Introduction
The ubiquitin proteasome system (UPS) is the main
pathway for protein degradation in eukaryotic cells, participating
in the degradation of >80% of proteins (1,2). The
UPS is highly selective and serves a critical role in maintaining
protein homeostasis. The output of the ubiquitin system is mainly
manifested in two forms, namely, the control of protein turnover by
providing proteasomal and lysosomal targeting signals, and the
control of cellular signaling networks by the regulation of protein
interactions and activity (3–5).
Ubiquitination is one of the most common and
important post-translational modifications, and can alter the
stability, localization and activity of target proteins (6,7).
Ubiquitination is most commonly demonstrated in the binding of
ubiquitin molecules and polyubiquitin chains to lysine residues of
substrate proteins. Ubiquitin itself has eight ubiquitination
sites, seven lysine residues (K6, K11, K27, K29, K33, K48 and K63)
and an amino-terminal methionine (M1), all of which can be involved
in the formation of polyubiquitin chains (8). There are various forms of
polyubiquitination; together with homotypic and heterotypic chains,
the complexity of ubiquitin coding can be enriched via association
with ubiquitin-like molecules, such as small ubiquitin-related
modifier, neural precursor cell expressed developmentally
downregulated 8 and interferon-stimulated gene product 15, and can
be modified by phosphorylation and acetylation (8–11).
The most dominant and abundant forms of polyubiquitination are K48-
and K63-polyubiquitination (12).
Ubiquitination requires the sequential action of three enzymes,
ubiquitin-activating enzyme (E1), ubiquitin-binding enzyme (E2) and
ubiquitin-ligase (E3). E1 catalyzes the ATP-dependent activation of
ubiquitin and the formation of a thioester bond between the
ubiquitin C-terminus and the catalytic cysteine on E1. Ubiquitin is
then transferred to a catalytic cysteine of one of the ~40 E2s and
then to the substrate via E3 (13).
Currently, ~100 human deubiquitinating enzymes
(DUBs) have been identified and they can be classified into seven
subfamilies, including six cysteine protease families
[ubiquitin-specific proteases (USPs), ubiquitin C-terminal
hydrolases, ovarian tumor proteases, Machado-Joseph disease
proteases, zinc finger-containing ubiquitin peptidase 1 and motif
interacting with Ub-containing novel DUB family] and one
metalloprotease family (JAB1/MPN/Mov34 metalloprotease). DUBs, as
key factors in the deubiquitination process, can catalyze the
separation of the isopeptide bond between the glycine site of
ubiquitin and the lysine site of the target protein to facilitate
deubiquitination (14).
Ubiquitin-specific peptidase 44 (USP44) has been
extensively studied as a member of the USP family since it was
first identified in 2004 (15).
The deubiquitination activity of USP44 has been previously
reported, as has its cooperation with E3S (a ubiquitin ligase) to
regulate the function and stability of target proteins. USP44 has
been reported to be involved in the regulation of various
physiological functions and pathological processes, including
sister chromatid separation, stem cell differentiation and tumor
progression (16–18). In the present review, a
comprehensive overview of recent advances regarding USP44, focusing
on its physiological roles in various cellular activities and its
pathophysiological roles in tumor progression, is presented, and
its potential therapeutic potential is highlighted.
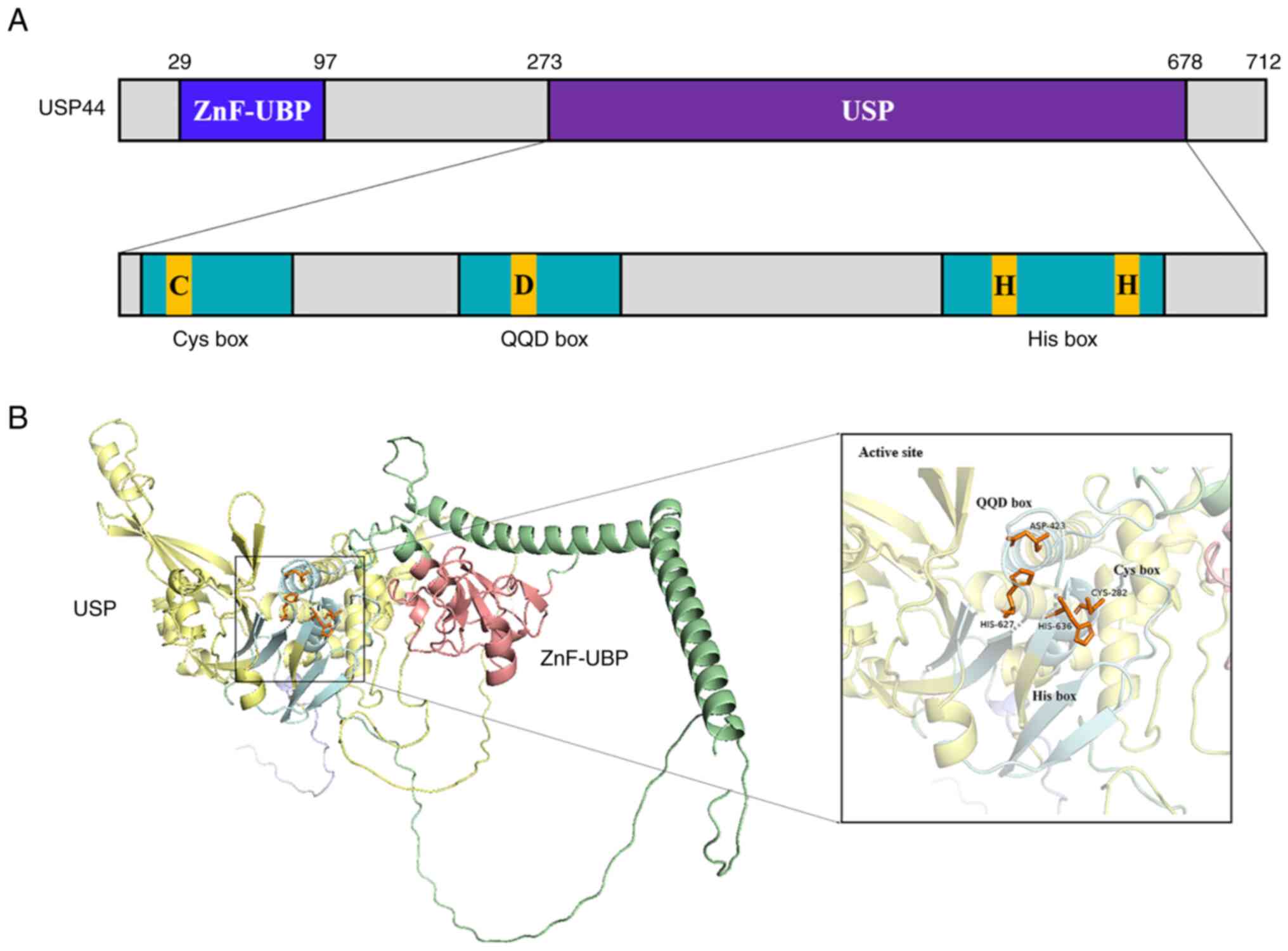
Properties of USP44
USP44 is a USP gene located on human
chromosome 12 and composed of five exons. Previous sequence
analysis of USP44 reported that its open reading frame (ORF)
was 2,139 bp and it encoded 712 amino acids, with a total molecular
weight of ~80 kDa. USP44 consists of ZnF-UBP and USP domains
(19,20). The ZnF-UBP domain is located at
amino acid residues 29–97, with the conserved catalytic USP domain
at amino acid residues 273–678. The catalytic domain of USP44 is
similar to that of other family members, consisting of a Cys box,
an Asp-containing motif and a His box (15,21)
(Fig. 1). These structures contain
a highly conserved cysteine residue, an aspartic acid residue and a
histidine residue, respectively (22–25).
Furthermore, USP44 also has a highly conserved centrin-binding
domain that is closely related to the centrosome distribution of
USP44 and its function in the prevention of chromosomal hysteresis
(26).
USP44 is mainly located in the nucleus, which may be
associated with the involvement of USP44 in chromosome-related
activities and its close association with chromosomes. For example,
USP44 is recruited to nuclear receptor corepressor (N-COR) target
loci to participate in gene expression regulation (27) and when DNA double-strand breaks
(DSBs) occur, USP44 is recruited to participate in the dynamic
regulation of damage repair (28).
Furthermore, a low level of USP44 expression in the cytoplasm has
also been reported previously (29), and recently, it has been
demonstrated that a part of USP44 in the cytoplasm can migrate to
transmembrane proteins to participate in the regulation of immune
response under the recruitment of viral infection signals (30). In these processes, the binding
domain of USP44 appears to be the key, unelucidated element. A
previous study reported that when the binding site between USP44
and the target protein was catalytically inactivated, the
recruitment of USP44 in the target protein region was significantly
reduced (26). This suggests that
the binding site of USP44 may largely determine the distribution of
USP44.
The activation of catalytic activity is the core
role of USP44. Similar to other domain-like DUBs (31,32),
USP44 needs to bind to partner proteins to achieve its full
enzymatic activity. It has been reported that for the identified
downstream factor histone H2B K120 (H2Bub1), the recombinant USP44
protein alone has no catalytic activity and only when USP44 is
bound to the N-COR complex can the enzyme activity of USP44 be
activated (27). Notably, N-COR
can also localize USP44, which suggests that the localization and
activation of USP44 may occur simultaneously (27). This also provides a new supplement
and explanation to the suggestion that the activation of the
catalytic activity of USP44 depends on its exclusive localization.
