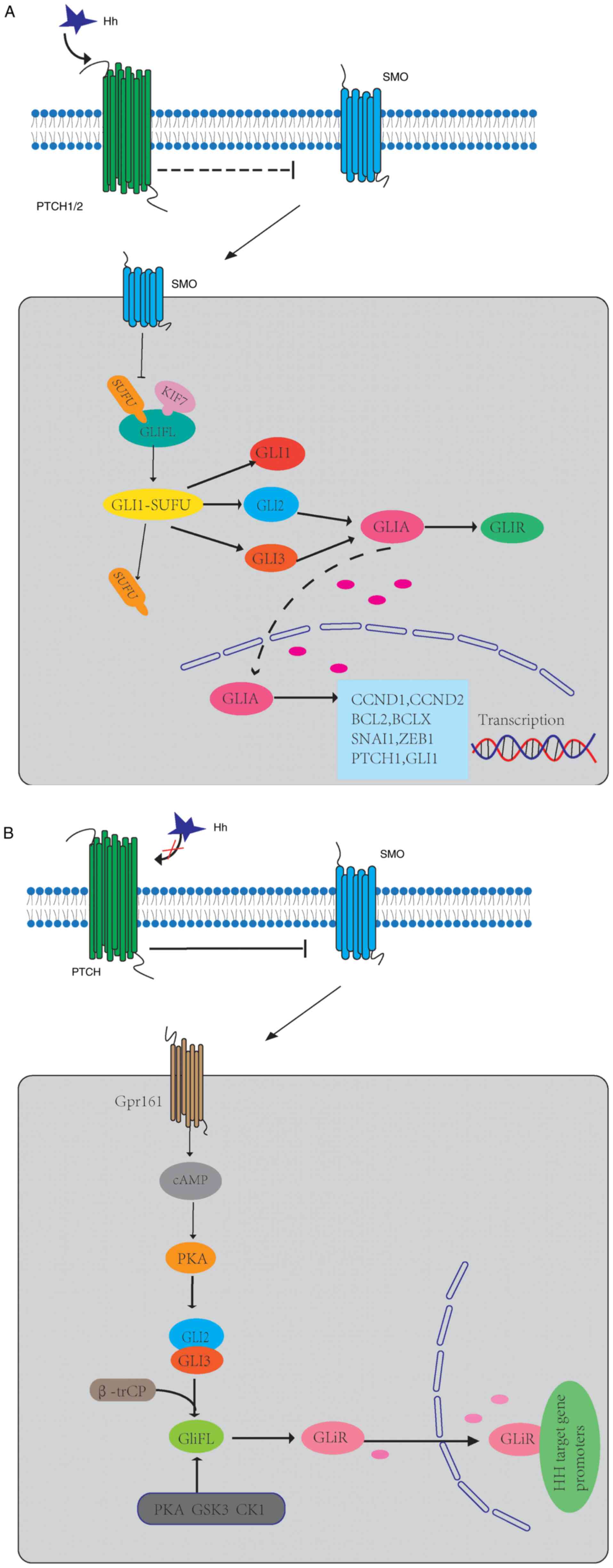
transcription factors (GLI1, GLI2 and GLI3) (31,32).
GLI2 and GLI3 constitute GliFL, which act as both GLI activators
(GLIA) and GLI inhibitors (GLIR) (33,34).
After activation of SMO, GLI2/3 P1-6 clusters were dephosphorylated
and separated from SUFU (35),
which facilitates the transfer of GLIA into the nucleus and the
initiation of the transcription of target genes, and their pathway
genes (PTCH1, GLI1) (36,37). GLI1 is the main HH target gene and
its expression further promotes the activation of the HH signaling
pathway at the transcriptional level (38). KIF7, in turn, coordinates HH
signaling at the top of the PC and avoids GLI3 from cracking into
an inhibited form in response to HH (39). This GLI transcription factor signal
transduction pathway is the canonical HH signaling pathway
(Fig. 1A).
When HH ligand is absent, PTCH inhibits the activity
of SMO by inhibiting the translocation of SMO in PC (40). GLIFL is phosphorylated by protein
kinase A (PKA), glycogen synthase kinase 3 (GSK3) and casein kinase
1 (CK1), and then recognized by β-trCP and cleaved into GLIR
(41,42). This results in the proteolytic
cleavage of GLIFL into the form of a C-terminal truncated repressor
known as a GLIR (33). SUFU is a
negative regulator that binds to GLI proteins and prevents them
from migrating to the nucleus (43). GLIR enters the nucleus, binds to HH
target gene promoters and inhibits their expression (Fig. 1B).
HH signaling, canonical and non-canonical signaling,
exist in parallel, and the mechanisms are complex. Non-canonical HH
signaling is the most common HH-dependent reaction process,
independent of GLI transcription factors or PC (44). Non-canonical HH signaling pathways
may be divided into type I (independent of SMO) and type II
(dependent on SMO) (45).
The canonical HH pathway is related to tumorigenesis
and detransformation development. In adults, this signal
abnormality has a key role in promoting the proliferation and
differentiation of numerous tumor types. Its carcinogenic
mechanisms mainly include abnormal cell differentiation,
neovascularization, epithelial-mesenchymal transition (EMT) and
enhanced invasiveness (46–48).
Initially, HH signaling was mainly studied in brain cancer,
skeletal muscle and skin cancer (49–51).
However, in recent years, studies have indicated that this pathway
is abnormal in numerous tumor types, including stomach, pancreas,
lung and breast tumors (52–54).
As HH signaling is activated in various types of cancer and
contributes to cancer proliferation, progression and invasiveness,
the HH signaling pathway is anticipated to provide new targets for
cancer therapy.
The tumor microenvironment/stroma is closely related
to tumorigenesis, metastasis and invasion (55,56).
The tumor microenvironment/stroma is mainly composed of endothelial
cells, adipocytes, immune cells and cancer-associated fibroblasts
(CAFs) (57). CAFs are able to
secrete soluble factors to stimulate cancer cells, thereby
triggering tumor metastasis and chemotherapy resistance (58–60).
Recombinant human Sonic HH N-terminal peptide (rhSHH) enhances HH
signaling, accompanied by increased mRNA and protein levels of
matrix metalloproteinase-2 (MMP2) and MMP9. Furthermore, the
protein expression of GLI1 was positively associated with the
protein expression of MMP-2 and −9, which promoted the adhesion and
invasion of GBM cells (60). It
has been reported that gap junctions have a role in tumor growth
and progression. Torrisi et al (61) modulated SHH signaling and connexin
43 (CX43)-based intercellular communication in an in vitro
model. Modulation of SMO with the use of a known agonist (i.e.,
taxamine) and a known antagonist (i.e., cyclodopamine) affected
CX43 expression levels and thus affected related functions. In
addition, SMO activation also promoted cell proliferation and
migration. Of note, inhibition of the CX43 channel prevented the
SMO-induced effects (61).
Therefore, further exploration of the mechanisms of
the HH signaling pathway in the tumor microenvironment may lead to
better targeting of this pathway to fight cancer.
In the development of GBM disease, PC serve as cell
antennae to transmit and regulate a variety of signaling pathways
and SHH is one of the most important pathways. SHH levels are
significantly increased in GBM cells compared with normal brain
tissue and SHH overexpression induced neuroectodermal angiogenesis
during mouse embryonic development (62–64).
A study has indicated that Fms-related tyrosine kinase 1 (FLT1) is
significantly increased in GBM cells and overexpression of FLT1
increased the expression of SHH in cells (64). Knockdown of SHH reduced the
migration and invasion mediated by FLT1 overexpression, while
overexpression of SHH restored the migration and invasive ability
of FLT1 knockdown (64). FLT1 is a
tyrosine kinase receptor that binds VEGF-A with several times the
affinity of other kinases inserted into domain receptors and has
been reported to promote tumor growth and metastasis (65). VEGF-A is one of the key factors
promoting tumor angiogenesis and activation of the VEGF-A pathway
requires the binding of VEGF-A to its receptor FLT1 to generate
downstream signals to stimulate the proliferation and development
of tumor cells and provide tumor blood vessels for the growth and
metastasis of GBM (64,66). In addition, brain
tumorigenic-initiating cells produce DHH ligands to realize the
paracrine DHH/PTCH2 signaling cascade, transmit high permeability
and angiogenesis, and also promote GBM growth (6). Chen et al (67) reported that C-terminal binding
protein 2 (CtBP2) expression was increased and zinc finger and BTB
domain containing 18 (ZBTB18) expression was decreased in GBM
tissues, and the two were negatively correlated. CtBP2 short
hairpin (sh)RNA interacts with ZBTB18 to block cells in G0/G1
phase, inhibit the SHH-Gli1 pathway and reduce the tumor volume
(67). However, whether this
effect is exerted by increasing SHH gene expression has remained to
be elucidated. Therefore, targeted FLT1 or CtBP2 therapy may be a
promising direction to develop anti-metastasis agents.
GBM develops through a complex interlocking
signaling pathway. RhSHH enhances the HH signaling pathway, which
increases the production of MMP-2 and −9 through the PI3K/AKT
pathway, thereby regulating migration and invasion of basal
membrane cells and promoting GBM cell adhesion, invasion and
migration (60). By contrast,
triggering the vasoactive intestinal peptide receptor system is
triggered to reduce GBM cell migration and invasion through
PKA-dependent PI3K/AKT and SHH/GLI1 pathway blocking (68). Similarly, Henao-Restrepo et
al (69) reported that
PI3K/AKT/mammalian target of rapamycin complex 1 (mTORC1) and
SHH/GLI signaling pathway proteins were expressed differently in
human gliomas with different tumor types and grades, suggesting
that the activation of these signaling networks is related to the
occurrence and development of high-grade gliomas.
Multiple studies have indicated that the SHH
signaling pathway promotes the plasticity of cancer cells by
regulating the adhesion between cells and the extracellular matrix,
thus increasing the motility and aggressiveness of cells, leading
to poor prognosis of patients (60,65,67,68).
