Introduction
Physiologically, cell cycle progression and cell
proliferation are under precise and coordinated control, whereas
uncontrolled cell proliferation caused by abnormal cell cycle
progression is a key feature of cancer development/progression.
Understanding the progression and regulation of the cell cycle is
of significant importance for improving cancer treatments (1). The cell cycle consists of a G1 phase,
S phase (DNA synthesis), G2 phase, and M phase (mitosis), and each
step is jointly regulated by cyclin proteins and related
cyclin-dependent kinases (CDKs) (2). To date, eight types of cyclin
proteins, cyclin A to H, have been identified in mammalian cells
(2,3), and can be further divided into
multiple sub-types depending on their functions. It has been widely
shown that cyclin genes play regulatory roles in a variety of
cancers (4), including urinary
malignant tumors (5,6), digestive tract malignant tumors
(7,8), reproductive malignant tumors
(9,10), and respiratory malignant tumors
(11), amongst others.
Breast cancer is caused by the uncontrolled
proliferation of breast epithelial tissue cells and is affected by
various carcinogenic factors, the environment, genetics, and other
factors (12). Based on the
presence or absence of marker proteins including estrogen receptor
(ER), progesterone receptor (PR), human epidermal growth factor 2
(HER-2), and Ki-67, breast cancer can be classified into four
subtypes, namely luminal A, luminal B, HER-2 enriched, and
triple-negative (13). According
to the 2020 Global Cancer Statistics (14), breast cancer not only ranks highest
amongst the most common malignant tumors in women worldwide, but
also surpasses lung cancer as the most commonly diagnosed cancer.
Breast cancer is an extremely heterogeneous malignant tumor with
inter-tumor and intra-tumor variability (15). Therefore, novel molecular
mechanisms and biomarkers for improving the detection of
early-stage breast cancer and management of breast cancer are
needed, with a long-term goal of improving individualized
therapy.
Although there are several causes underlying the
development of breast cancer, the cyclin family of genes has
consistently been shown to play a pivotal role in aberrant cell
cycle progression. CyclinA genes are divided into CCNA1 and CCNA2.
CCNA1 exerts differential effects in different diseases (10,16),
whereas there are fewer studies on the role of CCNA1 in breast
cancer (17). CyclinA2 is widely
upregulated in a variety of cancers (18) and plays a significant role in
regulating the cell cycle (19,20).
CCNB1 is expressed in almost all tissues in humans
and is highly expressed in a variety of cancers (21). It binds to CDK1 to form a complex,
a key factor regulating the G2-to-M transition and mitotic
progression (22). In solid
tumors, the expression of CCNB1 is considered a substantial
prognostic parameter (4,23). CyclinB2 also binds to CDK1 to form
a complex, which inhibits the G2-to-M transition, thereby inducing
cell cycle arrest (24). The
expression of CCNB2, an oncogene, is upregulated in several types
of malignant tumors (7,24–26),
and its upregulated expression is associated with a poor prognosis.
CyclinB3 possesses homology with cyclinA and cyclinB1/2, and is
expressed in animals, insects, and human tissue (27). CCNB3 is more interrelated with
BCOR, which encodes the BCL6 co-repressor for co-transcriptional
expression, and does not appear to exhibit any obvious specificity
in its upregulation regarding cancer type (28–30).
In contrast with other the cyclin genes, the
function of CCNC-encoding cyclinC remains largely unknown. It binds
to CDK3 and regulates the cell cycle in the G1 and G2 phases, and
stimulates the reactivation of the cell cycle from a resting state
(31,32). CCNC was shown to be upregulated
(33) and increased cell
proliferation in 82.6% of breast cancer cases (34).
D-type cyclins can bind to CDK4/6 and phosphorylate
various substrates involved in the G1-to-S phase transition
(35). In >50% of breast cancer
subtypes, cyclinD1 protein expression is upregulated (36–38)
and this in turn reduces the efficacy of treatments (39). The specificity of CCND1 expression
for the differential diagnosis of benign and malignant mesothelial
hyperplasia has been shown to approach 100% (40). CCND2 is one of the essential
factors affecting endocrine resistance in breast cancer (41). Hypermethylation of CCND2
significantly increases the risk of death and is deemed an
independent factor of a poor prognosis in triple-negative breast
cancer (42,43). A meta-analysis showed that the
upregulated expression of CCND3 was related to poorer overall
survival (OS) in breast cancer and bladder cancer patients
(44).
Overexpression of cyclinE1 and cyclinE2 accelerates
cell cycle progression by shortening the G1-to-S phase transition
period, and promoting cell proliferation and tumorigenesis,
resulting in poorer survival rates (45–48).
CCNF is expressed in all stages of the cell cycle, and accumulates
in the S phase, reaching a peak in the G2 phase and gradually
declining in the M phase (3). Its
expression in different types of cancer varies (49), for example, its expression is low
in liver cancer (50), high in
gastric cancer (51), and unknown
in breast cancer (52).
CCNG1-encoding cyclinG1 was identified as a target
of p53-regulated transcription (53). Several studies have shown that
CCNG1, which is hypothesized to be an estrogen regulatory gene
(54), is downregulated in breast
cancer (55–57). CCNH typically binds to CDK7 and
promotes cancer cell migration during carcinogenesis (58). It has been shown that high
expression of CCNH is correlated with a poor prognosis in patients
with lung cancer (58) and
gastrointestinal cancer (8).
However, the related role of CCNH in breast cancer has been
difficult to determine.
All types of cyclin genes are associated in some
manner with the prognosis and/or drug tolerance in breast cancer.
Thus far, certain cyclin genes have been verified as biomarkers in
the prognostic prediction, evaluation of curative effects, and
exploration of drug tolerance mechanisms. Yet, the role/effects of
the remainder of the cyclin genes in breast cancer are incompletely
understood. Therefore, in this study, bioinformatics methods were
used to analyze data from publicly available databases to
investigate the expression and mutation of different cyclin genes
in breast cancer, attempting to excavate novel biomarkers.
Material and methods
Cell culture
All cell lines used in the present study were
obtained from ATCC, including MCF-10A (normal breast tissue cells),
MCF-7 (hormone receptor-positive breast cancer cells), MDA-MB-231,
MDA-MB-468, and BT-549 (triple negative breast cancer cell lines),
and SK-BR-3 (HER2 positive breast cancer cells). All breast cancer
cell lines were maintained in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (NEWZERUM, Ltd.), and
100 U/ml penicillin and 100 mg/ml streptomycin (Gibco; Thermo
Fisher Scientific, Inc.). MCF-10A cells were cultured in DMEM/F12
supplemented with 5% (vol/vol) horse serum, 20 ng/ml EGF, 100 ng/ml
cholera toxin, 0.01 mg/ml insulin, and 500 ng/ml hydrocortisone
(MCF-10A specific medium; CM-0525-125; Procell Life Science &
Technology Co., Ltd.). All cell lines were maintained at 37°C in a
humidified incubator supplied with 5% CO2.
Reverse transcription-quantitative
(RT-q)PCR
Cells were plated in 6-well plates and cells in the
logarithmic growth phase were used for RNA extraction. Total RNA
was extracted using an ESscience RNA-Quick Purification Kit. Total
RNA concentration and purity were analyzed in duplicate using a
NanoDrop One (Thermo Fisher Scientific, Inc.; cat. no. AZY1705838).
cDNA was synthesized from qualified 1,000 ng RNA using an RT-PCR
reverse transcription kit (Hifair® III 1st Strand cDNA
Synthesis SuperMix for qPCR (gDNA digester plus), Shanghai Yeasen
Biotechnology Co., Ltd.). The reverse transcription temperature
protocol was: 25°C for 5 min, 55°C for 15 min, and 85°C for 5 min,
as per the manufacturer's protocol. cDNA was stored at −20°C until
required or kept on ice if used immediately. qPCR was performed
using SYBR Green PCR reagents (Hieff UNICON® qPCR SYBR
Green Master Mix; Shanghai Yeasen Biotechnology Co., Ltd.) on a
ROCHE LightCycler 480 II detection system. The thermocycling
conditions were: 95°C for 10 min; followed by 40 cycles at 95°C for
10 sec and 60°C for 30 sec. GAPDH was used as the internal
control, and relative mRNA levels were calculated using the
2−∆∆cq method (59).
Primers (sequences provided in Table
I) were designed using Primer Bank (https://pga.mgh.harvard.edu/primerbank/) and were
synthesized by Guangzhou IGE Biotechnology, Ltd. Primer targets
were confirmed using NCBI blast (https://www.ncbi.nlm.nih.gov/tools/primer-blast/primertool.cgi?ctg_time=1656555059&job_key=6OI3EQEuDIYruAm9BN0tj37GPL1T1SegUg).
 | Table I.Sequences of the primers used. |
Table I.
Sequences of the primers used.
| Gene symbol | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| CCNA1 |
GAGGTCCCGATGCTTGTCAG |
GTTAGCAGCCCTAGCACTGTC |
| CCNA2 |
CGCTGGCGGTACTGAAGTC |
GAGGAACGGTGACATGCTCAT |
| CCNB1 |
AATAAGGCGAAGATCAACATGGC |
TTTGTTACCAATGTCCCCAAGAG |
| CCNB2 |
CCGACGGTGTCCAGTGATTT |
TGTTGTTTTGGTGGGTTGAACT |
| CCNB3 |
ATGAAGGCAGTATGCAAGAAGG |
CATCCACACGAGGTGAGTTGT |
| CCNC |
CCTTGCATGGAGGATAGTGAATG |
AAGGAGGATACAGTAGGCAAAGA |
| CCND1 |
GCTGCGAAGTGGAAACCATC |
CCTCCTTCTGCACACATTTGAA |
| CCND2 |
ACCTTCCGCAGTGCTCCTA |
CCCAGCCAAGAAACGGTCC |
| CCND3 |
TACCCGCCATCCATGATCG |
AGGCAGTCCACTTCAGTGC |
| CCNE1 |
GCCAGCCTTGGGACAATAATG |
CTTGCACGTTGAGTTTGGGT |
| CCNE2 |
TCAAGACGAAGTAGCCGTTTAC |
TGACATCCTGGGTAGTTTTCCTC |
| CCNF |
CCCCGAAGATGTGCTCTTTCA |
GCCTTCATTGTAGAGGTAGGCT |
| CCNG1 |
GAGTCTGCACACGATAATGGC |
GTGCTTGGGCTGTACCTTCA |
| CCNG2 |
TCTCGGGTTGTTGAACGTCTA |
GTAGCCTCAATCAAACTCAGCC |
| CCNH |
TGTTCGGTGTTTAAGCCAGCA |
TCCTGGGGTGATATTCCATTACT |
| GAPDH |
GGAGCGAGATCCCTCCAAAAT |
GGCTGTTGTCATACTTCTCATGG |
Bioinformatics analysis
Oncomine (60), an
online large-scale tumor gene chip database, can be used to analyze
the transcriptional expression levels of cyclin genes in clinical
breast cancer samples compared with normal breast tissues (Fold
change=2, P<0.001, top 10% gene rank, for screening out samples
with relatively significant differential expression in cyclin
genes).
