Introduction
Several epidemiological studies have reported
potential associations between chronic exposure to power frequency
(50-60 Hz) magnetic fields (MFs) and increased risk of a number of
pathologies, including amyotrophic lateral sclerosis, brain tumors
or childhood and adult leukemia (1–8).
Based primarily on epidemiological evidence on childhood leukemia,
the International Agency for Research on Cancer (IARC) classified
extremely low frequency [(ELF) MFs: 3 Hz-3 kHz)] as possible
carcinogens to humans, class 2B (9). In addition, there is in vitro
experimental evidence that provides partial support to the
epidemiological data, as it reveals that exposure to ELF MFs can
affect different cellular processes involved in cancer promotion
(10,11). While it is accepted that ELF fields
cannot directly damage the DNA molecule (12), exposure to a 50-Hz MF at flux
densities as low as 0.1-1 mT has been reported to induce changes in
DNA integrity (13–15) and it has been proposed that
alterations in genes related to DNA repair, observed in acute
leukemia patients, could be associated to chronic exposure of ELF
MFs (16). This type of evidence
has led to a number of hypotheses on potential mechanisms through
which ELF fields could indirectly affect the DNA structure
(17–20).
Tumor suppressor genes such as TP53 are involved in
various processes associated with cell division, including gene
expression regulation, cell cycle control, cell death programming
or genome stability (21). Loss of
activity of these genes can cause inability of processes
controlling cell proliferation and occasionally lead to the
development of neoplasms and to their evolution towards more
aggressive tumor processes (22).
Due to its central role in coordinating the cellular responses of a
wide range of stressors, the TP53 gene has been described as a
‘genome guardian’ (23) or a ‘cell
guardian’ (24). When DNA damage
occurs, levels of the protein encoded by TP53 rapidly increase and
the cell cycle stops at phase G1-S, allowing the cell a time span
for DNA repair systems to act (25). Such a response does not occur in
tumor cells whose TP53 gene is inactivated due to mutation,
protein-protein interaction or reaggregation (26,27).
In addition, inactivation of this tumor suppressor gene, which is a
frequent event in tumorigenesis, has also been reported to be due
to alterations in the conformation of the wild-type (wt) p53
protein that do not necessarily involve mutations (28–30).
Other factors potentially capable of compromising the functions of
p53 are the inactivation of co-activators of the wt protein, the
inactivation of downstream targets of p53 or the cytoplasmic
retention of p53. Indeed, cytoplasmic sequestration of wt p53 has
been observed in undifferentiated neuroblastoma, colon carcinoma
and breast cancer cells (31,32).
In addition to exerting the aforementioned described
transcription-dependent functions at the nuclear domain, p53
interacts with cytoplasmic proteins such as those of the Bcl-2
family, leading to permeabilization of the mitochondrial membrane
and increased apoptosis (33–35).
Moreover, in contrast to wt p53, endogenous missense mutants of p53
are unable to form complexes with endogenous Bcl-2 in human cancer
cells, which renders them unable to induce apoptosis (36).
Recently, the mitogen-activated protein kinase
(MAPK) has been reported to integrate extracellular signals related
to p53 expression and its role in cell cycle regulation (37). In previous studies by the authors,
it was demonstrated that intermittent exposure to a weak, 50-Hz MF,
in addition to increasing free radical levels in NB69 human
neuroblastoma cells, promoted the activation of the MAPK-ERK1/2 and
-p38 transduction pathways, as well as that of the EGF receptor.
Such activations, some of which are stress-dependent, induce cell
cycle changes that lead to a significant increase in the
proliferation of NB69 cells (38–40).
Based on these data, the present study investigated whether in
vitro exposure to a 50-Hz, 100-µT MF can affect the expression of
protein p53 in NB69 cells. Two conformational specific anti-p53
antibodies were used to that end: pAb240, which specifically
recognizes the unfolded p53 tertiary structure, and pAb1801, which
recognizes the folded, wt isoform (41–43).
The results revealed that MF exposure causes changes in both gene
and protein expression of wt p53, as well as overexpression of
unfolded p53, together with changes in its nuclear/cytoplasmic
distribution. Additionally, MF exposure induced significant
overexpression of the anti-apoptotic protein Bcl-2.
Materials and methods
Cell culture
The neuroblastoma cell line NB69 (lot no.
03I019/2008; cat. no. 99072802) was purchased from the European
Collection of Authenticated Cell Cultures (ECACC). The cells were
periodically tested for Mycoplasma contamination (PCR) and response
to chemical and physical treatments. Cells were maintained in
Dulbecco's modified Eagle's medium (DMEM; cat. no. BE12-614F;
BioWhittaker; Lonza Group, Ltd.) supplemented with 10%
heat-inactivated foetal bovine serum (FBS; product code 11573397),
2 mM L-glutamine (product code 11539876), 100 U/ml penicillin, 100
U/ml streptomycin and 0.25 µg/ml of amphotericin B (product code
11580486; all Gibco BRL; Invitrogen; Thermo Fisher Scientific,
Inc.) and incubated in a 95% air-5% CO2 humidified
atmosphere (Forma Scientific incubators; Thermo Fisher Scientific,
Inc.). In each experimental run 4.5×104 cells
ml−1 were seeded either directly on the bottom of 60-mm
plastic Petri dishes (cat. no. 150288; Nunc, LabClinics, S.A.) or
on glass coverslips placed inside the dishes, and cultured for 4
days in the described incubation conditions before MF- or
sham-exposure.
Magnetic field exposure
The cultures were exposed to a 50-Hz, sine wave,
vertically polarized MF, at a magnetic flux density
(BAC) of 100 µT root mean square (rms). The MF exposure
set-up has been previously described (44). Briefly, current flow was supplied
by a wave generator (Newtronic, Model 200MSTPC; TER Calibration
Ltd.) that has a 3.53 mA DC offset (BDC =15 µT rms). The
generator was connected to two identical coil pairs, both set in
Helmholtz configuration. The current in the coils was monitored
using a multimeter (Hewlett Packard, model 974A; Hewlett Packard
Company) and the induced MF was routinely verified with two
magnetometers (EFA-3; Wandel and Goltermann GmbH & Co.
Elektronische Meûtechnik; and EMDEX II; Enertech Consultants). One
coil pair was placed inside each of the two magnetically shielded
chambers (Co-netic® metal; Amuneal Manufacturing Corp.)
located within two identical CO2 incubators. The
background MF inside the shielded chambers was BAC,
0.04±0.03 µT rms; and BDC, 0.05±0.04 µT rms. No increase
of temperature at the sample location was observed using two Pt100
Thermocouple probes (Fluke, Model 52; Adler Instruments) when the
coils were energized to produce the desired magnetic flux density
of 100 µT rms. In each experimental run, Petri dishes (5 per
experimental group) containing cell samples were stacked in the
central region of the Helmholtz coil gap, which ensured uniformity
of MF exposure. Only one set of coils was energized in each
experimental run. The samples in the unenergized set were
considered sham-exposed controls. Following a random sequence, both
coil sets and incubators were alternatively used for MF- or
sham-exposure. The protein expression of signaling markers was
analyzed at various time intervals after the MF- or sham-exposure
onset: 30, 60, 90 or 120 min.
