Introduction
Non-small cell lung cancer (NSCLC) is the most
common type of cancer, accounting for ~85% of all cases of lung
cancer, and is a major cause of cancer-associated mortality
worldwide (1). In the USA in 2020,
there were estimated to be 228,820 new cases of lung cancer and
140,730 new deaths from lung cancer (2). The 5-year survival rate of patients
with metastatic NSCLC is estimated to be 6% (3). Despite recent advancements in medical
treatment, the 5-year recurrence rate of NSCLC remains high at
>20% (4), indicating that it is
important to elucidate the molecular mechanisms underlying the
progression of NSCLC.
Ferroptosis is an iron-dependent form of oxidative
cell death triggered by the accumulation of lethal levels of
lipid-based reactive oxygen species (ROS) and lipid peroxidation
(5,6). Studies have demonstrated that
ferroptosis is critical in tumourigenesis, cancer development and
therapy (7–10). For example, ferroptosis is induced
in pancreatic cancer cells by artesunate (11), in Huh-7 hepatocellular carcinoma
cells by sorafenib (12) and in
human gastric cancer cells by erastin (13). Moreover, sorafenib has been reported
to induce ferroptosis in NSCLC cells (14).
Hypoxia plays a critical role in tumourigenesis,
metastasis and chemotherapy resistance in solid tumours (15–17).
The lack of oxygen enhances the expression and stability of
hypoxia-inducible factors (HIFs) such as HIF-1α (18). HIF-1α increases the expression of
vascular endothelial growth factor (VEGF) and promotes the
proliferation and migration of cancer cells (19). Furthermore, HIF-1α is detectable in
50% of solid tumours with high proliferative characteristics
(17,20). Therefore, HIF-1α is a potential
target for cancer therapy. Additionally, recent studies have shown
that HIF-1α inhibits ferroptosis in hepatocellular carcinoma
(21) and gastric cancer (22). However, the association between
HIF-1α and ferroptosis during the development of NSCLC remains
unclear.
The Hippo-Yes-associated protein (YAP) pathway
influences a wide variety of tumour types, including NSCLC
(23). YAP1, a transcriptional
activator of Hippo signalling, binds to and maintains the stability
of HIF-1α protein in the nuclei of tumour cells (24). YAP1 also promotes the proliferation,
invasion, migration and chemoresistance of cancer cells (25,26).
Wang and Liu (27) found that
knockdown of the long non-coding RNA GHET1 inhibited the
hypoxia-induced upregulation of HIF-1α expression and nuclear
translocation of YAP, thereby reducing the proliferation and
invasion of triple-negative breast cancer cells. Furthermore, YAP
has been shown to promote ferroptosis in cancer cells including
renal cell carcinoma (28), liver
cancer (29) and colon cancer
(30). Therefore, we hypothesised
that HIF-1α blocks ferroptosis in NSCLC by activating the Hippo-YAP
pathway.
In the present study, the proliferation, invasion,
ferroptosis and oxidative stress levels of NSCLC cells were
investigated after the silencing HIF-1α with or without treatment
with the ferroptosis antagonist ferrostatin-1 (Fer-1) or
ferroptosis agonist erastin. Expression levels of the Hippo-YAP
pathway-associated protein YAP1 and ferroptosis marker protein
glutathione peroxidase 4 (GPX4) were also detected. HIF-1α and
ferroptosis were investigated as a therapeutic target and strategy,
respectively, for the treatment of NSCLC.
Materials and methods
Cell culture
SW900 (cat. no. HTB-59) and A549 (cat. no.
CRM-CCL-185) human NSCLC and BEAS-2B (cat. no. CRL-9609) human
bronchial epithelial cell lines acquired from the American Type
Culture Collection were used in the study. The SW900 and A549 cells
were maintained in RPMI-1640 (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 4.5 g/l glucose, 4 mmol/l L-glutamine, 10%
foetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin-streptomycin (Gibco; Thermo Fisher Scientific,
Inc.). The BEAS-2B cells were cultured in RPMI-1640 containing 10%
FBS and 1% penicillin-streptomycin. All cells were cultured in
triplicate in 12-well plates (1×105/cm2) at
37°C with 5% CO2.
Cell transfection
Small interfering RNA (siRNA) specific to HIF-1α
(si-HIF-1α, 5′-CGAUGGAAGCACUAGACAAAG-3′), siRNA negative control
(si-NC, 5′-CACUGAUUUCAAAUGGUGCUAUU-3′) and a sequence for the
overexpression (oe) of YAP1 (oe-YAP1; Shanghai GeneChem Co., Ltd.)
were cloned into lentiviral vectors (Lenti-Mix: pMDLg/pRRE, pVSV-G,
pRSV-Rev; Wuhan GeneCreate Biological Engineering Co., Ltd.).
Lentivirus packaging plasmids (50 ng/µl) expressing si-HIF-1α,
si-NC, oe-YAP1 and negative control for YAP1 overexpression (oe-NC)
were constructed and transfected into SW900 and A549 cells using
HighGene transfection reagent (ABclonal Biotech Co., Ltd.) at 37°C
for 72 h. The mRNA expression and protein levels of HIF-1α and YAP1
were confirmed using reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) and western blotting, respectively.
Ferroptosis induction or
inhibition
To induce ferroptosis, NSCLC cells were treated with
5 µg/ml erastin (Beijing Solarbio Science & Technology Co.,
Ltd.) for 24 h. Treatment with Fer-1 (10 µmol/l; ABclonal Biotech
Co., Ltd.) for 24 h was used to inhibit ferroptosis (7,31). The
treatments were performed at 37°C in the presence of 5%
CO2.
Cell Counting Kit-8 (CCK-8) assay
The proliferation of SW900 and A549 cells was
detected using a CCK-8 assay (Beyotime Institute of Biotechnology).
Cells (2×10 cells/well) were seeded into 96-well plates (100
µl/well) and cultured for 24, 48, 72 and 96 h. The cells were then
incubated with 10 µl CCK-8 solution for 2 h. A microplate reader
(Wuxi Hiwell-Diatek Instruments Co., Ltd.) was used to determine
the optical density at 450 nm (OD450).
Flow cytometry assay
NSCLC cell apoptosis was detected using an Annexin
V-FITC Apoptosis Detection Kit (Beyotime Institute of
Biotechnology). In brief, 2×105 cells were resuspended
in 500 µl binding buffer and incubated with 5 µl Annexin V-FITC for
30 min at 4°C in the dark, followed by incubation with 5 µl
propidium iodide for 5 min at 25°C. The apoptosis rate was then
detected using flow cytometry with a CytoFLEX S instrument (Beckman
Coulter, Inc.) and Cell Quest software (version 5.2.1; BD
Biosciences).