GEPIA (61) is
primarily used for differential expression analysis between cancer
and normal tissues, and for correlation analysis between gene
expression and clinical pathological stage. The OS and risk-score
analyses of breast cancer patients were assessed using Kaplan-Meier
Plotter (62) and METABRIC using
the auto-select best cutoff. cBioportal (63) was further used to analyze the
genomic profiles of cyclin genes in TCGA, and analyze the
relationship amongst genes through protein-protein interaction
(PPI) analysis.
Database for annotation,
visualization, and integrated discovery (DAVID) knowledgebase
DAVID contains species-specific gene/protein
identifiers and their annotations from a variety of public genomic
resources such as NCBI, Gene Ontology (GO), and Kyoto Encyclopedia
of Genes and Genomes (KEGG), amongst others, to allow the
incorporation of a diverse range of arrays of functional and
sequencing annotations, greatly enriching the level of biological
information available for a required gene (e.g. gene ID, pathways,
etc.) (64). In this study, the GO
and KEGG enrichment analysis of cyclin genes by DAVID were used to
explore the functions and mechanisms of cyclin genes, R-4.0.4
(65,66) was used to draw figures.
Statistical analysis
A one-way ANOVA was used to compare the differences
between multiple groups; a post-hoc Dunnett's multiple comparisons
test was used to compare the mean of each column with the mean of
the control group (MCF-10A). All statistical analyses were
performed in GraphPad Prism version 9 (GraphPad Software, Inc.).
For all bioinformatics analysis, all analyses were performed by the
specific tools used in the corresponding website. P<0.05 was
considered to indicate a statistically significant difference.
Results
Transcriptional expression of the
cyclin genes in breast cancer patients
The transcriptional expression levels of the cyclin
genes in breast cancer and normal breast tissues were compared
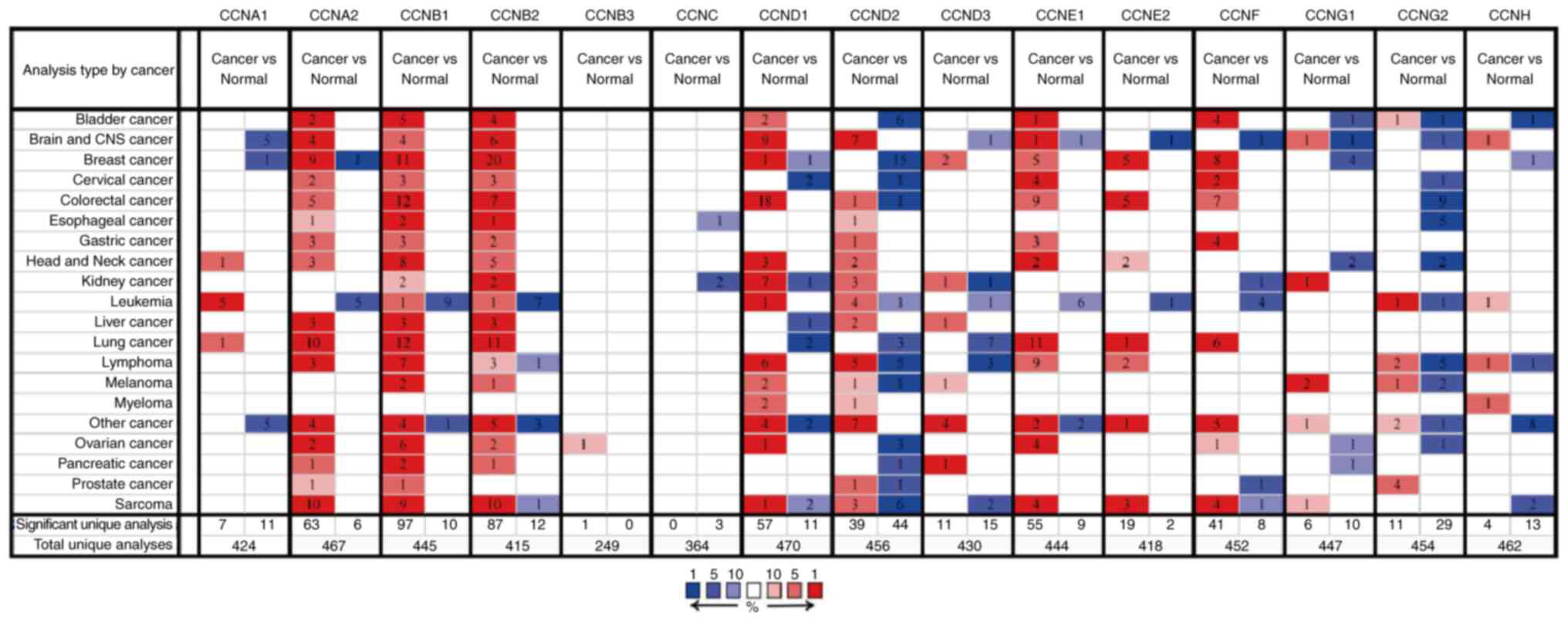
using data obtained from Oncomine (Fig. 1). CCNA2, CCNB1, CCNB2, CCND1,
CCND3, CCNE1, CCNE2, and CCNF expression was upregulated
in the tumor tissues; CCND3 and CCNE1 expression was
moderately upregulated, whereas the rest of the genes exhibited
significant upregulation. Of note, CCNA2 mRNA expression in
one of the datasets was significantly lower in the breast cancer
tissues. Similarly, CCND1 mRNA expression was slightly
downregulated in one of the datasets. The mRNA expression levels of
CCNA1, CCND2, CCNG1, and CCNH were downregulated in
the tumor tissues, particularly CCND2. Of note, the
transcriptional expression of CCNB3, CCNC, and CCNG2
had not been collected in the Oncomine database.
Relationship between the mRNA
expression of cyclin genes and clinicopathological stages of breast
cancer
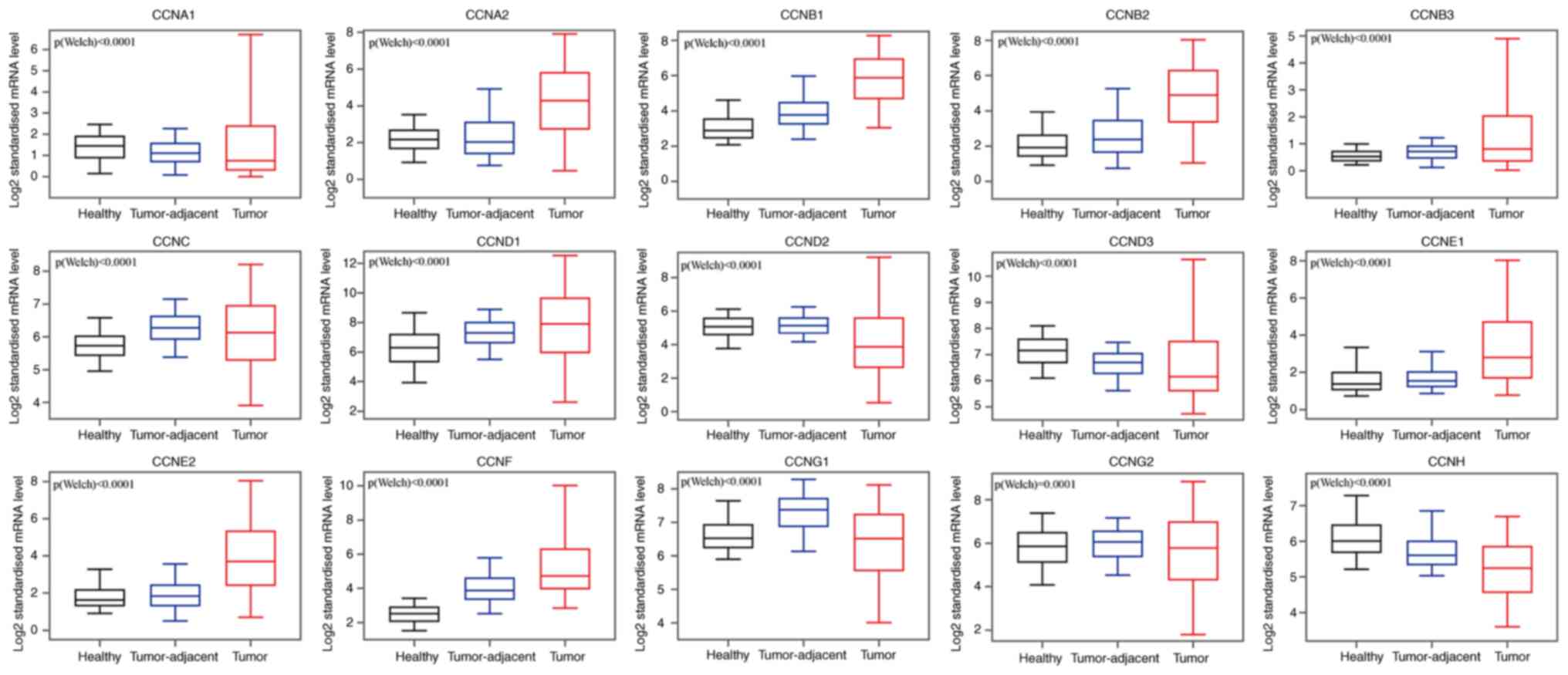
Comparative analysis of the transcriptional
expression of cyclin genes in breast cancer tissues, tumor-adjacent
tissues, and normal breast tissues in the GEPIA database
demonstrated that the mRNA expression of CCNA2, CCNB1, CCNB2,
CCNB3, CCNC, CCND1, CCNE1, CCNE2, and CCNF in breast
cancer and tumor-adjacent tissues was higher than that in the
normal breast tissues (Fig. 2),
whereas CCNA1, CCND2, CCND3, CCNG1, and CCNH mRNA
expression was higher in the normal tissues compared with the other
tissues. CCNG2 transcriptional expression did not differ
significantly between tumors and normal tissues, and in both, its
expression was lower compared with the tumor-adjacent tissues. We
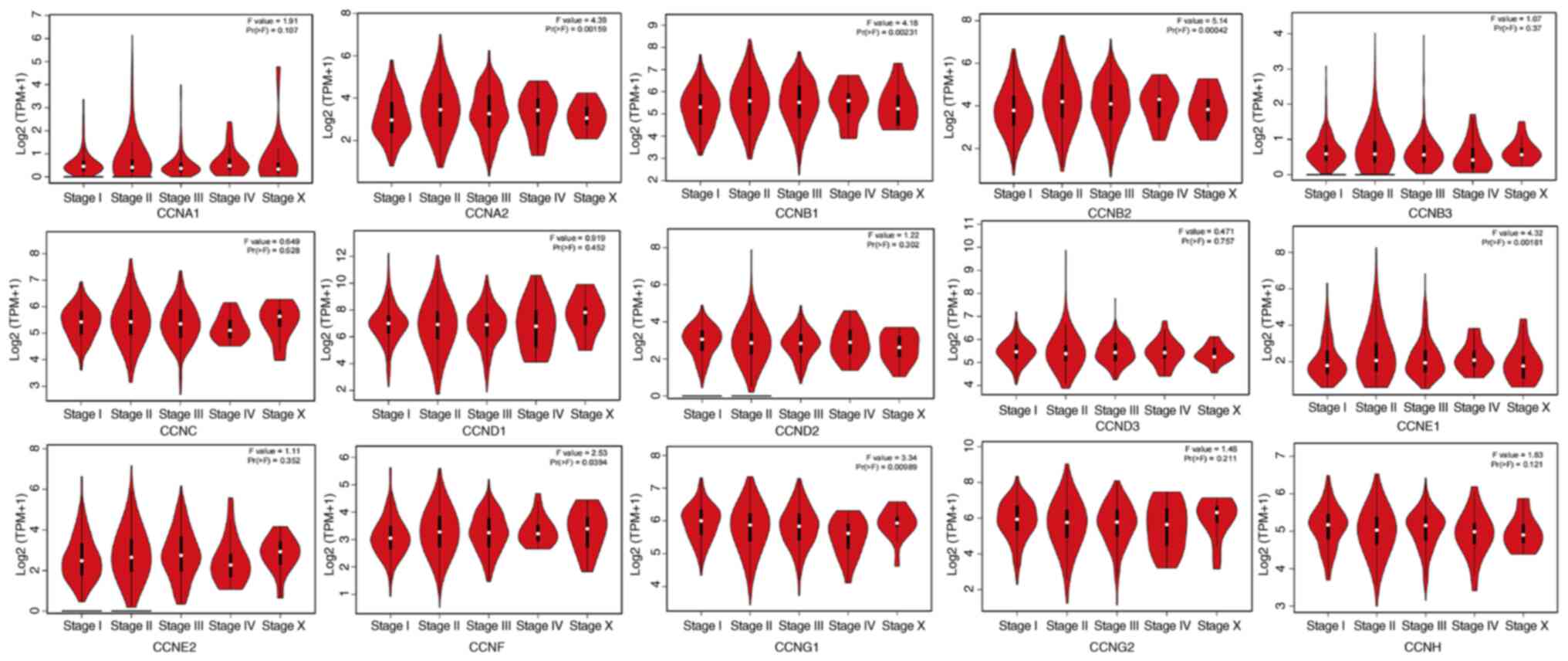
also analyzed the transcriptional expression of cyclin genes in the
different clinical stages of breast cancer (Fig. 3). CCNA2, CCNB1, CCNB2, CCNE1,
CCNF and CCNG1 expression differed significantly, and
the gene expression was highest in stage IV; however, the other
genes did not exhibit differential expression based on stage.
