CRC is the third most common cancer type in the
world (after breast cancer and lung cancer), and the second leading
cause of cancer-related death worldwide (after lung cancer)
(1). Although the incidence of CRC
in individuals aged 65 years and older has decreased in most
countries (2), the incidence in
individuals under the age of 50 years has been increasing worldwide
(3). CRC is a heterogeneous
neoplastic disease caused by the malignant degeneration of the
external epithelium of the large intestine. Its development follows
the sequential accumulation of various carcinogenic mutations,
namely the adenoma-carcinoma sequence, with dysregulation of
signalling pathways such as WNT/β-catenin, p53 and TGF-β-Smads
(4).
Common pathogenic factors of CRC include genetic
mutation, poor diet, obesity and sedentary lifestyle (5,6). CRC
is usually an asymptomatic disease until it progresses to an
advanced stage. As a result, numerous patients with CRC are
clinically diagnosed with advanced disease (7). Since adenomas take at least 5–10 years
to develop into CRC, early endoscopic screening is crucial for the
diagnosis of CRC. In recent decades, although the understanding of
CRC has markedly improved at the molecular and genetic levels, it
has not brought an equivalent level of clinical benefits to
patients. Furthermore, the clinical outcomes and therapeutic
responses of patients with CRC differ significantly from the
expected outcomes. Numerous drugs have been found to be effective
in in vitro cancer models but have eventually failed in
clinical trials, indicating that there are marked differences
between the existing research model system and clinical practice
(8). The current landscape of
antitumor drug development relies heavily on traditional models,
such as two-dimensional cell lines (2D) and patient-derived tumor
xenografts (PDXs). However, these models have inherent limitations
that hinder their effectiveness. Cell lines, for instance, fail to
preserve the genetic information and heterogeneity of tumors during
prolonged passaging. In addition, PDX models experience tumor
evolution specific to mice and they differ in genetic
characteristics and growth environments from those in human tumor
patients. Furthermore, PDX models suffer from a low success rate,
high cost and time-consuming procedures (9). Therefore, it is urgent to establish a
model that may accurately reflect the genetic diversity and
specificity of cancer to better aid in the clinical diagnosis and
treatment of colon cancer.
In recent years, tumor organoids have emerged as a
highly promising tool in the field of cancer research. Organoids, a
3D structured in vitro culture system containing
self-renewing stem cells, have organization and function similar to
those of an organ. This system overcomes many of the limitations of
traditional models. These organoids more closely resemble the
cellular composition, behaviour and physiology of natural tissues.
Currently, researchers are able to generate cancer-like organoids
derived from mouse or human tumor tissues. Under appropriate
culture conditions, the organoid forms a three-dimensional
structure similar to a mouse or human tumor and mostly maintains
the tumor tissue structure, gene profile and heterogeneity observed
in the original tumor tissue. Tumor organoids are able to
accurately report the drug response of the corresponding patient to
determine a more effective treatment plan for individual patients
(10,11). In this paper, the history of
organoid culture systems, their application in CRC and the
challenges they face as preclinical models in CRC research were
reviewed.
Currently, preclinical cancer models mainly include
cancer cell lines (CCLs), PDXs and organoids (12). CCLs are still widely used models for
large-scale drug screening. However, due to their inability to
accurately simulate the microenvironment of the original tumor
growing in 3D, they do not accurately represent the heterogeneous
characteristics of cancer cells in vivo (13). There is also a lack of corresponding
cell lines derived from normal tissues as a reference control
(14). There are significant
differences in cell morphology and drug susceptibility between 2D
cell lines and 3D cell models (15). PDXs can, to a certain extent,
preserve cell interactions and capture tumor heterogeneity in 3D
environments; however, they take a long time and have a low success
rate (13,16).
Recently, a novel 3D culture technique has
facilitated the development of organoid models and numerous
researchers have similar definitions of organoids. Organoids refer
to stem cells (including embryonic stem cells, adult stem cells,
induced pluripotent stem cells, or tumor stem cells) embedded in
extracellular matrix (ECM) produced in a specific environment and
have specific organ structures and functions (17–20).
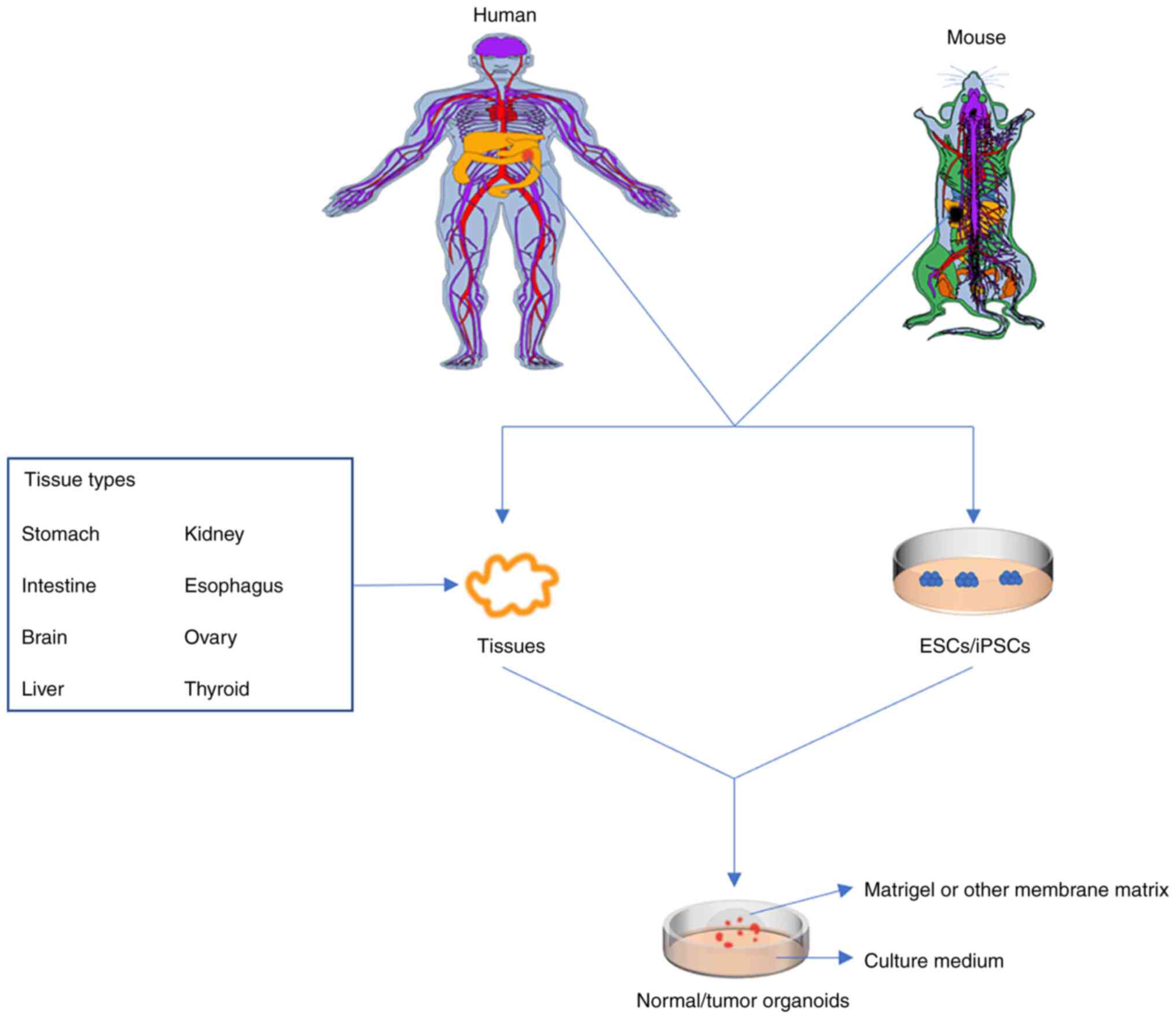
Organoids may be created from almost any tissue, such as brain
(21), intestine (22), thyroid gland (23), mammary gland (24), stomach (25), liver (26), ovaries (27), kidney (28) and oesophagus (29). Successfully constructed organoids
may effectively mimic the morphological structure and epigenetics
of both normal tissues and cancer tissues. It has been established
that the general culture method of organoids involves the use of
ECM hydrogels, such as Matrigel or basement membrane extract (BME),
the addition of various growth factors to various tissues, the use
of a gas-liquid interface, and the coculture of immune cells to
mimic the tumor microenvironment to simulate the matrix environment
for 3D culture. Organoid culture in vitro may be used for
intervention studies prior to clinical treatment, such as those for
drug sensitivity screening, new drug development, personalized
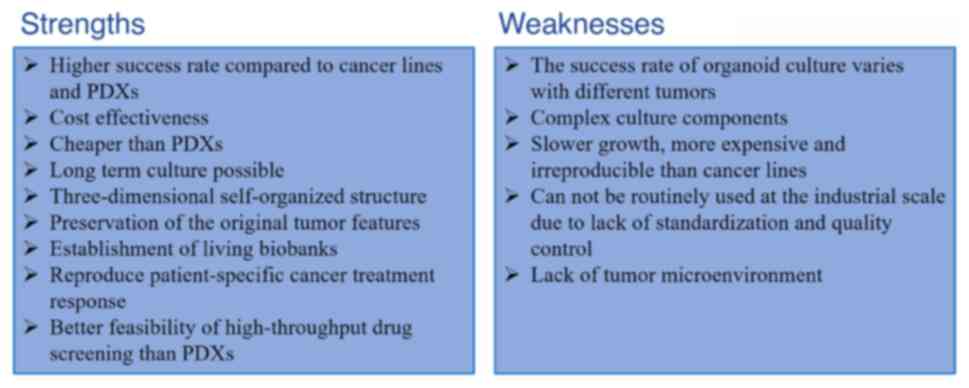
medicine and regenerative medicine (Fig. 1). Of note, intestinal organoids are
thought to be superior models to CCLs and PDXs for investigating
cancer genetics, cancer processes and antitumor drug activity,
since they make up for the inadequacies of existing models
(30) (Fig. 2).