Western blot analyses
Total protein extraction and immunoblotting were
performed as previously described (45). Briefly, whole cell proteins were
prepared by lysing the cells in hypotonic lysis buffer (45). Protein content was determined by
BCA protein assay kit (Pierce; Thermo Fisher Scientific, Inc.).
Equal protein volumes (60 µg) were separated from each of the
samples, using 10% SDS/PAGE gels, and transferred to nitrocellulose
membranes (Hybond ECL; GE Healthcare; Cytiva) by semidry transfer.
The membranes were then incubated at room temperature with mouse
monoclonal antibodies against proteins p53 (1:1,000; product no.
2524; Cell Signaling Technology, Inc.) or Bcl-2 (1:1,000; product
no. MBS3017024; MyBiosource, Inc.). Anti-human β-actin (1:5,000;
product no. A-5441, Sigma-Aldrich; Merck KGaA) was used as a
loading control. Following washing, the membranes were incubated
with anti-mouse IgG, horseradish peroxidase-linked whole antibody
(product no. NA931; GE Heathcare; Cytiva) or with the
fluorescent-labeled secondary antibody, IRDye 800CW, goat
anti-mouse IgG (1:15,000; product no. 926-32350; LI-COR
Biosciences), for 1 h at room temperature. For chemiluminescence
detection and visualization of the immunoreactive bands, the
enhanced detection kit ECL (RPN2132; GE Healthcare; Cytiva) and the
ProXima image densitometer (Isogen Life Science B.V.) were used.
The Odyssey infrared imaging system LI-COR was used to detect the
signal from bands marked with the fluorescent secondary antibody.
The blots were analyzed by densitometric assay using PDI Quantity
One-4.5.2 software (Bio-Rad Laboratories, Inc.). At least four
experimental replicates were conducted for each of the studied
proteins. Three MF-exposed dishes vs. three sham-exposed dishes per
experimental run and per exposure period were used.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) for p53 mRNA expression
Total RNA from NB69 cells was extracted with TRIzol
reagent (product no. T9424; Sigma-Aldrich; Merck KGaA) according to
the manufacturer's protocol, at 90 min of MF- or sham-exposure
onset. The RNA quality and quantity were assessed using NanoDrop
ND-1000 (NanoDrop Technologies; Thermo Fisher Scientific, Inc.) and
agarose gel electrophoresis (1%). A total amount of 500 ng RNA was
reverse-transcribed using a Primer Script RT™ Reagent
Kit (cat. no. RR037A; Takara Bio, Inc.). LightCycler 480 SYBR-Green
I Master mix (product no. 04887352001) and LightCycler thermal
cycler 480 II (both from Roche Applied Science) were used for
real-time PCR. All determinations were triple analyzed, and the
thermocycler protocol consisted of 5 min preincubation at 95°C,
followed by 45 cycles at 95°C for 10 sec, 60°C for 15 sec, and 72°C
for 10 sec. The following primers were used: TP53 forward,
5′-CAGCACATGACGGAGGTTGT-3′ and reverse,
5′-TCATCCAAATACTCCACACGC-3′; and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) forward 1.1, 5′-GAAGGTGAAGGTCGGAGTC-3′, and
reverse, 5′-GAAGATGGTGATGGGATTTC-3′. The melting curves were
evaluated and the PCR reaction products were separated on a 2%
agarose gel to confirm the presence of a single product. In all
cases, the efficiency of the amplification reaction was tested by
serial dilutions of cDNA, ensuring that the efficiency was linear
and close to 2. The results were analyzed using the relative
quantification method described by Pfaffl (46), which takes into account the
efficiency of the reaction for the target gene and the invariable
control. GAPDH was used as an invariant endogenous control. For
comparison and statistical analysis, data obtained were normalized
to the expression of the control group.
Immunofluorescence
After MF- or sham-exposure of the samples cultured
on coverslips, the expression of p53 and Bcl-2 were characterized
by indirect immunofluorescence and computer-assisted image
analysis. Cells were fixed with 4% formaldehyde for 20 min at 4°C,
permeabilized with ethanol/acetic acid for 20 min at −4°C and
blocked with PBS containing 10% goat serum (cat no. 31872;
Invitrogen; Thermo Fisher Scientific, Inc.) for 1 h at room
temperature. Coverslips were incubated overnight at 4°C with two
different p53 primary mouse monoclonal antibodies: p53-pAb1801
(1:100; cat. no. AHO0122, Invitrogen; Thermo Fisher Scientific,
Inc.) or p53-pAb240 (1:100; cat. no. 13-4100; Zymed Laboratories,
Inc.; Thermo Fisher Scientific, Inc.). The conformational changes
in p53 can be assessed by monitoring the reactivity with
conformation-specific antibodies pAb1801 and pAb240, which allow
discrimination between the functional, wt protein and its unfolded
isoform (47,48). Antibody p53-pAb1801 recognizes an
epitope between amino acids 32 and 79 of both wt and mutant-like
type p53 protein (49). The
conformationally altered mutant-like type isoform is specifically
recognized by antibody pAb240 (43,50).
This antibody recognizes a primary epitope that is cryptic in the
wt conformation and becomes exposed when the protein changes its
conformation towards an unfolded phenotype (51). Secondary anti-mouse-IgG conjugated
to AlexaFluor® 568 (cat. no. A11031; Molecular Probes;
Thermo Fisher Scientific, Inc.) was administered for 1 h at room
temperature to reveal p53 expression. The cell nuclei were
counterstained with Hoechst 33342 (product no. B2261; Bisbenzimide;
Sigma-Aldrich; Merck KGaA) added to the mounting medium. The mouse
monoclonal Bcl-2 primary antibody (1:100; product no. MBS3017024;
MyBiosource, Inc.) and a biotinylated secondary antibody (1:100)
were administered overnight at 4°C and 1 h at room temperature,
respectively. Immunostaining was enhanced through the ABC method
(cat. no. PK-6102; Vectastain ABC kit; Vector Laboratories, Ltd.),
and revealed with 3′3′-diaminobenzidine (DAB; product no. D5905;
Sigma-Aldrich; Merck KGaA). The cell nuclei were counterstained
with methyl-green for 5 min at room temperature. Images were
captured with a Nikon microscope (Eclipse TE300; Nikon Corporation)
and analyzed by computer imaging AnalySIS software (version 2007;
Olympus Soft-Imaging Systems GmbH). In each of at least 4
experimental runs, 4 coverslips were studied per experimental
condition: MF- or sham-exposed controls. A total of 15 microscopic
fields per coverslip were randomly selected for analysis. The total
number of nuclei and the percent of wt p53-, unfolded p53- or
Bcl-2-positive cells per microscopic field were recorded. The
cytoplasmic or nuclear location of protein p53 was determined by
computer-assisted image-analysis (AnalySIS software, version 2007;
Olympus Soft-Imaging Systems GmbH).