Statistical analysis of the The Cancer Genome Atlas (TCGA) dataset
(TCGA-Glioblastoma June 2016) suggested that SHH upregulation was
associated with decreased overall survival (64).
Hedgehog-interacting protein (HHIP), which is
located on chromosome 4q31.21-31.3, is defined as an antagonist of
SHH, IHH and DHH. Chang et al (9) were the first to demonstrate that the
expression of HHIP determined by immunohistochemistry is an
independent prognostic marker of favorable outcomes in patients
with GBM.
Although GLI1 was originally identified as the
amplified gene in malignant human gliomas (70), GLI1 amplification is infrequent in
most cancers such as GBM (71).
However, since GLI1 is a vital downstream target of the HH pathway,
the mRNA level of GLI1 is a reliable indicator of HH pathway
activity, this suggests that control of GLI1 protein conversion is
critical for GLI-dependent transcription and regulation of the HH
signaling pathway (53). And GLI1
protein levels are upregulated in a variety of cancers, and high
levels of GLI1 are often associated with tumor progression
(72,73). Low GLI1 mRNA levels were similarly
negatively correlated with survival in patients with GBM. GLI1 mRNA
expression in GBM was significantly lower than in patients with
high-HH-medulloblastoma (MB) but significantly higher than in
patients with low-HH-MB, and GLI1 mRNA expression is a single
continuous distribution, rather than being discrete high/low
clusters (5,74). GLI1 promotes the nuclear import of
GLI1 into GBM multiforme cells through its transcription factor
Forkhead box M1 (FOXM1), thereby increasing the expression of its
target genes (75).
It was observed that both the activation of
metabolic glutamate receptor subtype 4 and naringin are able to
inhibit the expression of GLI-1 in cells and affect HH signaling
pathway transduction, thus inhibiting cell proliferation and
promoting cell apoptosis to inhibit the growth of GBM cells
(77,78). These may be potential drug targets
for controlling GBM cell growth by blocking HH signaling.
The HH, mTOR, Notch and Wnt/β-catenin signaling
pathways are important signaling pathways that regulate GSC
stemness and self-renewal (27,89,90),
However, the self-renewal and abnormal differentiation of GSCs and
their ability to promote the formation of drug resistance to RT and
chemotherapy are the main reasons for the recurrence and invasion
of GBM after conventional treatment (91,92).
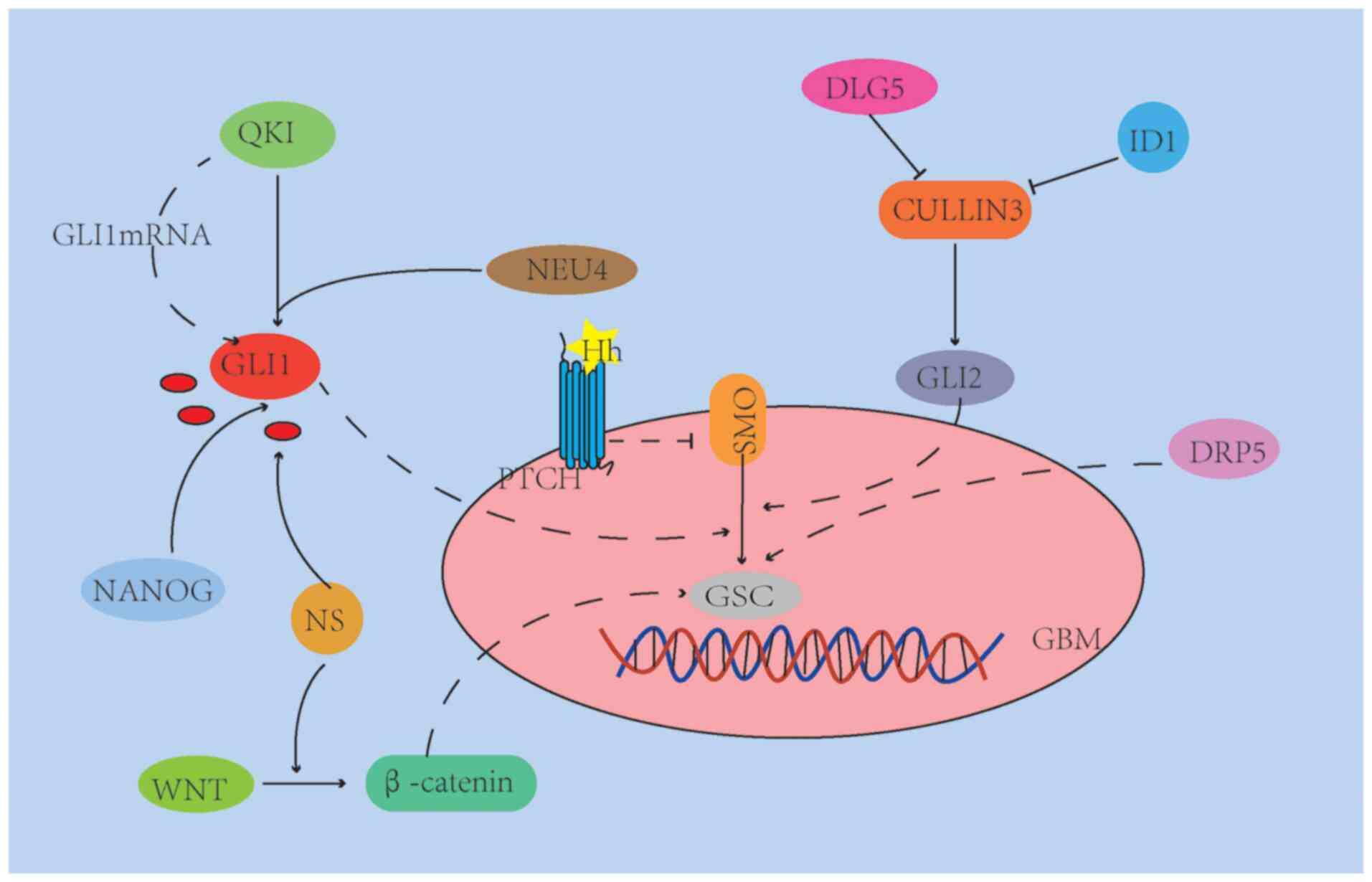
The mechanisms of how GSCs during invasion through the HH pathway,
particularly in the face of complex and changing brain tissue
anatomy, are presented in Fig.
2.
In summary, the neurobiology and basement membrane
invasiveness of neural stem cells involves multiple molecular
pathways that are interrelated. Therefore, targeting
cross-signaling pathways (e.g., Wnt/HH signaling) and specific
markers may be a better therapeutic approach for GSCs.
The increased understanding of the key role of HH
signaling in cancer has led to the development of pathway-specific
inhibitors and the reuse of existing drugs that regulate HH/GLI
(Table I). Drugs currently used in
the clinic target SMO; among them, Vismodegib and Sonidegib have
been approved by the US Food and Drug Administration for the
treatment of basal cell carcinoma (BCC) (100,101) and medulloblastoma (101). However, mutations leading to drug
resistance may occur, and thus, compounds that inhibit HH signaling
downstream of SMO are urgently required and further research on the
effects of HH/GLI pathway modulators in combination with anticancer
drugs should be performed in order to provide evidence to pave the
road for the future use of the combination of HH/GLI inhibitors and
anticancer drugs.
The mechanisms of GBM cell migration and invasion
are complex and involve a series of mechanisms, including adhesion
of GBM cells to the extracellular matrix (ECM) and ECM remodeling
and degradation (102). As with
other malignant tumor types, the growth, metastasis and invasion of
GBM also depend on tumor angiogenesis. Although gliomas are
characterized by hypervascularization, there are unavoidable
disadvantages to anti-angiogenesis, such as reactive resistance
mediated by the tumor microenvironment, and invasion and metastasis
of tumor cells activated by hypoxia responses (103,104). During invasion and metastasis,
GBM cells lose the polarized phenotype of epithelial cells and
acquired mesenchymal characteristics, which is referred to as EMT
(105). EMT is an active,
drug-resistant, low-proliferative transient state that is
frequently a feature of cancer as a whole but is seen in GBM in
particular (106–108). Tubasatin A, a histone deacetylase
6 (HDAC6) inhibitor, reduced the expression of mesenchymal markers
in GBM cells and contributed to the reversal of EMT (109). Feng et al (110) developed a pegylated poly (lactic
acid) based nano-drug delivery system (nanoparticles) and modified
CK peptides on its surface via GYG connectors to promote
multitargeted delivery of Paclitaxel vasculogenic mimicry channels,
tumor neovascularization and glioma cells. Similarly, Kast et
al (111) proposed the EIS
regimen (combination of itraconazole, metformin, naproxen,
pirfenidone, quetiapine and rifampicin) that was able to safely and
effectively block EMT of GBM. GBM progression may be inhibited by
targeting tumor angiogenesis and EMT. Although these animal models
are not perfect, they may be used to explore the effectiveness of
new treatments for GBM prior to clinical phase I/II studies.
The HH signaling pathway is closely related to PC
function, and thus, inhibiting PC function may help inhibit GBM
proliferation, malignant development and treatment resistance
(112). A previous study reported
that the development of resistance to acquired kinase inhibitors is
associated with upregulation of PC, uncontrolled PC length and
abnormal activation of SHH signaling. Knockdown of KIF7 was
observed to control the length and integrity of the PC and
re-sensitize GBM cells (113). In
addition, Dynarrestin was able to reversely inhibit intraflagellar
transport of SMO flux in PC and inhibit HH pathway-dependent
neuronal precursors and tumor cell proliferation (23). Therefore, Dynarrestin is a
promising compound for the pharmacochemical development of
anticancer drugs.