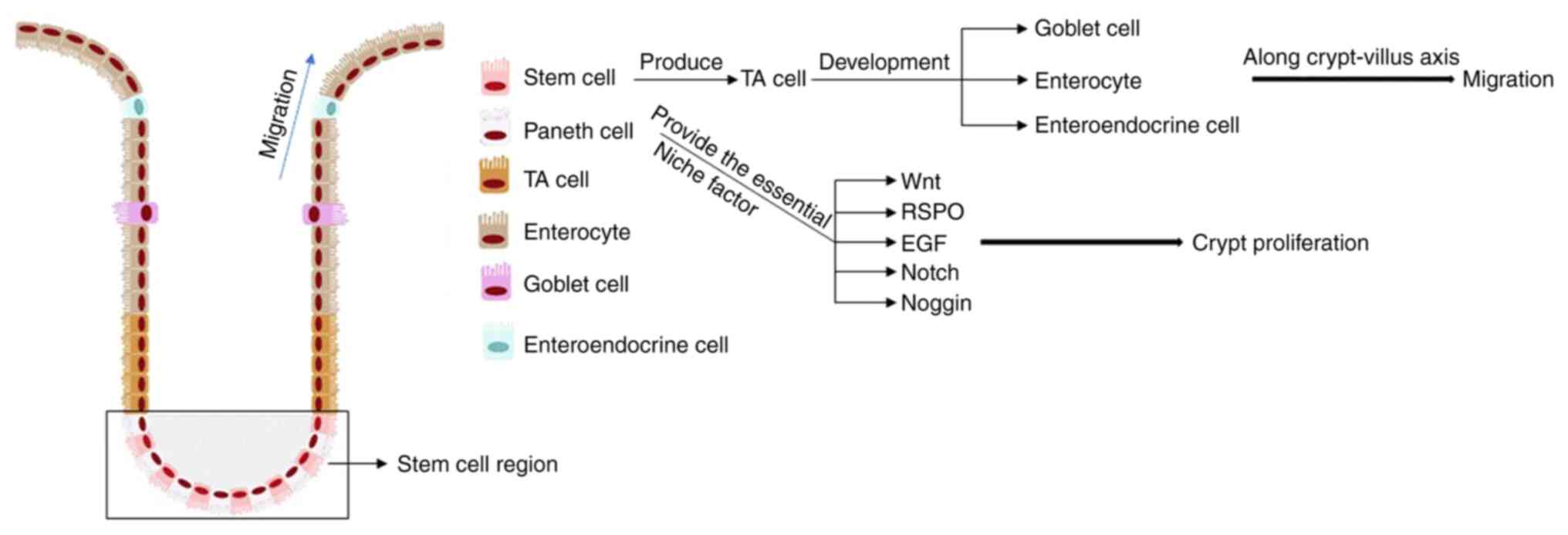
The mammalian intestinal epithelium consists of a
single columnar epithelium containing resorptive and secretory
cells that allow them to absorb nutrients and protect the intestine
(31). The intestinal epithelium is
the most rapidly self-renewing tissue in adult mammals and the
villus-crypt structure is completely renewed every 4–5 days. The
villi-crypt structure is the basic building block of the intestinal
epithelium, which is proliferated and differentiated by intestinal
stem cells at the base of the crypt into mature intestinal
functional cells (32,33) (Fig.
3). However, the specific location of intestinal stem cells is
not well defined, so it is difficult to study their role. Barker
et al (34) found that
leucine-rich G-protein-paired receptor 5 (Lgr5, Wnt target gene)
was a marker gene of intestinal stem cells. Subsequently, Sato
et al (35) found normal
mouse intestinal stem cells at the base of the crypt and
differentiated them into self-organized ‘mini small intestine’,
establishing the first 3D organoid. The characteristics of the
crypts remained after 8 months of culture (35). They went on to successfully create
organoids from normal human cells and human tumor colon epithelial
cells, and improve the colon culture system (long-term culture
requires niacin amide, a small molecule inhibitor of Alk and p38
inhibitors) (Table I) (36–42).
In addition to their presence in the gut, Lgr5+ stem cells may be
found in other tissues and organs, e.g., the liver (43), pancreas (44), ovaries (45), stomach (46), kidneys (47) and lung (48,49).
At present, almost all human tissues may be cultured in
vitro to produce organoids (50).
Intestinal organoids may also be derived from the
progressive differentiation of human pluripotent stem cells
(51,52). Spence et al (53) were the first to successfully
construct intestinal organoids using human pluripotent stem cells.
They first induced PSC differentiation into a defined endoderm by
activin A (TGF-β molecule) and then treated them with a medium
containing a combination of fibroblast growth factor 4 and Wnt3A to
form post intestinal spheres. Finally, this was transferred to
cultures that a known to facilitate the formation of organoids,
resulting in polarized columnar epithelial cells containing goblet
cells, Pan's cells and intestinal endocrine cells (53). Transient activation of bone
morphogenetic protein signalling is essential for organoid
development in the hindgut (54).
Of note, CRC organoids have received growing recognition in studies
of intestinal organoids.
Patient-derived colorectal carcinoids are mainly
derived from surgical resection specimens, biopsies and stem cells.
Organoid culture medium for normal human colon tissue was used with
minor modifications for CRC organoid culture (Table I). There are currently no
standardized methods for CRC organoid culture The current methods
mainly involve the derivation of organoids on polystyrene-coated
polydimethylsiloxane (PDMS) microporous matrix or on flat matrix
gel. The former method is well suited for high-throughput clonal
culture and subsequent retrieval of individual organoids. However,
polystyrene-coated PDMS may impair organoid development. The latter
method enables imaging and tracking of organoids, similar to the
traditional 3D culture environment, which has the disadvantages of
low throughput and poor reproducibility (13). Matrix is a prerequisite for cell
growth, proliferation and differentiation. In addition, researchers
have found that matrix gels are not suitable for high-throughput
drug screening and developed type I collagen gels instead of matrix
gels for organoid culture (62,63).