Statistical analysis
All experimental procedures and analyses were
conducted blindly for treatment. Data were normalized and expressed
as the means ± standard error (SEM) of at least three independent
experimental runs. Statistical analyses were performed with
Graph-Pad Prism 6.01 software (GraphPad Software, Inc.). Unpaired
two-tailed Student's t-test or the one-way ANOVA plus Bonferroni
post hoc test, were used when comparing two samples or multiple
samples, respectively. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of a MF on p53 protein
expression levels
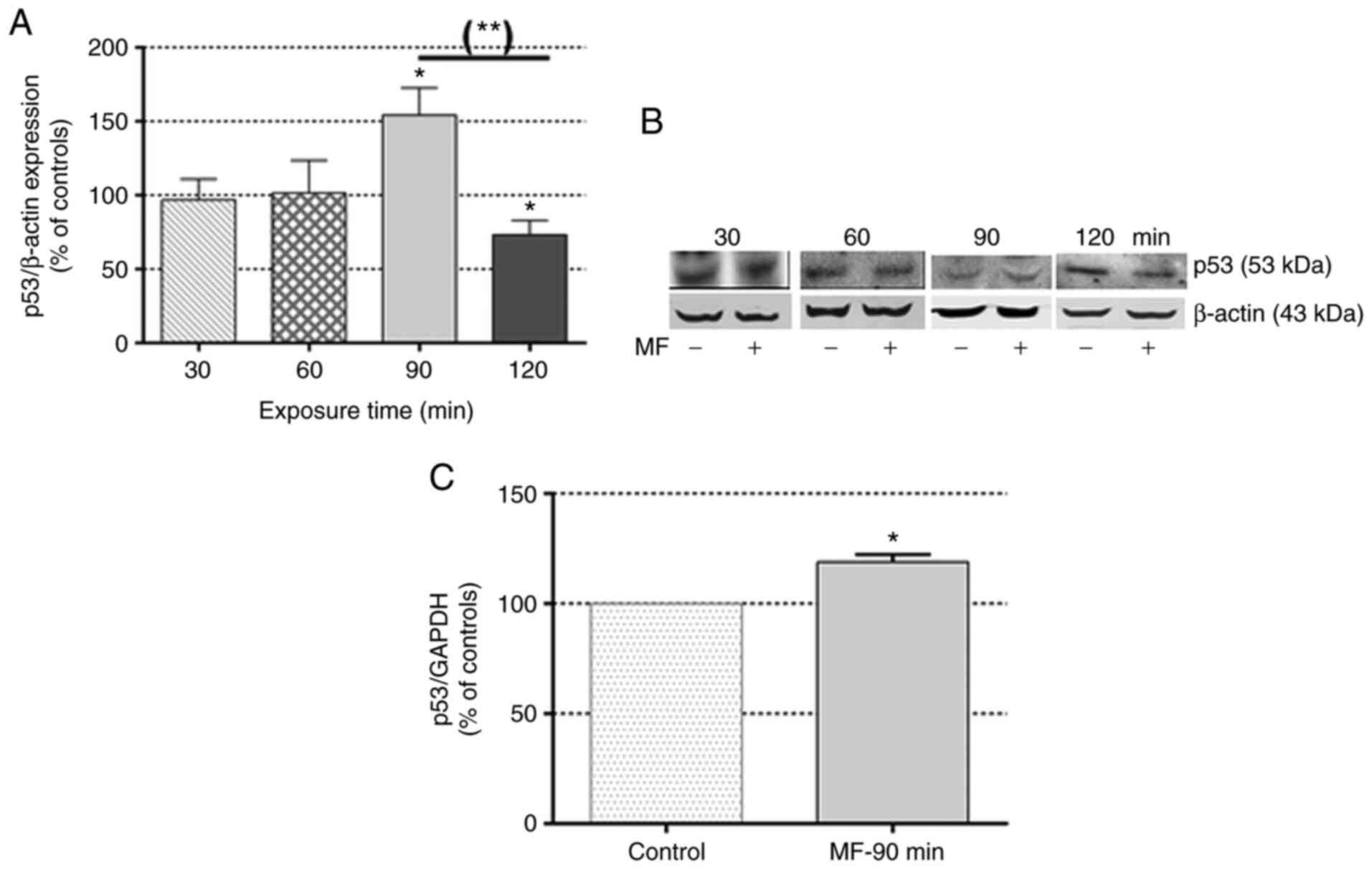
The immunoblot analysis revealed that at exposure
intervals t >60 min, the MF caused transient changes in p53
expression (Fig. 1A and B), with
significant overexpression observed at 90 min (54.40±18.30% over
the controls) followed by underexpression at 120 min (73.40±9.70%
of the controls). In addition, the RT-qPCR analysis revealed
significant overexpression of p53 mRNA (18.90±3.30% over the
controls) in samples exposed to the MF for 90 min (Fig. 1C), which is consistent with the
overexpression of p53 transcription factor revealed by
immunoblotting at the same time interval.
Effect of a MF on the number of cells
expressing p53
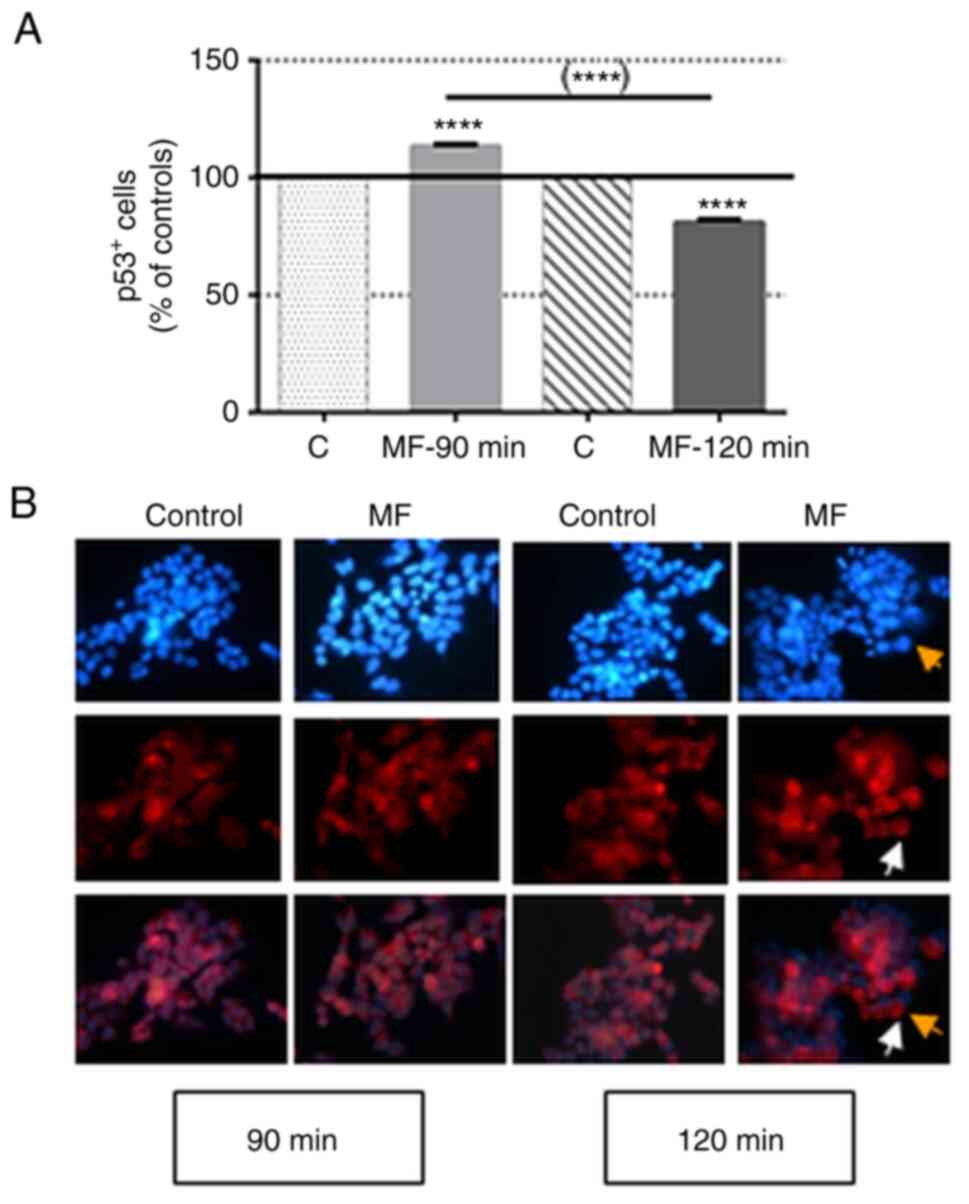
The immunocytochemical results, summarised in
Fig. 2, revealed that a 90-min
exposure to a MF significantly increased the rate of p53-positive
cells (p53+; 13.40±0.76% over the sham-exposed
controls), while after 120 min of exposure there was a significant
decrease in the rate of p53+ cells (81.09±1.28% of that
in the controls). These results are consistent with those obtained
by immunoblotting and RT-qPCR analysis on samples exposed to the
field during the same interval.