Transwell assay
Cell invasion was evaluated using Matrigel-coated
Transwell chambers (Corning, Inc.). NSCLC cells
(1×106/ml) were seeded into the upper chambers filled
with serum-free RPMI-1640. The lower chambers were filled with
complete RPMI-1640, and the cells cultured for 48 h at 37°C with 5%
CO2. Cells that adhered to the surface of the lower
chamber were fixed and stained with 1% crystal violet (Beyotime
Institute of Biotechnology) at room temperature for 20 min. Cells
were counted in digital photographs captured using a CKX53
microscope (Olympus Corporation).
RNA isolation and RT-qPCR
RNA was isolated from NSCLC and BEAS-2B cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). First-strand cDNA was synthesised using a Fastingking gDNA
Dispelling RT SuperMix kit (Tiangen Biotech Co., Ltd.) according to
the manufacturer's protocol. SYBR Green PCR Master Mix (Xiamen Life
Internet Technology Co., Ltd.) and an MX3000P Fast RT-PCR
instrument (Agilent Technologies, Inc.) were used for qPCR
analysis. The thermocycling conditions were as follows: 95°C for 3
min; followed by 40 cycles of 95°C for 12 sec and 62°C for 40 sec.
The expression levels of HIF-1α and YAP1 were determined using the
2−∆∆Cq method (32).
GAPDH was used as an internal reference gene. The specific primers
used for qPCR analysis were as follows: HIF-1α forward,
5′-AGAGGTTGAGGGACGGAGAT-3′ and reverse, 5′-GCACCAAGCAGGTCATAGGT-3′;
YAP1 forward, 5′-TGACCCTCGTTTTGCCATGA-3′ and reverse
5′-GTTGCTGCTGGTTGGAGTTG-3′; GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse,
5′-ATGGTGGTGAAGACGCCAGT-3′.
Western blot analysis
Proteins were isolated from NSCLC and BEAS-2B cells
using a radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology). The isolated protein samples were
quantified using a bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology) and then protein samples (25 µg) were
separated using 10% sodium dodecyl sulphate-polyacrylamide gel
electrophoresis (Beyotime Institute of Biotechnology).
Immunoblotting analysis was performed using polyvinylidene fluoride
membranes (Beyotime Institute of Biotechnology). The membranes were
blocked with 5% non-fat milk for 6 min at room temperature. For the
immunoblotting analysis, the membranes were incubated with specific
primary antibodies, namely anti-HIF-1α (1:500; cat. no. ab51608;
Abcam), anti-GPX4 (1:5,000; cat. no. ab125066; Abcam), anti-YAP1
(1:5,000; cat. no. ab52771; Abcam), anti-phosphorylated (p)-YAP1
(1:25,000; cat. no. ab76252; Abcam) and anti-GAPDH (1:2,500; cat.
no. ab9485; Abcam) antibodies, at 4°C overnight. Horseradish
peroxidase-tagged goat anti-rabbit IgG H&L (1:10,000; cat. no.
A0516; Beyotime Institute of Biotechnology) was used as the
secondary antibody and incubated with the membranes at room
temperature for 1 h. An enhanced chemiluminescence system (Pierce;
Thermo Fisher Scientific, Inc.) and automatic digital gel image
analysis system (Tanon 3500; Tanon Science & Technology Co.,
Ltd.) were used to analyse the target proteins.
Enzyme-linked immunosorbent assay
The levels of malondialdehyde (MDA), glutathione