To be precise, the activation of USP44 activity depends on partner
proteins, which act as an anchor for localization as well as a
switch for catalytic activity. Moreover, it should be noted that
the catalytic activity of USP44 for other downstream factors, such
as centriole protein centrin 2 (CETN2) has not been reported to be
associated with the N-COR complex (26), which suggests that there may be
other complexes or mechanisms that mediate the activation of USP44
activity or that CETN2 itself serves three roles, namely, anchor
point, activation point and functional protein.
As part of post-translational modifications, the
ultimate biological function of deubiquitinating enzymes depends on
the cellular function of downstream proteins (33,34).
Therefore, targeting downstream proteins is the main pathway by
which USP44 participates in numerous cellular activities.
Furthermore, the cellular functions in which USP44 is involved are
often redundant and there are numerous deubiquitinating enzymes
with similar functions that can compensate, to a certain extent,
for the effects caused by USP44 defects (28,35).
These points suggest that it is not the USP44 defect, but the
change of downstream protein expression caused by the USP44 defect,
that is the main cause of specific pathological activity.
Roles of USP44 in cellular events
Anaphase initiation
The spindle assembly checkpoint serves a pivotal
role in the regulation of the precise separation of sister
chromosomes, as well as the initiation of anaphase. The spindle
assembly checkpoint monitors the connection of the spindle
microtubules to the centromere and the tension between sister
chromatids produced by the microtubules (36). When the centromere is not yet bound
to the microtubule or the tension threshold is not reached at the
checkpoint, the checkpoint is activated and mitotic arrest
deficient (MAD)1 interacts with closed (C)-MAD2 to form a stable
complex C-MAD2-MAD1. This complex is then used as a template to
recruit free open (O)-MAD2. By inducing the conformational
transition from O-MAD2 to C-MAD2, the C-MAD2 subunit bound to MAD1
catalyzes the binding of C-MAD2 to cell division cycle 20 (CDC20),
which then combines with the budding uninhibited by
benzimidazole-related 1 (BUBR1)-BUB3 dimer to form the mitotic
checkpoint complex (MCC) (37).
When microtubules are properly connected to centromeres,
anaphase-promoting complex/cyclosome (APC/C) ubiquitinates cyclin B
and securin by catalyzing the binding of polyubiquitin chains
consisting of Lys11, 48 and 63 to cyclin B and securin (38–40).
APC/C is a 1.5-MDa protein complex that is a large ubiquitin ligase
consisting of >10 subunits. Like all E3 enzymes, APC/C uses
ubiquitin residues that are activated by E1 and then transferred to
E2 enzymes [ubiquitin-conjugating enzyme E2 D1 (UBCH5) and UB
Ubiquitin-conjugating enzyme E2 C (UBCH10)] (41). An important pathway after
ubiquitination is 26S proteasome-mediated degradation (42). APC/C-mediated ubiquitination of
cyclin B and securin promotes their rapid destruction by the
proteasome, which initiates sister chromatid separation (43,44)
(Fig. 2).
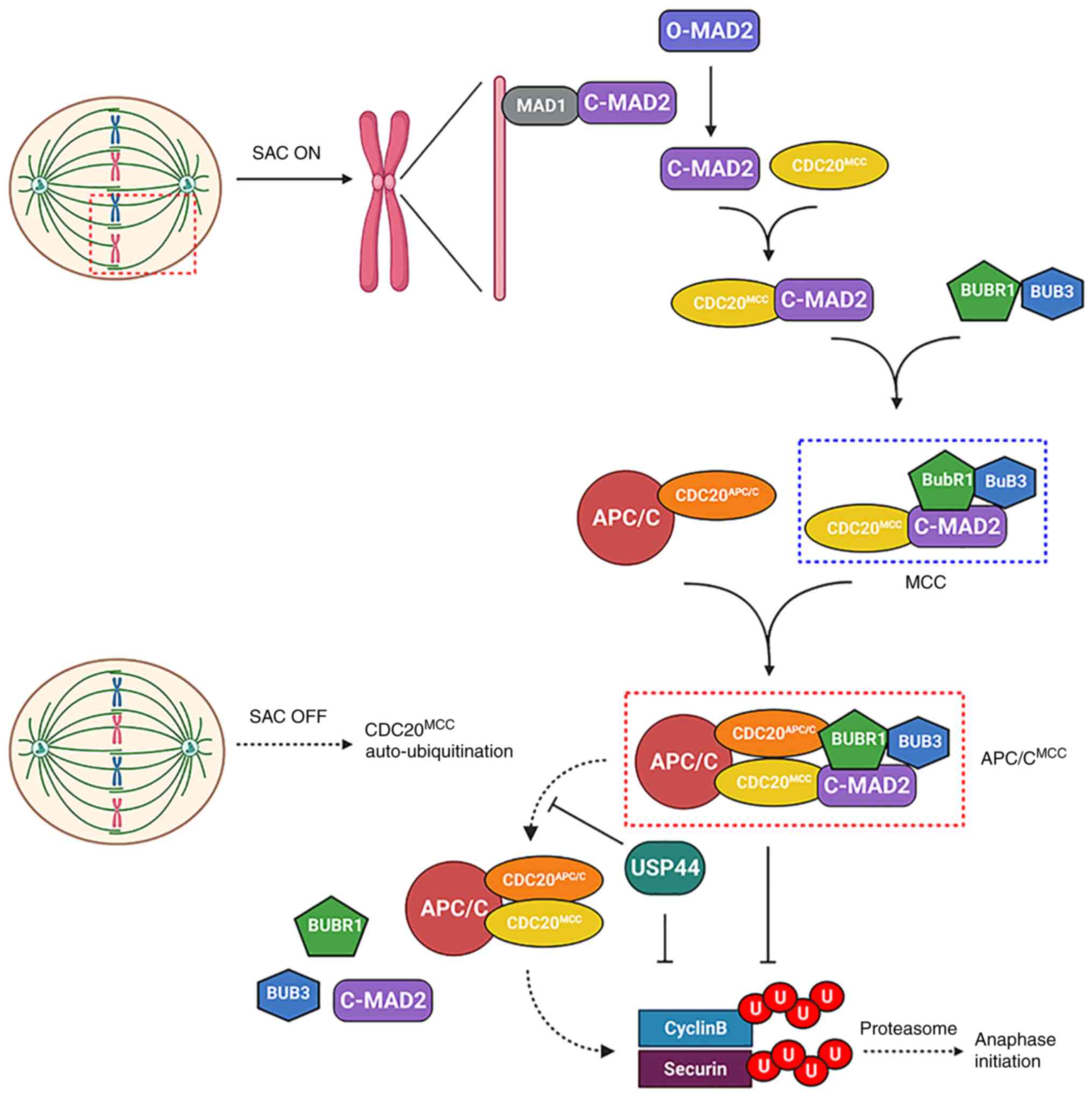 | Figure 2.Dynamic balance of APC/C
ubiquitination and USP44 deubiquitination controls anaphase
initiation. When the centromere is not yet bound to the microtubule
or the threshold tension is not reached at the checkpoint, the
checkpoint is activated. By inducing the conformational transition
from O-MAD2 to C-MAD2, the C-MAD2 subunit bound to MAD1 catalyzes
the binding of C-MAD2 to CDC20, which then combines with the
BUBR1-BUB3 dimer to form MCC. This complex inhibits the activity of
the APC/C through binding to the co-activating subunit CDC20
(CDC20APC/C). When microtubules are properly connected
to centromeres, the auto-ubiquitination of CDC20MCC
leads to the reactivation of APC/C activity, which leads to
anaphase initiation through the ubiquitination of cyclin B and
securin. In this process, USP44 can delay anaphase initiation not
only by direct stabilization of cyclin B and securin, but also by
stabilization of the binding of C-MAD2 and BUBR1 to CDC20 by
deubiquitination of CDC20MCC. MCC, mitotic checkpoint
complex; APC/C, anaphase-promoting complex/cyclosome; USP44,
ubiquitin-specific peptidase 44; O, open; C, closed; CDC20, cell
division cycle 20; MAD2, mitotic arrest deficient 2; BUBR1, budding
uninhibited by benzimidazole-related 1; BUB3, budding uninhibited
by benzimidazole 3; SAC, spindle assembly checkpoint. |
The regulation of APC/C is the core component of the
spindle assembly checkpoint mechanism. Previous studies by
Stegmeier et al (16) and
Reddy et al (45) reported
that USP44 may be a key regulatory factor in the physical
checkpoint of the spindle and may directly antagonize
UBCH10-induced APC/C-driven C-MAD2-CDC20 checkpoint complex
decomposition by promoting CDC20 deubiquitination. This pathway
leads to C-MAD2 disengagement and APC/C activation. By adjusting
this ubiquitination-deubiquitination switch, USP44 prevents
premature APC/C activation. Previous studies have reported that
APC/CMCC has two CDC20 sites, CDC20APC/C and
CDC20MCC (44,46). Alfieri et al (47) reported that UBCH10 mediated the
catalysis of intramolecular CDC20MCC ubiquitination via
cryogenic electron microscopy reconstruction of
APC/CMCC. Based on these findings, USP44 is likely to
stabilize the binding of C-MAD2 and BUBR1 to CDC20 via the
deubiquitination of CDC20MCC (Fig. 2). However, there is no clear
evidence that USP44 affects CDC20APC/C in this
process.
Centrosome separation
Increased frequency of lagged chromosomes has been
reported in USP44-deficient mouse models. Incomplete separation of
centrosomes and morphological changes of the spindle have been
demonstrated to be the main causes of this phenomenon (26). The ability of USP44 to bind to
CETN2 through highly conserved motifs and its deubiquitinating
activity are both reported to be key to ensuring accurate
chromosome separation (26).