HH signaling has been reported to be abnormally
activated in >30% of solid tumor types, including GBM (62,114). Abnormal activation of the SHH
pathway is associated with GBM resistance to temozolomide (TMZ) and
the reason is the high expression of methylguanine
methyltransferase (MGMT), which reverses the effects of TMZ on DNA
(115,116) and confirms cell protection from
TMZ-induced death by silencing three genes: MutS homolog 2 (a DNA
repair protein involved in MMR), PTCH2 and chloride channel
accessory 2 (a type 1 transmembrane protein that inhibits the Wnt
pathway) (24). Resistance to TMZ
was only slightly reversed by MGMT inhibitor O6-benzylamine, but a
marked further enhancement was achieved by addition of Honokiol
(117). Furthermore, the invasion
of GBM was reported to be associated with the presence of CSCs and
the SHH pathway has an important role in the maintenance and
proliferation of CSCs (118,119). After inhibiting SHH, LDE225
slowed down the growth of GBM and downregulated PTCH1 and GLI1
in vivo (120). CSCs
preferentially activate the DNA damage checkpoint response and
exhibit enhanced DNA repair ability; thus, SHH signaling via GLI1
in CSC has a role in GBM resistance to TMZ (121).
Glabrescione B is the first small molecule to bind
to GLI1 zinc fingers, impelling GLI1 activity by interfering with
its interaction with DNA. Thus, it inhibits the ability of
HH-dependent tumor stem cells to self-renew and cladogenesis. The
determination of the structural requirements for GLI1/DNA
interactions highlights their relevance to drug interference with
GLI signaling (10). Melamed et
al (122) developed
polyethylene imine-coated spherical nucleic acid nanoparticles
(PEI-SNAs) targeting GLI1. GLI1 PEI-SNAs bind scavenger receptors
on GBM cells and undergo endocytosis in a pit/lipid
raft/dynein-dependent manner, promoting the silencing of HH pathway
genes and downstream target genes. These genes promote an
aggressive, drug-resistant GBM phenotype. GLI1 PEI-SNAs not only
significantly increased the sensitivity of nerve spheres to
chemotherapy, but also further impaired the formation of dry nerve
spheres (123). Arsenic trioxide
also significantly reduced the cladogenesis of tumor neuroglobules
by inhibiting the HH pathway, inhibiting the proliferation of GBM
neuroglobules and promoting apoptosis (124). The combination of the SHH
inhibitor GANT-61 with TMZ increased the cytotoxicic effect of TMZ
and the combination of GANT-61 with TMZ increased the production of
reactive oxygen species in GBM cells, suggesting that inhibition of
the SHH pathway may sensitize GBM cells to the effects of TMZ by
increasing oxidative stress (114,124). GANT-61 induced apoptosis and
autophagy in GBM cells by increasing the expression of light chain
3II and lysed Caspase-3 and −9 (125). Furthermore, GLI inhibition
combined with TMZ increased the apoptosis rate of glioma stem cells
by 6.8-fold, thereby reducing the size and number of nerve spheres
grown from glioma stem cells (115). Yin et al (126) reported that the combination of
tumor suppressor gene miR-326 and curcumin significantly inhibited
the SHH/GLI1 pathway of glioma cells, independent of the P53
status, significantly increased apoptosis and reduced the
proliferation and migration of glioma cells. Similarly, Ji et
al (127) reported that a
novel PI3K inhibitor, XH30, inhibited tumor growth that was
resistant to TMZ. In terms of the mechanism, the role of XH30 may
be to reverse the activation of GLI1 induced by SHH by atypical HH
signaling and to reduce GLI1 activation by insulin-like growth
factor 1 (127). Thus, XH30 may
be a novel treatment option for TMZ-resistant GBM.
Traditional treatments for GBM include systemic
chemotherapy, RT and surgery. Han et al (128) synthesized three phosphorylated
peptides derived from GLI2 and combined them with the
cell-penetrating peptide TAT-[47–57]AYGRKKRRQRRR. The three mixed
phosphorylated polypeptides derived from GLI2 significantly
increased the level of GLI2 phosphorylation and decreased the
transcriptional activation of GLI2, and the radiation sensitization
of GBM cells was significantly higher than that in the control
group (128). HDAC6 was
upregulated in GSCs and inhibited HDAC6 down-regulated
glioma-associated oncogene GLI1 and PTCH1/2 receptors, as well as
SHH signaling components, expression and activity, thereby
inhibiting GSC proliferation, inducing differentiation and
increasing the apoptosis rate through the SHH/GLI1 signaling
pathway (109,129). Inhibition of HDAC6 by Tubasatin A
enhanced the radiosensitivity of GBM tumor cells. The mechanism may
be that HDAC6 inhibits checkpoint kinase (CHK)1 degradation induced
by down-regulation of X-linked inhibitor of apoptosis, which
reduced the DNA damage repair ability of GSCs, leading to increased
radiosensitivity (109,130).
In summary, target genes associated with the SHH/GLI
pathway provide promising new drug targets for inhibiting GBM
proliferation, as well as overcoming drug resistance and radiation
resistance of GSCs.
The steroidal alkaloid cyclopamine, an antagonist of
the HH coreceptor SMO, acts as an inhibitor of the HH pathway
(131). To limit the toxicity of
cyclodopamine to HH-dependent non-tumor cells, cyclodopamine
precursor drugs [e.g., cyclodopamine glucuronoside precursor drugs
(CGP-2) and 1b] are commonly used (132,133). It was indicated that CGP-2
inhibits the HH pathway more effectively than conventional TMZ
adjuvants (131). In the presence
of β-glucuronidase, the activated prodrug 1b was toxic and
downregulated the HH target gene GLI1 in C6 cells and C6-CSCs
(132). In U251 cells, tyramine
not only inhibited the HH/GLI1 signal transduction pathway, leading
to decreased MGMT expression, but also induced cell apoptosis by
activating caspase-3 cleavage, thus leading to increased
sensitivity of GBM to TMZ (133).
However, the combination of acepromazine and TMZ enhanced the
dryness and drug resistance of GBM cells by inducing the expression
of SOX-2 and OCT-4 and may lead to tumor recurrence in patients
(134). Therefore, the best
therapeutic strategy is to first inhibit the SHH pathway and then
administer TMZ (134,135). Liu et al (136) found that the combination of
capsulated propylamine and TMZ had synergistic cytotoxic effects
and was more likely to inhibit the ability to induce apoptosis and
eliminate neuroglobin formation by inhibiting GLI1 expression.
Therefore, MCyp may be used as a tumor stem cell inhibitor to
prevent tumor recurrence. Future efforts should be made to
investigate the possibility of using HH pathway inhibitors prior to
conventional chemotherapy in patients with GBM. Future efforts
should focus on the efficacy of HH pathway inhibitors prior to
systemic chemotherapy in patients with GBM.
The key to antitumor immunity is that
antigen-presenting cells (APCs) engulf tumor cells. TMZ may induce
an endoplasmic reticulum stress response, and the combination of
CD47 blocker and TMZ may produce significant prophagocytosis
(139,140). Increased tumor cell phagocytosis,
enhanced antigenic cross-presentation in APC and activation of
cyclic GMP-AMP interferon gene synthase stimulation leads to more
efficient T-cell effects. This connection between innate and
adaptive responses inhibits GBM growth while also activating immune
checkpoints. Sequential administration of an anti-programmed cell
death protein 1 (anti-PD1) antibody overcomes this potential
adaptive resistance (140).
However, the mechanism by which anti-PD1 antibodies reverse GBM
resistance through HH signaling remains to be elucidated. It has
been reported that GANT-61 is able to reduce the expression of
PD-L1 and the proliferation of tumor cells in vivo and in
vitro by using organic compound drugs for human gastric cancer.
Of note, anti-PD-L1 antibodies induced apoptosis of tumor cells in
organs of GLI2-expressing mice. Studies suggested that GLI2
expressed in gastric cancer cells is an internal regulator of PD-L1
and promotes tumor growth by inhibiting the anti-tumor response
(141,142). In summary, the HH pathway may
become a new immunotherapy target for GBM after further study.
The biological treatment of GBM has been studied for
numerous years, but the treatment of deadly cancers still poses a
great challenge. GBMs are highly invasive and susceptible to drug
resistance, resulting in a high mortality rate, and GBM accounts
for 2.9% of cancer-related deaths (143).
A key treatment issue for GBM is the high degree of
heterogeneity within the tumor. This heterogeneity further
complicates the differences among patients with GBM. In addition to
heterogeneity, GBM also has GSCs that contribute to tumor
proliferation, maintenance and drug resistance (144,145), and GSCs may respond differently
to TMZ or ionizing radiation (146). All of this makes routine
treatment difficult. Further research is required on the impact of
GBM heterogeneity on modern therapies, including molecular
immunotherapy and personalized therapy. The lymphocytes present in
GBM have an increased proportion of CD4+T cells and FOXP3+
regulatory T cells may induce signaling pathways that inhibit
immune responses (147,148), e.g., the expression of IDO enzyme
and STAT3 signals (149,150). However, GBM tumor-infiltrating
effector lymphocytes were observed to be rare (151,152). This may also be the reason why a
clinical trial of immune checkpoint blocking using the anti-PD1
antibody nivolumab (NCT02017717) used in patients with newly
diagnosed or relapsed unmethylated GBM (153), have not been successful. The
currently used immunotherapy for GBM may be broadly divided into
vaccine therapy, immune checkpoint blocking, oncolytic virus
therapy and chimeric antigen receptor T-cell therapy (154–156). In addition to immunotherapy, EGFR
using tyrosine kinase inhibitors (TKI), VEGF TKI and targeted
therapies for the PI3K/mTOR pathway have also been explored in GBM.
However, a phase 3 trial of deatuxizumab mandolin in combination
with standard therapy for the treatment of newly diagnosed
EGFR-amplified GBM was terminated early for being ineffective
(157), and mTOR inhibitors such
as everolimus (NCCTGN057K) and Taxiolimus (EORTC26082) also proved
to lack efficacy in phase 2 trials (158,159). A phase 2 trial of regorafenib
(REGOMA) in a relapsed setting indicated a therapeutic OS benefit
compared to lomustine, but the drug had minimal activity; thus,
VEGF monotherapy may have a limited effect in a non-selected
population (160).