(GSH) and ROS in A549 cells were determined using corresponding
commercial enzyme-linked immunosorbent assay kits (cat. no. BC0025
for MDA, Beijing Solarbio Science & Technology Co., Ltd.; cat.
no. E-BC-K030-M for GSH, Elabscience Biotechnology, Inc.; and cat.
no. CA1410 for ROS, Beijing Solarbio Science & Technology Co.,
Ltd.). A microplate reader was used to measure the OD450
value.
Detection of ferrous iron
(Fe2+) levels
The concentration of Fe2+ in A549 cells
was determined using an iron colorimetric assay kit (cat. no.
K390-100; BioVision, Inc.). A microplate reader was used to
determine the OD450 value.
Statistical analysis
All experiments were performed in triplicate.
Statistical analyses were conducted using GraphPad Prism 7.0
software (GraphPad Software, Inc.). Experimental data are expressed
as the mean ± standard deviation. Differences between two groups
were analysed using an unpaired t-test. Differences among three or
more experimental groups were analysed using one-way analysis of
variance, followed by Tukey's test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Upregulation of HIF-1α expression in
NSCLC cells
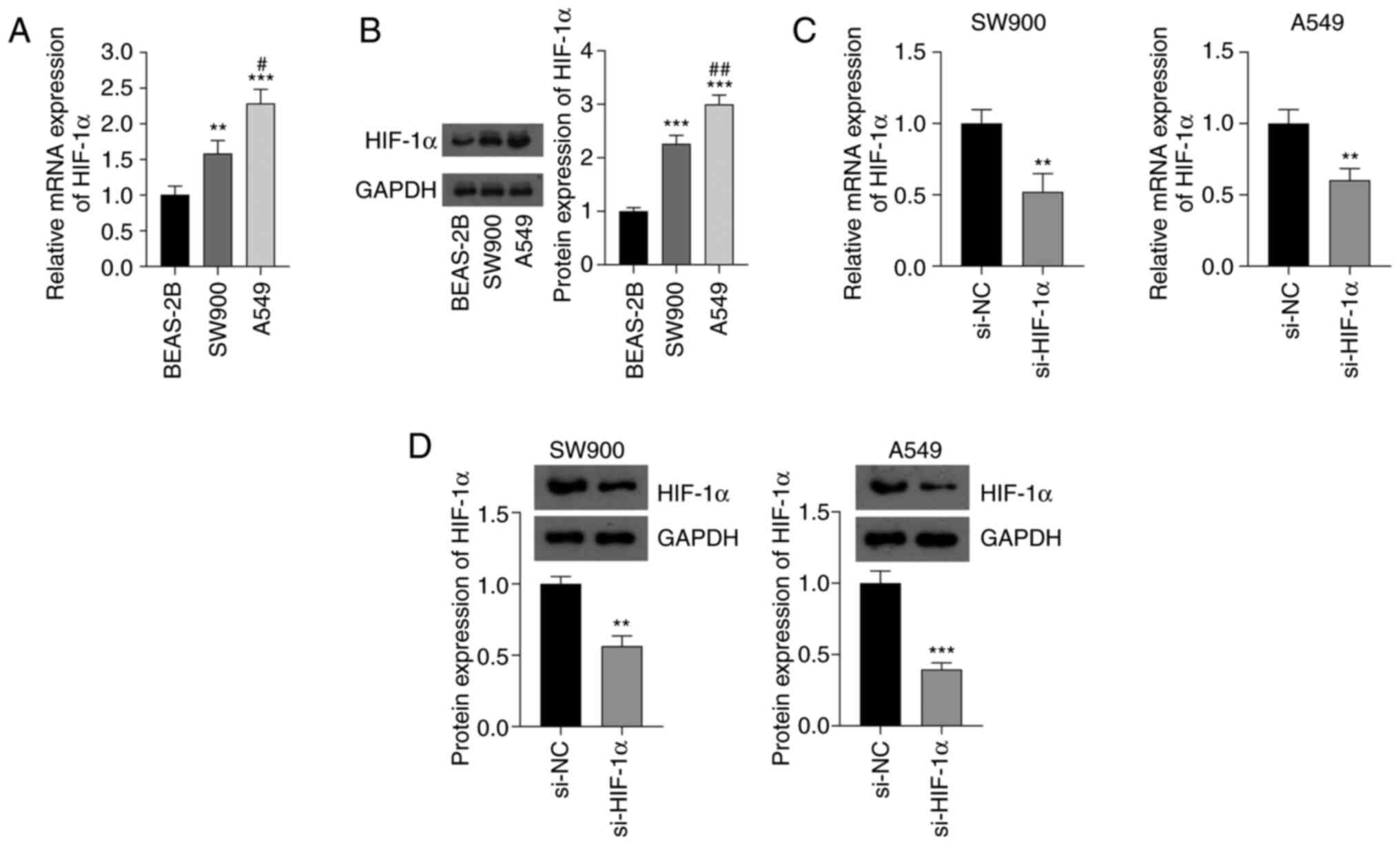
The expression levels of HIF-1α in SW900 and A549
NSCLC cell lines and BEAS-2B normal bronchial epithelial cells were
detected using RT-qPCR and western blotting. The mRNA and protein
expression levels of HIF-1α in the NSCLC cells were significantly
higher than those in the normal cells (P<0.01). In addition, the
mRNA and protein expression levels of HIF-1α in A549 cells were
higher than those in SW900 cells (P<0.05; Fig. 1A and B). The knockdown of HIF-1α in
NSCLC cells by transfection with si-HIF-1α significantly decreased
the mRNA and protein expression levels of HIF-1α compared with the
respective levels in cells transfected with si-NC (P<0.01;
Fig. 1C and D).
si-HIF-1α inhibits malignant
progression and induces ferroptosis in NSCLC cells
The effects of HIF-1α silencing on the malignant
progression of NSCLC were investigated. The CCK-8 assay showed that
HIF-1α silencing significantly suppressed the proliferation of
SW900 and A549 cells compared with the proliferation in the
respective si-NC group (P<0.05; Fig.
2A). Additionally, the flow cytometry results showed that the
silencing of HIF-1α significantly increased the apoptosis of SW900
and A549 cells (P<0.01; Fig.
2B). Moreover, the knockdown of HIF-1α significantly decreased
the invasion ability of SW900 and A549 cells compared with that of
the SW900 and A549 cells transfected with si-NC (P<0.01;
Fig. 2C). Based on the
aforementioned results, A549 cells were selected for further
analysis.
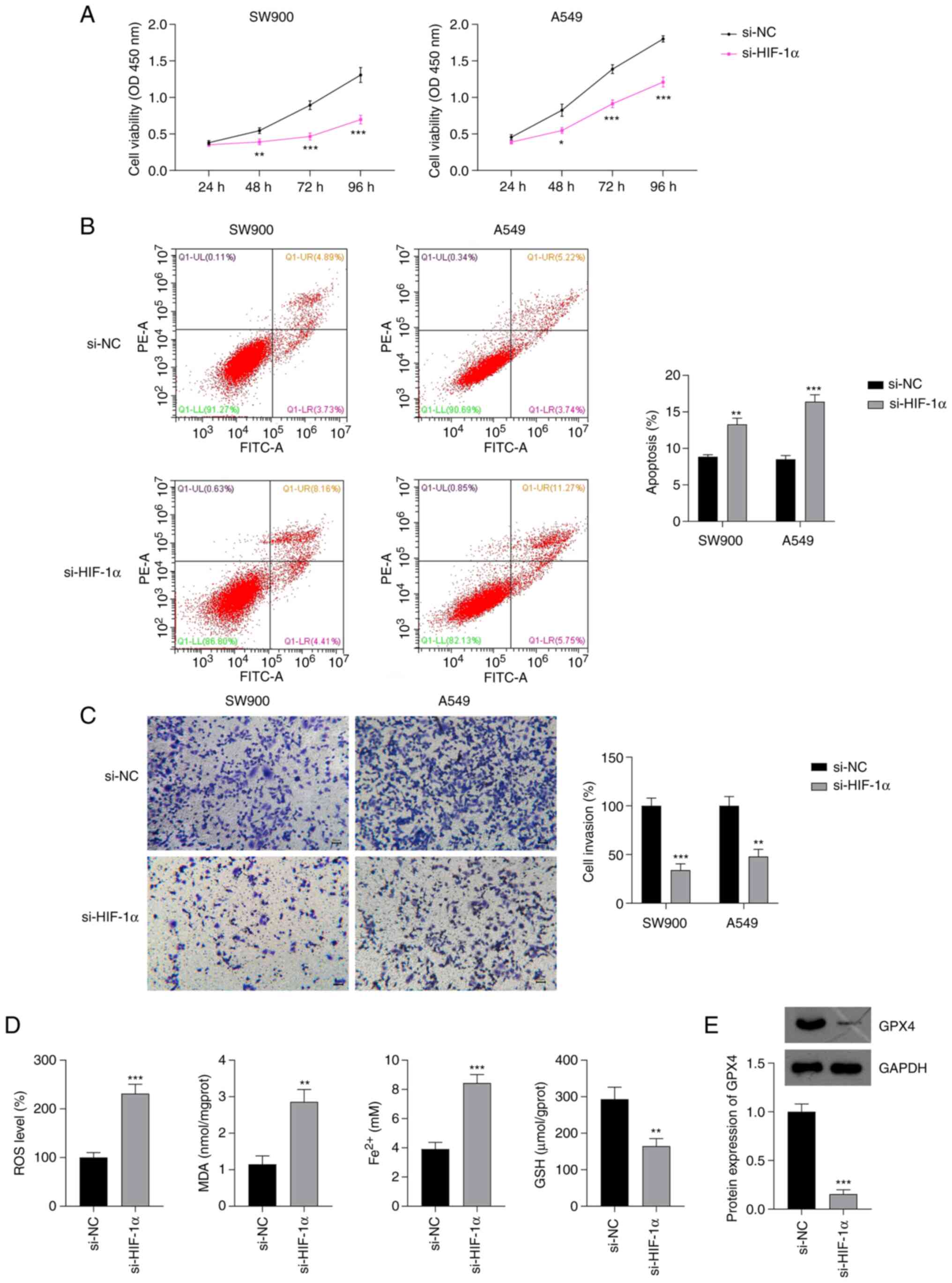 | Figure 2.Effect of si-HIF-1α on the
proliferation, invasion and ferroptosis of NSCLC cells. Results of
(A) Cell Counting Kit-8, (B) flow cytometry and (C) Transwell
invasion assays of NSCLC cells transfected with si-HIF-1α. (D)
Levels of ROS, MDA, Fe2+ and GSH in A549 cells
transfected with si-HIF-1α. Scale bar, 50 µm. (E) Protein
expression of GPX4 in A549 cells transfected with si-HIF-1α.