DNA repair
Non-homologous end joining (NHEJ) is the main way to
repair DNA DSBs. The cellular response to DSBs is characterized by
a rapid accumulation of repair factors and signaling factors in the
vicinity of the lesion (48–50).
The recruitment of numerous factors in the chromatin region around
DSBs requires a ubiquitination cascade. However, this process is
opposed by USP44. USP44 can counteract the ubiquitination of
histone H2A mediated by ring finger protein (RNF)168 to inhibit the
recruitment of downstream repair factors (28). USP44 can also promote Ku80
degradation by stabilizing E3 ubiquitin ligase tripartite
motif-containing protein 25, which inhibits downstream factor
recruitment and ultimately inhibits NHEJ-mediated DNA repair
(51).
The nucleotide excision repair pathway of DNA repair
is responsible for correcting helix-distorting DNA lesions that are
caused by chemical damage or exposure to ultraviolet light
(52,53). In this process, USP44 stabilizes
the accumulation of DNA damage-binding protein 2 on DNA lesions by
deubiquitination, allowing sufficient time for the metastasis of
the lesions, which guarantees the smooth progress of DNA repair
(54).
Immune response
Lin et al (35) reported that, in mouse models, USP44
was not necessary for normal lymphoid development and that USP44
deficiency did not significantly affect the B-cell mediated immune
response. However, the role of USP44 in T lymphocytes has been
reported. FOXP3 is essential for the function of regulatory T cells
in immune homeostasis (55). USP44
has been reported to be a new FOXP3 deubiquitinase and has been
demonstrated to stabilize immune function in models of inflammatory
disease and cancer. Compared with wild-type regulatory T cells
(Tregs), Tregs lacking the USP44 gene had a weaker inhibitory
function (56).
Mediator of IRF3 activation (MITA), which is known
as a key adaptor protein, is responsible for sensing the second
messenger cyclic GMP-AMP, which is synthesized upon DNA virus
infection and activation of the induction of type I interferons
(IFNs) and proinflammatory cytokines (57). It has been reported that USP44 in
the cytoplasm is recruited to MITA to perform deubiquitination
after herpes simplex virus 1 (HSV-1) infection and that USP44
inhibits proteasome-mediated degradation of MITA by selectively
removing the polyubiquitin chain connected by K48 in MITA.
Moreover, gene transcription of IFNs and proinflammatory cytokines
in response to HSV-1 was reported to be inhibited in THP-1, bone
marrow-derived macrophage and murine lung fibroblast cells with
defective USP44 expression (30).
These results suggested that USP44 serves an important role in the
regulation of the natural immune response to DNA viruses.
Stem cell differentiation
Chromatin modification serves a key role in cell
differentiation. It has been reported that changes in histone H2B
ubiquitination patterns are essential for the maintenance of stem
cell differentiation potential to differentiation progression
(58,59). USP44 has recently been reported to
be the deubiquitination enzyme involved in this process. USP44, as
a regulator of H2Bub1 expression, is downregulated during embryonic
stem cell differentiation and, together with RNF20, regulates the
dynamics of H2B mono-ubiquitination patterns during stem cell
differentiation (17). USP44
affects the ability of embryonic stem cells to differentiate;
however, a study has demonstrated that USP44 is not a necessary
gene for growth and development, and mice lacking USP44 can still
grow and develop normally, which may be related to functional
redundancy (26).
Autophagy
Autophagy has been reported to be critical for
maintaining cell homeostasis, as it serves a role in clearing
abnormal proteins or factors that are no longer needed (60). Previously, H2B ubiquitination
regulation has been reported to be one of the mechanisms regulating
autophagy: Reduction of H2Bub1 can lead to activation of autophagy
(61). As aforementioned, USP44 is
responsible for the decrease in H2Bub1, and the autophagy process
is indeed inhibited when the interaction between USP44 and H2Bub1
is inhibited. These results suggest that USP44 can indeed affect
autophagy by the regulation of the expression of H2Bub1 (62).
USP44 in pathophysiological conditions
Aneuploidy
Deletion of USP44 has been reported to lead to
chromosome mis-segregation and aneuploidy. USP44 defect-mediated
aneuploidy is considered to have two mechanisms, namely, spindle
morphology change and mitotic timing change.
Physical changes in spindle morphology are
considered to be the main mechanism that affects aneuploidy
development. Incomplete centrosome separation and abnormal spindle
geometry caused by USP44 deletion are the main causes of mitotic
errors and aneuploidy (26).
It is known that the development of aneuploidy in
mitosis is closely related to the spindle assembly checkpoint
mechanism (63–65). The dynamic balance between APC/C
ubiquitination and USP44 deubiquitination regulates the initiation
of anaphase (16). The disruption
of this balance invalidates the spindle assembly checkpoint
mechanism. A weakened spindle assembly checkpoint allows cells with
unattached or misaligned kinetochores to proceed from metaphase to
anaphase, yielding daughter cells with an abnormal chromosome
number. However, in a previous study, in the absence of USP44,
there was no evidence reported for the accelerated degradation of
cyclin B1, even though there was a significant, moderate deficiency
in mitotic checkpoint activity (26). The USP44 defect initiates the
process that leads to aneuploidy through the spindle mechanism but
stops it partway. This may be associated with the fact that other
DUBs take over the basic functions associated with checkpoints
after the loss of USP44. Furthermore, USP44 upregulation is also
associated with aneuploidy (66).
Overexpression of USP44 can also lead to mitotic errors and
aneuploidy elevation (67) but
there is a lack of research to explain this phenomenon.
According to the aforementioned results, the
abnormal expression of USP44 is closely associated with the
development of aneuploidy but its complex regulatory mechanism
needs to be further evaluated.
Cancer
USP44 is a multifunctional factor in cancer
progression; it mediates tumorigenesis and tumor development
through different pathways in different tumors and is closely
related to the function of substrate proteins (68) (Fig.
3; Table I).
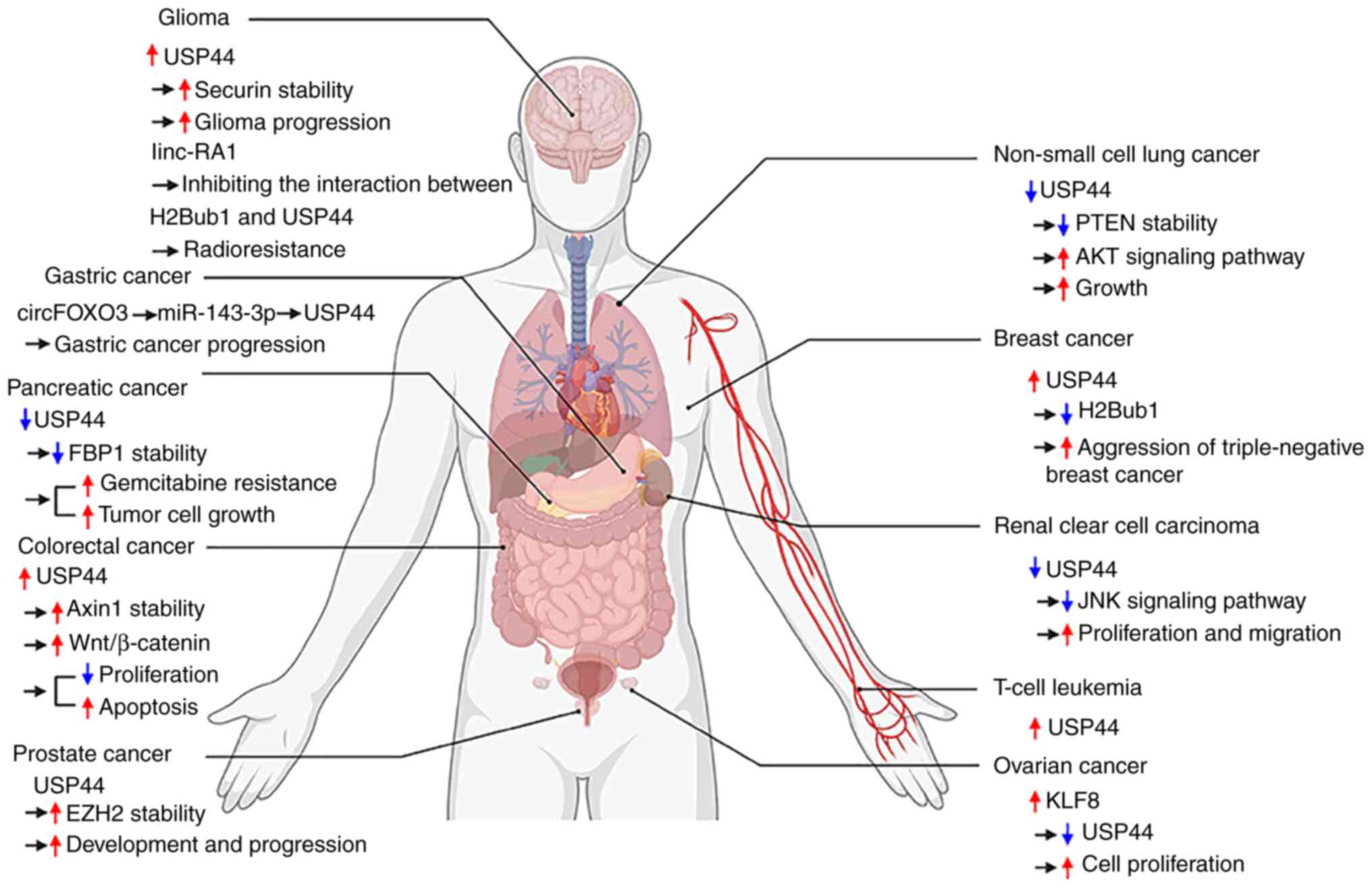 | Figure 3.USP44 is closely related to the
occurrence and progression of tumors. USP44 is closely related to
the occurrence and progression of tumors, including glioma, gastric
cancer, pancreatic cancer, colorectal cancer, prostate cancer,
non-small cell lung cancer, breast cancer, renal clear cell
carcinoma, T cell leukemia and ovarian cancer. Red arrows indicate
increased protein expression, pathway activation, increased
capacity or disease progression. The blue arrow indicates the
opposite to the red arrow. Black arrows indicate process
progression. USP44, ubiquitin-specific peptidase 44; EZH2, zeste
homolog 2; H2Bub1, histone H2B K120; FBP, fructose bisphosphatase
1. |
 | Table I.Summary of the downstream proteins
and functions targeted by USP44. |
Table I.