Effect of a MF on the number of cells
expressing wt or unfolded p53
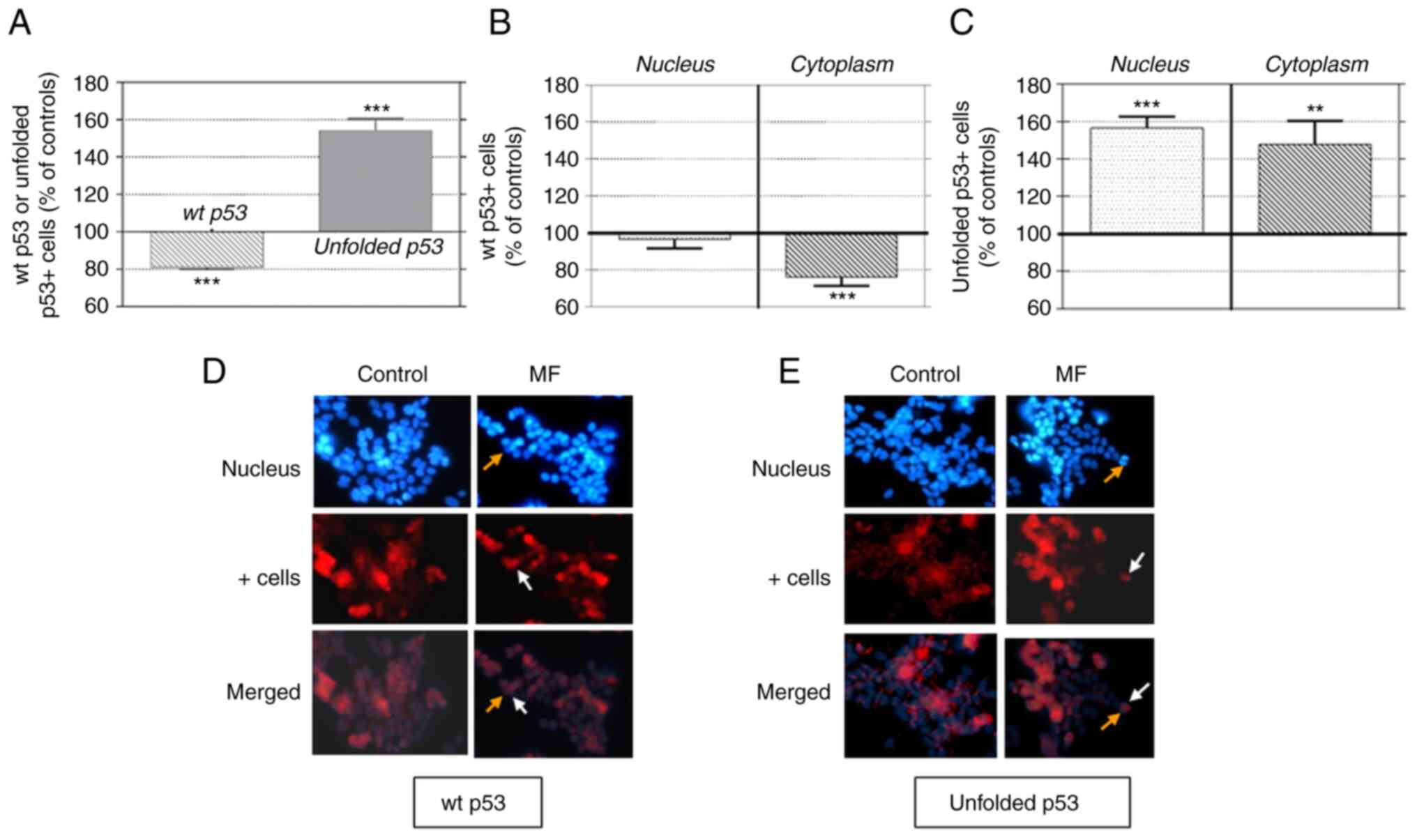
The results of the immunocytochemical analysis of
the number of cells expressing p53 protein after 120 min of MF
exposure is presented in Fig. 3.
Two types of antibodies: PAb1801, which recognizes wt p53, and
Pab240, which recognizes the unfolded isoform of p53 but not wt
p53, were used to analyze the cellular distribution (nuclear or
cytoplasmic) of these isoforms. The results summarized in Fig. 3A revealed that both isoforms were
present in control cells, with rates of 65.26±2.5 and 15.10±1.7%
for wt p53 and unfolded p53, respectively. MF exposure induced
underexpression of wt p53 (18.9±1.3% below the controls) and
increased the expression of the unfolded isoform (54.1±6.3% above
the controls). These results are consistent with the corresponding
immunoblotting data.
The decrease in the rate of wt p53+ cells
was located exclusively in the cytoplasm (76.26±2.16% of that in
the controls; Fig. 3B), with no
significant changes in nuclear labeling. By contrast, the increased
labeling of unfolded p53+ (Fig. 3C) was located both at the nuclear
and cytoplasmic levels (56.6±6.1 and 47.92±12.5% over the controls,
respectively). The photomicrographs in Fig. 3D and E illustrated these MF effects
on labeling distribution.
Effects of a MF on Bcl-2
expression
The immunoblotting results revealed that 60 and 90
min of MF exposure induced equivalent increases (35.20±11.40 and
39.20±8.20% over the corresponding controls, respectively) in the
levels of Bcl-2 expression (Fig. 4A
and B). The immunocytochemical analysis confirmed this effect
(Fig. 4C and D), revealing
significant increases over the controls in the number of
Bcl-2+ cells after MF exposure periods of 30 min
(19.1±3.6% over the controls), 60 min (27.4±1.4%), 90 min
(27.0±1.6%) and 120 min (28.6±2.5%).