Correlation between the
transcriptional expression of the cyclin genes and the survival
period of patients with breast cancer
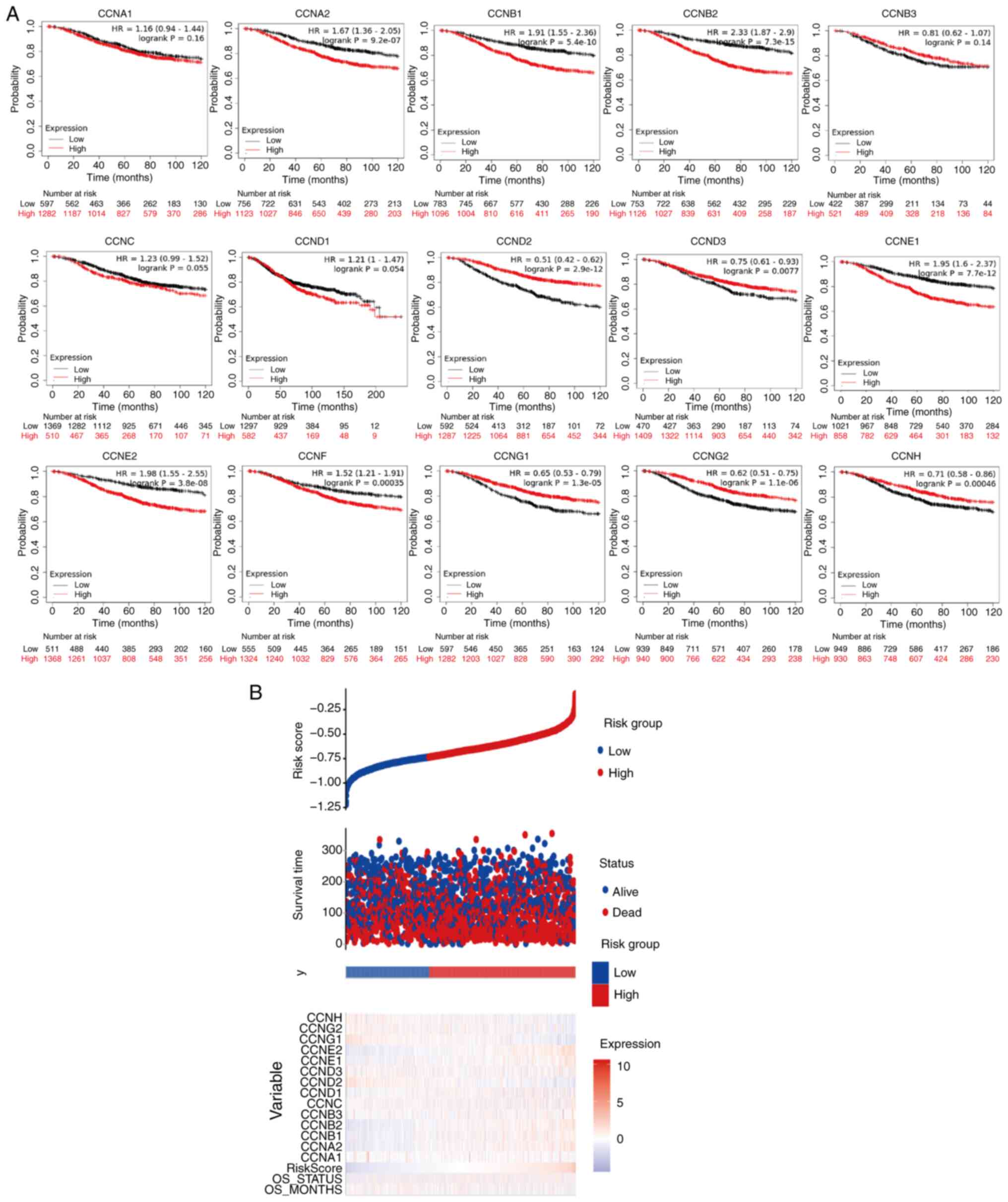
To further explore the role of cyclin genes in the
survival of breast cancer patients, Kaplan-Meier Plotter was used
(Fig. 4A). The results showed that
lower expression of CCNA2, CCNB1, CCNB2, CCNE1, CCNE2, and
CCNF and higher expression of CCND2, CCND3, CCNG1,
CCNG2, and CCNH was significantly correlated with a
better OS (P<0.05). However, high expression of CCNB3
mRNA was not associated with OS in patients (P=0.13). The curve
trend showed that OS was longer if the transcriptional expression
of CCNC was further reduced (P=0.064). The risk-score
analysis provided basic evidence for the above survival analysis
(Fig. 4B), the only thing lacking
was the analysis of CCNF, but from the results, the
relationship between low CCNF mRNA expression and high
survival was unexpected.
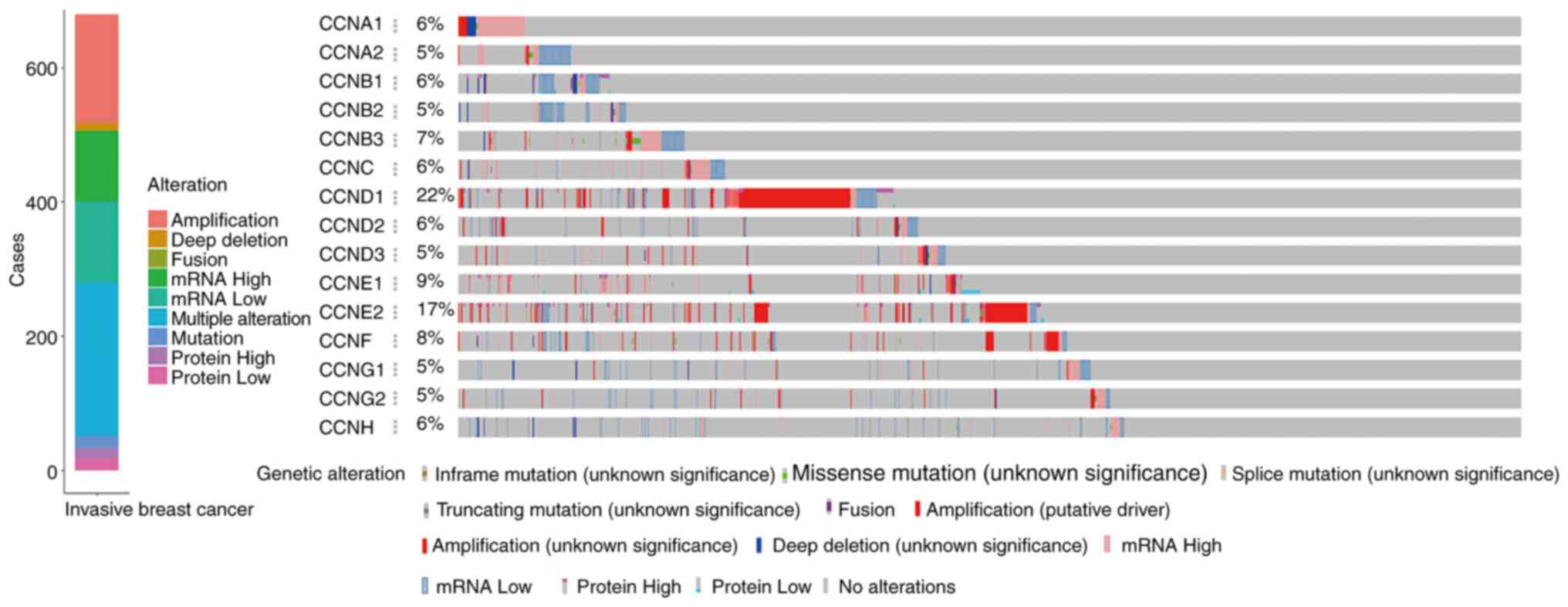
Alterations of expression of the
cyclin genes in breast cancer and correlation analysis
The online cBioportal tools were used to analyze the
alterations of cyclin genes and the related correlation in invasive
breast cancer from TCGA. Among the samples from 1,084 breast cancer
patients, various alterations were detected in 679 samples
(Fig. 5). Mutations including
inframe, missense, splicing, and truncations occurred in 18
samples, fusion-mutations occurred only in 1 sample, amplification
occurred in 161 samples, deep deletions occurred in 12 samples,
upregulated mRNA expression was observed in 105 samples,
downregulated mRNA expression was observed in 121 samples,
upregulated protein expression was observed in 15 samples,
downregulated protein expression was observed in 18 samples, and
multiple alterations were observed in 228 samples.