The combination of patient-derived CRC organoids with the
orthotopic transplantation model enables the tumor to grow in the
natural colon environment of mice, more accurately simulating the
development of CRC and liver metastasis (64). Researchers have successfully
introduced mutations into organoids derived from normal intestinal
epithelial cells to prepare CRC organoids for disease studies
(56). Thanks to the significant
improvements of methods in organoid cultures, CRC organoids have
been widely applied in preclinical studies of CRC.
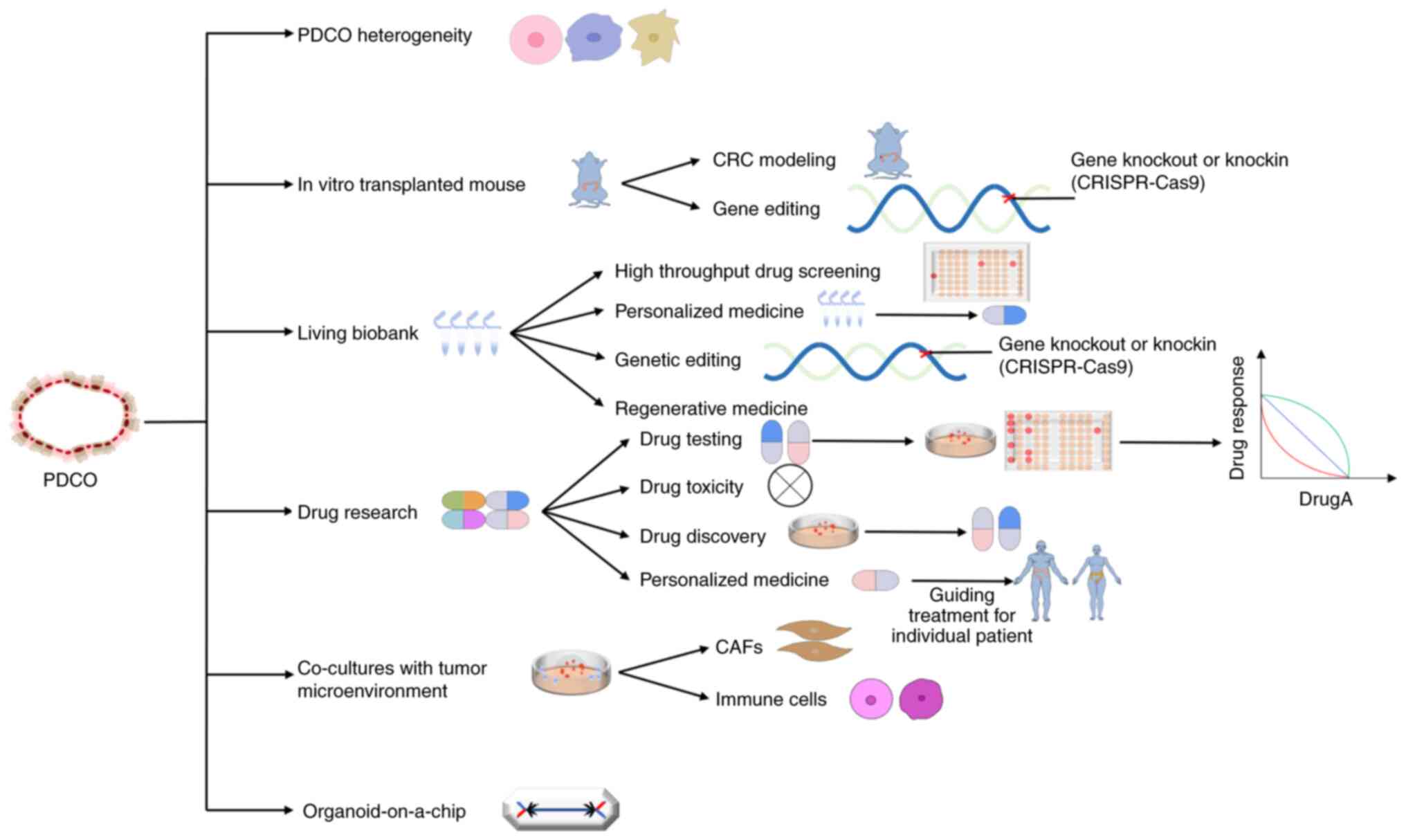
Currently, organoids are considered to be the best
preclinical models, as they may maintain genetic diversity while
being capable of long-term stable expansion, cryopreservation and
simulation of the link between cell polarity and tissue changes
during cancer progression. Organoid models have applications in the
study of cell heterogeneity, the construction of disease models,
drug screening, drug toxicity testing, the invention of new drugs
and the practice of personalized medicine. With the combination of
CRISPR-Cas gene editing technology, organoid models have become a
superior and unique tool for analysing gene function (65). Intestinal organoids are the first
successfully cultured organoids and are currently the most
developed and well-researched organoids. Studies based on
intestinal tumor organoids are also more extensive. The types of
CRC studies that have been conducted using intestinal tumor
organoids as models are summarized in Fig. 4 and limitations of the application
of CRC organoids are discussed below.
Epigenetic and environmental factors are primary
determinants of tumor heterogeneity which may be further subdivided
into two main categories: Intratumor heterogeneity and intertumor
heterogeneity (66). The cancer
stem cell theory demonstrates the existence of intron clonal
heterogeneity, which further complicates the heterogeneity of CRC
tumors (67,68). In recent years, CRC organoids
constructed from tissues of different parts of the same tumor or
tissues of different malignancy degrees of the same tumor have
largely maintained the characteristics of the original samples.
These organoids have been studied by cancer genomics (41,69),
multiregion sequencing (70),
single-cell derived cloning (71,72)
and single-cell sequencing (73,74) to
reveal the heterogeneity of CRC.
Tumor heterogeneity is specifically responsible for
the multiple capabilities and biological characteristics of the
tumor, which make it more prone to metastasis, recurrence and drug
resistance (75,76). Kim et al (77) performed whole-exome sequencing from
a multiregion-generating organoid tumor model in 12 patients,
revealing intraregional tumor heterogeneity in individual patients.
They tested 24 drugs and found that tumors with high intratumor
heterogeneity showed elevated expression of oncogenic features,
thereby reducing their susceptibility to related inhibitors
(77). Jeong et al (78) constructed organoid models for
different sites of multifocal CRC, performed whole-exome
sequencing, evaluated 25 medications to capture patients'
intertumoral heterogeneity and identified clonal relationships
between multifocal CRCs in terms of biological processes and
therapeutic responses. After constructing primary and metastatic
CRC organoids, transcriptomics and single-cell sequencing analysis
revealed differences in cell composition between primary and
metastatic foci (79). Using CRC
organoids to study tumor heterogeneity can help reveal possible
mechanisms of metastasis of CRC and its potential role in
predicting the chemotherapeutic response and clinical prognosis of
patients (70,80). Furthermore, gene editing techniques
have also been applied to study CRC organoids.
As previously mentioned, in addition to organoids
that may be prepared from patients with CRC, with the development
of gene editing techniques, organoids derived from normal
intestinal epithelial cells with introduced mutations have been
developed to model mutations at all stages of cancer in
vitro. By using CRISPR-Cas technology, Matano et al
(56) knocked out adenomatous
polyposis coli (APC), SMAD4 and TP53 tumor suppressor genes and
knocked in KRAS and phosphatidylinositol-4,5-bisphosphate 3-kinase
in normal epithelial organoids, allowing five mutant organoids to
be cultured in a medium free of ecological factors. In addition,
these organoids were transplanted into the renal capsule and spleen
of nude mice to form tumors. They were found to have different
abilities of tumor formation, invasion and metastasis, but failed
to colonize in the liver (56).