It is necessary to study new targets for the
treatment of GBM. HH signaling has emerged as an attractive target
for cancer therapy and several HH inhibitors have been designed. To
date, SMO inhibitors were proven to have satisfactory efficacy in
BCC and medulloblastoma (100,101), but clinical trials for other
cancer types, such as colorectal, pancreatic or lung cancers, have
yielded poor results (161–163). In preclinical studies, compared
with HH and SMO inhibitors, GLI inhibitors had better anticancer
efficacy (164,165). In addition, GLI inhibitors
effectively inhibited the growth of numerous GLI-dependent cancers
by targeting the GLI-regulated SMO-independent pathway (166). As for inhibitors of GLI1 and GLI2
transcription factors, the anticancer drug arsenic trioxide is
currently the only drug undergoing clinical trials in solid tumors
and hematological malignancies (167). Although the use of HH inhibitors
in GBM has not been extensively investigated, numerous studies
suggested that HH inhibitors in combination with conventional
therapies may markedly increase efficacy and reduce the incidence
of drug resistance (124–126,133,137). Of course, this also requires a
large number of clinical trials to further verify whether HH
inhibitors are beneficial to the therapeutic efficacy of GBM.
Epigenetic regulators interact with drivers of GBM
stem cell-like cell proliferation. These drivers include Notch, HH
and WNT pathways. Previous studies suggested that these signaling
pathways may perform cross-talk with SHH signaling pathways
(27,71,80-81,89,90), which means that these signaling pathways may
be activated simultaneously in different tumor types. WNT/β-catenin
interacts with the SHH pathway through GLI1 and GLI2 by regulating
the expression of secreted crimp-related proteins. SHH signaling
was inhibited by GSK3β, a component of the WNT signaling pathway.
In certain tumor types, upregulation of the WNT signaling pathway
occurs sequentially when the SHH pathway is inhibited (168). In addition, the synergistic
effect of the inhibition of the SHH and PI3K/AKT/mTOR signaling
pathways may inhibit the proliferation of glioblastoma-initiating
cells (GICs), tumor growth and the formation of neural spheres and
clones, and induce cell apoptosis (169). Combined drug action targeting two
pathways or inhibition at the intersection of two pathways may be a
good breakthrough point for targeted therapy.
Current conventional therapies for GBM are
ineffective due to drug resistance issues and resistance may be
overcome through a combination of HH inhibitors or multilevel HH
signaling cascades, such as combinations of multiple targeted HH
drugs and multi-target HH inhibitors. In addition to pioneering new
approaches based on existing scientific theories, the effectiveness
of evaluating these therapies in clinical trials requires to be
further improved. This includes increasing the number of patients
with GBM in phase I trials of HH pathway inhibitors, thereby
providing more complete clinical trial data for the development of
more effective targeted therapeutic strategies.
Not applicable.
Funding: No funding was received.
Not applicable.
HW performed the literature search and selection.
XX was responsible for the conception, analysis and design of the
study. HW and XX were major contributors in writing of the
manuscript. DW and JP participated in the coordination of the study
and reviewed the manuscript. BT and YG were responsible for the
revision of the manuscript. QL and ZG were responsible for the
literature search and design of the study. All authors read and
approved the final manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Ostrom QT, Gittleman H, de Blank PM,
Finlay JL, Gurney JG, McKean-Cowdin R, Stearns DS, Wolff JE, Liu M,
Wolinsky Y, et al: American brain tumor association adolescent and
young adult primary brain and central nervous system tumors
diagnosed in the United States in 2008-2012. Neuro Oncol. 18 (Suppl
1):i1–i50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ostrom QT, Gittleman H, Truitt G, Boscia
A, Kruchko C and Barnholtz-Sloan JS: CBTRUS statistical report:
Primary brain and other central nervous system tumors diagnosed in
the United States in 2011-2015. Neuro Oncol. 20 (Supp l4):iv1–iv86.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vadgaonkar R, Epari S, Chinnaswamy G,
Krishnatry R, Tonse R, Gupta T and Jalali R: Distinct demographic
profile and molecular markers of primary CNS tumor in 1873
adolescent and young adult patient population. Childs Nerv Syst.
34:1489–1495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Husson O, Zebrack B, Block R, Embry L,
Aguilar C, Hayes-Lattin B and Cole S: Personality traits and
health-related quality of life among adolescent and young adult
cancer patients: The role of psychological distress. J Adolesc
Young Adult Oncol. 6:358–362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chandra V, Das T, Gulati P, Biswas NK,
Rote S, Chatterjee U, Ghosh SN, Deb S, Saha SK, Chowdhury AK, et
al: Hedgehog signaling pathway is active in GBM with GLI1 mRNA
expression showing a single continuous distribution rather than
discrete high/low clusters. PLoS One. 10:e01163902015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Azzi S, Treps L, Leclair HM, Ngo HM,
Harford-Wright E and Gavard J: Desert Hedgehog/Patch2 axis
contributes to vascular permeability and angiogenesis in
glioblastoma. Front Pharmacol. 6:2812015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takezaki T, Hide T, Takanaga H, Nakamura
H, Kuratsu J and Kondo T: Essential role of the Hedgehog signaling
pathway in human glioma-initiating cells. Cancer Sci.
102:1306–1312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang J, Guo C, Li J, Liang Z, Wang Y, Yu
A, Liu R, Guo Y, Chen J and Huang S: EN1 regulates cell growth and
proliferation in human glioma cells via Hedgehog signaling. Int J
Mol Sci. 23:11232022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chang L, Zhang P, Zhao D, Liu H, Wang Q,
Li C, Du W, Liu X, Zhang H, Zhang Z and Jiang C: The Hedgehog
antagonist HHIP as a favorable prognosticator in glioblastoma.
Tumour Biol. 37:3979–3986. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Infante P, Mori M, Alfonsi R, Ghirga F,
Aiello F, Toscano S, Ingallina C, Siler M, Cucchi D, Po A, et al:
Gli1/DNA interaction is a druggable target for Hedgehog-dependent
tumors. EMBO J. 34:200–217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 World Health Organization
classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cancer Genome Atlas Research Network, .
Comprehensive genomic characterization defines human glioblastoma
genes and core pathways. Nature. 455:1061–1068. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Parsons DW, Jones S, Zhang X, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Siu IM, Gallia GL, et
al: An integrated genomic analysis of human glioblastoma
multiforme. Science. 321:1807–1812. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ceccarelli M, Barthel FP, Malta TM,
Sabedot TS, Salama SR, Murray BA, Morozova O, Newton Y, Radenbaugh
A, Pagnotta SM, et al: Molecular profiling reveals biologically
discrete subsets and pathways of progression in diffuse glioma.
Cell. 164:550–563. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Q, Hu B, Hu X, Kim H, Squatrito M,
Scarpace L, deCarvalho AC, Lyu S, Li P, Li Y, et al: Tumor
evolution of glioma-intrinsic gene expression subtypes associates
with immunological changes in the microenvironment. Cancer Cell.
32:42–56.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sturm D, Bender S, Jones DT, Lichter P,
Grill J, Becher O, Hawkins C, Majewski J, Jones C, Costello JF, et
al: Paediatric and adult glioblastoma: Multiform (epi)genomic
culprits emerge. Nat Rev Cancer. 14:92–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Capper D, Jones DTW, Sill M, Hovestadt V,
Schrimpf D, Sturm D, Koelsche C, Sahm F, Chavez L, Reuss DE, et al:
DNA methylation-based classification of central nervous system
tumours. Nature. 555:469–474. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Torrisi F, Alberghina C, D'Aprile S,
Pavone AM, Longhitano L, Giallongo S, Tibullo D, Di Rosa M, Zappalà
A, Cammarata FP, et al: The hallmarks of glioblastoma:
Heterogeneity, intercellular crosstalk and molecular signature of
invasiveness and progression. Biomedicines. 10:8062022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ostrom QT, Cioffi G, Gittleman H, Patil N,
Waite K, Kruchko C and Barnholtz-Sloan JS: CBTRUS statistical
report: Primary brain and other central nervous system tumors
diagnosed in the United States in 2012-2016. Neuro Oncol 21 (Suppl
5):v1-v100; 2019, View Article : Google Scholar
|
|
20
|
Roa W, Brasher PM, Bauman G, Anthes M,
Bruera E, Chan A, Fisher B, Fulton D, Gulavita S, Hao C, et al:
Abbreviated course of radiation therapy in older patients with
glioblastoma multiforme: A prospective randomized clinical trial. J
Clin Oncol. 22:1583–1588. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Suchorska B, Weller M, Tabatabai G, Senft
C, Hau P, Sabel MC, Herrlinger U, Ketter R, Schlegel U, Marosi C,
et al: Complete resection of contrast-enhancing tumor volume is
associated with improved survival in recurrent glioblastoma-results
from the DIRECTOR trial. Neuro Oncol. 18:549–556. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stupp R, Wong ET, Kanner AA, Steinberg D,
Engelhard H, Heidecke V, Kirson ED, Taillibert S, Liebermann F,
Dbalý V, et al: NovoTTF-100A versus physician's choice chemotherapy
in recurrent glioblastoma: A randomised phase III trial of a novel
treatment modality. Eur J Cancer. 48:2192–2202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Höing S, Yeh TY, Baumann M, Martinez NE,
Habenberger P, Kremer L, Drexler HCA, Küchler P, Reinhardt P,
Choidas A, et al: Dynarrestin, a novel inhibitor of cytoplasmic
dynein. Cell Chem Biol. 25:357–369.e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rocha CRR, Reily Rocha A, Molina Silva M,
Rodrigues Gomes L, Teatin Latancia M, Andrade Tomaz M, de Souza I,
Karolynne Seregni Monteiro L and Menck CFM: Revealing temozolomide
resistance mechanisms via genome-wide CRISPR libraries. Cells.