Summary of the downstream proteins
and functions targeted by USP44.
| Downstream
protein | Name | Function | (Refs.) |
|---|
| FBP1 |
Fructose-1,6-bisphosphatase | 1. Downregulation
of USP44 in pancreatic cancer mediates gemcitabine resistance
through FBP1. | (70) |
|
|
| 2. USP44 inhibits
tumor cell growth in pancreatic cancer through the FBP1-MAPK
signaling pathway. |
|
| N-COR | Nuclear receptor
co-repressor complex | 1. USP44
contributes to N-COR functions in regulating gene expression and in
modulating invasiveness of triple-negative breast cancer
cells. | (27) |
|
|
| 2. N-COR activates
deubiquitination of USP44. |
|
| TBL1X | Transducin β-like
1X | TBL1× is required
for USP44 to associate with N-COR. |
|
| TBL1XR1 | Transducin β-like
1X-related protein 1 | TBL1XR1 is required
for USP44 to associate with N-COR. |
|
| EZH2 | Enhancer of zeste
homolog 2 | USP44 promotes the
development and progression of prostate cancer by stabilizing
EZH2. | (90) |
| Securin | / | Overexpression of
USP44 promotes glioma progression by stabilizing securin. | (85) |
| Cyclin B1 | / | Antagonistic APC/C
function. | (16) |
| Axin 1 | / | USP44 suppresses
proliferation and enhances apoptosis in colorectal cancer cells by
inactivating Wnt/β-catenin signaling pathway via Axin 1
deubiquitination. | (74) |
| MITA | Mediator of IRF3
activation | USP44 is involved
in the regulation of innate immune responses to DNA viruses by
deubiquitinating MITA to prevent its degradation by
proteasomes. | (30) |
| CETN2 | Centrin-2 | Adjusting spindle
geometry, interpolar distance and centrosome separation. | (26) |
| FOXP3 | Forkhead box
protein P3 | USP44 collaborates
with USP7 to deubiquitinate and stabilize Foxp3 expression, thereby
promoting | (56) |
|
|
| Treg-mediated
immunosuppression. |
|
| PTEN | Phosphatase and
tensin homolog | Stabilize PTEN to
inhibit the AKT signaling pathway and thereby inhibit the growth of
non-small cell cancer cells. | (18) |
| Unknown | Unknown | USP44 inhibits the
JNK signaling pathway in renal carcinoma. | (75) |
| H2Bub1 | Histone H2B
K120 | 1. Regulates stem
cell differentiation. | (17) |
|
|
mono-ubiquitination | 2. Regulation of
H2BuB1-mediated autophagy is involved in the regulation of
radiation resistance of glioma. | (62) |
|
|
| 3. Opposite
phenotypes in different subtypes of breast cancer. | (84) |
|
|
| 4. Aggression of
triple-negative breast cancer. | (27) |
|
|
| 5. USP44-mediated
removal of H2Bub1 contributes to inhibition of N-COR target
genes. |
|
| CDC20 | Cell division cycle
20 homologue | Offsets
APC/C-driven decomposition of the MAD2-CDC20 complex and regulates
anaphase initiation. | (16) |
| DDB2 | DNA damage-binding
protein 2 | Participates in
nucleotide excision repair. | (54) |
Fructose bisphosphatase 1 (FBP1) is one of the key
enzymes in the gluconeogenesis process, which contributes to the
conversion of fructose-1,6-bisphosphatase to fructose-6-phosphate
and negatively regulates aerobic glycolysis (69). Studies reported that USP44 was
downregulated in pancreatic cancer, which was accompanied by the
downregulation of FBP1 and changes in glucose metabolism, mediating
the chemotherapy resistance to gemcitabine (70). Furthermore, FBP1 loss was
accompanied by the upregulation of ERK phosphorylation and changes
in cell proliferation (71).
Further experiments demonstrated that the FBP1-MAPK pathway was
regulated by USP44 and served an important role in the regulation
of the growth of the pancreatic cancer (70). These results indicate that USP44
may be a potential therapeutic target for pancreatic cancer.
In human tumor cells, CpG island (CGI) methylation
of promoter region 5 is involved in the regulation of the
expression of numerous genes (72), and USP44 is no exception. Recently,
the use of the combined bisulfite restriction analysis assay
demonstrated that transcriptional silencing of USP44 in CRC cell
lines was associated with CGI hypermethylation (73). This result was supported by the
re-expression of USP44 in four fully methylated CRC cell lines
(RKO, SW620, HCT116 and DDD-1) using the DNA methyltransferase
inhibitor decitabine (73). The
decreased expression of USP44 in colorectal cancer was also
reported in the study by Huang et al (74), where USP44, as a tumor suppressor,
was demonstrated to inhibit the Wnt/β-catenin pathway and promote
the apoptosis of colorectal cancer cells through the
deubiquitination of axin 1.
Renal clear cell carcinoma is the most common type
of renal carcinoma. Zhou et al (75) reported that USP44 was downregulated
in renal clear cell carcinoma and that its expression level was
negatively correlated with the grade and stage of renal cancer. It
was also demonstrated that downregulation of USP44 promoted cell
proliferation and migration through the JNK pathway in renal clear
cancer cells. Furthermore, Tang et al (76) constructed a more accurate
prognostic model using USP44 methylation as one of the
prognostic variables for renal clear cell carcinoma. These findings
suggest an important role for USP44 methylation in renal
clear cell carcinoma.
Among patients with lung cancer, non-small cell lung
cancer (NSCLC) is the most common type, accounting for ~80% of all
cases, and ~75% of patients are reported to have a poor 5-year
survival rate (77,78). A previous study has reported that
USP44 expression deficiency can lead to a significant increase in
the incidence of lung cancer (26). Subsequently, Zhang et al
(18) reported that the prognosis
of patients with lung cancer and low USP44 expression is poor and
the overexpression of USP44 may inhibit the progression of lung
cancer by stabilizing PTEN protein by the inhibition of AKT signal
transduction in lung cancer cells. These results indicate that
USP44 may be a potential therapeutic target for NSCLC.
For breast cancer, cancer stem cell (CSC) subclones
are often used as models. Liu et al (79) generated ‘mammospheres’ from breast
cancer cells to evaluate the role of USP44 in CSCs. Using
vasculogenic mimicry (VM), a newly defined tumor blood supply
pattern that has been reported to be closely associated with tumor
aggressiveness (80–82), as a bridge, the relationship
between USP44 and tumor aggressiveness was assessed. According to
the results of the study, USP44 inhibited breast CSCs with a
centrosomal amplification phenotype to form multipolar spindles,
but promoted the formation of a bipolar spindle, which was closely
associated with VM. After USP44 knockdown, multipole spindle
formation was induced, VM was inhibited and the ability of
mammosphere-derived MCF-7 AURKA cells to cross endothelial cells
was markedly reduced, which suggested a close relationship between
the four (79). Moreover, breast
cancer with USP44+ CSC subclones were significantly
associated with poor overall survival (OS) and disease-free
survival (DFS) times. The mean OS and DFS periods were 70.298
months (95% CI, 61.510-79.086) and 53.206 months (95% CI,
45.624-60.788), respectively, for patients with USP44+
CSC subclones; however, for patients without USP44+ CSC
subclones, the mean OS and DFS periods were 117.552 months (95% CI.
109.561-125.544) and 95.087 months (95% CI, 86.446-103.728),
respectively (79). These results
suggested that USP44 appeared to promote the progression of breast
cancer as an oncogenic factor. Furthermore, the downregulation of
USP44 in triple-negative breast cancer cells can impair the
aggressiveness of breast cancer cells, which also supports the
carcinogenic effect of USP44 (27). However, Chen et al (83) reported contrary results and
suggested that USP44 acts as a tumor suppressor in breast cancer to
limit tumor progression. The study demonstrated that patients with
breast cancer with high expression levels of USP44 had a better
prognosis and that the overexpression of USP44 in breast cancer
cells inhibited the malignancy of breast cancer, but the reasons
for this remain elusive. However, Tarcic et al (84) reported new insight into the
phenomenon of the aforementioned differential functional
expression, with different subtypes of breast cancer cells, which
demonstrated the opposite effects on proliferation after knocking
down USP44 to increase H2Bub1 levels. These results suggest that
USP44 serves different regulatory roles in different breast cancer
subtypes. Therefore, we hypothesized that these different results
may be caused by phenotypic differences between breast cancer stem
cells and breast cancer cells. Further research is needed to
evaluate this.
The role of USP44 in glioma, prostate cancer and
gastric cancer is different from the aforementioned roles. USP44 is
highly expressed in high-grade gliomas with a poor prognosis.
Downregulation of USP44 expression can inhibit proliferation,
migration and invasion of established glioma cell lines and induce
apoptosis (85).