As shown in the specific variation analysis, the
mRNA expression of CCNA1, CCNC, CCND3, CCNE1, and
CCNB3 was upregulated, and that of CCNA2, CCNB1, CCNB2,
CCND2, CCNG1, CCNG2, and CCNH was downregulated. Gene
amplifications were more pronounced with CCND1, CCNE2, and
CCNF. Furthermore, a fusion mutation was observed in the
CCNF gene.
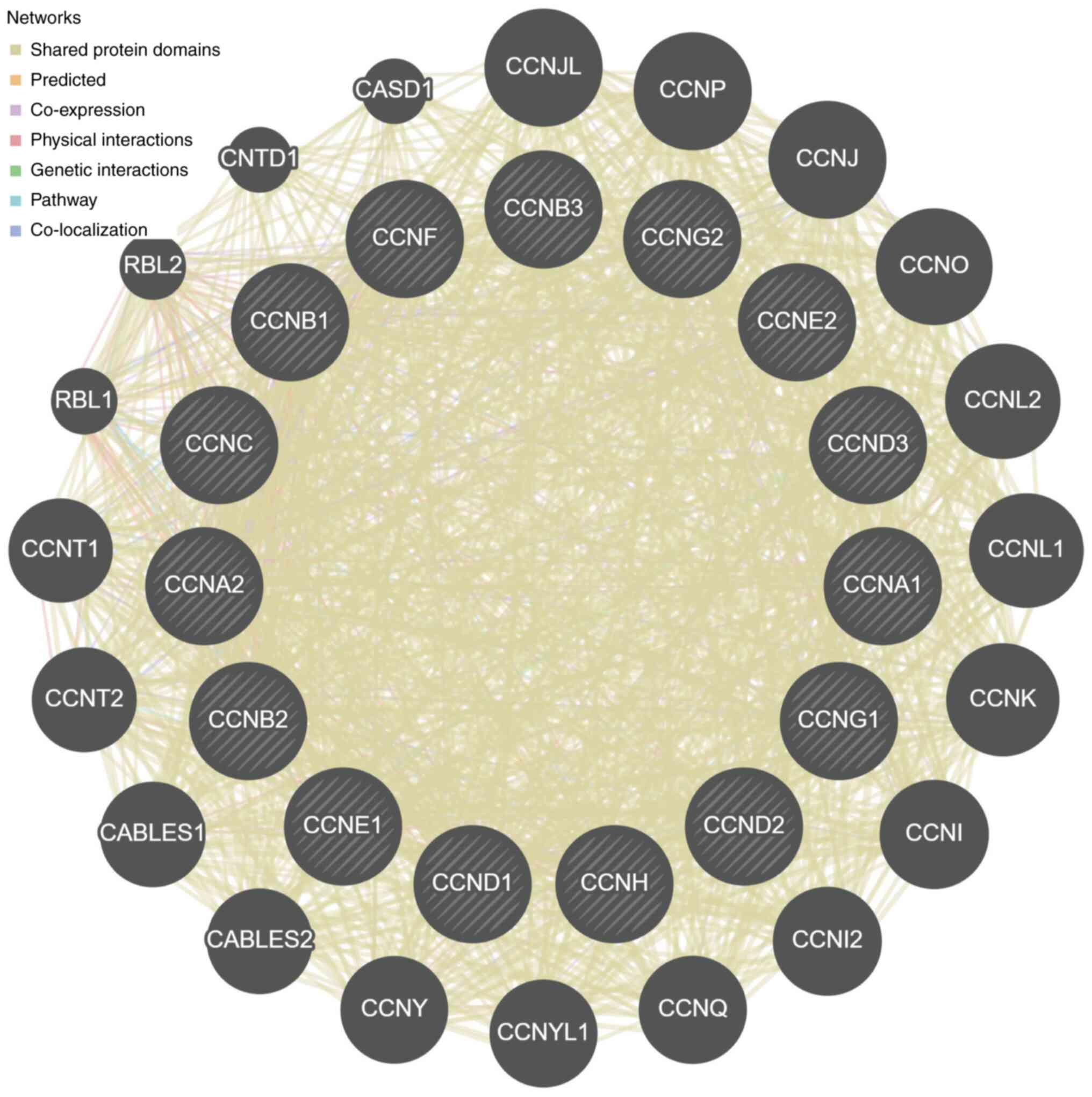
Based on the correlation analysis among the cyclin
genes (Fig. 6), for cyclin genes
and other interacting genes, the majority of shared protein domains
accounted for 69.48% of the total interacting network nodes,
predicted and co-expression accounted for 14.21 and 9.12%,
respectively, and the remaining accounted for <10%.
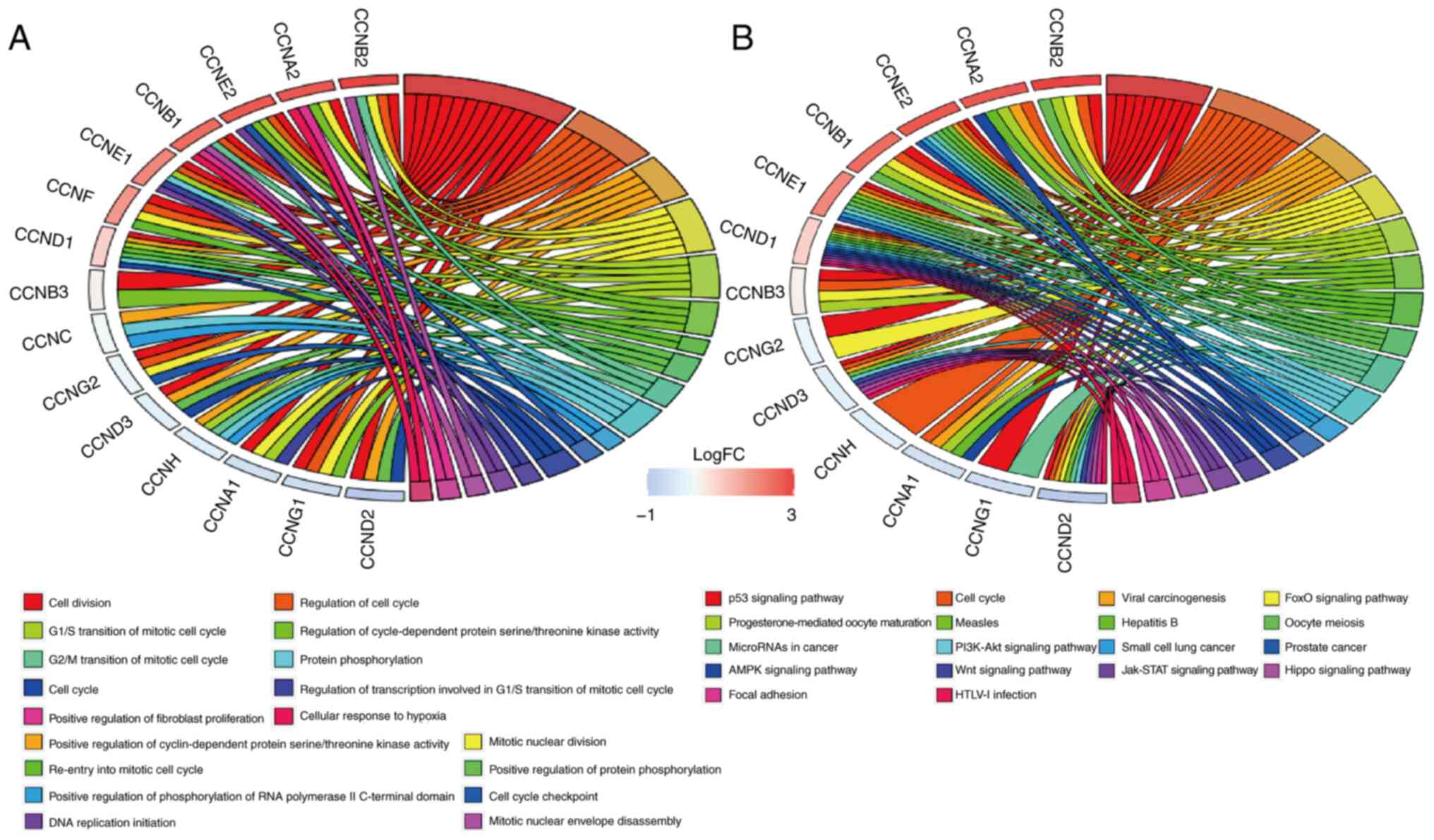
Cyclin gene functions and molecular
signaling pathways based on GO and KEGG enrichment analysis
Functional involvement of cyclin genes such as
biological processes (BP) was predicted using GO enrichment
analysis. The results showed that cell division (GO: 0051301),
regulation of cell cycle (GO: 0051726), positive regulation of
cyclin-dependent protein serine/threonine kinase activity (GO:
0045737), G1/S transition of mitotic cell cycle (GO: 0000082), and
regulation of cyclin-dependent protein serine/threonine kinase
activity (GO: 0000079) were significantly affected by cyclin genes
in BP (Fig. 7A).
KEGG enrichment analysis indicated that there were
18 molecular signaling pathways that cyclin genes participated in
(Fig. 7B). Signaling pathways
closely associated with breast cancer included the Wnt signaling
pathway (map04310), PI3K-Akt signaling pathway (map04151), p53
signaling pathway (map04115), microRNAs in cancer (map05206),
JAK-STAT signaling pathway (map04630), hippo signaling pathway
(map04390), cell cycle (map04110), and AMPK signaling pathway
(map04152); the cyclin genes were mostly involved in cell cycle
regulation and p53 signaling pathway.