9:25732020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ebrahimi A, Larijani L, Moradi A and
Ebrahimi MR: Hedgehog signalling pathway: Carcinogenesis and
targeted therapy. Iran J Cancer Prev. 6:36–43. 2013.PubMed/NCBI
|
|
26
|
Jin X, Jeon HM, Jin X, Kim EJ, Yin J, Jeon
HY, Sohn YW, Oh SY, Kim JK, Kim SH, et al: The ID1-CULLIN3 axis
regulates intracellular SHH and WNT signaling in glioblastoma stem
cells. Cell Rep. 16:1629–1641. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huynh DL, Koh H, Chandimali N, Zhang JJ,
Kim N, Kang TY, Ghosh M, Gera M, Park YH, Kwon T and Jeong DK:
BRM270 inhibits the proliferation of CD44 positive pancreatic
ductal adenocarcinoma cells via downregulation of sonic Hedgehog
signaling. Evid Based Complement Alternat Med. 2019:86204692019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Marigo V and Tabin CJ: Regulation of
patched by sonic Hedgehog in the developing neural tube. Proc Natl
Acad Sci USA. 93:9346–9351. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Plotnikova OV, Golemis EA and Pugacheva
EN: Cell cycle-dependent ciliogenesis and cancer. Cancer Res.
68:2058–2061. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mastronardi FG, Dimitroulakos J,
Kamel-Reid S and Manoukian AS: Co-localization of patched and
activated sonic Hedgehog to lysosomes in neurons. Neuroreport.
11:581–585. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rubin LL and de Sauvage FJ: Targeting the
Hedgehog pathway in cancer. Nat Rev Drug Discov. 5:1026–1033. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alexandre C, Jacinto A and Ingham PW:
Transcriptional activation of Hedgehog target genes in Drosophila
is mediated directly by the cubitus interruptus protein, a member
of the GLI family of zinc finger DNA-binding proteins. Genes Dev.
10:2003–2013. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pan Y, Bai CB, Joyner AL and Wang B: Sonic
Hedgehog signaling regulates Gli2 transcriptional activity by
suppressing its processing and degradation. Mol Cell Biol.
26:3365–3377. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang B, Fallon JF and Beachy PA:
Hedgehog-regulated processing of Gli3 produces an
anterior/posterior repressor gradient in the developing vertebrate
limb. Cell. 100:423–434. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Niewiadomski P, Kong JH, Ahrends R, Ma Y,
Humke EW, Khan S, Teruel MN, Novitch BG and Rohatgi R: Gli protein
activity is controlled by multisite phosphorylation in vertebrate
Hedgehog signaling. Cell Rep. 6:168–181. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gonnissen A, Isebaert S and Haustermans K:
Targeting the Hedgehog signaling pathway in cancer: Beyond
smoothened. Oncotarget. 6:13899–13913. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Skoda AM, Simovic D, Karin V, Kardum V,
Vranic S and Serman L: The role of the Hedgehog signaling pathway
in cancer: A comprehensive review. Bosn J Basic Med Sci. 18:8–20.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sabol M, Trnski D, Musani V, Ozretić P and
Levanat S: Role of GLI transcription factors in pathogenesis and
their potential as new therapeutic targets. Int J Mol Sci.
19:25622018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Endoh-Yamagami S, Evangelista M, Wilson D,
Wen X, Theunissen JW, Phamluong K, Davis M, Scales SJ, Solloway MJ,
de Sauvage FJ and Peterson AS: The mammalian Cos2 homolog Kif7
plays an essential role in modulating Hh signal transduction during
development. Curr Biol. 19:1320–1326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Denef N, Neubüser D, Perez L and Cohen SM:
Hedgehog induces opposite changes in turnover and subcellular
localization of patched and smoothened. Cell. 102:521–531. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tschaikner P, Enzler F, Torres-Quesada O,
Aanstad P and Stefan E: Hedgehog and Gpr161: Regulating cAMP
signaling in the primary cilium. Cells. 9:1182020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Price MA and Kalderon D: Proteolysis of
the Hedgehog signaling effector cubitus interruptus requires
phosphorylation by glycogen synthase kinase 3 and casein kinase 1.
Cell. 108:823–835. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Méthot N and Basler K: Suppressor of fused
opposes Hedgehog signal transduction by impeding nuclear
accumulation of the activator form of cubitus interruptus.
Development. 127:4001–4010. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Brennan D, Chen X, Cheng L, Mahoney M and
Riobo NA: Noncanonical Hedgehog signaling. Vitam Horm. 88:55–72.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Robbins DJ, Fei DL and Riobo NA: The
Hedgehog signal transduction network. Sci Signal. 5:re62012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Awasthi A, Woolley AG, Lecomte FJ, Hung N,
Baguley BC, Wilbanks SM, Jeffs AR and Tyndall JD: Variable
expression of GLIPR1 correlates with invasive potential in melanoma
cells. Front Oncol. 3:2252013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang K, Pan L, Che X, Cui D and Li C:
Sonic Hedgehog/GLI1 signaling pathway inhibition restricts cell
migration and invasion in human gliomas. Neurol Res. 32:975–980.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lo HW, Zhu H, Cao X, Aldrich A and
Ali-Osman F: A novel splice variant of GLI1 that promotes
glioblastoma cell migration and invasion. Cancer Res. 69:6790–6798.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Reifenberger J, Wolter M, Weber RG,
Megahed M, Ruzicka T, Lichter P and Reifenberger G: Missense
mutations in SMOH in sporadic basal cell carcinomas of the skin and
primitive neuroectodermal tumors of the central nervous system.
Cancer Res. 58:1798–1803. 1998.PubMed/NCBI
|
|
50
|
Lee Y, Miller HL, Jensen P, Hernan R,
Connelly M, Wetmore C, Zindy F, Roussel MF, Curran T, Gilbertson RJ
and McKinnon PJ: A molecular fingerprint for medulloblastoma.
Cancer Res. 63:5428–5437. 2003.PubMed/NCBI
|
|
51
|
Lim CB, Prêle CM, Cheah HM, Cheng YY,
Klebe S, Reid G, Watkins DN, Baltic S, Thompson PJ and Mutsaers SE:
Mutational analysis of Hedgehog signaling pathway genes in human
malignant mesothelioma. PLoS One. 8:e666852013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Berman DM, Karhadkar SS, Maitra A, Montes
De Oca R, Gerstenblith MR, Briggs K, Parker AR, Shimada Y, Eshleman
JR, Watkins DN and Beachy PA: Widespread requirement for Hedgehog
ligand stimulation in growth of digestive tract tumours. Nature.
425:846–851. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Noman AS, Uddin M, Rahman MZ, Nayeem MJ,
Alam SS, Khatun Z, Wahiduzzaman M, Sultana A, Rahman ML, Ali MY, et
al: Overexpression of sonic Hedgehog in the triple negative breast
cancer: Clinicopathological characteristics of high burden breast
cancer patients from Bangladesh. Sci Rep. 6:188302016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Watkins DN, Berman DM, Burkholder SG, Wang
B, Beachy PA and Baylin SB: Hedgehog signalling within airway
epithelial progenitors and in small-cell lung cancer. Nature.
422:313–317. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Riobo-Del Galdo NA, Lara Montero Á and
Wertheimer EV: Role of Hedgehog signaling in breast cancer:
Pathogenesis and therapeutics. Cells. 8:3752019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hanahan D and Coussens LM: Accessories to
the crime: Functions of cells recruited to the tumor
microenvironment. Cancer Cell. 21:309–322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Petersen OW, Rønnov-Jessen L, Howlett AR
and Bissell MJ: Interaction with basement membrane serves to
rapidly distinguish growth and differentiation pattern of normal
and malignant human breast epithelial cells. Proc Natl Acad Sci
USA. 89:9064–9068. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Martinez-Outschoorn U, Sotgia F and
Lisanti MP: Tumor microenvironment and metabolic synergy in breast
cancers: Critical importance of mitochondrial fuels and function.
Semin Oncol. 41:195–216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Martinez-Outschoorn UE, Lin Z, Ko YH,
Goldberg AF, Flomenberg N, Wang C, Pavlides S, Pestell RG, Howell
A, Sotgia F and Lisanti MP: Understanding the metabolic basis of
drug resistance: Therapeutic induction of the Warburg effect kills
cancer cells. Cell Cycle. 10:2521–2528. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chang L, Zhao D, Liu HB, Wang QS, Zhang P,
Li CL, Du WZ, Wang HJ, Liu X, Zhang ZR and Jiang CL: Activation of
sonic Hedgehog signaling enhances cell migration and invasion by
induction of matrix metalloproteinase-2 and −9 via the
phosphoinositide-3 kinase/AKT signaling pathway in glioblastoma.
Mol Med Rep. 12:6702–6710. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Torrisi F, Alberghina C, Lo Furno D,
Zappalà A, Valable S, Li Volti G, Tibullo D, Vicario N and Parenti
R: Connexin 43 and Sonic Hedgehog pathway interplay in glioblastoma
cell proliferation and migration. Biology (Basel).
10:7672021.PubMed/NCBI
|
|
62
|
Cherepanov SA, Cherepanova KI, Grinenko
NF, Antonova OM and Chekhonin VP: Effect of Hedgehog signaling
pathway activation on proliferation of high-grade gliomas. Bull Exp
Biol Med. 161:674–678. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Rowitch DH, S-Jacques B, Lee SM, Flax JD,
Snyder EY and McMahon AP: Sonic Hedgehog regulates proliferation
and inhibits differentiation of CNS precursor cells. J Neurosci.
19:8954–8965. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Jiang K, Wang YP, Wang XD, Hui XB, Ding
LS, Liu J and Liu D: Fms related tyrosine kinase 1 (Flt1) functions
as an oncogene and regulates glioblastoma cell metastasis by
regulating sonic Hedgehog signaling. Am J Cancer Res. 7:1164–1176.