In prostate cancer, USP44 promotes disease
progression by stabilizing enhancer of zeste homolog 2 (EZH2). EZH2
is an enzymatic catalytic subunit of polycomb repressive complex 2
(one of the two polycomb group protein core complexes) that can
alter gene expression by histone H3 lysine 27 trimethylation
(86). EZH2 has been reported to
promote the progression of prostate cancer by influencing several
factors associated with the cell cycle, autophagy and apoptosis
(87–89). The introduction of EZH2 into USP44
knockdown PC3 and DU145 cells significantly rescued USP44
knockdown-induced suppression of wound healing, migration and
invasion activity (90).
In gastric cancer, USP44 expression levels were also
upregulated. The mean and median expression rates of USP44 in
carcinoma were 39.6 and 36.0%, respectively, which were higher than
the 14.6 and 11.3%, respectively, in normal mucosa (66). Further studies demonstrated that
the upregulation of USP44 in GC was due to the interaction between
circFoxO3 and miR-143-3p, which promoted GC proliferation and
migration (91). Furthermore,
USP44 also has a clinical impact on the induction of DNA aneuploidy
and the poor prognosis of gastric cancer. The proportion of DNA
aneuploidy in gastric cancer with high USP44 expression levels was
significantly higher than that in gastric cancer with low USP44
expression levels. The 5-year OS and progression-free survival
rates of gastric cancer with high USP44 expression levels were 36.8
and 32.7%, respectively, which were significantly lower than those
of the low USP44 expression levels group (50.5 and 47.4%,
respectively) (66). These results
suggest that USP44 can not only inhibit tumors as a protective
factor, but can also promote tumor progression as an oncogenic
factor.
Regulation of USP44 expression
As presented in Fig.
4, USP44 is expressed to different degrees in numerous tissues
and organs, among which the lungs and kidneys show the highest
expression levels, whereas the bile duct and liver show lower
expression levels. This indicates that USP44 expression not only
has certain universality but that it also has certain tissue
specificity. However, it has not been clarified whether USP44 is
universally expressed in all tissues throughout the life cycle,
which demonstrates the need for the elucidation of the
macro-regulatory mechanism of USP44. Furthermore, the expression of
USP44 also demonstrates a complex trend in the tumor environment.
Compared with those in the corresponding normal tissues, the
expression levels of USP44 are significantly downregulated in most
tumors, such as lung cancer, kidney cancer and pancreatic cancer,
but upregulated in a few tumors, such as cholangiocarcinoma. These
results suggest that there may be complex tissue-dependent
regulation of USP44 in physiological and pathological conditions.
The regulatory mechanisms reported so far are elaborated on
next.
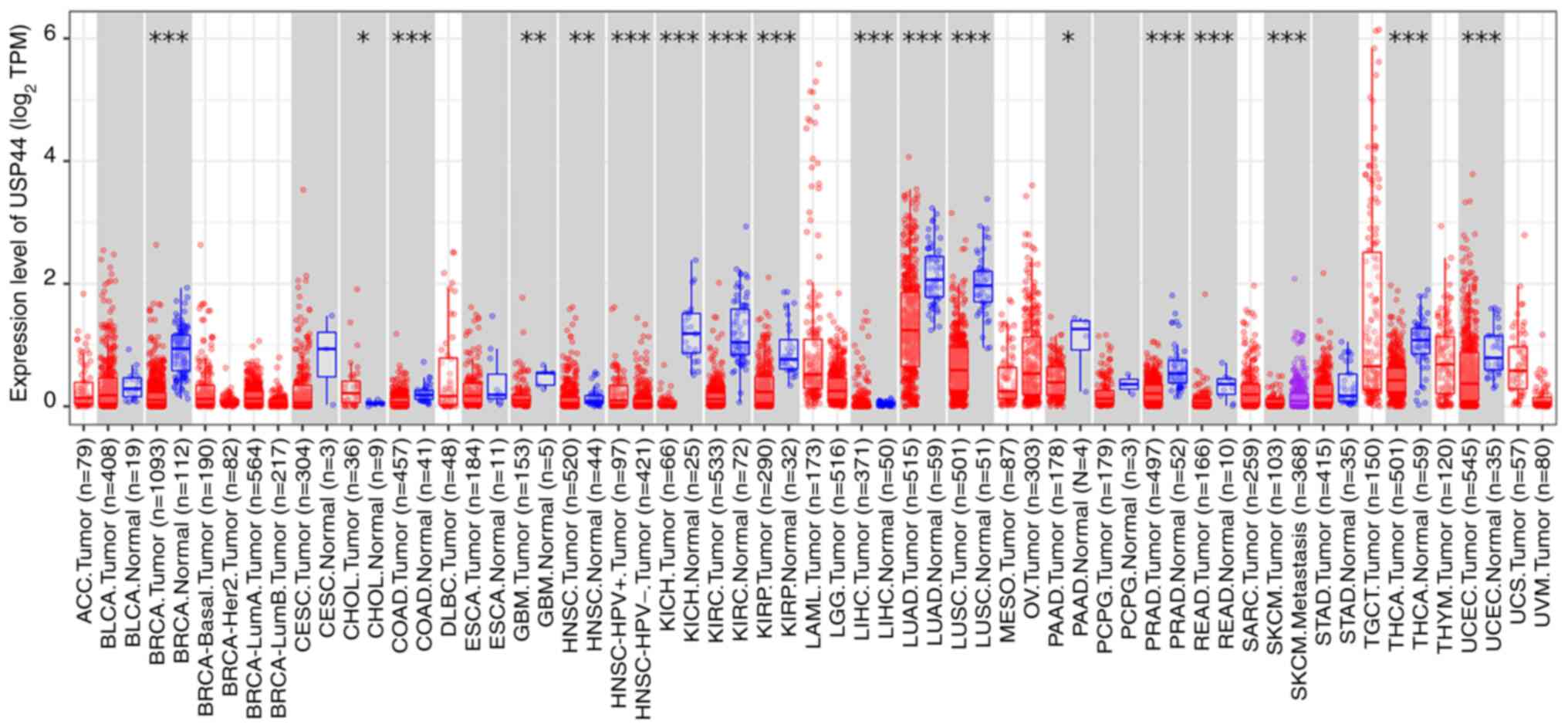 | Figure 4.Expression of USP44 in normal tissues
and corresponding tumors. *P<0.05, **P<0.01 and ***P<0.001
vs. normal. Gene expression data were provided by the open database
TIMER2.0 (http://timer.cistrome.org/). USP44,
ubiquitin-specific peptidase 44; TPM, transcripts per million; ACC,
adrenocortical carcinoma; BLCA, bladder urothelial carcinoma; BRCA,
breast invasive carcinoma; Her2, human epidermal growth factor
receptor 2; LumA, Luminal A; LumB, Luminal B; CESC, cervical and
endocervical cancer; CHOL, cholangiocarcinoma; COAD, colon
adenocarcinoma; DLBC, diffuse large B-cell lymphoma; ESCA,
esophageal carcinoma; GBM, glioblastoma multiforme; HNSC, head and
neck squamous cell carcinoma; HPV, human papillomavirus; KICH,
kidney chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP,
kidney renal papillary cell carcinoma; LAML, acute myeloid
leukemia; LGG, lower grade glioma; LIHC, liver hepatocellular
carcinoma; LUAD, liver hepatocellular carcinoma; LUSC, lung
squamous cell carcinoma; MESO, mesothelioma; OV, ovarian serous;
PAAD, pancreatic adenocarcinoma; PCPG, pheochromocytoma and
paraganglioma; PRAD, prostate adenocarcinoma; READ, rectum
adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous melanoma; STAD,
stomach adenocarcinoma; TGCT, testicular germ cell tumors; THCA,
thyroid carcinoma; THYM, thymoma; UCEC, uterine corpus endometrial
carcinoma; UCS, uterine carcinosarcoma; UVM, uterine
carcinosarcoma. |
USP44 is regulated by differentiation signals. SOX2,
Nanog and OCT4 are well known to be pluripotent transcription
factors that maintain the state of embryonic stem cells (92,93).
USP44 was reported as a direct target of OCT4 by Boyer et al
(94). The expression of USP44 was
downregulated during the differentiation of OCT4 knockdown-induced
embryonic cancer cells, which confirmed the regulatory role of OCT4
on USP44 expression at the functional level (95). Moreover, the high expression levels
of USP44 in embryonic stem cells, pluripotent stem cells and
germinal organs/cells, but low expression levels in differentiation
and somatic tissues, is consistent with the aforementioned results
(96). Furthermore, upstream
signaling that mediates the upregulation of USP44 expression during
Treg differentiation has also been reported. TGF-β/SMAD signaling
promotes the upregulation of USP44 expression by driving conserved
SMAD binding sites on the USP44 promoter (56).
The expression of USP44 is regulated via epigenetic
mechanisms. A previous study reported that USP44 not only serves a
key role in regulating proteasomal-mediated protein degradation but
also self-regulates through K48- and K63-linked polyubiquitination
degradation pathways (29).
Moreover, cell cycle-dependent changes in USP44 are also associated
with ubiquitination. USP44 is mainly localized to the nucleus. With
the end of the previous cell division, USP44 expression is rapidly
upregulated and reaches a peak in G1/S phase. After
that, USP44 expression begins to be downregulated, and when cells
enter mitosis, with nuclear envelope rupture, USP44 is rapidly
released into the cytoplasm and USP44 expression is further
decreased. Until anaphase, with nuclear envelope recombination,
USP44 recovers its tight association with chromosomes and its
expression is resumed shortly after mitotic exit (67). During this process, the proteasome
inhibitor MG132 stabilized USP44 levels both before and during
mitosis, which suggested that USP44 may be ubiquitinated and
degraded by the proteasome before and during mitosis (67). However, the E3 ligase mediating
ubiquitination of USP44 has not been reported.