*P<0.05, **P<0.01 and ***P<0.001 vs. si-NC. si-HIF-1α,
small interfering RNA to hypoxia-inducible factor 1α; NSCLC,
non-small cell lung cancer; ROS, reactive oxygen species; MDA,
malondialdehyde; Fe2+, ferrous iron; GSH, glutathione;
GPX4, glutathione peroxidase 4; si-NC, small interfering RNA
negative control; OD, optical density. |
Lipid peroxidation, GSH depletion and iron
accumulation are reported to be critical events in ferroptosis
(33). To explore whether HIF-1α
expression affects ferroptosis in NSCLC, the levels of ROS, MDA,
GSH and Fe2+ in A549 cells were measured. si-HIF-1α
transfection significantly increased the levels of ROS, MDA and
Fe2+ and decreased the production of GSH in A549 cells,
compared with the levels in A549 cells transfected with si-NC
(P<0.01; Fig. 2D). GPX4 is a key
indicator of ferroptosis (34) and
has a regulatory relationship with HIF-1α (35). Therefore, the expression of GPX4 in
NSCLC cells was evaluated. Western blotting showed that the
knockdown of HIF-1α in A549 cells significantly downregulated GPX4
expression compared with that in the si-NC group (P<0.001;
Fig. 2E).
si-HIF-1α inhibits Hippo-YAP pathway
activation in NSCLC cells
The underlying mechanism by which si-HIF-1α induced
ferroptosis in NSCLC cells was then investigated. The Hippo-YAP
pathway regulates ferroptosis in cancer (36). Thus, cell viability was examined, as
well as the expression of the Hippo-YAP pathway regulator YAP1 and
ferroptosis marker GPX4 in A549 cells using CCK-8, RT-qPCR and
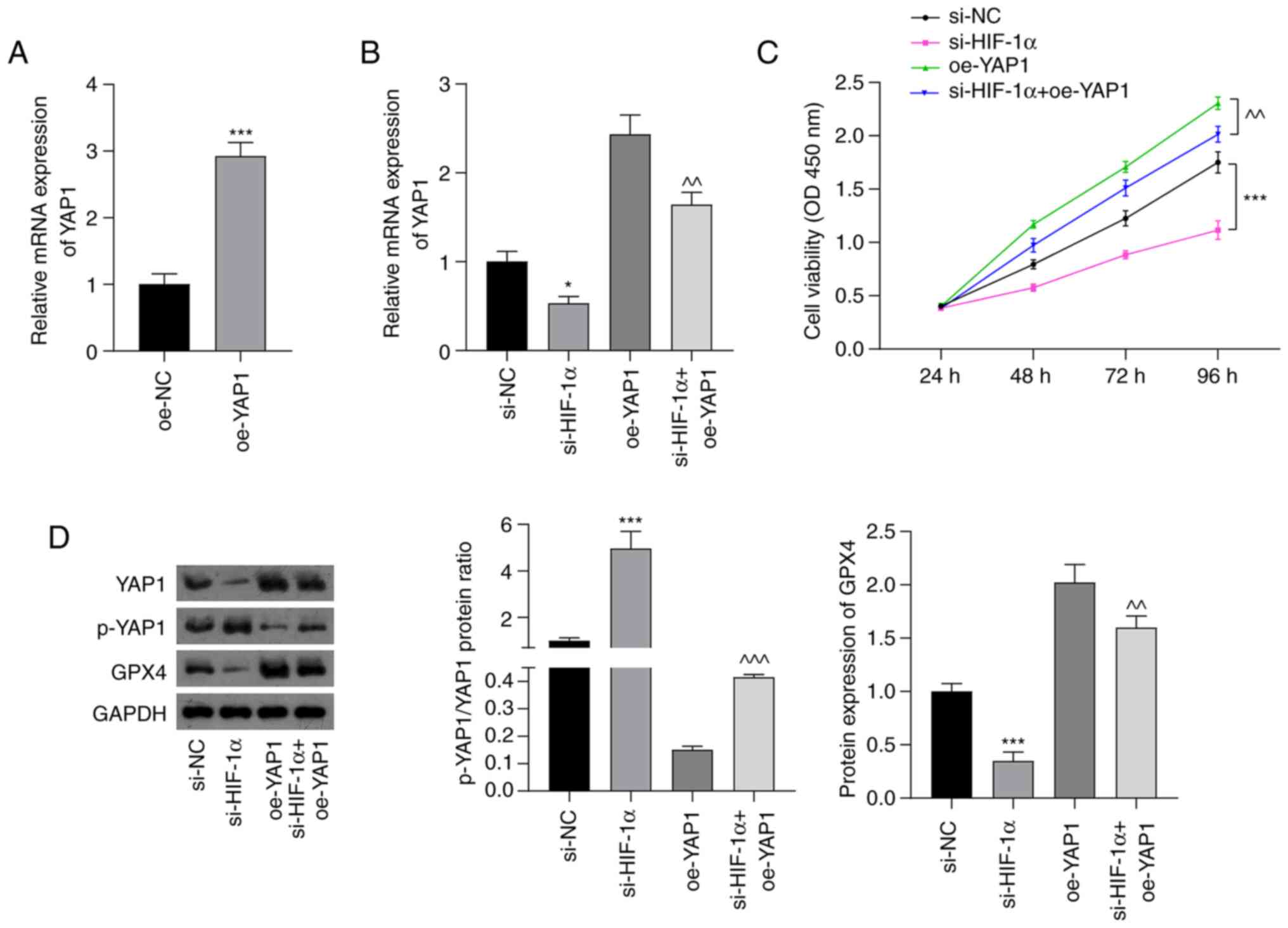
western blotting assays. As shown in Fig. 3A, YAP1 expression in A439 cells was
significantly increased following transfection with oe-YAP1
compared with that in cells transfected with oe-NC (P<0.001).