Drost et al (57) used
CRISPR-Cas to target APC, P53, KRAS and SMAD4 in human intestinal
stem cells. They removed one growth factor from the culture medium
to select mutated organoids and then transplanted triple/quadruple
mutated organoids into immunodeficient mice to induce tumor growth.
Both triple- and quadruple-mutant organoids showed high
proliferation rates, while only quadruple-mutant organoids appeared
as solid tumor masses (57).
Fumagalli et al (81)
demonstrated that the continuous accumulation of oncogenic
mutations in the Wnt, EGFR, P53 and TGF-β signalling pathways
promoted effective tumor growth, migration and metastasis using an
in situ transplantation model of human colon organoids with
different combinations of CRC mutations. Roper et al
(64) injected lentivirus into the
intestinal mucosa of mice to generate CRC organoids of APC, P53 and
KRAS and injected these organoids into the colon mucosa of mice to
form an in situ transplantation model. After 12 weeks, 1/3
of the mice developed liver metastases. Human CRC organoids were
then injected into the intestinal mucosa of mice and it was found
that the in situ transplantation of patient-derived CRC
organoids effectively mimicked both primary and metastatic human
CRC (64). The establishment of a
living biobank significantly facilitates the study of CRC
organoids.
To date, researchers have established live organoids
for various tumors, including locally advanced rectal cancer
(82), CRC, liver metastasis of CRC
(80), breast cancer (83), bladder cancer (84), nasopharyngeal cancer (85), pancreatic cancer (86), non-small cell lung cancer (87), gastric cancer (88) and glioblastoma (89). For the first time, van de Wetering
et al (41) successfully
established a living biobank for CRC, including 22 tumor organoids
and 19 normal adjacent organoids from 20 patients. Genomic analysis
has indicated that organoids may display the genomic
characteristics of primary colon cancer and mutation analysis has
demonstrated that the spectrum of genetic changes in organoids was
highly consistent with the large-scale mutation analysis of CRC
(41). Vlachogiannis et al
(90) used 110 fresh biopsy samples
from 71 patients in four prospective phase I/II clinical trials to
construct a living CRC organoid biobank. Histological evaluation
revealed significant morphological similarities between organoids
and patient-derived tissues. Organoid sequencing, along with drug
responses, demonstrated a high degree of similarity in organoid
phenotypes and genotypes to tumors derived from the patient.
Molecular profiling of organoids is matched with the results of
drug screening, which may predict the clinical response of patients
to various medications (90). These
results suggest that, compared with traditional biobanks, organoids
may better simulate the primary tumor and reduce tumor
heterogeneity. It may be used for preclinical high-throughput drug
screening, enabling patients to undergo drug testing without
participation, screening appropriate medications for patients,
forecasting patients' responses to drugs and providing more
possibilities for individualized treatment of tumor patients. In
addition, organoid biobanks can also be used for gene editing and
other research, novel drug development or regenerative medicine
(91).
The advantages of using patient-derived organoids
(PDOs) as preclinical models have been extensively studied over the
last few years, particularly in four settings: i) Drug screening,
ii) drug toxicity testing, iii) drug discovery and iv)
individualized therapy.
How to determine the sensitivity of patients to
chemotherapeutic drugs is critical to patient treatment and it has
been proven that CRC organoids are valuable in individualized drug
sensitivity tests (92). Kong et
al (93) developed
organopharmacogenomic data and a web-based calculation method to
accurately predict drug response in 144 patients with CRC receiving
5-fluorouracil (5-FU), and they found that the expression levels of
components of the ‘BH3-only protein’ pathway correlated with the
sensitivity of CRC organs to 5-FU. Pasch et al (94) verified the response of CRC organoids
to chemotherapy with different concentrations of 5-FU and
oxaliplatin. The results indicated that multiphoton imaging was
used to prospectively predict the response to FOLFOX chemotherapy
in a patient with metastatic CRC receiving FOLFOX (5-FU, leucovorin
and oxaliplatin) chemotherapy, identifying those who did not
respond to standard chemotherapy for CRC. Ooft et al
(95) used 67 colon cancer
organoids to accurately predict the drug response of >80% of
patients receiving irinotecan. However, they failed to predict the
outcome of 5-FU and oxaliplatin, and failed to identify patients
who may benefit from treatment. Chen et al (96) administered different concentrations
of 5-FU and oxaliplatin to CRC organoids to investigate drug
susceptibility and the test data were consistent with clinical
data. Then, through single-cell sequencing technology, it was
speculated that driver genes Stathmin 1, vascular endothelial
growth factor A and N-myc downstream-regulated gene 1 and
transcription factors [E2F transcription factor 1, breast cancer
susceptibility gene 1, MYB proto-oncogene like 2, caudal-type
homeobox 1 (CDX1) and CDX2] were potential oxaliplatin resistance
targets (96). Lv et al
(97) used locally advanced rectal
cancer organoids to observe whether the sensitivity of irinotecan
could predict complete response and survival. The results suggested
that patient-derived locally advanced rectal cancer organoids are
able to predict response to irinotecan-based neoadjuvant
chemoradiotherapy (97).
Drug toxicity is a major cause of failure and
withdrawal of effective potential drug candidates from the
pharmaceutical pipeline (98).
Compared to other preclinical models, patient-derived tumor
organoids better mimic the original tumor tissue and their toxic
response to drugs can help predict drug safety for patients. It
also allows the study of drug effects in mice and the subsequent
generation of normal and tumor organoids to evaluate their
potential systemic toxicity and prioritize their future preclinical
assessment (99,100).
CRC organoids and living organoid biobanks can
provide an accurate summary of the heterogeneous genetic and
morphological composition of cancer cells in the initial tumor, and
their responses to drugs are highly consistent with patients'
clinical responses, providing great hope for the development of
novel drugs (105).
Personalized medicine is an evolving concept in
oncology. The goal is to determine the most appropriate treatment
based on the genetic, transcriptome, environmental and lifestyle
features of each patient (110).
Research has indicated that personalized medicine may achieve
significant improvements in treatment response (111,112). The high consistency between the
organoid model and the original tumor in terms of morphology,
genotype and mutation characteristics, as well as physiological and
pathological transformation, makes it possible to achieve precise
personalized therapy.
In general, the use of organoids to guide the
personalized treatment of CRC has promising applications. The
establishment of the CRC organoid biobank broadens the diagnosis
and treatment vision of CRC, which is helpful in promoting the
implementation of personalized diagnosis and treatment.
Organoids are powerful tools for studying human
development and disease. However, there are still numerous
limitations and challenges.
Single-cell sequencing may provide novel solutions
to the question of whether organoids may serve as a true model of
human biology, whether organoid models need to be optimized and
whether organoids may be used in basic biology (genetic and
pharmacological perturbations) and biomedical research (drug
development and personalized medicine). Studies have developed a
suspension culture technique for cancer organoids. This technique
simplifies organoid cell culture and extends organoid applications
to include routine use in large-scale perturbation screening
(117,118). The efficiency of CRC organoids
generation still needs to be further studied.
The success rate of CRC organoid creation is high;
however, there is a need for improved efficiency, reduced time and
decreased costs associated with organoid generation. One of the
main problems is the limited number of cancer cells contained in
biopsies. The success rate of culture may be further improved by
obtaining multiple biopsies and direct evaluation by pathologists
(119). CRC organoids with
different histological subtypes have been described to be
successfully constructed and clinical parameters, such as tumor
size or presence of ulcers, were not associated with the occurrence
of contamination (55,80,120).
However, the establishment of organoids from tumors with rare CRC
histologic subtypes (e.g., poorly differentiated adenocarcinoma,
mucinous adenocarcinoma and neuroendocrine carcinoma) is more
difficult (33). Furthermore, a
limitation of the application is organoid culture.