2017.PubMed/NCBI
|
|
65
|
Hamerlik P, Lathia JD, Rasmussen R, Wu Q,
Bartkova J, Lee M, Moudry P, Bartek J Jr, Fischer W, Lukas J, et
al: Autocrine VEGF-VEGFR2-neuropilin-1 signaling promotes glioma
stem-like cell viability and tumor growth. J Exp Med. 209:507–520.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kloepper J, Riedemann L, Amoozgar Z, Seano
G, Susek K, Yu V, Dalvie N, Amelung RL, Datta M, Song JW, et al:
Ang-2/VEGF bispecific antibody reprograms macrophages and resident
microglia to anti-tumor phenotype and prolongs glioblastoma
survival. Proc Natl Acad Sci USA. 113:4476–4481. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chen L, Wang L, Qin J and Wei DS: CtBP2
interacts with ZBTB18 to promote malignancy of glioblastoma. Life
Sci. 262:1184772020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Bensalma S, Turpault S, Balandre AC, De
Boisvilliers M, Gaillard A, Chadéneau C and Muller JM: PKA at a
cross-road of signaling pathways involved in the regulation of
glioblastoma migration and invasion by the neuropeptides VIP and
PACAP. Cancers (Basel). 11:1232019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Henao-Restrepo J, Caro-Urrego YA,
Barrera-Arenas LM, Arango-Viana JC and Bermudez-Munoz M: Expression
of activator proteins of SHH/GLI and PI3K/Akt/mTORC1 signaling
pathways in human gliomas is associated with high grade tumors. Exp
Mol Pathol. 122:1046732021. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kinzler KW, Bigner SH, Bigner DD, Trent
JM, Law ML, O'Brien SJ, Wong J and Vogelstein B: Identification of
an amplified, highly expressed gene in a human glioma. Science.
236:70–73. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Bigner SH, Wong AJ, Mark J, Muhlbaier LH,
Kinzler KW, Vogelstein B and Bigner DD: Relationship between gene
amplification and chromosomal deviations in malignant human
gliomas. Cancer Genet Cytogenet. 29:165–170. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
ten Haaf A, Bektas N, von Serenyi S, Losen
I, Arweiler EC, Hartmann A, Knuchel R and Dahl E: Expression of the
glioma-associated oncogene homolog (GLI) 1 in human breast cancer
is associated with unfavourable overall survival. BMC Cancer.
9:2982009. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zhou A, Lin K, Zhang S, Ma L, Xue J,
Morris SA, Aldape KD and Huang S: Gli1-induced deubiquitinase USP48
aids glioblastoma tumorigenesis by stabilizing Gli1. EMBO Rep.
18:1318–1330. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Kim Y, Do IG, Hong M and Suh YL: Negative
prognostic effect of low nuclear GLI1 expression in glioblastomas.
J Neurooncol. 133:69–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Xue J, Zhou A, Tan C, Wu Y, Lee HT, Li W,
Xie K and Huang S: Forkhead box M1 is essential for nuclear
localization of glioma-associated oncogene homolog 1 in
glioblastoma multiforme cells by promoting importin-7 expression. J
Biol Chem. 290:18662–18670. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhu H, Carpenter RL, Han W and Lo HW: The
GLI1 splice variant TGLI1 promotes glioblastoma angiogenesis and
growth. Cancer Lett. 343:51–61. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang Z, Zheng X, Luan Y and Liu Y, Li X,
Liu C, Lu H, Chen X and Liu Y: Activity of metabotropic glutamate
receptor 4 suppresses proliferation and promotes apoptosis with
inhibition of Gli-1 in human glioblastoma cells. Front Neurosci.
12:3202018. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Sargazi ML, Juybari KB, Tarzi ME,
Amirkhosravi A, Nematollahi MH, Mirzamohammdi S, Mehrbani M and
Mehrabani M and Mehrabani M: Naringenin attenuates cell viability
and migration of C6 glioblastoma cell line: A possible role of
Hedgehog signaling pathway. Mol Biol Rep. 48:6413–6421. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chantaravisoot N, Wongkongkathep P, Loo
AJ, Mischel SP and Tamanoi F: Significance of filamin A in mTORC2
function in glioblastoma. Mol Cancer. 14:e1272015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Maiti S, Mondal S, Satyavarapu EM and
Mandal C: mTORC2 regulates Hedgehog pathway activity by promoting
stability to Gli2 protein and its nuclear translocation. Cell Death
Dis. 8:e29262017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Tanigawa S, Fujita M, Moyama C, Ando S, Ii
H, Kojima Y, Fujishita T, Aoki M, Takeuchi H, Yamanaka T, et al:
Inhibition of Gli2 suppresses tumorigenicity in glioblastoma stem
cells derived from a de novo murine brain cancer model. Cancer Gene
Ther. 28:1339–1352. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zaphiropoulos PG, Undén AB, Rahnama F,
Hollingsworth RE and Toftgård R: PTCH2, a novel human patched gene,
undergoing alternative splicing and up-regulated in basal cell
carcinomas. Cancer Res. 59:787–792. 1999.PubMed/NCBI
|
|
83
|
Larsson NG, Wang J, Wilhelmsson H, Oldfors
A, Rustin P, Lewandoski M, Barsh GS and Clayton DA: Mitochondrial
transcription factor A is necessary for mtDNA maintenance and
embryogenesis in mice. Nat Genet. 18:231–236. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Rudin CM, Hann CL, Laterra J, Yauch RL,
Callahan CA, Fu L, Holcomb T, Stinson J, Gould SE, Coleman B, et
al: Treatment of medulloblastoma with Hedgehog pathway inhibitor
GDC-0449. N Engl J Med. 361:1173–1178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Scales SJ and de Sauvage FJ: Mechanisms of
Hedgehog pathway activation in cancer and implications for therapy.
Trends Pharmacol Sci. 30:303–312. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Marjanovic Vicentic J, Drakulic D, Garcia
I, Vukovic V, Aldaz P, Puskas N, Nikolic I, Tasic G, Raicevic S,
Garros-Regulez L, et al: SOX3 can promote the malignant behavior of
glioblastoma cells. Cell Oncol (Dordr). 42:41–54. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Saito A, Kanemoto S, Zhang Y, Asada R,
Hino K and Imaizumi K: Chondrocyte proliferation regulated by
secreted luminal domain of ER stress transducer BBF2H7/CREB3L2. Mol
Cell. 53:127–139. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Iwamoto H, Matsuhisa K, Saito A, Kanemoto
S, Asada R, Hino K, Takai T, Cui M, Cui X, Kaneko M, et al:
Promotion of cancer cell proliferation by cleaved and secreted
luminal domains of ER stress transducer BBF2H7. PLoS One.
10:e01259822015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Guen VJ, Chavarria TE, Kröger C, Ye X,
Weinberg RA and Lees JA: EMT programs promote basal mammary stem
cell and tumor-initiating cell stemness by inducing primary
ciliogenesis and Hedgehog signaling. Proc Natl Acad Sci USA.
114:E10532–E10539. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Brandner S: Nanog, Gli, and p53: A new
network of stemness in development and cancer. EMBO J.
29:2475–2476. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Rich JN and Eyler CE: Cancer stem cells in
brain tumor biology. Cold Spring Harb Symp Quant Biol. 73:411–420.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
de Groot JF, Fuller G, Kumar AJ, Piao Y,
Eterovic K, Ji Y and Conrad CA: Tumor invasion after treatment of
glioblastoma with bevacizumab: Radiographic and pathologic
correlation in humans and mice. Neuro Oncol. 12:233–242. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Han B, Wang R, Chen Y, Meng X, Wu P, Li Z,
Duan C, Li Q, Li Y, Zhao S, et al: QKI deficiency maintains glioma
stem cell stemness by activating the SHH/GLI1 signaling pathway.
Cell Oncol (Dordr). 42:801–813. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Yuan Y, Zhang M, Yan G, Ma Q, Yan Z, Wang
L, Yang K and Guo D: Nanog promotes stem-like traits of
glioblastoma cells. Front Biosci (Landmark Ed). 26:552–565. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Silvestri I, Testa F, Zappasodi R, Cairo
CW, Zhang Y, Lupo B, Galli R, Di Nicola M, Venerando B and Tringali
C: Sialidase NEU4 is involved in glioblastoma stem cell survival.
Cell Death Dis. 5:e13812014. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Kundu S, Nandhu MS, Longo SL, Longo JA,
Rai S, Chin LS, Richardson TE and Viapiano MS: The scaffolding
protein DLG5 promotes glioblastoma growth by controlling sonic
Hedgehog signaling in tumor stem cells. Neuro Oncol: noac001.
2022.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Park MG, Seo S, Ham SW, Choi SH and Kim H:
Dihydropyrimidinase-related protein 5 controls glioblastoma stem
cell characteristics as a biomarker of proneural-subtype
glioblastoma stem cells. Oncol Lett. 20:1153–1162. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Mondal S, Bhattacharya K and Mandal C:
Nutritional stress reprograms dedifferention in glioblastoma
multiforme driven by PTEN/Wnt/Hedgehog axis: A stochastic model of
cancer stem cells. Cell Death Discov. 4:1102018. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Yan GN, Yang L, Lv YF, Shi Y, Shen LL, Yao
XH, Guo QN, Zhang P, Cui YH, Zhang X, et al: Endothelial cells
promote stem-like phenotype of glioma cells through activating the
Hedgehog pathway. J Pathol. 234:11–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Sekulic A, Migden MR, Oro AE, Dirix L,
Lewis KD, Hainsworth JD, Solomon JA, Yoo S, Arron ST, Friedlander
PA, et al: Efficacy and safety of vismodegib in advanced basal-cell
carcinoma. N Engl J Med. 366:2171–2179. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Robinson GW, Orr BA, Wu G, Gururangan S,
Lin T, Qaddoumi I, Packer RJ, Goldman S, Prados MD, Desjardins A,
et al: Vismodegib exerts targeted efficacy against recurrent sonic
Hedgehog-subgroup medulloblastoma: Results from phase II pediatric
brain tumor consortium studies PBTC-025B and PBTC-032. J Clin
Oncol. 33:2646–2654. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Wong R, Turlova E, Feng ZP, Rutka JT and
Sun HS: Activation of TRPM7 by naltriben enhances migration and
invasion of glioblastoma cells. Oncotarget. 8:11239–11248. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Shojaei F, Lee JH, Simmons BH, Wong A,
Esparza CO, Plumlee PA, Feng J, Stewart AE, Hu-Lowe DD and
Christensen JG: HGF/c-Met acts as an alternative angiogenic pathway
in sunitinib-resistant tumors. Cancer Res. 70:10090–10100. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Lu KV, Chang JP, Parachoniak CA, Pandika
MM, Aghi MK, Meyronet D, Isachenko N, Fouse SD, Phillips JJ,
Cheresh DA, et al: VEGF inhibits tumor cell invasion and
mesenchymal transition through a MET/VEGFR2 complex. Cancer Cell.