RT-qPCR analysis showed that suppression of HIF-1α significantly
downregulated the mRNA expression of YAP1 (P<0.05). Moreover,
the silencing of HIF-1α attenuated the oe-YAP1-induced upregulation
of YAP1 mRNA expression (P<0.01; Fig. 3B). The silencing of HIF-1α also
attenuated the increase in cell viability induced by oe-YAP1
transfection (P<0.01; Fig. 3C).
As shown in Fig. 3D, the
p-YAP1/YAP1 ratio was significantly upregulated in the si-HIF-1α
group compared with the si-NC group (P<0.001). The inhibitory
effect of oe-YAP1 on the p-YAP1/YAP1 ratio was significantly
attenuated by HIF-1α silencing (P<0.001). Furthermore, si-HIF-1α
transfection significantly suppressed GPX4 expression compared with
that in the si-NC group (P<0.001), and the increase in GPX4
expression induced by YAP1 overexpression was significantly
weakened by HIF-1α silencing (P<0.01).
si-HIF-1α promotes ferroptosis by
inhibiting Hippo-YAP pathway activation in NSCLC cells
To confirm that si-HIF-1α-induced ferroptosis plays
an important role in NSCLC treatment, the proliferation and
invasion of A549 cells, and the oxidative stress and iron
accumulation in these were cells investigated after treatment with
Fer-1 or erastin. As shown in Fig.
4A-C, Fer-1 significantly attenuated the inhibition of cell
viability and invasion in A549 cells induced by si-HIF-1α; by
contrast, erastin treatment augmented the si-HIF-1α-induced
suppression of A549 cell proliferation and invasion (P<0.05).
The enzyme-linked immunosorbent assay results indicated that Fer-1
significantly inhibited the si-HIF-1α-induced production of ROS,
MDA and Fe2+ and reduction in GSH levels in A549 cells
(P<0.01; Fig. 4D). By contrast,
erastin treatment increased the production of ROS, MDA and
Fe2+ and further reduced GSH levels in
si-HIF-1α-transfected A549 cells (P<0.01; Fig. 4D). Additionally, RT-qPCR and western
blot analyses showed that Fer-1 and erastin treatment did not alter
the HIF-1α mRNA and protein levels of the A549 cells. However,
Fer-1 significantly increased the levels of YAP1 mRNA and GPX4
protein and decreased the p-YAP1/YAP1 ratio compared with those in
the si-HIF-1α group (P<0.05), while erastin augmented the
si-HIF-1α-induced downregulation of YAP1 mRNA and GPX4 protein
expression and upregulation of the p-YAP1/YAP1 ratio (P<0.01;
Fig. 4E and F).
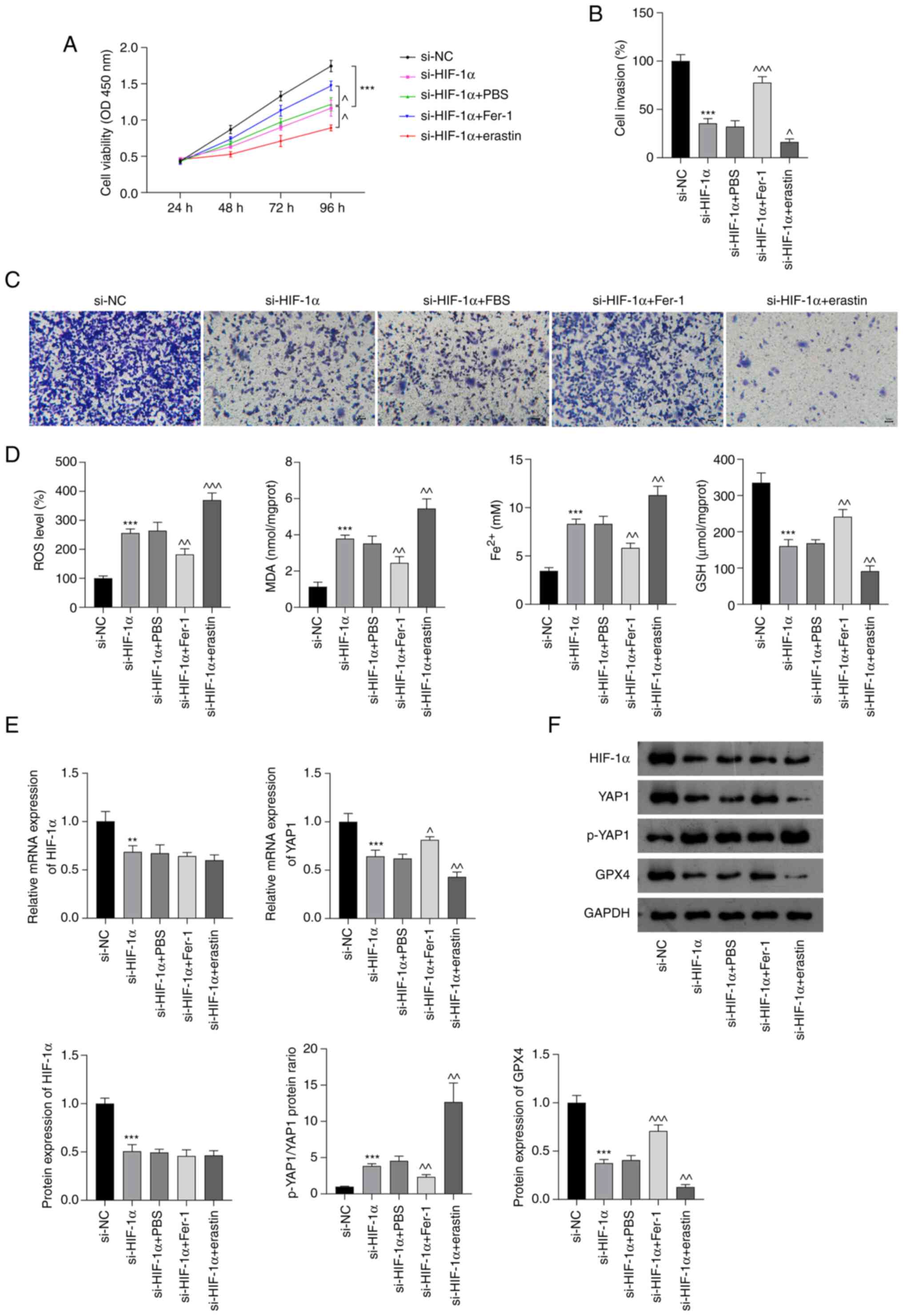 | Figure 4.si-HIF-1α promotes ferroptosis by
inhibiting Hippo-YAP pathway activation in non-small cell lung
cancer cells. (A) Cell Counting Kit-8 assay results. (B) Quantified
Transwell invasion assay results and (C) representative images of
invaded A549 cells. Scale bar, 50 µm. (D) ROS, MDA, Fe2+
and GSH levels in A549 cells. (E) Relative mRNA expression of
HIF-1α and YAP1 in A549 cells. (F) Protein levels of HIF-1α,
p-YAP1/YAP1 and GPX4 in A549 cells. **P<0.01 and ***P<0.001
vs. si-NC. ^P<0.05, ^^P<0.01 and
^^^P<0.001 vs. si-HIF-1α. HIF-1α, hypoxia-inducible
factor 1 α; si-HIF-1α, small interfering RNA to HIF-1α; YAP,
Yes-associated protein; p-, phosphorylated; ROS, reactive oxygen
species; MDA, malondialdehyde; Fe2+, ferrous iron; GSH,
glutathione; GPX4, glutathione peroxidase 4; PBS,
phosphate-buffered saline; Fer-1, ferrostatin-1; si-NC, small
interfering RNA negative control; OD, optical density. |
Discussion
In recent years, ferroptosis has emerged as a
strategy for cancer treatment. The present study evaluated the
ability of HIF-1α silencing to induce ferroptosis in NSCLC cells
and investigated whether the underlying mechanism involves the
inhibition of Hippo-YAP pathway activation.