The organoid culture of CRC is relatively expensive
and the Wnt3A conditioned medium is not suitable for long-term
storage. As a result of acylation between Wnt and Fzd interactions,
the hydrophobic Wnt protein needs to be purified. Janda et
al (121) developed
water-soluble Wnt agonists to replace Wnt3A and they established an
economical and clear recombinant culture reagent. Traditional
organoid generation methods produce organoids with a closed
sac-like structure and uncontrolled self-organization formation,
which limits the lifespan, morphology, experimental operation and
homeostasis of organoids. The different morphologies of organoids
may lead to different responses to drug therapy. To solve this
problem, researchers use bioengineering technology to control the
shape of organoids (122,123). Most of the Matrigel used for
organoid culture is of animal origin. Matrigel is a basal membrane
extract purified from mouse sarcoma cells by Engelbreth-Holm-Swarm.
However, ECMs of animal origin are characterized by batch-to-batch
variability and lack of tissue-specific ECM components (124–126). To overcome the limitations of ECMs
of animal origin, hydrogels are being developed to replace Matrigel
and BME (127–129). The establishment of living
organoid biobanks still needs to be further studied.
Not all organoid biobanks maintain tumor
heterogeneity in individual patients and the reason why organoids
reproduce tumor heterogeneity in individuals is unknown (130). There are many limitations to the
establishment of living organoid biobanks in CRC. First, the
generation efficiency of the organoid biobank is low, which may be
caused by: i) Pollution of tumor organoids (overgrowth of normal
organoids or tissue sample contamination); ii) optimization of
medium formula; iii) few tumor cells; and iv) quality control
problems. Second, the coculture system with other cell types is not
clearly defined, so it is difficult to reproduce the tumor
microenvironment. Third, standardized procedures for living
organoids have not been defined. Fourth, there are ethical issues,
such as informed consent, commercialization, and the manufacture
and safety of patentable products (131–134). The toxicity of drugs to the liver
is often overlooked.
Researchers commonly utilize normal intestinal
organoids as models for studying drug toxicity. However, it is
essential to recognize and address the potential role of liver
toxicity in chemotherapy, which is often ignored by these studies
(104,135,136). Combining normal liver and
intestinal organoids with CRC organoids from patients in a single
organoid microarray system may provide a better understanding of
drug toxicity (137).
Another important limitation of organoid models is
that they usually contain only tumor epithelium, which cannot fully
reproduce the diversity of cell types in the tumor microenvironment
(including nonvalvular cells, such as immune cells and stroma), and
the heterogeneity of cell types in the tumor microenvironment has
an important impact on the development and treatment of tumors
(138,139). Therefore, numerous studies have
attempted to develop next-generation tumor organoids by
cocultivation with nonepithelial stromal cells, such as
cancer-associated fibroblasts (CAFs), enteric nervous system cells,
endothelial cells and immune cells. However, a holistic view of
these cells in vivo is still lacking, limiting their ability
to fully mimic the primary tumor. It is difficult to determine the
therapeutic efficacy of individual cell populations (140,141).
CAFs are a major part of the tumor microenvironment
and successful implementations of a coculture system of organoids
and fibroblasts for CRC have been reported. Mosa et al
(142) constructed a
WNT-independent CRC organoid-inflammatory CAF coculture model and
found that EMT markers were significantly upregulated, while
cancer-associated myofibroblasts restored this phenotype. The
results suggested that tumor growth and malignancy are modulated
differently by different fibroblast subtypes under the influence of
Wnt signalling (142). Naruse
et al (143) constructed a
coculture system of CRC organoids and CAFs, and performed
sequencing that showed that the coculture system of CRC organoids
and paired CAFs was able to partially replicate the tumor
microenvironment. In addition, Luo et al (128) found that CAFs were able to
maintain the proliferation of CRC organoids cultured in hydrogels
and restore the unique biological pathway that was present not only
in organoid cultures but also in patient tissues.
The interaction of immune cells with tumors in the
tumor microenvironment was examined by coculturing normal or tumor
intestinal epithelial organoids with immune cells. Noel et
al (144) established the
first human macrophage-intestinal organic-coculture system to
elucidate human innate immune processes and cellular communication,
and to examine host-pathogen interactions by accurately
representing what happens in human intestinal epithelial cells.
Neal et al (145) used the
liquid-gas interface method to reconstruct the tumor
microenvironment and successfully retained a variety of immune
cells except for T cells in the organoid culture system.
Single-cell sequencing and immunohistochemical analysis
demonstrated that organoid tumor-infiltrating lymphocytes
accurately retained the original tumor T-cell receptor profile and
successfully mimicked immune checkpoint blockade. The
organoid-based global proliferation of primary tumor epithelial
cells and endogenous immune stroma should enable immuno-oncology
studies in tumor microenvironments (145). Dijkstra et al (146) constructed an organoid-autologous
T-cell coculture system for CRC with mismatch repair defects and
found that the system amplified tumor-reactive T cells, providing a
possibility for the generation of patient-specific T-cell products.
T cells may be used to evaluate the killing efficiency of matched
tumor organoids and to allow the establishment of an in
vitro testing system for T-cell-based immunotherapies at the
individual patient level (147).
Frenkel et al (148)
studied the interaction between lymphatic vessels and CRC by
coculturing lymphatic endothelial cells (LECs) with CRC organoids
and found that LECs formed a permeable vascular structure on the
extracellular matrix, resulting in a longer and more stable
lifespan of the cocultivation model.
Patient-derived CRC organoids are usually closed
sacs and lack the tissue-tissue interface between epithelial tumor
cells and the surrounding vascularis and stroma, which are crucial
for cancer control and progression. It is challenging to provide
organoid nutrients, supply oxygen and remove waste. Therefore,
bioengineered devices or scaffolds are needed to reconstruct
organoids that are more representative of the internal environment
(149).
Organ chips, also referred to as microfluidic cell
culture devices, are developed from microfluidic chips and contain
continuously perfused microcavities, in which live cells are used
to simulate the functional units of human tissues and organs in
vitro. In recent years, organ chip technology has been greatly
developed, and currently, organ chips of different structures may
be constructed, providing a new method for drug screening (150,151). Organoid technology is combined
with chip technology to form organ-on-a-chip technology. Combining
the advantages of these two technologies, organ-on-a-chip may be
used as a more predictive preclinical model that is widely
applicable to drug discovery, personalized medicine and
regenerative medicine (152,153). However, at present, microfluidic
organ chips mostly rely on organoids formed from human pluripotent
stem cells, and there are few studies on organ chips for CRC
(154,155).
Organ chips open up numerous possibilities for the
use of microfluidic devices, such as specifying the shape of
organoids, coculture of CRC organoids with tumor microenvironments
and drug screening (156).
Gjorevski et al (122) used
microfluidics to control the initial shape of intestinal organoids,
helping to control the self-organization of organoids. Frenkel
et al (148) injected
immortalized lymphatic endothelial cells into a microfluidic chip
with an independent extracellular matrix to form a perfectible
vascular structure, and introduced mouse colon cancer organoids
into lymphatic vessels to establish a stable coculture model for
studying lymphatic vessel formation and tumor cell metastasis.
Rajasekar et al (157)
successfully designed a microfluidic platform for
infusion-vascularized colon organoids that may guide organoid
development without the use of physical structures to artificially
define and limit biological structure and remodelling, and this
system is expected to be used for CRC organoids. Pinho et al
(158) found that, compared with
conventional PDO culture, no significant differences were verified
in the organoid response to 5-FU treatment on-chip and on-plate.
However, the size and efficiency of colony formation of the
organoid were significantly increased.
CRC is a worldwide health concern. The lack of good
in vitro models has been a limitation to improving clinical
treatment for a long time. Organoid technology has become a new
strategy to solve this issue and may further deepen the
understanding of the occurrence and development of CRC. Organoid
models, which may be combined with biobanks, CRISPR,
high-throughput screening, chip technology and xenotransplantation,
can make a significant contribution to the further development of
organoids. A promising correlation exists between CRC organoids and
patient responses to cancer treatment.