22:21–35. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Li L and Li W: Epithelial-mesenchymal
transition in human cancer: Comprehensive reprogramming of
metabolism, epigenetics, and differentiation. Pharmacol Ther.
150:33–46. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Chaffer CL, San Juan BP, Lim E and
Weinberg RA: EMT, cell plasticity and metastasis. Cancer Metastasis
Rev. 35:645–654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Tang H, Massi D, Hemmings BA, Mandalà M,
Hu Z, Wicki A and Xue G: AKT-ions with a TWIST between EMT and MET.
Oncotarget. 7:62767–62777. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Yeung KT and Yang J:
Epithelial-mesenchymal transition in tumor metastasis. Mol Oncol.
11:28–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Urdiciain A, Erausquin E, Meléndez B, Rey
JA, Idoate MA and Castresana JS: Tubastatin A, an inhibitor of
HDAC6, enhances temozolomide-induced apoptosis and reverses the
malignant phenotype of glioblastoma cells. Int J Oncol.
54:1797–1808. 2019.PubMed/NCBI
|
|
110
|
Feng X, Yao J, Gao X, Jing Y, Kang T,
Jiang D, Jiang T, Feng J, Zhu Q, Jiang X and Chen J:
Multi-targeting peptide-functionalized nanoparticles recognized
vasculogenic mimicry, tumor neovasculature, and glioma cells for
enhanced anti-glioma therapy. ACS Appl Mater Interfaces.
7:27885–27899. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Kast RE, Skuli N, Karpel-Massler G,
Frosina G, Ryken T and Halatsch ME: Blocking
epithelial-to-mesenchymal transition in glioblastoma with a sextet
of repurposed drugs: The EIS regimen. Oncotarget. 8:60727–60749.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Li M, Zhang J, Zhou H and Xiang R: Primary
cilia-related pathways moderate the development and therapy
resistance of glioblastoma. Front Oncol. 11:7189952021. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Jenks AD, Vyse S, Wong JP, Kostaras E,
Keller D, Burgoyne T, Shoemark A, Tsalikis A, de la Roche M,
Michaelis M, et al: Primary cilia mediate diverse kinase inhibitor
resistance mechanisms in cancer. Cell Rep. 23:3042–3055. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Carpenter RL and Lo HW: Hedgehog pathway
and GLI1 isoforms in human cancer. Discov Med. 13:105–113.
2012.PubMed/NCBI
|
|
115
|
Honorato JR, Hauser-Davis RA, Saggioro EM,
Correia FV, Sales-Junior SF, Soares LOS, Lima LDR, Moura-Neto V,
Lopes GPF and Spohr TCLS: Role of sonic Hedgehog signaling in cell
cycle, oxidative stress, and autophagy of temozolomide resistant
glioblastoma. J Cell Physiol. 235:3798–3814. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Melamed JR, Morgan JT, Ioele SA, Gleghorn
JP, Sims-Mourtada J and Day ES: Investigating the role of
Hedgehog/GLI1 signaling in glioblastoma cell response to
temozolomide. Oncotarget. 9:27000–27015. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Lai IC, Shih PH, Yao CJ, Yeh CT, Wang-Peng
J, Lui TN, Chuang SE, Hu TS, Lai TY and Lai GM: Elimination of
cancer stem-like cells and potentiation of temozolomide sensitivity
by honokiol in glioblastoma multiforme cells. PLoS One.
10:e01148302015. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Sarkaria JN, Kitange GJ, James CD, Plummer
R, Calvert H, Weller M and Wick W: Mechanisms of chemoresistance to
alkylating agents in malignant glioma. Clin Cancer Res.
14:2900–2908. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Munoz JL, Rodriguez-Cruz V, Walker ND,
Greco SJ and Rameshwar P: Temozolomide resistance and tumor
recurrence: Halting the Hedgehog. Cancer Cell Microenviron.
2:e7472015.PubMed/NCBI
|
|
120
|
Hung HC, Liu CC, Chuang JY, Su CL and Gean
PW: Inhibition of sonic Hedgehog signaling suppresses glioma
stem-like cells likely through inducing autophagic cell death.
Front Oncol. 10:12332020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Chen J, Fu X, Wan Y, Wang Z, Jiang D and
Shi L: miR-125b inhibitor enhance the chemosensitivity of
glioblastoma stem cells to temozolomide by targeting Bak1. Tumour
Biol. 35:6293–6302. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Melamed JR, Ioele SA, Hannum AJ, Ullman VM
and Day ES: Polyethylenimine-spherical nucleic acid nanoparticles
against Gli1 reduce the chemoresistance and stemness of
glioblastoma cells. Mol Pharm. 15:5135–5145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Ding D, Lim KS and Eberhart CG: Arsenic
trioxide inhibits Hedgehog, notch and stem cell properties in
glioblastoma neurospheres. Acta Neuropathol Commun. 2:312014.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Wang J, Huang S, Tian R, Chen J, Gao H,
Xie C, Shan Y, Zhang Z, Gu S and Xu M: The protective autophagy
activated by GANT-61 in MYCN amplified neuroblastoma cells is
mediated by PERK. Oncotarget. 9:14413–14427. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Carballo GB, Ribeiro JH, Lopes GPF, Ferrer
VP, Dezonne RS, Pereira CM and Spohr TCLSE: GANT-61 induces
autophagy and apoptosis in glioblastoma cells despite their
heterogeneity. Cell Mol Neurobiol. 41:1227–1244. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Yin S, Du W, Wang F, Han B, Cui Y, Yang D,
Chen H, Liu D, Liu X, Zhai X and Jiang C: MicroRNA-326 sensitizes
human glioblastoma cells to curcumin via the SHH/GLI1 signaling
pathway. Cancer Biol Ther. 19:260–270. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Ji M, Zhang Z, Lin S, Wang C, Jin J, Xue
N, Xu H and Chen X: The PI3K inhibitor XH30 enhances response to
temozolomide in drug-resistant glioblastoma via the noncanonical
Hedgehog signaling pathway. Front Pharmacol. 12:7492422021.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Han L, Tang L, Jiang Z and Jiang Y:
Enhanced radiosensitization of human glioblastoma multiforme cells
with phosphorylated peptides derived from Gli2. Neuropeptides.
70:87–92. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Urdiciain A, Erausquin E, Zelaya MV, Zazpe
I, Lanciego JL, Meléndez B, Rey JA, Idoate MA, Riobo-Del Galdo NA
and Castresana JS: Silencing of histone deacetylase 6 decreases
cellular malignancy and contributes to primary cilium restoration,
epithelial-to-mesenchymal transition reversion, and autophagy
inhibition in glioblastoma cell lines. Biology (Basel).
10:4672021.PubMed/NCBI
|
|
130
|
Yang W, Liu Y, Gao R, Yu H and Sun T:
HDAC6 inhibition induces glioma stem cells differentiation and
enhances cellular radiation sensitivity through the SHH/Gli1
signaling pathway. Cancer Lett. 415:164–176. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Clement V, Sanchez P, de Tribolet N,
Radovanovic I and Ruiz i Altaba A: Hedgehog-GLI1 signaling
regulates human glioma growth, cancer stem cell self-renewal, and
tumorigenicity. Curr Biol. 17:165–172. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Balbous A, Renoux B, Cortes U, Milin S,
Guilloteau K, Legigan T, Rivet P, Boissonnade O, Martin S, Tripiana
C, et al: Selective release of a cyclopamine glucuronide prodrug
toward stem-like cancer cell inhibition in glioblastoma. Mol Cancer
Ther. 13:2159–2169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Bensalma S, Chadeneau C, Legigan T, Renoux
B, Gaillard A, de Boisvilliers M, Pinet-Charvet C, Papot S and
Muller JM: Evaluation of cytotoxic properties of a cyclopamine
glucuronide prodrug in rat glioblastoma cells and tumors. J Mol
Neurosci. 55:51–61. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Wang K, Chen D, Qian Z, Cui D, Gao L and
Lou M: Hedgehog/Gli1 signaling pathway regulates MGMT expression
and chemoresistance to temozolomide in human glioblastoma. Cancer
Cell Int. 17:1172017. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Carballo GB, Matias D, Ribeiro JH, Pessoa
LS, Arrais-Neto AM and Spohr TCLSE: Cyclopamine sensitizes
glioblastoma cells to temozolomide treatment through sonic Hedgehog
pathway. Life Sci. 257:1180272020. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Liu YJ, Ma YC, Zhang WJ, Yang ZZ, Liang
DS, Wu ZF and Qi XR: Combination therapy with micellarized
cyclopamine and temozolomide attenuate glioblastoma growth through
Gli1 down-regulation. Oncotarget. 8:42495–42509. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Chen J, Lv H, Hu J, Ji M, Xue N, Li C, Ma
S, Zhou Q, Lin B, Li Y, et al: CAT3, a novel agent for
medulloblastoma and glioblastoma treatment, inhibits tumor growth
by disrupting the Hedgehog signaling pathway. Cancer Lett.