Hypoxia is a common condition in the tumour
environment due to the high proliferation rate of tumour cells
(37–39). HIF-1α is an oncoprotein induced by
intratumoural hypoxia (40), which
promotes tumour cell proliferation, metastasis and migration
(15–17,39).
VEGF is a key contributor to angiogenesis, and the inhibition of
HIF-1α has been shown to decrease VEGF expression in malignant
glioma cells and reduce tumour growth (41). The present study confirmed that
HIF-1α is upregulated in NSCLC cells compared with normal bronchial
epithelial cells. Furthermore, the inhibition of HIF-1α
significantly decreased the proliferation and invasion of NSCLC
cells and increased their apoptosis. Therefore, the inhibition of
HIF-1α shows potential as a mechanism for NSCLC therapy.
As HIF-1α contributes to the inhibition of
ferroptosis in hepatocellular carcinoma, it has been reported as a
potential biomarker of poor post-operative outcomes in this disease
(21). Ferroptosis is a form of
iron-dependent, non-apoptotic cell death induced by lipid
peroxidation, system xc− inhibitors and ROS
accumulation (42,43). Hypoxia accelerates lipid
peroxidation, depletes intracellular GSH (44) and prevents ferroptosis (45,46).
These activities were evaluated in the present study following the
knockdown of HIF-1α. The results revealed that HIF-1α knockdown
significantly increased the ROS, MDA and Fe2+ levels and
decreased the GSH levels of NSCLC cells, suggesting that HIF-1α
silencing facilitates the ferroptosis of these cells. Further, GPX4
is a prominent regulator of ferroptosis in cancer cells, and GSH
depletion has been demonstrated to induce GPX4 inactivation and
ferroptosis (34). Additionally,
other studies have shown that the inhibition GPX4 leads to lipid
peroxidation and ferroptosis (42,47,48).
The present study showed that the transfection of NSCLC cells with
si-HIF-1α downregulated GPX4 expression, which is consistent with
the findings of previous research (35). In addition, the effects of Fer-1 and
erastin, which are classical antagonists and agonists of
ferroptosis, respectively were investigated. Fer-1 reversed the
effects of si-HIF-1α on proliferation, invasion, ROS production,
and Fe2+ and GPX4 levels in NSCLC cells, whereas erastin
enhanced these effects. These results support the hypothesis that
HIF-1α silencing inhibits malignant progression by promoting
ferroptosis in NSCLC cells.
The Hippo signalling pathway has tumour suppressing
effects, and YAP acts as an oncogene in most tumour cells (24,49).
Hypoxia promotes the nuclear localisation of YAP and decreases the
phosphorylation of YAP in cancer cells (24). In addition, YAP promotes the
proliferation and chemoresistance of cancer cells (26,50,51),
and also contributes to the invasion and migration of NSCLC cells
(26). A previous study has shown
that hypoxia enhances the binding of YAP to HIF-1α in the nucleus
and maintains the stability of HIF-1α protein (24). The results of the present study
showed that transfection with si-HIF-1α decreased the expression of
YAP1 in NSCLC cells, and attenuated the oe-YAP1-induced changes in
YAP1 and p-YAP1 levels, suggesting that the silencing of HIF-1α
inhibits activation of the Hippo-YAP pathway. Furthermore, the
upregulation of GPX4 induced by oe-YAP1 was attenuated by HIF-1α
silencing. These results indicate that the silencing of HIF-1α
promotes ferroptosis by suppressing Hippo-YAP signalling pathway
activation.
The present study had some limitations. First, the
pathological mechanisms affecting the growth of cancer cells are
complex, and ferroptosis is only one such mechanism; the other
mechanisms that may be involved were not evaluated. Second, HIF-1α
affects NSCLC cell ferroptosis and may be involved in multiple
pathways in addition to the Hippo-YAP pathway. Finally, additional
experimental data obtained from techniques such as confocal
microscopy or fluorescent probe analyses are required to confirm
the results.
In conclusion, HIF-1α expression is upregulated in
NSCLC cells. The silencing of HIF-1α inhibited the proliferation
and invasion of NSCLC cells and induced their ferroptosis; it also
suppressed activation of the Hippo-YAP pathway. Moreover, erastin
further enhanced the effect of si-HIF-1α whereas Fer-1 counteracted
the effect of si-HIF-1α. These results suggest that HIF-1α
silencing promotes ferroptosis in NSCLC cells via the inhibition of
Hippo-YAP pathway activation. Therefore, HIF-1α is a potential
target for NSCLC therapy.
Acknowledgements
Not applicable.