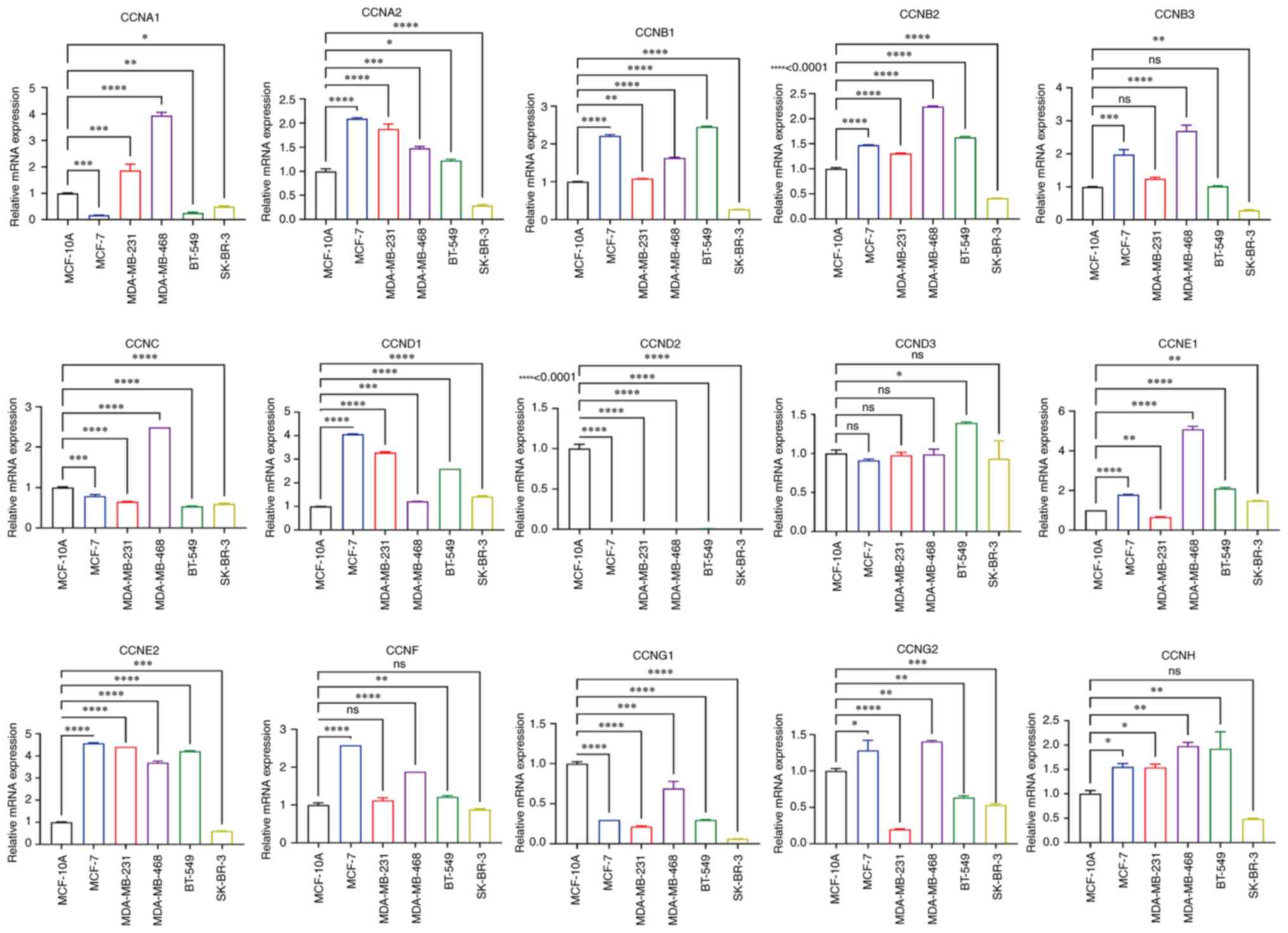
Expression of the cyclin genes in the
breast cancer cell lines
To verify cyclin gene expression in vitro,
RT-qPCR analysis of the breast cancer cell lines was performed
using MCF-10A cells as the standard control. The results
demonstrated that the mRNA expression levels of most of the cyclin
genes in SK-BR-3 were lower than that in control, except for
CCND1 and CCNE1 mRNA expression, whose expression was
slightly higher (Fig. 8). The mRNA
expression levels of CCNA2, CCNB1, CCNB2, CCNB3, CCND1, CCNE1,
CCNE2, and CCNF in the cancer cell lines were
significantly higher than that in MCF-10A cells, whereas
CCND3 and CCNG2 exhibited lower expression.
CCNA1 and CCNG2 mRNA expression varied based on the
breast cancer cell line, and CCND2 was barely expressed in
any of the cell lines. In summary, these results are generally
consistent with the gene expression analysis of the data from the
databases. However, the mRNA expression levels of CCNC and
CCNH in cell lines were lower and higher than that in
MCF-10A cells, respectively; their expression in tissues showed the
opposite trend compared with that in the cell lines.
Discussion
The cyclin genes play significant roles in a variety
of malignant tumors (4–11), and evaluation of the
transcriptional expression levels of the different cyclin genes may
be used to predict the prognosis and assess drug efficacy in
malignant tumors. However, studies assessing this are lacking due
to the numerous sub-types of cyclin genes and the high degree of
heterogeneity in breast cancer. Thus, the present study aimed to
analyze and discuss the value of the analysis of the diverse range
of cyclin genes in breast cancer at the transcriptional level,
using online public databases and bioinformatics methods to
identify potential biomarkers to improve survival and prognostic
prediction, and/or serve as therapeutic targets.
The functions of different cyclin genes vary in
enrichment analysis, and regulation, in general, is associated with
the phase of the cell cycle (67–72).
However, in the enrichment analysis of breast cancer-related
molecular signaling pathways, the regulation of CCNC and
CCNF was not involved. Therefore, certain cyclin genes,
especially CCNC and CCNF, may have greater value as
potential biomarkers based on our results.
First, the median transcriptional expression of
CCNA1 in breast cancer tissue was lower than that in
tumor-adjacent and normal breast tissues. The protein encoded by
CCNA1 is involved in the process of anthracycline
resistance, and the methylation status of CCNA1 and
CCND2 in normal tissues was lower in normal breast tissues
than that in tumors, and its expression in breast cancer was
downregulated after treatment (5,73,74).
These data indicated that high transcriptional expression of
CCNA1 and CCND2 may slow down the process of
acquisition of resistance to anthracyclines and prolong the effects
of the drug in breast cancer cells. Therefore, they may be used to
predict anthracycline/filomycin sensitivity. Secondly, there were
no significant differences in the mRNA expression levels of
CCNA1 between the clinical tumor stages of breast cancer.
The results showed that its high expression could significantly
increase the patients' RFS, while low expression increased the
patients' OS, although the difference was not significant. From the
analysis of TCGA data, CCNA1 mRNA was mainly highly
expressed in breast cancer caused by mutations. The contradictory
results may be due to the fact that mutations may frequently occur
due to alterations in breast cancer heterogeneity during disease
development or treatment (15).
Previous studies on the investigation of CCNA1 expression in
breast cancer are lacking; however, based on these results,
CCNA1 may exhibit potential as a biomarker for evaluating
drug tolerance in breast cancer.
CCNA2 is a relatively more reliable biomarker
for predicting the prognosis and assessing drug resistance. Gao
et al (18) confirmed that
CCNA2 was powerful in predicting the survival prognosis of
ER-positive breast cancer patients. Overexpression of CCNA2
mRNA could lead to Tamoxifen resistance. However, additional
evidence is required to understand the mechanism by which tamoxifen
resistance is reversed. In this study, the mRNA expression levels
of CCNA2 in breast cancer were significantly higher than
that in normal tissues, highlighting the potential of CCNA2
as a biomarker. Additionally, its expression in different clinical
stages of breast cancer varied significantly. Based on survival
length, lower transcriptional expression of CCNA2 was
significantly correlated with a better OS and RFS in cancer
patients, a result consistent with a previous study (75). Additionally, the mRNA expression
levels of CCNA2 in one dataset were significantly lower than
that in normal tissues, and analysis of data from TCGA also showed
that CCNA2 mRNA was lower in normal tissues. Furthermore,
CCNA2 mRNA expression was lower in SK-BR-3 cells than in
MCF-10A cells. Together, the results not only reflect the
heterogeneity of breast cancer (15), but also provide a novel direction
for the in-depth investigation of breast cancer. Thus, future
studies should analyze CCNA2 expression for predicting
prognosis and evaluating its clinical efficacy.
Similar to CCNA2, CCNB1, and CCNB2
show significant potential for predicting the prognosis and
assessing drug resistance. In both tumor tissues and cancer cell
lines, the mRNA expression levels of CCNB1 and CCNB2
were significantly higher than that in normal tissues and cells.
The low transcriptional expression of the two genes may indicate a
better OS in patients, consistent with previous studies (22,75–77).
It has also been found that their high transcriptional expression
can result in tamoxifen resistance and is positively correlated
with endocrine resistance (22,78).
Unlike CCNB1 and CCNB2, CCNB3 in
breast cancer has not been extensively studied and thus could not
be validated. Corresponding mRNA expression data was not available
in the Oncomine database. Transcriptional expression of
CCNB3 in breast cancer was slightly higher than that in the
normal tissues, and the in vitro analysis showed the same
trend. In TCGA analysis, the number of samples that showed
upregulated or downregulated expression of CCNB3 mRNA was
approximately the same. Although the P-value was >0.05, the
survival curve showed a trend such that the upregulated expression
of CCNB3 mRNA may predict an improved OS for patients. At
present, whether CCNB3 is suitable for future use to judge
the prognosis in patients with breast cancer and the efficacy of
drug response is still open for discussion, a definite correlation
between CCNB3 and breast cancer has been confirmed.
Additional studies are required to explore the biological function
of CCNB3.
Analysis of the GEPIA dataset showed that
CCNC mRNA expression in breast cancer was significantly
higher than that in normal tissues, similar to the primary
alteration in TCGA data. There were also a small number of
mutations with lower transcriptional expression. In the breast
cancer cell lines, CCNC mRNA expression was lower than that
in MCF-10A cells. Li et al (79) showed that CCNC regulates the
expression of the NOTCH1 oncogene to exert a tumor suppressor
effect. Thus far, a more direct explanation for the low expression
of CCNC mRNA is lacking. Tumor cells may adopt various means
to circumvent the inhibitory effects of CCNC to grow.
Additionally, CCNC is also involved in the transcriptional
regulation of SRC2 in human breast cancer, and their expression was
positively correlated (80). SRC
is upregulated in the early stages of breast intraductal carcinoma
compared with normal breast tissue. Decreased expression of SRC2
was associated with the progression of the disease and the
formation of invasive ductal carcinoma (81), thus it is highly likely that
CCNC mRNA expression may be similarly altered with SRC2. It
has been suggested that CCNC participates in the RB-E2F pathway
during the progression of breast cancer (31), and its expression may also vary as
the disease progresses. Therefore, it may serve as a potential
therapeutic target for repressing the cell cycle and a novel
prognostic predictor.