Although CRC organoids have shown some limitations
and gaps in their application to cancer modelling and personalized
medicine, with the continuous optimization of organoid technology,
CRC organoid models will become an indispensable tool for
preclinical and clinical research.
Not applicable.
This study was supported by the National Natural Science
Foundation of China (grant no. 81703022), Jiangsu Province Key
Youth Talents Project (grant no. QNRC2016262), Gusu Health Talents
Training Project (grant no. GSWS2019078), the guiding project of
Jiangsu Provincial Health Committee (grant no. Z2021077), the
project of Suzhou Medical health science and technology innovation
(grant no. SKY2022029) and the project of Taicang Science and
Technology Bureau (grant no. TC2018JCYL20).
Not applicable.
CY, WWX and RW were involved in the conception,
writing and editing of the manuscript. YH, KY, XS, GHW and XHX
critically contributed to the drafting of sections falling within
their expertise, and reviewed and corrected the manuscript. All
authors have read and approved the final manuscript. Data
authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global Cancer Statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD, Goding Sauer A,
Fedewa SA, Butterly LF, Anderson JC, Cercek A, Smith RA and Jemal
A: Colorectal cancer statistics, 2020. CA Cancer J Clin.
70:145–164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wong MCS, Huang J, Lok V, Wang J, Fung F,
Ding H and Zheng ZJ: Differences in incidence and mortality trends
of colorectal cancer worldwide based on sex, age, and anatomic
location. Clin Gastroenterol Hepatol. 19:955–966.e61. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nakayama M, Wang D, Kok SY, Oshima H and
Oshima M: Genetic alterations and microenvironment that drive
malignant progression of colorectal cancer: Lessons from mouse and
organoid models. J Cancer Prev. 27:1–6. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jeyakumar A, Dissabandara L and Gopalan V:
A critical overview on the biological and molecular features of red
and processed meat in colorectal carcinogenesis. J Gastroenterol.
52:407–418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen H, Zheng X, Zong X, Li Z, Li N, Hur
J, Fritz CD, Chapman W Jr, Nickel KB, Tipping A, et al: Metabolic
syndrome, metabolic comorbid conditions and risk of early-onset
colorectal cancer. Gut. 70:1147–1154. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Roney MSI, Lanagan C, Sheng YH, Lawler K,
Schmidt C, Nguyen NT, Begun J and Kijanka GS: IgM and IgA augmented
autoantibody signatures improve early-stage detection of colorectal
cancer prior to nodal and distant spread. Clin Transl Immunology.
10:e13302021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
DeStefanis RA, Kratz JD, Olson AM, Sunil
A, DeZeeuw AK, Gillette AA, Sha GC, Johnson KA, Pasch CA, Clipson
L, et al: Impact of baseline culture conditions of cancer organoids
when determining therapeutic response and tumor heterogeneity. Sci
Rep. 12:52052022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ben-David U, Ha G, Tseng YY, Greenwald NF,
Oh C, Shih J, McFarland JM, Wong B, Boehm JS, Beroukhim R and Golub
TR: Patient-derived xenografts undergo mouse-specific tumor
evolution. Nat Genet. 49:1567–1575. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu H, Jiao Y, Qin S, Zhao W, Chu Q and Wu
K: Organoid technology in disease modelling, drug development,
personalized treatment and regeneration medicine. Exp Hematol
Oncol. 7:302018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li M and Izpisua Belmonte JC:
Organoids-Preclinical models of human disease. N Engl J Med.
380:569–579. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng D, Gleyzer R, Tai WH, Kumar P, Bian
Q, Isaacs B, da Rocha EL, Cai S, DiNapoli K, Huang FW and Cahan P:
Evaluating the transcriptional fidelity of cancer models. Genome
Med. 13:732021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Janakiraman H, Zhu Y, Becker SA, Wang C,
Cross A, Curl E, Lewin D, Hoffman BJ, Warren GW, Hill EG, et al:
Modeling rectal cancer to advance neoadjuvant precision therapy.
Int J Cancer. 147:1405–1418. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Veninga V and Voest EE: Tumor organoids:
Opportunities and challenges to guide precision medicine. Cancer
Cell. 39:1190–1201. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fontoura JC, Viezzer C, Dos Santos FG,
Ligabue RA, Weinlich R, Puga RD, Antonow D, Severino P and Bonorino
C: Comparison of 2D and 3D cell culture models for cell growth,
gene expression and drug resistance. Mater Sci Eng C Mater Biol
Appl. 107:1102642020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao H, Yan C, Hu Y, Mu L, Huang K, Li Q,
Li X, Tao D and Qin J: Sphere-forming assay vs. organoid culture:
Determining long-term stemness and the chemoresistant capacity of
primary colorectal cancer cells. Int J Oncol. 54:893–904.
2019.PubMed/NCBI
|
|
17
|
Lehmann R, Lee CM, Shugart EC, Benedetti
M, Charo RA, Gartner Z, Hogan B, Knoblich J, Nelson CM and Wilson
KM: Human organoids: A new dimension in cell biology. Mol Biol
Cell. 30:1129–1137. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Joshi R, Castro De Moura M, Piñeyro D,
Alvarez-Errico D, Arribas C and Esteller M: The DNA methylation
landscape of human cancer organoids available at the American type
culture collection. Epigenetics. 15:1167–1177. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lancaster MA and Knoblich JA:
Organogenesis in a dish: Modeling development and disease using
organoid technologies. Science. 345:12471252014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shirure VS, Hughes CCW and George SC:
Engineering Vascularized Organoid-on-a-Chip Models. Annu Rev Biomed
Eng. 23:141–167. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lancaster MA and Knoblich JA: Generation
of cerebral organoids from human pluripotent stem cells. Nat
Protoc. 9:2329–2340. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sugimoto S and Sato T: Establishment of 3D
intestinal organoid cultures from intestinal stem cells. Methods
Mol Biol. 1612:97–105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saito Y, Onishi N, Takami H, Seishima R,
Inoue H, Hirata Y, Kameyama K, Tsuchihashi K, Sugihara E, Uchino S,
et al: Development of a functional thyroid model based on an
organoid culture system. Biochem Biophys Res Commun. 497:783–789.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mazzucchelli S, Piccotti F, Allevi R,
Truffi M, Sorrentino L, Russo L, Agozzino M, Signati L, Bonizzi A,
Villani L and Corsi F: Establishment and morphological
characterization of patient-derived organoids from breast cancer.
Biol Proced Online. 21:122019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Seidlitz T, Merker SR, Rothe A, Zakrzewski
F, von Neubeck C, Grützmann K, Sommer U, Schweitzer C, Schölch S,
Uhlemann H, et al: Human gastric cancer modelling using organoids.
Gut. 68:207–217. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Broutier L, Andersson-Rolf A, Hindley CJ,
Boj SF, Clevers H, Koo BK and Huch M: Culture and establishment of
self-renewing human and mouse adult liver and pancreas 3D organoids
and their genetic manipulation. Nat Protoc. 11:1724–1743. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kopper O, de Witte CJ, Lõhmussaar K,
Valle-Inclan JE, Hami N, Kester L, Balgobind AV, Korving J, Proost
N, Begthel H, et al: An organoid platform for ovarian cancer
captures intra- and interpatient heterogeneity. Nat Med.
25:838–849. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Takasato M, Er PX, Chiu HS, Maier B,
Baillie GJ, Ferguson C, Parton RG, Wolvetang EJ, Roost MS, Chuva de
Sousa Lopes SM and Little MH: Kidney organoids from human iPS cells
contain multiple lineages and model human nephrogenesis. Nature.
526:564–568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X, Francies HE, Secrier M, Perner J,
Miremadi A, Galeano-Dalmau N, Barendt WJ, Letchford L, Leyden GM,
Goffin EK, et al: Organoid cultures recapitulate esophageal
adenocarcinoma heterogeneity providing a model for clonality
studies and precision therapeutics. Nat Commun. 9:29832018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Christin JR and Shen MM: Modeling tumor
plasticity in organoid models of human cancer. Trends Cancer.