Funding
This study was supported by the Project of Taizhou Science and
Technology Bureau (grant no. 21ywb51).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SZ, JM and YC contributed to the conception and the
design of the study. SZ, JM and JZ were responsible for the
acquisition, analysis and interpretation of the data. SZ, JZ and YC
confirm the authenticity of all the raw data. SZ and JM contributed
to manuscript drafting and critical revisions of intellectual
content. SZ obtained the funding. YC approved the final manuscript
for publication. All authors have read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
NSCLC
|
non-small cell lung cancer
|
|
ROS
|
reactive oxygen species
|
|
VEGF
|
vascular endothelial growth factor
|
|
YAP1
|
Yes-associated protein 1
|
|
GPX4
|
glutathione peroxidase 4
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
CCK-8
|
Cell Counting Kit-8
|
|
OD
|
optical density
|
|
MDA
|
malondialdehyde
|
|
GSH
|
glutathione
|
References
|
1
|
Fashoyin-Aje LA, Fernandes LL, Sridhara R,
Keegan P and Pazdur R: Demographic composition of lung cancer
trials: FDA analysis. J Clin Oncol. 36 (15_suppl):S90882018.
View Article : Google Scholar
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Salgia R, Pharaon R, Mambetsariev I, Nam A
and Sattler M: The improbable targeted therapy: KRAS as an emerging
target in non-small cell lung cancer (NSCLC). Cell Rep Med.
2:1001862021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Goodgame B, Viswanathan A, Zoole J, Gao F,
Miller CR, Subramanian J, Meyers BF, Patterson AG and Govindan R:
Risk of recurrence of resected stage I non-small cell lung cancer
in elderly patients as compared with younger patients. J Thorac
Oncol. 4:1370–1374. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Imai H, Matsuoka M, Kumagai T, Sakamoto T
and Koumura T: Lipid peroxidation-dependent cell death regulated by
GPx4 and ferroptosis. Curr Top Microbiol Immunol. 403:143–170.
2017.PubMed/NCBI
|
|
7
|
Tang X, Ding H, Liang M, Chen X, Yan Y,
Wan N, Chen Q, Zhang J and Cao J: Curcumin induces ferroptosis in
non-small-cell lung cancer via activating autophagy. Thoracic
Cancer. 12:1219–1230. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nie J, Lin B, Zhou M, Wu L and Zheng T:
Role of ferroptosis in hepatocellular carcinoma. J Cancer Res Clin
Oncol. 144:2329–2337. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liang C, Zhang X, Yang M and Dong X:
Recent progress in ferroptosis inducers for cancer therapy. Adv
Mater. 31:e19041972019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shen Z, Song J, Yung BC, Zhou Z, Wu A and
Chen X: Emerging strategies of cancer therapy based on ferroptosis.
Adv Mater. 30:e17040072018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Eling N, Reuter L, Hazin J, Hamacher-Brady
A and Brady NR: Identification of artesunate as a specific
activator of ferroptosis in pancreatic cancer cells. Oncoscience.
2:517–532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li ZJ, Dai HQ, Huang XW, Feng J, Deng JH,
Wang ZX, Yang XM, Liu YJ, Wu Y, Chen PH, et al: Artesunate
synergizes with sorafenib to induce ferroptosis in hepatocellular
carcinoma. Acta Pharmacol Sin. 42:301–310. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hao S, Yu J, He W, Huang Q, Zhao Y, Liang
B, Zhang S, Wen Z, Dong S, Rao J, et al: Cysteine dioxygenase 1
mediates erastin-induced ferroptosis in human gastric cancer cells.
Neoplasia. 19:1022–1032. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Yan H, Xu X, Liu H, Wu C and Zhao L:
Erastin/sorafenib induces cisplatin-resistant non-small cell lung
cancer cell ferroptosis through inhibition of the Nrf2/xCT pathway.
Oncol Lett. 19:323–333. 2020.PubMed/NCBI
|
|
15
|
Li H, Jia Y and Wang Y: Targeting HIF-1α
signaling pathway for gastric cancer treatment. Pharmazie. 74:3–7.
2019.PubMed/NCBI
|
|
16
|
Qin Y, Liu HJ, Li M, Zhai DH, Tang YH,
Yang L, Qiao KL, Yang JH, Zhong WL, Zhang Q, et al: Salidroside
improves the hypoxic tumor microenvironment and reverses the drug
resistance of platinum drugs via HIF-1α signaling pathway.
EBioMedicine. 38:25–36. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pezzuto A and Carico E: Role of HIF-1 in
cancer progression: Novel insights. A review. Curr Mol Med.
18:343–351. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park SY, Jeong KJ, Lee J, Yoon DS, Choi
WS, Kim YK, Han JW, Kim YM, Kim BK and Lee HY: Hypoxia enhances
LPA-induced HIF-1alpha and VEGF expression: Their inhibition by
resveratrol. Cancer Lett. 258:63–69. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim DH, Sung B, Kang YJ, Hwang SY, Kim MJ,
Yoon JH, Im E and Kim ND: Sulforaphane inhibits hypoxia-induced
HIF-1α and VEGF expression and migration of human colon cancer
cells. Int J Oncol. 47:2226–2232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma Z, Xiang X, Li S, Xie P, Gong Q, Goh BC
and Wang L: Targeting hypoxia-inducible factor-1, for cancer
treatment: Recent advances in developing small-molecule inhibitors
from natural compounds. Semin Cancer Biol. 80:379–390. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao J, Zeng G, Lin E, Cai C, Li P, Zou B
and Li J: Combined HIF-1α and SHH Up-Regulation Is a Potential
Biomarker to Predict Poor Prognosis in Postoperative Hepatocellular
Carcinoma. J Invest Surg. 35:1660–1667. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin Z, Song J, Gao Y, Huang S, Dou R,
Zhong P, Huang G, Han L, Zheng J, Zhang X, et al: Hypoxia-induced
HIF-1α/lncRNA-PMAN inhibits ferroptosis by promoting the
cytoplasmic translocation of ELAVL1 in peritoneal dissemination
from gastric cancer. Redox Biol. 52:1023122022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang G, Dai S, Chen Y, Wang H, Chen T,
Shu Q, Chen S, Shou L and Cai X: Aqueous extract of Taxus chinensis
var. mairei regulates the Hippo-YAP pathway and promotes apoptosis
of non-small cell lung cancer via ATF3 in vivo and in vitro. Biomed
Pharmacother. 138:1115062021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Li Y, Ma Y, Yang L, Wang T, Meng
X, Zong Z, Sun X, Hua X and Li H: Yes-associated protein (YAP)
binds to HIF-1α and sustains HIF-1α protein stability to promote
hepatocellular carcinoma cell glycolysis under hypoxic stress. J
Exp Clin Cancer Res. 37:2162018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li C, Wang S, Xing Z, Lin A, Liang K, Song
J, Hu Q, Yao J, Chen Z, Park PK, et al: A ROR1-HER3-lncRNA