CCND1, an oncogene, is currently the most
extensively studied gene related to endocrine resistance (39,82).
Its expression was significantly increased in hormone
receptor-positive breast cancer (38) and was closely related to mechanisms
of endocrine resistance. CCND1 has been used in
HER-2-positive breast cancer as a continuous variable marker to
predict the therapeutic benefit of trastuzumab (83). In addition, CCND1 had a
different relationship with the aggressiveness of tumors and its
low transcriptional expression may enhance the migration of breast
cancer subgroups (43,84). In this study, CCND1 mRNA
expression in breast cancer was significantly increased. Its low
expression was shown to correlate with a better RFS in patients
with breast cancer, consistent with previous studies (36,37).
Analysis of the Oncomine dataset showed that the expression of
CCND1 mRNA in breast cancer was lower than that in normal
tissues, and the same trend of expression in breast cancer samples
was found in TCGA, providing evidence for why the low expression of
CCND1 can enhance the migration of breast cancer sub-types.
Based on the above findings, it is understandable that CCND1
may serve as a biomarker for breast cancer diagnosis, prognostic
prediction, drug efficacy, and resistance.
CCND2 mRNA expression was significantly lower
in breast cancer samples compared with the normal tissues in
several databases including Oncomine, GEPIA, and TCGA. Its high
expression was typically associated with a better OS and RFS,
consistent with previous reports (42,85).
Further studies showed that CCND2 methylation was negatively
correlated with its gene expression (43,86).
The integrated analysis of the correlation between CCND2
methylation and gene expression may be used to improve the
predictive performance of breast cancer outcomes, which may be
conducive to the expansion of the potential clinical applications
of CCND2.
The Oncomine dataset showed that CCND3
expression was highly upregulated in breast cancer and its mRNA
expression was upregulated in the TCGA dataset. Analysis of the
GEPIA dataset illustrated that CCND3 mRNA expression was
high in normal breast tissues, and the RT-qPCR results showed that
CCND3 mRNA expression in MCF-10A cells was higher than that
in other cancer cell lines. Based on the survival analysis it was
shown that upregulated expression of CCND3 mRNA was
associated with a better OS. Conversely, Justenhoven et al
(87) found that the low
expression of E2F2, CCND1, and CCND3 genes reduced
the levels of factors that downregulate HER-2, thereby allowing
upregulated expression of HER-2 in tumors, and it is hypothesized
that these three gene sub-types are potential indicators of HER-2
status in breast cancer. Combining all the results of the databases
together, it can be suggested that CCND3 plays various
functions in different breast cancer sub-types. Since the current
exploration of CCND3 is still limited, further research is required
to better understand the potential of CCND3 as a possible biomarker
for prognostic prediction and improving drug efficacy.
The expression of CCNE1 and CCNE2,
two subtypes of the CCNE gene, was approximately the same.
The expression of both of these genes was upregulated in breast
cancer, a result that was confirmed by RT-qPCR. Patients with low
transcriptional levels of both genes in breast cancer had a
significantly better OS. One difference between the two subtypes
was that the expression of CCNE1 was correlated with the
clinical tumor stage. The clinical trials PALPMA-3 and POP
(88) demonstrated that the
efficacy of Palbociclib decreased when CCNE1 mRNA expression
was increased. Therefore, CCNE1 mRNA is predictive of
metastatic tumors. Previously it has been shown that the
amplification of CCNE1 was associated with a poorer OS of
metastatic triple-negative breast cancer, and it was speculated
that this phenomenon may be caused by CCNE1
upregulation-induced chemotherapeutic resistance (89). Moreover, both CCNE1 and
CCNE2 could be used as independent prognostic markers for
patients with lymph node-negative breast cancer (90). Thus, as typical oncogenes,
CCNE1 and CCNE2 may be employed as factors for the
prognostic prediction and for countering drug resistance mechanisms
in breast cancer.
The functional involvement of CCNF in breast
cancer remains unclear. Studies have used CCNF as a target
gene for RNAi targeting in breast cancer (91,92).
CCNF may be involved in the non-KEGG-enriched molecular
signaling pathways to regulate drug resistance in breast cancer.
Additionally, the CCNF protein possesses a unique characteristic,
an F-BOX homologous sequence, which is suggestive of CCNF
possessing a similar function to that of F-BOX. As for F-BOX, high
mRNA expression of its subtypes (FBXO1, FBXO31, and
FBXO5) may serve as biomarkers for predicting prognosis, and
their expression was significantly correlated with a poor prognosis
in patients with breast cancer (93). Therefore, it was hypothesized that
CCNF may be used as a biomarker similar to F-BOX. In this study,
CCNF mRNA expression was upregulated in breast cancer, and
it was primarily observed as gene amplification and the only
example of a fusion alteration found in TCGA in the present study.
The clinical stage of breast cancer was significantly related to
the differential expression of CCNF, and patients with low
CCNF mRNA expression had a significantly better OS.
Therefore, CCNF has potential as an advanced biomarker for
prognostic prediction and as a gene target for counteracting
therapeutic tolerance.
CCNG1 is the only cyclin gene that has both
positive and negative effects on cellular growth (94,95)
as the cyclinG1 protein encoded by CCNG1 is highly unstable.
Tian et al (96) found out
that the administration of estrogen and progesterone promoted cell
proliferation and upregulated the expression of cyclinG1 in MCF-7
cells. Estradiol and progesterone-mediated cell viability and
clonal ability were restricted after CCNG1 was knocked down
using shRNA, suggesting that estradiol and progesterone promote
cell proliferation in breast cancer partially by inducing cyclinG1
expression. In the present study, CCNG1 mRNA expression was
low in breast cancer, and cancer patients with upregulated
CCNG1 expression had a better OS. CCNG1 expression
was also significantly associated with the clinicopathological
stage. Taken together, despite the instability of the protein
encoded by CCNG1, it may still serve as a biomarker for
evaluating the prognosis of breast cancer and as a therapeutic
target in certain subtypes of breast cancer.
As an estrogen-regulated gene, CCNG2 has
previously been used as a target gene for judging the efficacy of
endocrine-based medicine, as low CCNG2 mRNA expression is
indicative of a poorer prognosis. In the present study,
CCNG2 mRNA expression in breast cancer tissues was higher
than that in normal tissues although the difference was not
statistically significant. RT-qPCR analysis showed that
CCNG2 mRNA expression was higher in MCF-7 cells and lower in
MDA-MB-231 and BT-549 cells, and patients with high transcriptional
expression of CCNG2 had a better OS. Consistent with a
previous study (57), CCNG2
expression was downregulated in highly aggressive breast cancer
subtypes with poor prognoses. Consequently, CCNG2 may
exhibit potential for the prognostic prediction of ER-positive
breast cancer and as a therapeutic target. However, additional
studies on CCNG2 in other subtypes of breast cancer are required to
examine its value as a biomarker.
Studies on the expression or the function of
CCNH in breast cancer are limited. Shahi et al
(97) performed whole-exome
sequencing of 492 cancer-related genes as well as high-risk genes
and mutation analysis of BRCA1 and BRCA2 negative breast cancer
patients. In total, 31 gene variants/genes were identified for
breast cancer susceptibility predisposition including CCNH.
However, CCNH has not been extensively studied with regard
to familial breast cancer. In the present study, CCNH
expression was low in breast cancer, and upregulated expression was
predictive of a better OS. It has been indirectly shown that the
increased expression of CCNH in ER-positive breast cancer
was associated with indicators of good prognosis (98). Yet, RT-qPCR analysis of breast
cancer cell lines showed higher CCNH mRNA expression than
that in MCF-10A. At present, there is currently no sufficient
evidence to explain this phenomenon. Thus, further studies are
required to determine the value of CCNH as a biomarker for
evaluating the prognosis and exploring the mechanisms of drug
action.
In conclusion, we systematically analyzed the
expression of the cyclin genes in breast cancer based on its value
in predicting prognosis and evaluating drug efficacy, enabling a
deeper understanding of the heterogeneity of breast cancer. Our
results indicated that CCNA2, CCNB1, CCNB2, CCND1, CCND2,
CCNE1, and CCNE2 may be mature and valid biomarkers in
predicting prognosis and judging efficacy. CCNG1 and
CCNG2 may be used as biological markers in specific types of
breast cancer. At present, there are few relevant studies on
CCNA1, CCNB3, CCNC, CCND3, CCNF, and CCNH in breast
cancer; however, the results of the present and previous studies
showed their potential as unique predictors of prognosis and
potential targets for countering therapeutic resistance. Together,
the discovery of novel biomarkers and gene targets will facilitate
the exploration of more personalized therapeutic strategies;
however, further studies are required to verify their biological
functions.
Acknowledgements
Not applicable.
Funding
This study was supported by funding from the National Nature
Science Foundation of China, The Third Affiliated Hospital of
Kunming Medical University: Yunnan Cancer Center, Kunming, Yunnan,
China (grant nos. 81760480 and 81960479).
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
NQL, WHC and JN confirm the authenticity of all the
raw data. NQL participated in the design of study, and was
responsible for collecting and interpreting data, performing the
bioinformatics analysis, visualizing the presentation, and drafting
and revising the original manuscript. WHC collected the data,
performed the bioinformatics analysis and drafted the original
manuscript. XW and JC participated in data curation, confirmed and
analyzed the data, and assisted in drafting the revised manuscript.
JN mainly provided the research conceptualization and funding
support, supervised and assisted in designing the study, and
participated in the revision of the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cai Z and Liu Q: Cell cycle regulation in
treatment of breast cancer. Adv Exp Med Biol. 1026:251–270. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Steeg PS and Zhou Q: Cyclins and breast
cancer. Breast Cancer Res Treat. 52:17–28. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bai C, Richman R and Elledge SJ: Human
cyclin F. EMBO J. 13:6087–6098. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ye C, Wang J, Wu P, Li X and Chai Y:
Prognostic role of cyclin B1 in solid tumors: A meta-analysis.
Oncotarget. 8:2224–2232. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee YS, Ryu SW, Bae SJ, Park TH, Kwon K,
Noh YH and Kim SY: Cross-platform meta-analysis of multiple gene
expression profiles identifies novel expression signatures in
acquired anthracycline-resistant breast cancer. Oncol Rep.
33:1985–1993. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miftakhova R, Hedblom A, Semenas J,
Robinson B, Simoulis A, Malm J, Rizvanov A, Heery DM, Mongan NP,
Maitland NJ, et al: Cyclin A1 and P450 aromatase promote metastatic
homing and growth of stem-like prostate cancer cells in the bone
marrow. Cancer Res. 76:2453–2464. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li R, Jiang X, Zhang Y, Wang S, Chen X, Yu
X, Ma J and Huang X: Cyclin B2 overexpression in human
hepatocellular carcinoma is associated with poor prognosis. Arch
Med Res. 50:10–17. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dorn J, Spatz H, Schmieder M, Barth TF,
Blatz A, Henne-Bruns D, Knippschild U and Kramer K: Cyclin H
expression is increased in GIST with very-high risk of malignancy.