8:161–163. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kakni P, Hueber R, Knoops K,
López-Iglesias C, Truckenmüller R, Habibovic P and Giselbrecht S:
Intestinal organoid culture in polymer film-based microwell arrays.
Adv Biosyst. 4:e20001262020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stevens CE and Leblond CP: Rate of renewal
of the cells of the intestinal epithelium in the rat. Anat Rec.
97:3731947.PubMed/NCBI
|
|
33
|
Li X, Larsson P, Ljuslinder I, Öhlund D,
Myte R, Löfgren-Burström A, Zingmark C, Ling A, Edin S and
Palmqvist R: Ex vivo organoid cultures reveal the importance of the
tumor microenvironment for maintenance of colorectal cancer stem
cells. Cancers (Basel). 12:9232020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Barker N, van Es JH, Kuipers J, Kujala P,
van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H,
Peters PJ and Clevers H: Identification of stem cells in small
intestine and colon by marker gene Lgr5. Nature. 449:1003–1007.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sato T, Vries RG, Snippert HJ, van de
Wetering M, Barker N, Stange DE, van Es JH, Abo A, Kujala P, Peters
PJ and Clevers H: Single Lgr5 stem cells build crypt-villus
structures in vitro without a mesenchymal niche. Nature.
459:262–265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jung P, Sato T, Merlos-Suárez A, Barriga
FM, Iglesias M, Rossell D, Auer H, Gallardo M, Blasco MA, Sancho E,
et al: Isolation and in vitro expansion of human colonic stem
cells. Nat Med. 17:1225–1227. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sato T, Stange DE, Ferrante M, Vries RG,
Van Es JH, Van den Brink S, Van Houdt WJ, Pronk A, Van Gorp J,
Siersema PD and Clevers H: Long-term expansion of epithelial
organoids from human colon, adenoma, adenocarcinoma, and Barrett's
epithelium. Gastroenterology. 141:1762–1772. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Grabinger T, Luks L, Kostadinova F,
Zimberlin C, Medema JP, Leist M and Brunner T: Ex vivo culture of
intestinal crypt organoids as a model system for assessing cell
death induction in intestinal epithelial cells and enteropathy.
Cell Death Dis. 5:e12282014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ganesh K, Wu C, O'Rourke KP, Szeglin BC,
Zheng Y, Sauvé CG, Adileh M, Wasserman I, Marco MR, Kim AS, et al:
A rectal cancer organoid platform to study individual responses to
chemoradiation. Nat Med. 25:1607–1614. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fujii M, Matano M, Nanki K and Sato T:
Efficient genetic engineering of human intestinal organoids using
electroporation. Nat Protoc. 10:1474–1485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
van de Wetering M, Francies HE, Francis
JM, Bounova G, Iorio F, Pronk A, van Houdt W, van Gorp J,
Taylor-Weiner A, Kester L, et al: Prospective derivation of a
living organoid biobank of colorectal cancer patients. Cell.
161:933–945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xie BY and Wu AW: Organoid culture of
isolated cells from patient-derived tissues with colorectal cancer.
Chin Med J (Engl). 129:2469–2475. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huch M, Dorrell C, Boj SF, van Es JH, Li
VS, van de Wetering M, Sato T, Hamer K, Sasaki N, Finegold MJ, et
al: In vitro expansion of single Lgr5+ liver stem cells induced by
Wnt-driven regeneration. Nature. 494:247–250. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huch M, Bonfanti P, Boj SF, Sato T,
Loomans CJ, van de Wetering M, Sojoodi M, Li VS, Schuijers J,
Gracanin A, et al: Unlimited in vitro expansion of adult bi-potent
pancreas progenitors through the Lgr5/R-spondin axis. EMBO J.
32:2708–2721. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Amsterdam A, Raanan C, Schreiber L,
Freyhan O, Schechtman L and Givol D: Localization of the stem cell
markers LGR5 and Nanog in the normal and the cancerous human ovary
and their inter-relationship. Acta Histochem. 115:330–338. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Koo BK, Stange DE, Sato T, Karthaus W,
Farin HF, Huch M, van Es JH and Clevers H: Controlled gene
expression in primary Lgr5 organoid cultures. Nat Methods. 9:81–83.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Barker N, Rookmaaker MB, Kujala P, Ng A,
Leushacke M, Snippert H, van de Wetering M, Tan S, Van Es JH, Huch
M, et al: Lgr5(+ve) stem/progenitor cells contribute to nephron
formation during kidney development. Cell Rep. 2:540–552. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kadur Lakshminarasimha Murthy P, Sontake
V, Tata A, Kobayashi Y, Macadlo L, Okuda K, Conchola AS, Nakano S,
Gregory S, Miller LA, et al: Human distal lung maps and lineage
hierarchies reveal a bipotent progenitor. Nature. 604:111–119.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Leung C, Tan SH and Barker N: Recent
advances in Lgr5+ stem cell research. Trends Cell Biol.
28:380–391. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Schutgens F and Clevers H: Human
organoids: Tools for understanding biology and treating diseases.
Annu Rev Pathol. 15:211–234. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Crespo M, Vilar E, Tsai SY, Chang K, Amin
S, Srinivasan T, Zhang T, Pipalia NH, Chen HJ, Witherspoon M, et
al: Colonic organoids derived from human induced pluripotent stem
cells for modeling colorectal cancer and drug testing. Nat Med.
23:878–884. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Daoud A and Múnera JO: Generation of human
colonic organoids from human pluripotent stem cells. Methods Cell
Biol. 159:201–227. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Spence JR, Mayhew CN, Rankin SA, Kuhar MF,
Vallance JE, Tolle K, Hoskins EE, Kalinichenko VV, Wells SI, Zorn
AM, et al: Directed differentiation of human pluripotent stem cells
into intestinal tissue in vitro. Nature. 470:105–109. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Múnera JO, Sundaram N, Rankin SA, Hill D,
Watson C, Mahe M, Vallance JE, Shroyer NF, Sinagoga KL,
Zarzoso-Lacoste A, et al: Differentiation of human pluripotent stem
cells into colonic organoids via transient activation of BMP
signaling. Cell Stem Cell. 21:51–64.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Fujii M, Shimokawa M, Date S, Takano A,
Matano M, Nanki K, Ohta Y, Toshimitsu K, Nakazato Y, Kawasaki K, et
al: A colorectal tumor organoid library demonstrates progressive
loss of niche factor requirements during tumorigenesis. Cell Stem
Cell. 18:827–838. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Matano M, Date S, Shimokawa M, Takano A,
Fujii M, Ohta Y, Watanabe T, Kanai T and Sato T: Modeling
colorectal cancer using CRISPR-Cas9-mediated engineering of human
intestinal organoids. Nat Med. 21:256–262. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Drost J, van Jaarsveld RH, Ponsioen B,
Zimberlin C, van Boxtel R, Buijs A, Sachs N, Overmeer RM, Offerhaus
GJ, Begthel H, et al: Sequential cancer mutations in cultured human
intestinal stem cells. Nature. 521:43–47. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yan HHN, Siu HC, Ho SL, Yue SSK, Gao Y,
Tsui WY, Chan D, Chan AS, Wong JWH, Man AHY, et al: Organoid
cultures of early-onset colorectal cancers reveal distinct and rare
genetic profiles. Gut. 69:2165–2179. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Weeber F, van de Wetering M, Hoogstraat M,
Dijkstra KK, Krijgsman O, Kuilman T, Gadellaa-van Hooijdonk CG, van
der Velden DL, Peeper DS, Cuppen EP, et al: Preserved genetic
diversity in organoids cultured from biopsies of human colorectal
cancer metastases. Proc Natl Acad Sci USA. 112:13308–13311. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang R, Mao Y, Wang W, Zhou X, Wang W, Gao
S, Li J, Wen L, Fu W and Tang F: Systematic evaluation of
colorectal cancer organoid system by single-cell RNA-Seq analysis.