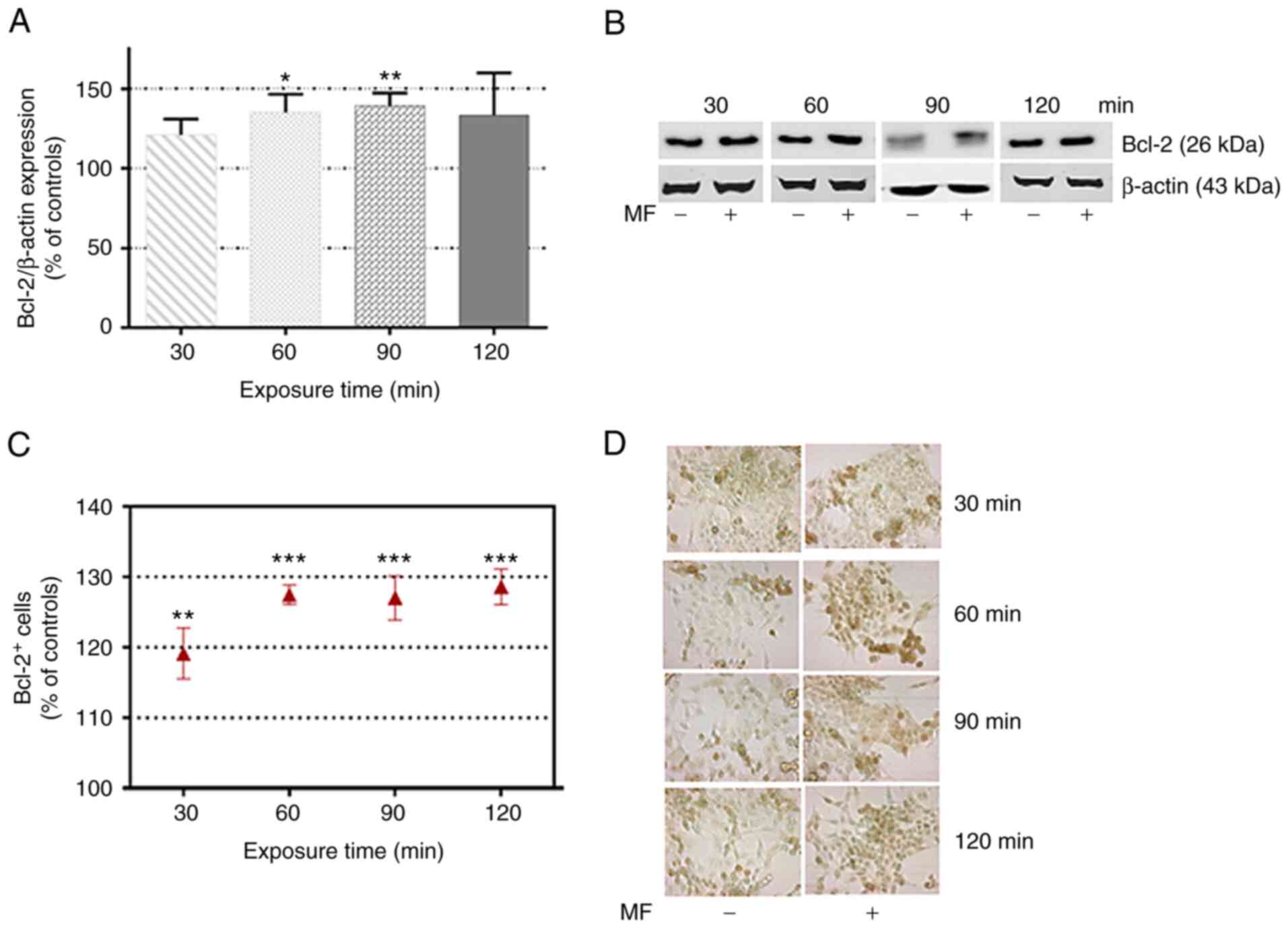 | Figure 4.Effects of a MF on Bcl-2 expression.
(A) Immunoblot analysis of Bcl-2 expression after 30, 60, 90 or 120
min of MF exposure. Data, normalized over controls, are the means ±
SEM of 4 experimental replicates per exposure time, with 3 MF- and
3 sham-exposed control samples per replicate. (B) Representative
blots for each of the tested intervals; β-actin was used as loading
control. (C) Immunocytochemical analysis of the number of Bcl-2+
cells after 30, 60, 90 or 120 min of MF exposure. Each point
represents the mean ± SEM of 4 experimental replicates, with 3 MF-
and 3 sham-exposed coverslips per replicate. A total of 15
microscopic fields per coverslips were analyzed. (D) Representative
images (magnification ×400) of the expression of the antiapoptotic
protein Bcl-2 (brown labeling). Nuclei were counterstained with
methyl green. *P<0.05, **P<0.01 and ***P<0.001 (unpaired
Student's t-test). MF, magnetic field. |
Discussion
In previous studies by the authors (38,39,44,45,52)
it was demonstrated that intermittent exposure to a 50-Hz, 100-µT
MF caused significant changes in the regulation and kinetics of the
cell cycle and in the proliferation of human neuroblastoma cells
NB69, and these responses were mediated by activation of the EGF
receptor and of the signaling pathways MAPK-p38 (free radical
dependent) and MAPK-ERK1/2 (free radical independent). These
findings, together with the fact that MF exposure increases the
overall level of free radicals (40), supports the hypothesis that the
effects of the field are partly caused through free radical
generation. The present study investigated the possibility that
protein p53, which acts as a nuclear transcription factor sensitive
to oxidative stress and, in addition to regulating cell cycle
control, promotes DNA repair (22), is one of the targets of the ELF
field. In control cells, the protein wt p53 exhibits low expression
levels due to its degradation through the ubiquitin-mediated
proteasome pathway. However, under stress conditions that lead to
DNA damage, p53 becomes stabilized and translocates to the nucleus,
where it accumulates. This leads to transcriptional activation of
several p53 target genes, including those encoding proteins
p21WAF1, BAX, and Mdm2, that can result in cell cycle arrest and
cell differentiation, senescence, or apoptosis (53,54).
Therefore, wt p53 plays a key role in the suppression of cell
proliferation control and in tumorigenesis. Thus, a potential,
MF-induced dysregulation or loss of p53 activity could alter cell
proliferation control mechanisms and eventually trigger processes
that can evolve into tumorigenesis.
The results of the present study revealed that in
the short term, MF exposure induced a transient effect on the
expression levels of wt p53 protein. Indeed, after 90 min of
exposure, overexpression of p53 gene and protein were observed,
followed 30 min later by significant underexpression of the wt
isoform. This underexpression occurred at the cytoplasmic domain
and coincided with significant nuclear and cytoplasmic
overexpression of the unfolded isoform of the protein.
Previous studies have also reported ELF MF-induced
changes in p53 expression, but in general relatively high magnetic
flux densities were applied in combination with chemical or
physical agents such as ciplastin (55) or X-rays (56). Other studies have reported no
changes in p53 expression in cardiac cells exposed continuously (60
min) or intermittently (75 min) to a weak MF of 100 µT at 50 Hz
(57). In the present study, short
exposure (t <90 min) to MF parameters equivalent to those used
by these previous authors did not induce significant changes in p53
expression, whereas longer exposure lapses (t ≥90 min) did.