BMC Cancer. 10:3502010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang KC, Yang J, Ng MC, Ng SK, Welch WR,
Muto MG, Berkowitz RS and Ng SW: Cyclin A1 expression and
paclitaxel resistance in human ovarian cancer cells. Eur J Cancer.
67:152–163. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chujan S, Kitkumthorn N, Siriangkul S and
Mutirangura A: CCNA1 promoter methylation: A potential marker for
grading Papanicolaou smear cervical squamous intraepithelial
lesions. Asian Pac J Cancer Prev. 15:7971–7975. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takashima S, Saito H, Takahashi N, Imai K,
Kudo S, Atari M, Saito Y, Motoyama S and Minamiya Y: Strong
expression of cyclin B2 mRNA correlates with a poor prognosis in
patients with non-small cell lung cancer. Tumour Biol.
35:4257–4265. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wiseman M: The second world cancer
research fund/American institute for cancer research expert report.
Food, nutrition, physical activity, and the prevention of cancer: A
global perspective. Proc Nutr Soc. 67:253–256. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harbeck N and Gnant M: Breast cancer.
Lancet. 389:1134–1150. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeng X, Liu C, Yao J, Wan H, Wan G, Li Y
and Chen N: Breast cancer stem cells, heterogeneity, targeting
therapies and therapeutic implications. Pharmacol Res.
163:1053202021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ochsenreither S, Majeti R, Schmitt T,
Stirewalt D, Keilholz U, Loeb KR, Wood B, Choi YE, Bleakley M,
Warren EH, et al: Cyclin-A1 represents a new immunogenic targetable
antigen expressed in acute myeloid leukemia stem cells with
characteristics of a cancer-testis antigen. Blood. 119:5492–5501.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang R, Nakamaki T, Lübbert M, Said J,
Sakashita A, Freyaldenhoven BS, Spira S, Huynh V, Müller C and
Koeffler HP: Cyclin A1 expression in leukemia and normal
hematopoietic cells. Blood. 93:2067–2074. 1999.PubMed/NCBI
|
|
18
|
Gao T, Han Y, Yu L, Ao S, Li Z and Ji J:
CCNA2 is a prognostic biomarker for ER+ breast cancer and tamoxifen
resistance. PLoS One. 9:e917712014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hein JB and Nilsson J: Interphase
APC/C-Cdc20 inhibition by cyclin A2-Cdk2 ensures efficient mitotic
entry. Nat Commun. 7:109752016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pei J, Zhang J, Yang X, Wu Z, Sun C, Wang
Z and Wang B: NEK5 promotes breast cancer cell proliferation
through up-regulation of cyclin A2. Mol Carcinog. 58:933–943. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding K, Li W, Zou Z, Zou X and Wang C:
CCNB1 is a prognostic biomarker for ER+ breast cancer. Med
Hypotheses. 83:359–564. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu HY, Liu YY, Yang F, Zhang L, Zhang FL,
Hu X, Shao ZM and Li DQ: Acetylation of MORC2 by NAT10 regulates
cell-cycle checkpoint control and resistance to DNA-damaging
chemotherapy and radiotherapy in breast cancer. Nucleic Acids Res.
48:3638–3656. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu AQ, Wang ZX, Wu W, Chen KY, Yan SR and
Mao ZB: Circular RNA CircCCNB1 sponges micro RNA-449a to inhibit
cellular senescence by targeting CCNE2. Aging (Albany NY).
11:10220–10241. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shubbar E, Kovács A, Hajizadeh S, Parris
TZ, Nemes S, Gunnarsdóttir K, Einbeigi Z, Karlsson P and Helou K:
Elevated cyclin B2 expression in invasive breast carcinoma is
associated with unfavorable clinical outcome. BMC Cancer. 13:12013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qian X, Song X, He Y, Yang Z, Sun T, Wang
J, Zhu G, Xing W and You C: CCNB2 overexpression is a poor
prognostic biomarker in Chinese NSCLC patients. Biomed
Pharmacother. 74:222–227. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gao Z, Man X, Li Z, Bi J, Liu X, Li Z, Zhu
Y, Zhang Z and Kong C: Expression profiles analysis identifies the
values of carcinogenesis and the prognostic prediction of three
genes in adrenocortical carcinoma. Oncol Rep. 41:2440–2452.
2019.PubMed/NCBI
|
|
27
|
Lozano JC, Perret E, Schatt P, Arnould C,
Peaucellier G and Picard A: Molecular cloning, gene localization,
and structure of human cyclin B3. Biochem Biophys Res Commun.
291:406–413. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Han H, Bertrand KC, Patel KR, Fisher KE,
Roy A, Muscal JA and Venkatramani R: BCOR-CCNB3 fusion-positive
clear cell sarcoma of the kidney. Pediatr Blood Cancer.
67:e281512020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoshida A, Arai Y, Hama N, Chikuta H,
Bando Y, Nakano S, Kobayashi E, Shibahara J, Fukuhara H, Komiyama
M, et al: Expanding the clinicopathologic and molecular spectrum of
BCOR-associated sarcomas in adults. Histopathology. 76:509–520.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shibayama T, Okamoto T, Nakashima Y, Kato
T, Sakurai T, Minamiguchi S, Kataoka TR, Shibuya S, Yoshizawa A,
Toguchida J and Haga H: Screening of BCOR-CCNB3 sarcoma using
immunohistochemistry for CCNB3: A clinicopathological report of
three pediatric cases. Pathol Int. 65:410–414. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ren S and Rollins BJ: Cyclin C/cdk3
promotes Rb-dependent G0 exit. Cell. 117:239–251. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miyata Y, Liu Y, Jankovic V, Sashida G,
Lee JM, Shieh JH, Naoe T, Moore M and Nimer SD: Cyclin C regulates
human hematopoietic stem/progenitor cell quiescence. Stem Cells.
28:308–317. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu W and Ji JY: Dysregulation of CDK8 and
cyclin C in tumorigenesis. J Genet Genomics. 38:439–452. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu YN, Yip GW, Tan PH, Thike AA, Matsumoto
K, Tsujimoto M and Bay BH: Y-box binding protein 1 is up-regulated
in proliferative breast cancer and its inhibition deregulates the
cell cycle. Int J Oncol. 37:483–492. 2010.PubMed/NCBI
|
|
35
|
Roy PG and Thompson AM: Cyclin D1 and
breast cancer. Breast. 15:718–127. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Elsheikh S, Green AR, Aleskandarany MA,
Grainge M, Paish CE, Lambros MB, Reis-Filho JS and Ellis IO: CCND1
amplification and cyclin D1 expression in breast cancer and their
relation with proteomic subgroups and patient outcome. Breast
Cancer Res Treat. 109:325–335. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He Q, Wu J, Liu XL, Ma YH, Wu XT, Wang WY
and An HX: Clinicopathological and prognostic significance of
cyclin D1 amplification in patients with breast cancer: A
meta-analysis. J BUON. 22:1209–1216. 2017.PubMed/NCBI
|
|
38
|
Villegas SL, Darb-Esfahani S, von
Minckwitz G, Huober J, Weber K, Marmé F, Furlanetto J, Schem C,
Pfitzner BM, Lederer B, et al: Expression of cyclin D1 protein in
residual tumor after neoadjuvant chemotherapy for breast cancer.
Breast Cancer Res Treat. 168:179–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shi Q, Li Y, Li S, Jin L, Lai H, Wu Y, Cai
Z, Zhu M, Li Q, Li Y, et al: LncRNA DILA1 inhibits cyclin D1
degradation and contributes to tamoxifen resistance in breast
cancer. Nat Commun. 11:55132020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pors J, Naso J, Berg K and Churg A: Cyclin
D1 immunohistochemical staining to separate benign from malignant
mesothelial proliferations. Mod Pathol. 33:312–318. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kwapisz D: Cyclin-dependent kinase 4/6
inhibitors in breast cancer: Palbociclib, ribociclib, and
abemaciclib. Breast Cancer Res Treat. 166:41–54. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hung CS, Wang SC, Yen YT, Lee TH, Wen WC
and Lin RK: Hypermethylation of CCND2 in lung and breast cancer is
a potential biomarker and drug target. Int J Mol Sci. 19:30962018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Callahan CL, Wang Y, Marian C, Weng DY,
Eng KH, Tao MH, Ambrosone CB, Nie J, Trevisan M, Smiraglia D, et
al: DNA methylation and breast tumor clinicopathological features:
The western New York exposures and breast cancer (WEB) study.
Epigenetics. 11:643–652. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ding ZY, Li R, Zhang QJ, Wang Y, Jiang Y,
Meng QY, Xi QL and Wu GH: Prognostic role of cyclin D2/D3 in
multiple human malignant neoplasms: A systematic review and
meta-analysis. Cancer Med. 8:2717–2729. 2019.PubMed/NCBI
|
|
45
|
Luhtala S, Staff S, Tanner M and Isola J:
Cyclin E amplification, over-expression, and relapse-free survival
in HER-2-positive primary breast cancer. Tumour Biol. 37:9813–9823.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Keyomarsi K, Tucker SL, Buchholz TA,
Callister M, Ding Y, Hortobagyi GN, Bedrosian I, Knickerbocker C,
Toyofuku W, Lowe M, et al: Cyclin E and survival in patients with
breast cancer. N Engl J Med. 347:1566–1575. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee C, Fernandez KJ, Alexandrou S, Sergio
CM, Deng N, Rogers S, Burgess A and Caldon CE: Cyclin E2 promotes
whole genome doubling in breast cancer. Cancers (Basel).
12:22682020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Peek GW and Tollefsbol TO: Combinatorial
PX-866 and raloxifene decrease Rb phosphorylation, cyclin E2
transcription, and proliferation of MCF-7 breast cancer cells. J
Cell Biochem. 117:1688–1696. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lindskog C: The potential clinical impact
of the tissue-based map of the human proteome. Expert Rev
Proteomics. 12:213–215. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fu J, Qiu H, Cai M, Pan Y, Cao Y, Liu L,
Yun J and Zhang CZ: Low cyclin F expression in hepatocellular
carcinoma associates with poor differentiation and unfavorable
prognosis. Cancer Sci. 104:508–515. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhao L, Jiang L, He L, Wei Q, Bi J, Wang
Y, Yu L, He M, Zhao L and Wei M: Identification of a novel cell
cycle-related gene signature predicting survival in patients with
gastric cancer. J Cell Physiol. 234:6350–6360. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Noh JM, Kim J, Cho DY, Choi DH, Park W and
Huh SJ: Exome sequencing in a breast cancer family without BRCA
mutation. Radiat Oncol J. 33:149–154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tamura K, Kanaoka Y, Jinno S, Nagata A,
Ogiso Y, Shimizu K, Hayakawa T, Nojima H and Okayama H: Cyclin G: A
new mammalian cyclin with homology to fission yeast Cig1. Oncogene.