381:391–403. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Ji M, Wang L, Chen J, Xue N, Wang C, Lai
F, Wang R, Yu S, Jin J and Chen X: CAT3, a prodrug of
13a(S)-3-hydroxyl-6,7-dimethoxyphenanthro[9,10-b]-indolizidine,
circumvents temozolomide-resistant glioblastoma via the Hedgehog
signaling pathway, independently of O6-methylguanine DNA
methyltransferase expression. Onco Targets Ther. 11:3671–3684.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Horrigan SK; Reproducibility Project, :
Cancer Biology: Replication study: The CD47-signal regulatory
protein alpha (SIRPa) interaction is a therapeutic target for human
solid tumors. Elife. 6:e181732017. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
von Roemeling CA, Wang Y, Qie Y, Yuan H,
Zhao H, Liu X, Yang Z, Yang M, Deng W, Bruno KA, et al: Therapeutic
modulation of phagocytosis in glioblastoma can activate both innate
and adaptive antitumour immunity. Nat Commun. 11:15082020.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Chakrabarti J, Holokai L, Syu L, Steele
NG, Chang J, Wang J, Ahmed S, Dlugosz A and Zavros Y: Hedgehog
signaling induces PD-L1 expression and tumor cell proliferation in
gastric cancer. Oncotarget. 9. 37439-37457; 2018, View Article : Google Scholar
|
|
142
|
Grund-Gröschke S, Stockmaier G and Aberger
F: Hedgehog/GLI signaling in tumor immunity-new therapeutic
opportunities and clinical implications. Cell Commun Signal.
17:1722019. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
SurveillanceEpidemiology, End Results
Program, . Cancer stat facts: Brain and other nervous system
cancer. 2019.https://seer.cancer.gov/statfacts/html/brain.htmlJuly
12–2019
|
|
144
|
Braun S, Oppermann H, Mueller A, Renner C,
Hovhannisyan A, Baran-Schmidt R, Gebhardt R, Hipkiss A, Thiery J,
Meixensberger J and Gaunitz F: Hedgehog signaling in glioblastoma
multiforme. Cancer Biol Ther. 13:487–495. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Filbin MG, Dabral SK, Pazyra-Murphy MF,
Ramkissoon S, Kung AL, Pak E, Chung J, Theisen MA, Sun Y,
Franchetti Y, et al: Coordinate activation of Shh and PI3K
signaling in PTEN-deficient glioblastoma: New therapeutic
opportunities. Nat Med. 19:1518–1523. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Lan X, Jörg DJ, Cavalli FMG, Richards LM,
Nguyen LV, Vanner RJ, Guilhamon P, Lee L, Kushida MM, Pellacani D,
et al: Fate mapping of human glioblastoma reveals an invariant stem
cell hierarchy. Nature. 549:227–232. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Han S, Zhang C, Li Q, Dong J, Liu Y, Huang
Y, Jiang T and Wu A: Tumour-infiltrating CD4(+) and CD8(+)
lymphocytes as predictors of clinical outcome in glioma. Br J
Cancer. 110:2560–2568. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Heimberger AB, Abou-Ghazal M, Reina-Ortiz
C, Yang DS, Sun W, Qiao W, Hiraoka N and Fuller GN: Incidence and
prognostic impact of FoxP3+ regulatory T cells in human gliomas.
Clin Cancer Res. 14:5166–5172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Wainwright DA, Chang AL, Dey M,
Balyasnikova IV, Kim CK, Tobias A, Cheng Y, Kim JW, Qiao J, Zhang
L, et al: Durable therapeutic efficacy utilizing combinatorial
blockade against IDO, CTLA-4, and PD-L1 in mice with brain tumors.
Clin Cancer Res. 20:5290–5301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Heimberger AB, Sun W, Hussain SF, Dey M,
Crutcher L, Aldape K, Gilbert M, Hassenbusch SJ, Sawaya R,
Schmittling B, et al: Immunological responses in a patient with
glioblastoma multiforme treated with sequential courses of
temozolomide and immunotherapy: Case study. Neuro Oncol. 10:98–103.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Jackson CM, Choi J and Lim M: Mechanisms
of immunotherapy resistance: Lessons from glioblastoma. Nat
Immunol. 20:1100–1109. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Sampson JH, Gunn MD, Fecci PE and Ashley
DM: Brain immunology and immunotherapy in brain tumours. Nat Rev
Cancer. 20:12–25. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Reardon DA, Brandes AA, Omuro A,
Mulholland P, Lim M, Wick A, Baehring J, Ahluwalia MS, Roth P, Bähr
O, et al: Effect of nivolumab vs bevacizumab in patients with
recurrent glioblastoma: The CheckMate 143 phase 3 randomized
clinical trial. JAMA Oncol. 6:1003–1010. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Chiocca EA, Nassiri F, Wang J, Peruzzi P
and Zadeh G: Viral and other therapies for recurrent glioblastoma:
Is a 24-month durable response unusual? Neuro Oncol. 21:14–25.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Iorgulescu JB, Reardon DA, Chiocca EA and
Wu CJ: Immunotherapy for glioblastoma: Going viral. Nat Med.
24:1094–1096. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Brown CE, Alizadeh D, Starr R, Weng L,
Wagner JR, Naranjo A, Ostberg JR, Blanchard MS, Kilpatrick J,
Simpson J, et al: Regression of glioblastoma after chimeric antigen
receptor T-cell therapy. N Engl J Med. 375:2561–2569. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Lassman A, Pugh S, Wang T, Aldape K, Gan
H, Preusser M, Vogelbaum M, Sulman E, Won M, Zhang P, et al:
ACTR-21. A randomized, double-blind, placebo-controlled phase 3
trial of depatuxizumab mafodotin (ABT-414) in epidermal growth
factor receptor (EGFR) amplified (AMP) newly diagnosed glioblastoma
(nGBM). Neuro Oncol. 21 (Suppl 6):vi172019. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Ma DJ, Galanis E, Anderson SK, Schiff D,
Kaufmann TJ, Peller PJ, Giannini C, Brown PD, Uhm JH, McGraw S, et
al: A phase II trial of everolimus, temozolomide, and radiotherapy
in patients with newly diagnosed glioblastoma: NCCTG N057K. Neuro
Oncol. 17:1261–1269. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Wick W, Gorlia T, Bady P, Platten M, van
den Bent MJ, Taphoorn MJ, Steuve J, Brandes AA, Hamou MF, Wick A,
et al: Phase II study of radiotherapy and temsirolimus versus
radiochemotherapy with temozolomide in patients with newly
diagnosed glioblastoma without MGMT promoter hypermethylation
(EORTC 26082). Clin Cancer Res. 22:4797–4806. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Lombardi G, De Salvo GL, Brandes AA, Eoli
M, Rudà R, Faedi M, Lolli I, Pace A, Daniele B, Pasqualetti F, et
al: Regorafenib compared with lomustine in patients with relapsed
glioblastoma (REGOMA): A multicentre, open-label, randomised,
controlled, phase 2 trial. Lancet Oncol. 20:110–119. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Kim EJ, Sahai V, Abel EV, Griffith KA,
Greenson JK, Takebe N, Khan GN, Blau JL, Craig R, Balis UG, et al:
Pilot clinical trial of Hedgehog pathway inhibitor GDC-0449
(vismodegib) in combination with gemcitabine in patients with
metastatic pancreatic adenocarcinoma. Clin Cancer Res.
20:5937–5945. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Berlin J, Bendell JC, Hart LL, Firdaus I,
Gore I, Hermann RC, Mulcahy MF, Zalupski MM, Mackey HM, Yauch RL,
et al: A randomized phase II trial of vismodegib versus placebo
with FOLFOX or FOLFIRI and bevacizumab in patients with previously
untreated metastatic colorectal cancer. Clin Cancer Res.
19:258–267. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Belani CP, Dahlberg SE, Rudin CM, Fleisher
M, Chen HX, Takebe N, Velasco MR Jr, Tester WJ, Sturtz K, Hann CL,
et al: Vismodegib or cixutumumab in combination with standard
chemotherapy for patients with extensive-stage small cell lung
cancer: A trial of the ECOG-ACRIN cancer research group (E1508).
Cancer. 122:2371–2378. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Zubčić V, Rinčić N, Kurtović M, Trnski D,
Musani V, Ozretić P, Levanat S, Leović D and Sabol M: GANT61 and
lithium chloride inhibit the growth of head and neck cancer cell
lines through the regulation of GLI3 processing by GSK3β. Int J Mol
Sci. 21:64102020. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Miyazaki Y, Matsubara S, Ding Q, Tsukasa
K, Yoshimitsu M, Kosai K and Takao S: Efficient elimination of
pancreatic cancer stem cells by Hedgehog/GLI inhibitor GANT61 in
combination with mTOR inhibition. Mol Cancer. 15:492016. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Pietrobono S, Gagliardi S and Stecca B:
Non-canonical Hedgehog signaling pathway in cancer: Activation of
GLI transcription factors beyond smoothened. Front Genet.
10:5562019. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Kim J, Lee JJ, Kim J, Gardner D and Beachy
PA: Arsenic antagonizes the Hedgehog pathway by preventing ciliary
accumulation and reducing stability of the Gli2 transcriptional
effector. Proc Natl Acad Sci USA. 107:13432–13437. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Yanai K, Nakamura M, Akiyoshi T, Nagai S,
Wada J, Koga K, Noshiro H, Nagai E, Tsuneyoshi M, Tanaka M and
Katano M: Crosstalk of Hedgehog and Wnt pathways in gastric cancer.
Cancer Lett. 263:145–156. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Nanta R, Shrivastava A, Sharma J, Shankar
S and Srivastava RK: Inhibition of sonic Hedgehog and PI3K/Akt/mTOR
pathways cooperate in suppressing survival, self-renewal and
tumorigenic potential of glioblastoma-initiating cells. Mol Cell
Biochem. 454:11–23. 2019. View Article : Google Scholar : PubMed/NCBI
|