Collectively, these data add to the body of evidence suggesting
that the exposure cycle and protocol, as well as other physical or
biological parameters, can critically influence the type and
magnitude of the MF-induced response (58,59).
Conversely, p53 plays a fundamental role in the
control of tumor formation. Indeed, the inactivation of p53
generally caused by TP53 gene mutations followed by loss of
functionality through a variety of non-mutational regulatory
failures, such as alterations in intracellular protein location, is
a common phenomenon reported in >50% of various human cancers
(60,61). In neuroblastomas and other human
primary tumors, this dysfunction is mediated by cytoplasmic
sequestration and nuclear exclusion of the wt protein (31,62).
The analysis of the present results does not allow
to determine whether the observed effects could be mediated by
potential MF-induced DNA alterations. However, the data revealed
that field exposure is capable of altering the wt p53 gene and
protein expression in the short term and inducing an overexpression
of the unfolded isoform of p53, which could contribute to the
subsequent proliferative effect observed in NB69 cells after longer
exposure intervals to identical MF parameters: 50 Hz and 100 µT
(38,39,44,44,52).
Such overexpression of the unfolded form of the protein could
result from MF-induced alterations in the conformation of the
malleable and conformationally labile, pAb240-reactive wt isoform,
as it occurs in response to chemical stressors (63). Indeed, it has been reported that in
vitro stimulation with serum, changes wt p53 to a mutant
conformation in murine fibroblasts (64) and that the formation of
heterooligomers of the wild and mutated types of p53 could lead to
unfolding of the wt protein (65).
Therefore, it is possible that some of the aforementioned
mechanisms have intervened in the field-induced conformational
changes of p53 that manifest as overexpression of the unfolded
isoform.
In addition, there is experimental evidence
indicating that mutations and defects in the folding of p53 disable
this protein from exerting its tumor suppressive functions in
cancer cells, while enabling it to actively intervene in various
stages of tumor progression and to promote resistance to anticancer
treatments (66,67). The present results indicate that,
in addition to inducing p53 gene and protein overexpression, MF
exposure could cause alterations in protein folding that are likely
to intervene in the field-induced proliferation promotion of
neuroblastoma cells.
On the other hand, the tumor suppressor protein p53
functions as a stress-sensitive transcription factor, and there is
compelling evidence that ELF-MFs affect cell physiology by altering
redox-related processes (59,68).
Specifically, in human neuroblastoma NB69 cells, exposure to a
100-µT MF at 50 Hz was revealed as a stress factor (38,40)
capable of increasing free radical levels and of dysregulating
mitogenic stress transduction and p53 regulation pathways, such as
the Jun NH2-terminal kinase pathway (69,70).
Under stress conditions, both wt and mutated p53 accumulate in the
cell, and only wt p53 returns to basal concentration levels once
normal conditions are restored. The fact that the levels of mutant
p53 remain elevated is attributed, at least in part, to lack of an
autoregulatory loop of wt p53, mediated by Mdm2 and other negative
regulators (61). A similar effect
could be involved in the presently reported transient
overexpression or accumulation of wt p53 induced by the MF, and in
the subsequent underexpression of the wild form and overexpression
or accumulation, both at the nuclear and cytoplasmic levels, of the
unfolded isoform of the protein. Therefore, as a whole, these data
suggest that the alterations observed in the expression of both
conformations of p53 could result from a MF-induced redox
modification of the protein, and that such alterations could
mediate the subsequent NB69 cell cycle dysregulation and cell
proliferation promotion reported in the aforementioned studies
(38,39,44,45,52).
These results also revealed that a 100-µT MF at 50
Hz causes transitory overexpression of the antiapoptotic protein
Bcl-2 at 60 and 90 min of exposure. Other authors have also
reported MF induction of Bcl-2 overexpression in various cell types
including MCF-7 (71), CHO-K1
(72), xrs5 (56) or leukemia K562 (73). However, those studies applied
higher density MF (1-5 mT) in combination with ionizing radiation
(X-rays; 1-12 Gy) or chemical agents. In most cases, exposure to
the MF inhibited the apoptosis induced by physical or chemical
agents, which resulted in overexpression of Bcl-2, an increased
number of cells in the G1 phase, and inhibition of p53
expression.
In addition to acting as a nuclear transcription
factor, wt p53 can directly interact with cytoplasmic proteins such
as Bcl-2, and neutralize their anti-apoptotic activity (34,35).
The MF-induced alterations in p53 described in the present study
could affect such an interaction and result in the observed
overexpression of free Bcl-2, which is capable of triggering
antiapoptotic and/or cell survival-promoting responses, as reported
in previous studies on NB69 cells exposed for longer periods to the
same MF stimulus (38,39,44,45,52).
In conclusion, the present results demonstrated that
exposure to a 50-Hz and 100-µT MF causes increased expression of
Bcl-2, accompanied by early overexpression (at 90 min of exposure)
of the TP53 gene and transient overexpression of the wt of the
corresponding protein. This transitoriness would be caused by
overexpression of the unfolded configuration of the p53 protein and
the corresponding underexpression of the wt p53 configuration, both
observed at 120 min of MF exposure. Such alterations could affect
the functions of p53 in the cytoplasmic and nuclear domains, thus
mediating the MF-induced dysregulations of DNA synthesis, the cell
cycle and cell proliferation in NB69 cells that have been described
in previous studies by the authors.
These results build on those of previous studies
(38,39,44,45,52)
and provide complementary information affording a more complete
picture of the cascade of effects involved in the proliferative
response of the NB69 cell line. These studies are not intended to
resolve current controversies about the potential carcinogenicity
of chronic exposure to weak ELF fields, nor are they intended to be
applied for the evaluation of real risk levels in cases of
carcinogenicity. Their purpose is to characterize the mechanisms
underlying the cell response to a physical stimulus that current
safety standards adjudge too weak to be biologically relevant.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the European
Defence Agency/Spanish Ministry of Defence, ‘Radiofrequency
Biological Effects’: grant no. MOU EUROPA ERG 101.013.
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MAM and MAT designed the study, conducted the
experiments, analyzed the data and wrote the manuscript. AU
designed the study, analyzed the data and also participated in the
writing of the manuscript. JMB conducted the RT-qPCR analyses and
also participated in the writing of the manuscript. MAM and MAT
confirm the authenticity of all the raw data. All authors read and
approved the manuscript, and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kheifets L, Ahlbom A, Crespi CM, Draper G,
Hagihara J, Lowenthal RM, Mezei G, Oksuzyan S, Schüz J, Swanson J,
et al: Pooled analysis of recent studies on magnetic fields and
childhood leukaemia. Br J Cancer. 103:1128–1135. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Saito T, Nitta H, Kubo O, Yamamoto S,
Yamaguchi N, Akiba S, Honda Y, Hagihara J, Isaka K, Ojima T, et al:
Power-frequency magnetic fields and childhood brain tumors: A
case-control study in Japan. J Epidemiol. 20:54–61. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Teepen JC and van Dijck JA: Impact of high
electromagnetic field levels on childhood leukemia incidence. Int J
Cancer. 131:769–778. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Koeman T, Slottje P, Schouten LJ, Peters
S, Huss A, Veldink JH, Kromhout H, van den Brandt PA and Vermeulen
R: Occupational exposure and amyotrophic lateral sclerosis in a
prospective cohort. Occup Environ Med. 74:578–585. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou H, Chen G, Chen C, Yu Y and Xu Z:
Association between extremely low-frequency electromagnetic fields
occupations and amyotrophic lateral sclerosis: A meta-analysis.