8:2113–2118. 1993.PubMed/NCBI
|
|
54
|
Zimmermann M, Arachchige-Don AP, Donaldson
MS, Patriarchi T and Horne MC: Cyclin G2 promotes cell cycle arrest
in breast cancer cells responding to fulvestrant and metformin and
correlates with patient survival. Cell Cycle. 15:3278–3295. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu D, Han B, Guo L and Fan Z: Molecular
mechanisms associated with breast cancer based on integrated gene
expression profiling by bioinformatics analysis. J Obstet Gynaecol.
36:615–621. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Knowles LM and Smith JW: Genome-wide
changes accompanying knockdown of fatty acid synthase in breast
cancer. BMC Genomics. 8:1682007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Miller LD, Smeds J, George J, Vega VB,
Vergara L, Ploner A, Pawitan Y, Hall P, Klaar S, Liu ET and Bergh
J: An expression signature for p53 status in human breast cancer
predicts mutation status, transcriptional effects, and patient
survival. Proc Natl Acad Sci USA. 102:13550–13555. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Mao L, Ling X and Chen J: Cyclin H
regulates lung cancer progression as a carcinoma inducer. Comput
Math Methods Med. 2021:66460772021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45((W1)):
W98–W102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Nagy Á, Munkácsy G and Győrffy B:
Pancancer survival analysis of cancer hallmark genes. Sci Rep.
11:60472021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hoadley KA, Yau C, Hinoue T, Wolf DM,
Lazar AJ, Drill E, Shen R, Taylor AM, Cherniack AD, Thorsson V, et
al: Cell-of-origin patterns dominate the molecular classification
of 10,000 tumors from 33 types of cancer. Cell. 173:291–304.e6.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
R Core Team: R: A language and environment
for statistical computing. R Foundation for Statistical Computing;
Vienna: 2012
|
|
66
|
RStudio Team, . RStudio: Integrated
Development for R. RStudio Inc. Boston, MA: 2015
|
|
67
|
Tang T, Guo C, Xia T, Zhang R, Zen K, Pan
Y and Jin L: LncCCAT1 promotes breast cancer stem cell function
through activating WNT/β-catenin signaling. Theranostics.
9:7384–7402. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Shao F, Pang X and Baeg GH: Targeting the
JAK/STAT signaling pathway for breast cancer. Curr Med Chem.
28:5137–5151. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Costa RLB, Han HS and Gradishar WJ:
Targeting the PI3K/AKT/mTOR pathway in triple-negative breast
cancer: A review. Breast Cancer Res Treat. 169:397–406. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Li X, Zeng Z, Wang J, Wu Y, Chen W, Zheng
L, Xi T, Wang A and Lu Y: MicroRNA-9 and breast cancer. Biomed
Pharmacother. 122:1096872020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Qiao K, Ning S, Wan L, Wu H, Wang Q, Zhang
X, Xu S and Pang D: LINC00673 is activated by YY1 and promotes the
proliferation of breast cancer cells via the miR-515-5p/MARK4/Hippo
signaling pathway. J Exp Clin Cancer Res. 38:4182019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lee MG, Kwon YS, Nam KS, Kim SY, Hwang IH,
Kim S and Jang H: Chaga mushroom extract induces autophagy via the
AMPK-mTOR signaling pathway in breast cancer cells. J
Ethnopharmacol. 274:1140812021. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Woo SH, Seo SK, An S, Choe TB, Hong SI,
Lee YH and Park IC: Implications of caspase-dependent proteolytic
cleavage of cyclin A1 in DNA damage-induced cell death. Biochem
Biophys Res Commun. 453:438–442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Klajic J, Busato F, Edvardsen H, Touleimat
N, Fleischer T, Bukholm I, Børresen-Dale AL, Lønning PE, Tost J and
Kristensen VN: DNA methylation status of key cell-cycle regulators
such as CDKNA2/p16 and CCNA1 correlates with treatment response to
doxorubicin and 5-fluorouracil in locally advanced breast tumors.
Clin Cancer Res. 20:6357–6366. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Deng JL, Xu YH and Wang G: Identification
of potential crucial genes and key pathways in breast cancer using
bioinformatic analysis. Front Genet. 10:6952019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tang J, Kong D, Cui Q, Wang K, Zhang D,
Gong Y and Wu G: Prognostic genes of breast cancer identified by
gene co-expression network analysis. Front Oncol. 8:3742018.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Jayanthi VSPKSA, Das AB and Saxena U:
Grade-specific diagnostic and prognostic biomarkers in breast
cancer. Genomics. 112:388–396. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhou H, Lv Q and Guo Z: Transcriptomic
signature predicts the distant relapse in patients with ER+ breast
cancer treated with tamoxifen for five years. Mol Med Rep.
17:3152–3157. 2018.PubMed/NCBI
|
|
79
|
Li N, Fassl A, Chick J, Inuzuka H, Li X,
Mansour MR, Liu L, Wang H, King B, Shaik S, et al: Cyclin C is a
haploinsufficient tumour suppressor. Nat Cell Biol. 16:1080–1091.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Bozickovic O, Hoang T, Fenne IS, Helland
T, Skartveit L, Ouchida M, Mellgren G and Sagen JV: Cyclin C
interacts with steroid receptor coactivator 2 and upregulates cell
cycle genes in MCF-7 cells. Biochim Biophys Acta. 1853:2383–2391.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kurebayashi J, Otsuki T, Kunisue H, Tanaka
K, Yamamoto S and Sonoo H: Expression levels of estrogen
receptor-alpha, estrogen receptor-beta, coactivators, and
corepressors in breast cancer. Clin Cancer Res. 6:512–518.
2000.PubMed/NCBI
|
|
82
|
Butt AJ, McNeil CM, Musgrove EA and
Sutherland RL: Downstream targets of growth factor and oestrogen
signalling and endocrine resistance: The potential roles of c-Myc,
cyclin D1 and cyclin E. Endocr Relat Cancer. 12 (Suppl 1):S47–S59.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Filipits M, Dafni U, Gnant M,
Polydoropoulou V, Hills M, Kiermaier A, de Azambuja E, Larsimont D,
Rojo F, Viale G, et al: Association of p27 and cyclin D1 expression
and benefit from adjuvant trastuzumab treatment in HER2-positive
early breast cancer: A TransHERA study. Clin Cancer Res.
24:3079–3086. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Tobin NP, Sims AH, Lundgren KL, Lehn S and
Landberg G: Cyclin D1, Id1 and EMT in breast cancer. BMC Cancer.
11:4172011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Fischer H, Chen J, Skoog L and Lindblom A:
Cyclin D2 expression in familial and sporadic breast cancer. Oncol
Rep. 9:1157–1161. 2002.PubMed/NCBI
|
|
86
|
Li Z, Heng J, Yan J, Guo X, Tang L, Chen
M, Peng L, Wu Y, Wang S, Xiao Z, et al: Integrated analysis of gene
expression and methylation profiles of 48 candidate genes in breast
cancer patients. Breast Cancer Res Treat. 160:371–383. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Justenhoven C, Pierl CB, Haas S, Fischer
HP, Hamann U, Baisch C, Harth V, Spickenheuer A, Rabstein S,
Vollmert C, et al: Polymorphic loci of E2F2, CCND1 and CCND3 are
associated with HER2 status of breast tumors. Int J Cancer.
124:2077–2081. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Turner NC, Liu Y, Zhu Z, Loi S, Colleoni
M, Loibl S, DeMichele A, Harbeck N, André F, Bayar MA, et al:
Cyclin E1 expression and palbociclib efficacy in previously treated
hormone receptor-positive metastatic breast cancer. J Clin Oncol.
37:1169–1178. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Zhao ZM, Yost SE, Hutchinson KE, Li SM,
Yuan YC, Noorbakhsh J, Liu Z, Warden C, Johnson RM, Wu X, et al:
CCNE1 amplification is associated with poor prognosis in patients
with triple negative breast cancer. BMC Cancer. 19:962019.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Sieuwerts AM, Look MP, Meijer-van Gelder
ME, Timmermans M, Trapman AM, Garcia RR, Arnold M, Goedheer AJ, de
Weerd V, Portengen H, et al: Which cyclin E prevails as prognostic
marker for breast cancer? Results from a retrospective study
involving 635 lymph node-negative breast cancer patients. Clin
Cancer Res. 12:3319–3328. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Seyhan AA, Varadarajan U, Choe S, Liu W
and Ryan TE: A genome-wide RNAi screen identifies novel targets of
neratinib resistance leading to identification of potential drug
resistant genetic markers. Mol Biosyst. 8:1553–1570. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Gupta ED, Pachauri M, Ghosh PC and Rajam
MV: Targeting polyamine biosynthetic pathway through RNAi causes
the abrogation of MCF 7 breast cancer cell line. Tumour Biol.
37:1159–1171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Wang X, Zhang T, Zhang S and Shan J:
Prognostic values of F-box members in breast cancer: An online
database analysis and literature review. Biosci Rep.
39:BSR201809492019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Piscopo DM and Hinds PW: A role for the
cyclin box in the ubiquitin-mediated degradation of cyclin G1.
Cancer Res. 68:5581–5590. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Liu F, Gao X, Yu H, Yuan D, Zhang J, He Y
and Yue L: Effects of expression of exogenous cyclin G1 on
proliferation of human endometrial carcinoma cells. Chin J Physiol.
56:83–89. 2013.PubMed/NCBI
|
|
96
|
Tian JM, Ran B, Zhang CL, Yan DM and Li
XH: Estrogen and progesterone promote breast cancer cell
proliferation by inducing cyclin G1 expression. Braz J Med Biol
Res. 51:1–7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Shahi RB, De Brakeleer S, Caljon B,
Pauwels I, Bonduelle M, Joris S, Fontaine C, Vanhoeij M, Van Dooren
S, Teugels E and De Grève J: Identification of candidate cancer
predisposing variants by performing whole-exome sequencing on index
patients from BRCA1 and BRCA2-negative breast cancer families. BMC
Cancer. 19:3132019. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Patel H, Abduljabbar R, Lai CF, Periyasamy
M, Harrod A, Gemma C, Steel JH, Patel N, Busonero C, Jerjees D, et
al: Expression of CDK7, cyclin H, and MAT1 is elevated in breast
cancer and is prognostic in estrogen receptor-positive breast
cancer. Clin Cancer Res. 22:5929–5938. 2016. View Article : Google Scholar : PubMed/NCBI
|