APC/C and Skp1-cullin1-F-box (SCF) complex are
important E3 enzymes in monomial processes of the cell cycle
(97). The possibility of their
functioning in the same manner as E3S of USP44 was considered.
APC/C is known to be active from mitosis to the subsequent
G1 phase. At the G1/S boundary, APC/C is
forcibly inactivated by numerous mechanisms and remains low until
mitosis (98). However, this could
not explain the reported trend of USP44 expression peaking and then
decreasing in the G1/S phase (67), so the possibility of SCF
functioning in the same manner as E3S of USP44 was considered. It
is known that substrates recognized by SCF are mostly
phosphorylated (99), and studies
have reported that phosphorylated USP44 does exist in the cell
cycle (16,100). Furthermore, USP44 has been
reported to interact directly with the WD40 repeat sequence [a
specific sequence of the substrate recognition domain of the FBXL
family (a sub-family of F-box proteins)] (27). This suggests that USP44 has a
structural basis for binding to F-box proteins. Therefore, it is
feasible for phosphorylated USP44 to be ubiquitinated by the SCF
complex during the cell cycle. Moreover, dephosphorylation of USP44
was reported to be one of the changes that occur at the exit from
cell division, which is consistent with the rapid increase in USP44
expression after the exit from cell division (100). In summary, it is hypothesized
that the SCF complex, rather than APC/C, mediated the
ubiquitination of USP44 during the cell cycle, but further research
is needed.
Apart from ubiquitination and the aforementioned
phosphorylation, promoter methylation is the most extensively
studied epigenetic regulatory mechanism in the regulation of USP44
expression. Tropel et al (96) reported that CpG 9 methylation might
be involved in USP44 transcriptional regulation and promoter
selection, thereby mediating the tissue-specific expression of
USP44. Promoter hypermethylation is an important mechanism for the
epigenetic silencing of tumor suppressor genes (101,102). A close association between
hypermethylation of the USP44 promoter and downregulation of USP44
expression has been reported in colon and breast cancer (73,83).
Moreover, Chen et al (61)
also reported that the DNA methyltransferases DNMT3a and DNMT3b
acting on the promoter of USP44 were the transcriptional silencers
of the USP44 gene. Finally, the circRNA-miRNA axis is also one of
the mechanisms of USP44 expression. CircFOXO3 upregulates USP44
expression by regulating miR-143-3p, which directly targets the
USP44 gene (91). However, there
is no subsequent study on the mechanism of circRNA regulating USP44
expression, and further research is needed to more comprehensively
explore the regulatory mechanisms.
Therapeutic potential of targeting
USP44
USP44 serves a central role in multiple tumor
regulatory networks. Moreover, multiple studies have clearly
demonstrated that targeting USP44 is the key to inhibiting the
progression of nasopharyngeal, colon and lung cancer (18,51,74),
and improve the sensitivity of pancreatic cancer and glioma
treatment (62,70). Therefore, according to its
expression characteristics, reversing USP44 expression is a
promising therapeutic strategy. The exact regulation of USP44
expression by a complex named KRIBB53
(2′,4-dihydroxy-3,4′,6′-trimethoxychalcone) has been demonstrated.
KRIBB53 inhibited the expression of the downstream protein USP44
(IC50, 15 µm) and the progression of teratoma by
inducing the proteasome-dependent degradation of OCT4, which
confirmed the feasibility of targeting USP44 in the treatment of
tumors (103). Furthermore, it
can be hypothesized that antagonizing promoter methylation of USP44
(e.g., antagonizing the key enzymes DNMT3a and DNMT3b), limiting
ubiquitination of USP44 itself and searching for partner proteins
may be the most likely directions for future drug development.
Conclusions
In the present study, the basic characteristics,
functions and regulatory mechanisms of USP44 were systematically
introduced, emphasizing not only its physiological functions in
various cellular activities and pathophysiological roles in related
diseases, especially tumors (Table
II), but also the significant effect of USP44 expression
inhibitors on tumor treatment, which fully reflect the feasibility
and importance of USP44 as a tumor therapeutic target. However, at
present, no substantial progress has been made in the development
of drugs that directly target USP44, and numerous problems remain
to be resolved. For example, the exact mechanisms by which USP44
achieves specificity for each substrate need to be elucidated, and
all possible conformational changes in USP44 and their roles in
USP44 activation and substrate specificity need to be further
evaluated. These are critical for the development of selective
agonists and inhibitors. Therefore, in-depth evaluation of the
structure, regulation and dynamics of USP44 is expected to be the
key to solve this complex situation.
| Table II.Comparison of physiological and
pathological functions of USP44. |
Table II.
Comparison of physiological and
pathological functions of USP44.
| Physiological
function | Causes | Pathological
effect |
|---|
| Accurate separation
of centrosomes | USP44
downregulation/C281A/W162A | Aneuploidy |
| Unknown | USP44
upregulation |
|
| Participation in
spindle assembly checkpoint mechanism | USP44
downregulation | Accelerated
anaphase |
| Stabilization of
immune function | USP44
downregulation/C282S | A weaker
immunosuppressive function of Tregs |
|
| USP44
downregulation/C282A | Impair innate
immunity to DNA viruses |
| Involved in
stem-cell differentiation | USP44
downregulation/C282A | Impair
differentiation and induction of genes |
| Regulation of
autophagy | Inhibited
interaction between USP44 and H2Bub1 | Radio-resistance of
glioma |
| Involved in DNA
repair | USP44
downregulation/C281A | Induce tumors |
| Regulation of
tumor-associated proteins | USP44
downregulation | The tumorigenesis
and development of non-small cell carcinoma, colorectal cancer,
renal clear cell carcinoma and breast cancer |
|
| USP44
downregulation/C282A | The progression and
drug resistance of pancreatic cancer |
|
| USP44
upregulation | The development of
glioma and breast cancer |
|
| USP44
upregulation/C282A | The tumorigenesis
of prostate cancer |
Acknowledgements
Not applicable.
Funding
The present study was supported by The National Natural Science
Foundation of China (grant no. 81341135), Jinhua Nonprofit
Technology Applied Research Projects of Zhejiang China (grant no.
2018-4-016), Jinhua Central Hospital Nonprofit Technology Applied
Research Projects of Zhejiang China (grant no, JY2020-5-04) and The
Youth Key Project of Shaoxing People's Hospital of Zhejiang China
(grant no. 2021YA07).
Availability of data and materials
Not applicable.
Authors' contributions
YL was involved in the conception of the study and
drafted the manuscript. CX and FT reviewed and edited the
manuscript. MY performed a literature search and study selection.
All authors read and approved the final manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rock KL, Gramm C, Rothstein L, Clark K,
Stein R, Dick L, Hwang D and Goldberg AL: Inhibitors of the
proteasome block the degradation of most cell proteins and the
generation of peptides presented on MHC class I molecules. Cell.
78:761–771. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kwon YT and Ciechanover A: The ubiquitin
code in the ubiquitin-proteasome system and autophagy. Trends
Biochem Sci. 42:873–886. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dikic I: Proteasomal and autophagic
degradation systems. Annu Rev Biochem. 86:193–224. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ronai ZA: Monoubiquitination in
proteasomal degradation. Proc Natl Acad Sci USA. 113:8894–8896.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martinez-Fonts K, Davis C, Tomita T,
Elsasser S, Nager AR, Shi Y, Finley D and Matouschek A: The
proteasome 19S cap and its ubiquitin receptors provide a versatile
recognition platform for substrates. Nat Commun. 11:4772020.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Varshavsky A: The ubiquitin system, an
immense realm. Annu Rev Biochem. 81:167–176. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Husnjak K and Dikic I: Ubiquitin-binding
proteins: Decoders of ubiquitin-mediated cellular functions. Annu
Rev Biochem. 81:291–322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yau R and Rape M: The increasing
complexity of the ubiquitin code. Nat Cell Biol. 18:579–586. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Swatek KN and Komander D: Ubiquitin
modifications. Cell Res. 26:399–422. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mevissen TET and Komander D: Mechanisms of
deubiquitinase specificity and regulation. Annu Rev Biochem.
86:159–192. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Komander D and Rape M: The ubiquitin code.
Annu Rev Biochem. 81:203–229. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim W, Bennett EJ, Huttlin EL, Guo A, Li
J, Possemato A, Sowa ME, Rad R, Rush J, Comb MJ, et al: Systematic
and quantitative assessment of the ubiquitin-modified proteome. Mol
Cell. 44:325–340. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morreale FE and Walden H: Types of
ubiquitin ligases. Cell. 165:248–248.e1. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Heride C, Urbé S and Clague MJ: Ubiquitin
code assembly and disassembly. Curr Biol. 24:R215–R220. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Quesada V, Díaz-Perales A,
Gutiérrez-Fernández A, Garabaya C, Cal S and López-Otín C: Cloning
and enzymatic analysis of 22 novel human ubiquitin-specific
proteases. Biochem Biophys Res Commun. 314:54–62. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stegmeier F, Rape M, Draviam VM, Nalepa G,
Sowa ME, Ang XL, McDonald ER III, Li MZ, Hannon GJ, Sorger PK, et
al: Anaphase initiation is regulated by antagonistic ubiquitination
and deubiquitination activities. Nature. 446:876–881. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fuchs G, Shema E, Vesterman R, Kotler E,
Wolchinsky Z, Wilder S, Golomb L, Pribluda A, Zhang F, Haj-Yahya M,
et al: RNF20 and USP44 regulate stem cell differentiation by
modulating H2B monoubiquitylation. Mol Cell. 46:662–673. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang YK, Tian WZ, Zhang RS, Zhang YJ and
Ma HT: Ubiquitin-specific protease 44 inhibits cell growth by
suppressing AKT signaling in non-small cell lung cancer. Kaohsiung
J Med Sci. 35:535–541. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Komander D, Clague MJ and Urbe S: Breaking
the chains: Structure and function of the deubiquitinases. Nat Rev
Mol Cell Biol. 10:550–563. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Belle JI and Nijnik A: H2A-DUBbing the
mammalian epigenome: Expanding frontiers for histone H2A
deubiquitinating enzymes in cell biology and physiology. Int J
Biochem Cell Biol. 50:161–174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu M, Li P, Li M, Li W, Yao T, Wu JW, Gu
W, Cohen RE and Shi Y: Crystal structure of a UBP-family
deubiquitinating enzyme in isolation and in complex with ubiquitin
aldehyde. Cell. 111:1041–1054. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wilkinson KD: Regulation of
ubiquitin-dependent processes by deubiquitinating enzymes. FASEB J.