PLoS One. 7:e483542012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bunch KJ, Keegan TJ, Swanson J, Vincent TJ
and Murphy MFG: Residential distance at birth from overhead
high-voltage powerlines: Childhood cancer risk in Britain
1962-2008. Br J Cancer. 110:1402–1408. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Turner MC, Benke G, Bowman JD, Figuerola
J, Fleming S, Hours M, Kincl L, Krewski D, McLean D, Parent ME, et
al: Occupational exposure to extremely low-frequency magnetic
fields and brain tumor risks in the INTEROCC study. Cancer
Epidemiol Biomarkers Prev. 23:1863–1872. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Carpenter DO: Extremely low frequency
electromagnetic fields and cancer: How source of funding affects
results. Environ Res. 178:1086882019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
International Agency for Research of
Cancer (IARC), . IARC monograph on the evaluation of carcinogenic
risks to humans. Vol 80. Non-Ionizing Radiation, Part 1, Static and
Extremely Low Frequency (ELF) Electric and Magnetic Fields. IARC
Press; Lyon: 2002, PubMed/NCBI
|
|
10
|
Santini MT, Rainaldi G and Indovina PL:
Cellular effects of extremely low frequency (ELF) electromagnetic
fields. Int J Radiat Biol. 85:294–313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saliev T, Begimbetova D, Masoud AR and
Matkarimov B: Biological effects of non-ionizing electromagnetic
fields: Two sides of a coin. Prog Biophys Mol Biol. 141:25–36.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Adair RK: Extremely low frequency
electromagnetic fields do not interact directly with DNA.
Bioelectromagnetics. 19:136–138. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Focke F, Schuermann D, Kuster N and Schär
P: DNA fragmentation in human fibroblasts under extremely low
frequency electromagnetic field exposure. Mutat Res. 683:74–83.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rageh MM, El-Gebaly RH and El-Bialy NS:
Assessment of genotoxic and cytotoxic hazards in brain and bone
marrow cells of newborn rats exposed to extremely low-frequency
magnetic field. J Biomed Biotechnol. 2012:7160232012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luukkonen J, Liimatainen A, Juutilainen J
and Naarala J: Induction of genomic instability, oxidative
processes, and mitochondrial activity by 50 Hz magnetic fields in
human SH-SY5Y neuroblastoma cells. Mutat Res. 760:33–41. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang Y, Jin X, Yan C, Tian Y, Tang J and
Shen X: Case-only study of interactions between DNA repair genes
(hMLH1, APEX1, MGMT, XRCC1 and XPD) and low-frequency
electromagnetic fields in childhood acute leukemia. Leuk Lymphoma.
49:2344–2350. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Porath D, Bezryadin A, de Vries S and
Dekker C: Direct measurement of electrical transport through DNA
molecules. Nature. 403:635–638. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wan C, Fiebig T, Kelley SO, Treadway CR,
Barton JK and Zewail AH: Femtosecond dynamics of DNA-mediated
electron transfer. Proc Natl Acad Sci USA. 96:6014–6019. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Giese B: Electron transfer through DNA and
peptides. Bioorg Med Chem. 14:6139–6143. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Blank M and Goodman R: DNA is a fractal
antenna in electromagnetic fields. Int J Radiat Biol. 87:409–415.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kastenhuber ER and Lowe SW: Putting p53 in
context. Cell. 170:1062–1078. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pitolli C, Wang Y, Mancini M, Shi Y,
Melino G and Amelio I: Do mutations turn p53 into an oncogene? Int
J Mol Sci. 20:62412019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lane DP: Cancer. p53, guardian of the
genome. Nature. 358:15–16. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Levine AJ: p53, the cellular gatekeeper
for growth and division. Cell. 88:323–331. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Williams AB and Schumacher B: p53 in the
DNA-damage-repair process. Cold Spring Harb Perspect Med.
6:a0260702016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lane DP: p53 and human cancers. Br Med
Bull. 50:582–599. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vieler M and Sanyal S: p53 isoforms and
their implications in cancer. Cancers (Basel). 10:2882018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Webley K, Bond JA, Jones CJ, Blaydes JP,
Craig A, Hupp T and Wynford-Thomas D: Posttranslational
modifications of p53 in replicative senescence overlapping but
distinct from those induced by DNA damage. Mol Cell Biol.
20:2803–2808. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ishimaru D, Maia LF, Maiolino LM, Quesado
PA, Lopez PC, Almeida FC, Valente AP and Silva JL: Conversion of
wild-type p53 core domain into a conformation that mimics a
hot-spot mutant. J Mol Biol. 333:443–451. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
de Oliveira GAP, Petronilho EC, Pedrote
MM, Marques MA, Vieira TCRG, Cino EA and Silva JL: The status of
p53 oligomeric and aggregation states in cancer. Biomolecules.
10:5482020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ostermeyer AG, Runko E, Winkfield B, Ahn B
and Moll UM: Cytoplasmically sequestered wild-type p53 protein in
neuroblastoma is relocated to the nucleus by a C-terminal peptide.
Proc Natl Acad Sci USA. 93:15190–15194. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wolff A, Technau A, Ihling C,
Technau-Ihling K, Erber R, Bosch FX and Brandner G: Evidence that
wild-type p53 in neuroblastoma cells is in a conformation
refractory to integration into the transcriptional complex.
Oncogene. 20:1307–1317. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Riley T, Sontag E, Chen P and Levine A:
Transcriptional control of human p53-regulated genes. Nat Rev Mol
Cell Biol. 9:402–412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Green DR and Kroemer G: Cytoplasmic
functions of the tumour suppressor p53. Nature. 458:1127–1130.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou X, Hao Q and Lu H: Mutant p53 in
cancer therapy-the barrier or the path. J Mol Cell Biol.
11:293–305. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tomita Y, Marchenko N, Erster S,
Nemajerova A, Dehner A, Klein C, Pan H, Kessler H, Pancoska P and
Moll UM: WT p53, but not tumor-derived mutants, bind to Bcl2 via
the DNA binding domain and induce mitochondrial permeabilization. J
Biol Chem. 281:8600–8606. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
De S, Campbell C, Venkitaraman AR and
Esposito A: Pulsatile MAPK signaling modulates p53 activity to
control cell fate decisions at the G2 checkpoint for DNA damage.
Cell Rep. 30:2083–2093.e5. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martínez MA, Úbeda A, Moreno J and Trillo
MÁ: Power frequency magnetic fields affect the p38 MAPK-mediated
regulation of NB69 cell proliferation implication of free radicals.
Int J Mol Sci. 17:5102016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Martínez MA, Úbeda A and Trillo MÁ:
Involvement of the EGF receptor in MAPK signaling activation by a
50 Hz magnetic field in human neuroblastoma cells. Cell Physiol
Biochem. 52:893–907. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Martínez MA, Úbeda A and Trillo MÁ: Role
of NADPH oxidase in MAPK signaling activation by a 50 Hz magnetic
field in human neuroblastoma cells. Electromagn Biol Med.