signalling axis modulates the Hippo-YAP pathway to regulate bone
metastasis. Nat Cell Biol. 19:106–119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu M, Chen Y, Li X, Yang R, Zhang L,
Huangfu L, Zheng N, Zhao X, Lv L, Hong Y, et al: YAP1 contributes
to NSCLC invasion and migration by promoting Slug transcription via
the transcription co-factor TEAD. Cell Death Dis. 9:4642018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y and Liu S: LncRNA GHET1 promotes
hypoxia-induced glycolysis, proliferation, and invasion in
triple-negative breast cancer through the Hippo/YAP signaling
pathway. Front Cell Dev Biol. 9:6435152021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang WH, Ding CC, Sun T, Rupprecht G, Lin
CC, Hsu D and Chi JT: The Hippo pathway effector TAZ regulates
ferroptosis in renal cell carcinoma. Cell Rep. 28:2501–2508.e4.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu G, Murshed A, Li H, Ma J, Zhen N, Ding
M, Zhu J, Mao S, Tang X, Liu L, et al: O-GlcNAcylation enhances
sensitivity to RSL3-induced ferroptosis via the YAP/TFRC pathway in
liver cancer. Cell Death Discov. 7:832021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ye S, Xu M, Zhu T, Chen J, Shi S, Jiang H,
Zheng Q, Liao Q, Ding X and Xi Y: Cytoglobin promotes sensitivity
to ferroptosis by regulating p53-YAP1 axis in colon cancer cells. J
Cell Mol Med. 25:3300–3311. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cao X, Li Y, Wang Y, Yu T, Zhu C, Zhang X
and Guan J: Curcumin suppresses tumorigenesis by ferroptosis in
breast cancer. PLoS One. 17:e02613702022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stockwell BR, Friedmann Angeli JP, Bayir
H, Bush AI, Conrad M, Dixon SJ, Fulda S, Gascón S, Hatzios SK,
Kagan VE, et al: Ferroptosis: A regulated cell death nexus linking
metabolism, redox biology, and disease. Cell. 171:273–285. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang WS, SriRamaratnam R, Welsch ME,
Shimada K, Skouta R, Viswanathan VS, Cheah JH, Clemons PA, Shamji
AF, Clish CB, et al: Regulation of ferroptotic cancer cell death by
GPX4. Cell. 156:317–331. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang Y, Tang H, Zheng J and Yang K: The
PER1/HIF-1alpha negative feedback loop promotes ferroptosis and
inhibits tumor progression in oral squamous cell carcinoma. Transl
Oncol. 18:1013602022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang R and Zhu G: A narrative review for
the Hippo-YAP pathway in cancer survival and immunity: the Yin-Yang
dynamics. Transl Cancer Res. 11:262–275. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tang T, Yang Z, Zhu Q, Wu Y, Sun K,
Alahdal M, Zhang Y, Xing Y, Shen Y, Xia T, et al: Up-regulation of
miR-210 induced by a hypoxic microenvironment promotes breast
cancer stem cell metastasis, proliferation, and self-renewal by
targeting E-cadherin. FASEB J. Sep 6–2018.(Epub ahead of print).
View Article : Google Scholar
|
|
38
|
Marhuenda E, Campillo N, Gabasa M,
Martínez-García MA, Campos-Rodríguez F, Gozal D, Navajas D, Alcaraz
J, Farré R and Almendros I: Effects of sustained and intermittent
hypoxia on human lung cancer cells. Am J Respir Cell Mol Biol.
61:540–544. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang L, Zhang W, Wang Y, Zou T, Zhang B,
Xu Y, Pang T, Hu Q, Chen M, Wang L, et al: Hypoxia-induced miR-214
expression promotes tumour cell proliferation and migration by
enhancing the Warburg effect in gastric carcinoma cells. Cancer
Lett. 414:44–56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pawlus MR and Hu CJ: Enhanceosomes as
integrators of hypoxia inducible factor (HIF) and other
transcription factors in the hypoxic transcriptional response. Cell
Signal. 25:1895–1903. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jensen RL, Ragel BT, Whang K and Gillespie
D: Inhibition of hypoxia inducible factor-1alpha (HIF-1alpha)
decreases vascular endothelial growth factor (VEGF) secretion and
tumor growth in malignant gliomas. J Neurooncol. 78:233–247. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Forcina GC and Dixon SJ: GPX4 at the
crossroads of lipid homeostasis and ferroptosis. Proteomics.
19:e18003112019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xia X, Fan X, Zhao M and Zhu P: The
relationship between ferroptosis and tumors: A novel landscape for
therapeutic approach. Curr Gene Ther. 19:117–124. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang X, Wu M, Zhang X, Li F, Zeng Y, Lin
X, Liu X and Liu J: Hypoxia-responsive nanoreactors based on
self-enhanced photodynamic sensitization and triggered ferroptosis
for cancer synergistic therapy. J Nanobiotechnology. 19:2042021.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fan Z, Yang G, Zhang W, Liu Q, Liu G, Liu
P, Xu L, Wang J, Yan Z, Han H, et al: Hypoxia blocks ferroptosis of
hepatocellular carcinoma via suppression of METTL14 triggered
YTHDF2-dependent silencing of SLC7A11. J Cell Mol Med.
25:10197–10212. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wu Y, Wang J, Zhao T, Chen J, Kang L, Wei
Y, Han L, Shen L, Long C, Wu S and Wei G: Di-(2-ethylhexyl)
phthalate exposure leads to ferroptosis via the HIF-1α/HO-1
signaling pathway in mouse testes. J Hazard Mater. 426:1278072022.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shin D, Kim EH, Lee J and Roh JL: Nrf2
inhibition reverses resistance to GPX4 inhibitor-induced
ferroptosis in head and neck cancer. Free Radic Biol Med.
129:454–462. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gong Y, Wang N, Liu N and Dong H: Lipid
peroxidation and GPX4 inhibition are common causes for
myofibroblast differentiation and ferroptosis. DNA Cell Biol.
38:725–733. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yamamura S, Goda N, Akizawa H, Kohri N,
Balboula AZ, Kobayashi K, Bai H, Takahashi M and Kawahara M:
Yes-associated protein 1 translocation through actin cytoskeleton
organization in trophectoderm cells. Dev Biol. 468:14–25. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Coelho MA, de Carné Trécesson S, Rana S,
Zecchin D, Moore C, Molina-Arcas M, East P, Spencer-Dene B, Nye E,
Barnouin K, et al: Oncogenic RAS signaling promotes tumor
immunoresistance by stabilizing PD-L1 mRNA. Immunity.
47:1083–1099.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhou Y, Yang R and Ma G: YAP1 knockdown
suppresses the proliferation, migration and invasion of human
nasopharyngeal carcinoma cells. Nan Fang Yi Ke Da Xue Xue Bao.
39:286–291. 2019.(In Chinese). PubMed/NCBI
|