11:1245–1256. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Amerik A, Swaminathan S, Krantz BA,
Wilkinson KD and Hochstrasser M: In vivo disassembly of free
polyubiquitin chains by yeast Ubp14 modulates rates of protein
degradation by the proteasome. EMBO J. 16:4826–4838. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
D'Andrea A and Pellman D: Deubiquitinating
enzymes: A new class of biological regulators. Crit Rev Biochem Mol
Biol. 33:337–352. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chung CH and Baek SH: Deubiquitinating
enzymes: Their diversity and emerging roles. Biochem Biophys Res
Commun. 266:633–640. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang Y, Foreman O, Wigle DA, Kosari F,
Vasmatzis G, Salisbury JL, van Deursen J and Galardy PJ: USP44
regulates centrosome positioning to prevent aneuploidy and suppress
tumorigenesis. J Clin Invest. 122:4362–4374. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lan X, Atanassov BS, Li W, Zhang Y,
Florens L, Mohan RD, Galardy PJ, Washburn MP, Workman JL and Dent
SYR: USP44 is an integral component of N-CoR that contributes to
gene repression by deubiquitinating histone H2B. Cell Rep.
17:2382–2393. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mosbech A, Lukas C, Bekker-Jensen S and
Mailand N: The deubiquitylating enzyme USP44 counteracts the DNA
double-strand break response mediated by the RNF8 and RNF168
ubiquitin ligases. J Biol Chem. 288:16579–16587. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suresh B, Ramakrishna S, Lee HJ, Choi JH,
Kim JY, Ahn WS and Baek KH: K48- and K63-linked polyubiquitination
of deubiquitinating enzyme USP44. Cell Biol Int. 34:799–808. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang HY, Liao BW, Xu ZS, Ran Y, Wang DP,
Yang Y, Luo WW and Wang YY: USP44 positively regulates innate
immune response to DNA viruses through deubiquitinating MITA. PLoS
Pathog. 16:e10081782020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lang G, Bonnet J, Umlauf D, Karmodiya K,
Koffler J, Stierle M, Devys D and Tora L: The tightly controlled
deubiquitination activity of the human SAGA complex differentially
modifies distinct gene regulatory elements. Mol Cell Biol.
31:3734–3744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Z, Jones A, Joo HY, Zhou D, Cao Y,
Chen S, Erdjument-Bromage H, Renfrow M, He H, Tempst P, et al:
USP49 deubiquitinates histone H2B and regulates cotranscriptional
pre-mRNA splicing. Genes Dev. 27:1581–1595. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Harrigan JA, Jacq X, Martin NM and Jackson
SP: Deubiquitylating enzymes and drug discovery: Emerging
opportunities. Nat Rev Drug Discov. 17:57–78. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng J, Guo J, North BJ, Wang B, Cui CP,
Li H, Tao K, Zhang L and Wei W: Functional analysis of
deubiquitylating enzymes in tumorigenesis and development. Biochim
Biophys Acta Rev Cancer. 1872:1883122019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin YH, Forster M, Liang Y, Yu M, Wang H,
Robert F, Langlais D, Pelletier J, Clare S and Nijnik A: USP44 is
dispensable for normal hematopoietic stem cell function, lymphocyte
development, and B-cell-mediated immune response in a mouse model.
Exp Hematol. 72:1–8. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu H: Regulation of APC-Cdc20 by the
spindle checkpoint. Curr Opin Cell Biol. 14:706–714. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sudakin V, Chan GK and Yen TJ: Checkpoint
inhibition of the APC/C in HeLa cells is mediated by a complex of
BUBR1, BUB3, CDC20, and MAD2. J Cell Biol. 154:925–936. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Garnett MJ, Mansfeld J, Godwin C,
Matsusaka T, Wu J, Russell P, Pines J and Venkitaraman AR: UBE2S
elongates ubiquitin chains on APC/C substrates to promote mitotic
exit. Nat Cell Biol. 11:1363–1369. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wickliffe KE, Lorenz S, Wemmer DE, Kuriyan
J and Rape M: The mechanism of linkage-specific ubiquitin chain
elongation by a single-subunit E2. Cell. 144:769–781. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Williamson A, Banerjee S, Zhu X, Philipp
I, Iavarone AT and Rape M: Regulation of ubiquitin chain initiation
to control the timing of substrate degradation. Mol Cell.
42:744–757. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chang L, Zhang Z, Yang J, McLaughlin SH
and Barford D: Atomic structure of the APC/C and its mechanism of
protein ubiquitination. Nature. 522:450–454. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ciechanover A: Proteolysis: From the
lysosome to ubiquitin and the proteasome. Nat Rev Mol. 6:79–87.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Meyer HJ and Rape M: Processive ubiquitin
chain formation by the anaphase-promoting complex. Semin Cell Dev
Biol. 22:544–550. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Primorac I and Musacchio A: Panta rhei:
The APC/C at steady state. J Cell Biol. 201:177–189. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Reddy SK, Rape M, Margansky WA and
Kirschner MW: Ubiquitination by the anaphase-promoting complex
drives spindle checkpoint inactivation. Nature. 446:921–925. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Izawa D and Pines J: The mitotic
checkpoint complex binds a second CDC20 to inhibit active APC/C.
Nature. 517:631–634. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Alfieri C, Chang L, Zhang Z, Yang J,
Maslen S, Skehel M and Barford D: Molecular basis of APC/C
regulation by the spindle assembly checkpoint. Nature. 536:431–436.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ciccia A and Elledge SJ: The DNA damage
response: Making it safe to play with knives. Mol Cell. 40:179–204.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bekker-Jensen S and Mailand N: Assembly
and function of DNA double-strand break repair foci in mammalian
cells. DNA Repair (Amst). 9:1219–1228. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lukas J, Lukas C and Bartek J: More than
just a focus: The chromatin response to DNA damage and its role in
genome integrity maintenance. Nat Cell Biol. 13:1161–1169. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen Y, Zhao Y, Yang X, Ren X, Huang S,
Gong S, Tan X, Li J, He S, Li Y, et al: USP44 regulates
irradiation-induced DNA double-strand break repair and suppresses
tumorigenesis in nasopharyngeal carcinoma. Nat Commun. 13:5012022.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kamileri I, Karakasilioti I and Garinis
GA: Nucleotide excision repair: New tricks with old bricks. Trends
Genet. 28:566–573. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Marteijn JA, Lans H, Vermeulen W and
Hoeijmakers JH: Understanding nucleotide excision repair and its
roles in cancer and ageing. Nat Rev Mol Cell Biol. 15:465–481.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang Y, Mandemaker IK, Matsumoto S,
Foreman O, Holland CP, Lloyd WR, Sugasawa K, Vermeulen W, Marteijn
JA and Galardy PJ: USP44 stabilizes DDB2 to facilitate nucleotide
excision repair and prevent tumors. Front Cell Dev Biol.
9:6634112021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sakaguchi S, Yamaguchi T, Nomura T and Ono
M: Regulatory T cells and immune tolerance. Cell. 133:775–787.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yang J, Wei P, Barbi J, Huang Q, Yang E,
Bai Y, Nie J, Gao Y, Tao J, Lu Y, et al: The deubiquitinase USP44
promotes Treg function during inflammation by preventing FOXP3
degradation. EMBO Rep. 21:e503082020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Luo WW and Shu HB: Delicate regulation of
the cGAS-MITA-mediated innate immune response. Cell Mol Immunol.
15:666–675. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen S, Li J, Wang DL and Sun FL: Histone
H2B lysine 120 monoubiquitination is required for embryonic stem
cell differentiation. Cell Res. 22:1402–1405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Karpiuk O, Najafova Z, Kramer F, Hennion
M, Galonska C, König A, Snaidero N, Vogel T, Shchebet A,
Begus-Nahrmann Y, et al: The histone H2B monoubiquitination
regulatory pathway is required for differentiation of multipotent
stem cells. Mol Cell. 46:705–713. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Klionsky DJ: Autophagy: From phenomenology
to molecular understanding in less than a decade. Nat Rev Mol Cell
Biol. 8:931–937. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen S, Jing Y, Kang X, Yang L, Wang DL,
Zhang W, Zhang L, Chen P, Chang JF, Yang XM and Sun FL: Histone H2B
monoubiquitination is a critical epigenetic switch for the
regulation of autophagy. Nucleic Acids Res. 45:1144–1158.