40:103–116. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bartek J, Iggo R, Gannon J and Lane DP:
Genetic and immunochemical analysis of mutant p53 in human breast
cancer cell lines. Oncogene. 5:893–899. 1990.PubMed/NCBI
|
|
42
|
Méplan C, Richard MJ and Hainaut P:
Metalloregulation of the tumor suppressor protein p53: Zinc
mediates the renaturation of p53 after exposure to metal chelators
in vitro and in intact cells. Oncogene. 19:5227–5236. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sabapathy K and Lane DP: Understanding p53
functions through p53 antibodies. J Mol Cell Biol. 11:317–329.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Trillo MA, Martínez MA, Cid MA, Leal J and
Úbeda A: Influence of a 50 Hz magnetic field and of
all-trans-retinol on the proliferation of human cancer cell lines.
Int J Oncol. 40:1405–1413. 2012.PubMed/NCBI
|
|
45
|
Martínez MA, Úbeda A, Cid MA and Trillo
MÁ: The proliferative response of NB69 human neuroblastoma cells to
a 50 Hz magnetic field is mediated by ERK1/2 signaling. Cell
Physiol Biochem. 29:675–686. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Milner J, Cook A and Sheldon M: A new
anti-p53 monoclonal antibody, previously reported to be directed
against the large T antigen of simian virus 40. Oncogene.
1:453–455. 1987.PubMed/NCBI
|
|
48
|
Gannon JV, Greaves R, Iggo R and Lane DP:
Activating mutations in p53 produce a common conformational effect.
A monoclonal antibody specific for the mutant form. EMBO J.
9:1595–1602. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nagata Y, Anan T, Yoshida T, Mizukami T,
Taya Y, Fujiwara T, Kato H, Saya H and Nakao M: The stabilization
mechanism of mutant-type p53 by impaired ubiquitination: The loss
of wild-type p53 function and the hsp90 association. Oncogene.
18:6037–6049. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Milner J: Flexibility: The key to p53
function? Trends Biochem Sci. 20:49–51. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Stephen CW and Lane DP: Mutant
conformation of p53. Precise epitope mapping using a filamentous
phage epitope library. J Mol Biol. 225:577–583. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Trillo MÁ, Martínez MA, Cid MA and Úbeda
A: Retinoic acid inhibits the cytoproliferative response to weak
50-Hz magnetic fields in neuroblastoma cells. Oncol Rep.
29:885–894. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ko LJ and Prives C: p53: Puzzle and
paradigm. Genes Dev. 10:1054–1072. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chen S, Gao R, Yao C, Kobayashi M, Liu SZ,
Yoder MC, Broxmeyer H, Kapur R, Boswell HS, Mayo LD and Liu Y:
Genotoxic stresses promote clonal expansion of hematopoietic stem
cells expressing mutant p53. Leukemia. 32:850–854. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Baharara J, Hosseini N and Farzin TR:
Extremely low frequency electromagnetic field sensitizes
cisplatin-resistant human ovarian adenocarcinoma cells via P53
activation. Cytotechnology. 68:1403–1413. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tian F, Nakahara T, Yoshida M, Honda N,
Hirose H and Miyakoshi J: Exposure to power frequency magnetic
fields suppresses X-ray-induced apoptosis transiently in
Ku80-deficient xrs5 cells. Biochem Biophys Res Commun. 292:355–361.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang Y, Liu X, Zhang Y, Wan B, Zhang J, He
W, Hu D, Yang Y, Lai J, He M and Chen C: Exposure to a 50 Hz
magnetic field at 100 µT exerts no DNA damage in cardiomyocytes.
Biol Open. 8:bio0412932019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kesari KK, Juutilainen J, Luukkonen J and
Naarala J: Induction of micronuclei and superoxide production in
neuroblastoma and glioma cell lines exposed to weak 50 Hz magnetic
fields. J R Soc Interface. Jan 1–2016.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Falone S, Santini S Jr, Cordone V, Di
Emidio G, Tatone C, Cacchio M and Amicarelli F: Extremely
low-frequency magnetic fields and redox-responsive pathways linked
to cancer drug resistance: Insights from co-exposure-based in vitro
studies. Front Public Health. 6:332018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Harris CC and Hollstein M: Clinical
implications of the p53 tumor-suppressor gene. N Engl J Med.
329:1318–1327. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Vijayakumaran R, Tan KH, Miranda PJ, Haupt
S and Haupt Y: Regulation of mutant p53 protein expression. Front
Oncol. 5:2842015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen L, Malcolm AJ, Wood KM, Cole M,
Variend S, Cullinane C, Pearson AD, Lunec J and Tweddle DA: p53 is
nuclear and functional in both undifferentiated and differentiated
neuroblastoma. Cell Cycle. 6:2685–2696. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sasaki M, Nie L and Maki CG: MDM2 binding
induces a conformational change in p53 that is opposed by
heat-shock protein 90 and precedes p53 proteasomal degradation. J
Biol Chem. 282:14626–14634. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Milner J and Watson JV: Addition of fresh
medium induces cell cycle and conformation changes in p53, a tumour
suppressor protein. Oncogene. 5:1683–1690. 1990.PubMed/NCBI
|
|
65
|
Milner J and Medcalf EA: Cotranslation of
activated mutant p53 with wild type drives the wild-type p53
protein into the mutant conformation. Cell. 65:765–774. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Do PM, Varanasi L, Fan S, Li C, Kubacka I,
Newman V, Chauhan K, Daniels SR, Boccetta M, Garrett MR, et al:
Mutant p53 cooperates with ETS2 to promote etoposide resistance.
Genes Dev. 26:830–845. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Schmidt V, Nagar R and Martinez LA:
Control of nucleotide metabolism enables mutant p53′s oncogenic
gain-of-function activity. Int J Mol Sci. 18:27592017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lai H: Exposure to static and
extremely-low frequency electromagnetic fields and cellular free
radicals. Electromagn Biol Med. 38:231–248. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Fuchs SY, Adler V, Pincus MR and Ronai Z:
MEKK1/JNK signaling stabilizes and activates p53. Proc Natl Acad
Sci USA. 95:10541–10546. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Buschmann T, Potapova O, Bar-Shira A,
Ivanov VN, Fuchs SY, Henderson S, Fried VA, Minamoto T,
Alarcon-Vargas D, Pincus MR, et al: Jun NH2-terminal kinase
phosphorylation of p53 on Thr-81 is important for p53 stabilization
and transcriptional activities in response to stress. Mol Cell
Biol. 21:2743–2754. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ding GR, Nakahara T, Tian FR, Guo Y and
Miyakoshi J: Transient suppression of X-ray-induced apoptosis by
exposure to power frequency magnetic fields in MCF-7 cells. Biochem
Biophys Res Commun. 286:953–957. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Nakahara T, Yaguchi H, Yoshida M and
Miyakoshi J: Effects of exposure of CHO-K1 cells to a 10-T static
magnetic field. Radiology. 224:817–822. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Brisdelli F, Bennato F, Bozzi A, Cinque B,
Mancini F and Iorio R: ELF-MF attenuates quercetin-induced
apoptosis in K562 cells through modulating the expression of Bcl-2
family proteins. Mol Cell Biochem. 397:33–43. 2014. View Article : Google Scholar : PubMed/NCBI
|