2017.PubMed/NCBI
|
|
62
|
Zheng J, Wang B, Zheng R, Zhang J, Huang
C, Zheng R, Huang Z, Qiu W, Liu M, Yang K, et al: Linc-RA1 inhibits
autophagy and promotes radioresistance by preventing H2Bub1/USP44
combination in glioma cells. Cell Death Dis. 11:7582020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Targa A and Rancati G: Cancer: A CINful
evolution. Curr Opin Cell Biol. 52:136–144. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Gordon DJ, Resio B and Pellman D: Causes
and consequences of aneuploidy in cancer. Nat Rev Genet.
13:189–203. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Simonetti G, Bruno S, Padella A, Tenti E
and Martinelli G: Aneuploidy: Cancer strength or vulnerability? Int
J Cancer. 144:8–25. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Nishimura S, Oki E, Ando K, Iimori M,
Nakaji Y, Nakashima Y, Saeki H, Oda Y and Maehara Y: High
ubiquitin-specific protease 44 expression induces DNA aneuploidy
and provides independent prognostic information in gastric cancer.
Cancer Med. 6:1453–1464. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang Y, van Deursen J and Galardy PJ:
Overexpression of ubiquitin specific protease 44 (USP44) induces
chromosomal instability and is frequently observed in human T-cell
leukemia. PLoS One. 6:e233892011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yuan T, Yan F, Ying M, Cao J, He Q, Zhu H
and Yang B: Inhibition of ubiquitin-specific proteases as a novel
anticancer therapeutic strategy. Front Pharmacol. 9:10802018.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Timson DJ: Fructose 1,6-bisphosphatase:
Getting the message across. Biosci Rep. 39:BSR201901242019.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yang C, Zhu S, Yang H, Deng S, Fan P, Li M
and Jin X: USP44 suppresses pancreatic cancer progression and
overcomes gemcitabine resistance by deubiquitinating FBP1. Am J
Cancer Res. 9:1722–1733. 2019.PubMed/NCBI
|
|
71
|
Jin X, Pan Y, Wang L, Ma T, Zhang L, Tang
AH, Billadeau DD, Wu H and Huang H: Fructose-1,6-bisphosphatase
inhibits ERK activation and bypasses gemcitabine resistance in
pancreatic cancer by blocking IQGAP1-MAPK interaction. Cancer Res.
77:4328–4341. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Teodoridis JM, Strathdee G and Brown R:
Epigenetic silencing mediated by CpG island methylation: Potential
as a therapeutic target and as a biomarker. Drug Resist Updat.
7:267–278. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Sloane MA, Wong JW, Perera D, Nunez AC,
Pimanda JE, Hawkins NJ, Sieber OM, Bourke MJ, Hesson LB and Ward
RL: Epigenetic inactivation of the candidate tumor suppressor USP44
is a frequent and early event in colorectal neoplasia. Epigenetics.
9:1092–1100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Huang T, Zhang Q, Ren W, Yan B, Yi L, Tang
T, Lin H and Zhang Y: USP44 suppresses proliferation and enhances
apoptosis in colorectal cancer cells by inactivating the
Wnt/β-catenin pathway via Axin1 deubiquitination. Cell Biol Int.
44:1651–1659. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhou J, Wang T, Qiu T, Chen Z, Ma X, Zhang
L and Zou J: Ubiquitin-specific protease-44 inhibits the
proliferation and migration of cells via inhibition of JNK pathway
in clear cell renal cell carcinoma. BMC Cancer. 20:2142020.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tang W, Cao Y and Ma X: Novel prognostic
prediction model constructed through machine learning on the basis
of methylation-driven genes in kidney renal clear cell carcinoma.
Biosci Rep. 40:BSR202016042020. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: Epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Xu D, Tian W, Jiang C, Huang Z and Zheng
S: The anthelmintic agent oxfendazole inhibits cell growth in
non-small cell lung cancer by suppressing c-Src activation. Mol Med
Rep. 19:2921–2926. 2019.PubMed/NCBI
|
|
79
|
Liu T, Sun B, Zhao X, Li Y, Zhao X, Liu Y,
Yao Z, Gu Q, Dong X, Shao B, et al: USP44+ cancer stem cell
subclones contribute to breast cancer aggressiveness by promoting
vasculogenic mimicry. Mol Cancer Ther. 14:2121–2131. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Sun B, Zhang S, Zhang D, Du J, Guo H, Zhao
X, Zhang W and Hao X: Vasculogenic mimicry is associated with high
tumor grade, invasion and metastasis, and short survival in
patients with hepatocellular carcinoma. Oncol Rep. 16:693–698.
2006.PubMed/NCBI
|
|
81
|
Sun T, Sun BC, Zhao XL, Zhao N, Dong XY,
Che N, Yao Z, Ma YM, Gu Q, Zong WK and Liu ZY: Promotion of tumor
cell metastasis and vasculogenic mimicry by way of transcription
coactivation by Bcl-2 and Twist1: A study of hepatocellular
carcinoma. Hepatology. 54:1690–1706. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Luo F, Yang K, Liu RL, Meng C, Dang RF and
Xu Y: Formation of vasculogenic mimicry in bone metastasis of
prostate cancer: Correlation with cell apoptosis and senescence
regulation pathways. Pathol Res Pract. 210:291–295. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Chen X, Wu X and Lei W: USP44
hypermethylation promotes cell proliferation and metastasis in
breast cancer. Future Oncol. 17:279–289. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Tarcic O, Granit RZ, Pateras IS, Masury H,
Maly B, Zwang Y, Yarden Y, Gorgoulis VG, Pikarsky E, Ben-Porath I
and Oren M: RNF20 and histone H2B ubiquitylation exert opposing
effects in Basal-Like versus luminal breast cancer. Cell Death
Differ. 24:694–704. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zou Y, Qiu G, Jiang L, Cai Z, Sun W, Hu H,
Lu C, Jin W and Hu G: Overexpression of ubiquitin specific
proteases 44 promotes the malignancy of glioma by stabilizing
tumor-promoter securin. Oncotarget. 8:58231–58246. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Duan R, Du W and Guo W: EZH2: A novel
target for cancer treatment. J Hematol Oncol. 13:1042020.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Nutt SL, Keenan C, Chopin M and Allan RS:
EZH2 function in immune cell development. Biol Chem. 401:933–943.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Yao Y, Hu H, Yang Y, Zhou G, Shang Z, Yang
X, Sun K, Zhan S, Yu Z, Li P, et al: Downregulation of enhancer of
zeste homolog 2 (EZH2) is essential for the induction of autophagy
and apoptosis in colorectal cancer cells. Genes (Basel). 7:832016.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ito T, Teo YV, Evans SA, Neretti N and
Sedivy JM: Regulation of cellular senescence by polycomb chromatin
modifiers through distinct DNA damage- and histone
methylation-dependent pathways. Cell Rep. 22:3480–3492. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Park JM, Lee JE, Park CM and Kim JH: USP44
promotes the tumorigenesis of prostate cancer cells through EZH2
protein stabilization. Mol Cells. 42:17–27. 2019.PubMed/NCBI
|
|
91
|
Xiang T, Jiang HS, Zhang BT and Liu G:
CircFOXO3 functions as a molecular sponge for miR-143-3p to promote
the progression of gastric carcinoma via upregulating USP44. Gene.
753:1447982020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wang Z, Oron E, Nelson B, Razis S and
Ivanova N: Distinct lineage specification roles for NANOG, OCT4,
and SOX2 in human embryonic stem cells. Cell Stem Cell. 10:440–454.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Yu J, Vodyanik MA, Smuga-Otto K,
Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA,
Ruotti V, Stewart R, et al: Induced pluripotent stem cell lines
derived from human somatic cells. Science. 318:1917–1920. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Boyer LA, Lee TI, Cole MF, Johnstone SE,
Levine SS, Zucker JP, Guenther MG, Kumar RM, Murray HL, Jenner RG,
et al: Core transcriptional regulatory circuitry in human embryonic
stem cells. Cell. 122:947–956. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Jung M, Peterson H, Chavez L, Kahlem P,
Lehrach H, Vilo J and Adjaye J: A data integration approach to
mapping OCT4 gene regulatory networks operative in embryonic stem
cells and embryonal carcinoma cells. PLoS One. 5:e107092010.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Tropel P, Jung L, André C, Ndandougou A
and Viville S: CpG island methylation correlates with the use of
alternative promoters for USP44 gene expression in human
pluripotent stem cells and testes. Stem Cells Dev. 26:1100–1110.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Mocciaro A and Rape M: Emerging regulatory
mechanisms in ubiquitin-dependent cell cycle control. J Cell Sci.
125:255–263. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Kernan J, Bonacci T and Emanuele MJ: Who
guards the guardian? Mechanisms that restrain APC/C during the cell
cycle. Biochim Biophys Acta Mol Cell Res. 1865:1924–1933. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Petroski MD and Deshaies RJ: Function and
regulation of cullin-RING ubiquitin ligases. Nat Rev Mol Cell Biol.
6:9–20. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Visconti R, Palazzo L, Della Monica R and
Grieco D: Fcp1-dependent dephosphorylation is required for
M-phase-promoting factor inactivation at mitosis exit. Nat Commun.
3:8942012. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Berdasco M and Esteller M: Aberrant
epigenetic landscape in cancer: How cellular identity goes awry.
Dev Cell. 19:698–711. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Esteller M: Epigenetic gene silencing in
cancer: The DNA hypermethylome. Hum Mol Genet. 16:R50–R59. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Jung J, Kim Y, Song J, Yoon YJ, Kim DE,
Kim JA, Jin Y, Lee YJ, Kim S, Kwon BM and Han DC: KRIBB53 binds to
OCT4 and enhances its degradation through the proteasome, causing
apoptotic cell death of OCT4-positive testicular germ cell tumors.
Carcinogenesis. 39:838–849. 2018. View Article : Google Scholar : PubMed/NCBI
|