Genome Biol. 23:1062022. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cristobal A, van den Toorn HWP, van de
Wetering M, Clevers H, Heck AJR and Mohammed S: Personalized
proteome profiles of healthy and tumor human colon organoids reveal
both individual diversity and basic features of colorectal cancer.
Cell Rep. 18:263–274. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Jabaji Z, Sears CM, Brinkley GJ, Lei NY,
Joshi VS, Wang J, Lewis M, Stelzner M, Martín MG and Dunn JC: Use
of collagen gel as an alternative extracellular matrix for the in
vitro and in vivo growth of murine small intestinal epithelium.
Tissue Eng Part C Methods. 19:961–969. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Brown JW and Mills JC: Implantable
synthetic organoid matrices for intestinal regeneration. Nat Cell
Biol. 19:1307–1308. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Roper J, Tammela T, Cetinbas NM, Akkad A,
Roghanian A, Rickelt S, Almeqdadi M, Wu K, Oberli MA,
Sánchez-Rivera FJ, et al: In vivo genome editing and organoid
transplantation models of colorectal cancer and metastasis. Nat
Biotechnol. 35:569–576. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Barbáchano A, Fernández-Barral A,
Bustamante-Madrid P, Prieto I, Rodríguez-Salas N, Larriba MJ and
Muñoz A: Organoids and colorectal cancer. Cancers (Basel).
13:26572021. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kapoor-Narula U and Lenka N: Cancer stem
cells and tumor heterogeneity: Deciphering the role in tumor
progression and metastasis. Cytokine. 157:1559682022. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zeuner A, Todaro M, Stassi G and De Maria
R: Colorectal cancer stem cells: From the crypt to the clinic. Cell
Stem Cell. 15:692–705. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Punt CJA, Koopman M and Vermeulen L: From
tumour heterogeneity to advances in precision treatment of
colorectal cancer. Nat Rev Clin Oncol. 14:235–246. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Pleguezuelos-Manzano C, Puschhof J,
Rosendahl Huber A, van Hoeck A, Wood HM, Nomburg J, Gurjao C,
Manders F, Dalmasso G, Stege PB, et al: Mutational signature in
colorectal cancer caused by genotoxic pks+ E. coli.
Nature. 580:269–273. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang R, Li J, Zhou X, Mao Y, Wang W, Gao
S, Wang W, Gao Y, Chen K, Yu S, et al: Single-cell genomic and
transcriptomic landscapes of primary and metastatic colorectal
cancer tumors. Genome Med. 14:932022. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Youk J, Kwon HW, Kim R and Ju YS:
Dissecting single-cell genomes through the clonal organoid
technique. Exp Mol Med. 53:1503–1511. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Roerink SF, Sasaki N, Lee-Six H, Young MD,
Alexandrov LB, Behjati S, Mitchell TJ, Grossmann S, Lightfoot H,
Egan DA, et al: Intra-tumour diversification in colorectal cancer
at the single-cell level. Nature. 556:457–462. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ono H, Arai Y, Furukawa E, Narushima D,
Matsuura T, Nakamura H, Shiokawa D, Nagai M, Imai T, Mimori K, et
al: Single-cell DNA and RNA sequencing reveals the dynamics of
intra-tumor heterogeneity in a colorectal cancer model. BMC Biol.
19:2072021. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Demmers LC, Kretzschmar K, Van Hoeck A,
Bar-Epraïm YE, van den Toorn HWP, Koomen M, van Son G, van Gorp J,
Pronk A, Smakman N, et al: Single-cell derived tumor organoids
display diversity in HLA class I peptide presentation. Nat Commun.
11:53382020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Greaves M: Evolutionary determinants of
cancer. Cancer Discov. 5:806–820. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
McGranahan N and Swanton C: Clonal
heterogeneity and tumor evolution: Past, present, and the future.
Cell. 168:613–628. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kim SC, Park JW, Seo HY, Kim M, Park JH,
Kim GH, Lee JO, Shin YK, Bae JM, Koo BK, et al: Multifocal organoid
capturing of colon cancer reveals pervasive intratumoral
heterogenous drug responses. Adv Sci (Weinh). 9:e21033602022.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Jeong N, Kim SC, Park JW, Park SG, Nam KH,
Lee JO, Shin YK, Bae JM, Jeong SY, Kim MJ and Ku JL: Multifocal
organoids reveal clonal associations between synchronous intestinal
tumors with pervasive heterogeneous drug responses. NPJ Genom Med.
7:422022. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Okamoto T, duVerle D, Yaginuma K, Natsume
Y, Yamanaka H, Kusama D, Fukuda M, Yamamoto M, Perraudeau F,
Srivastava U, et al: Comparative analysis of patient-matched PDOs
revealed a reduction in OLFM4-Associated clusters in metastatic
lesions in colorectal cancer. Stem Cell Reports. 16:954–967. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Mo S, Tang P, Luo W, Zhang L, Li Y, Hu X,
Ma X, Chen Y, Bao Y, He X, et al: Patient-Derived organoids from
colorectal cancer with paired liver metastasis reveal tumor
heterogeneity and predict response to chemotherapy. Adv Sci
(Weinh). 9:e22040972022. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Fumagalli A, Drost J, Suijkerbuijk SJE,
van Boxtel R, de Ligt J, Offerhaus GJ, Begthel H, Beerling E, Tan
EH, Sansom OJ, et al: Genetic dissection of colorectal cancer
progression by orthotopic transplantation of engineered cancer
organoids. Proc Natl Acad Sci USA. 114:E2357–E2364. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Yao Y, Xu X, Yang L, Zhu J, Wan J, Shen L,
Xia F, Fu G, Deng Y, Pan M, et al: Patient-Derived organoids
predict chemoradiation responses of locally advanced rectal cancer.
Cell Stem Cell. 26:17–26.e6. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Sachs N, de Ligt J, Kopper O, Gogola E,
Bounova G, Weeber F, Balgobind AV, Wind K, Gracanin A, Begthel H,
et al: A living biobank of breast cancer organoids captures disease
heterogeneity. Cell. 172:373–386.e10. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Mullenders J, de Jongh E, Brousali A,
Roosen M, Blom JPA, Begthel H, Korving J, Jonges T, Kranenburg O,
Meijer R and Clevers HC: Mouse and human urothelial cancer
organoids: A tool for bladder cancer research. Proc Natl Acad Sci
USA. 116:4567–4574. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang XW, Xia TL, Tang HC, Liu X, Han R,
Zou X, Zhao YT, Chen MY and Li G: Establishment of a
patient-derived organoid model and living biobank for
nasopharyngeal carcinoma. Ann Transl Med. 10:5262022. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Beato F, Reverón D, Dezsi KB, Ortiz A,
Johnson JO, Chen DT, Ali K, Yoder SJ, Jeong D, Malafa M, et al:
Establishing a living biobank of patient-derived organoids of
intraductal papillary mucinous neoplasms of the pancreas. Lab
Invest. 101:204–217. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Li YF, Gao Y, Liang BW, Cao XQ, Sun ZJ, Yu
JH, Liu ZD and Han Y: Patient-derived organoids of non-small cells
lung cancer and their application for drug screening. Neoplasma.
67:430–437. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Yan HHN, Siu HC, Law S, Ho SL, Yue SSK,
Tsui WY, Chan D, Chan AS, Ma S, Lam KO, et al: A Comprehensive
human gastric cancer organoid biobank captures tumor subtype
heterogeneity and enables therapeutic screening. Cell Stem Cell.
23:882–897.e11. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Jacob F, Salinas RD, Zhang DY, Nguyen PTT,
Schnoll JG, Wong SZH, Thokala R, Sheikh S, Saxena D, Prokop S, et
al: A patient-derived glioblastoma organoid model and biobank
recapitulates inter- and intra-tumoral heterogeneity. Cell.
180:188–204.e22. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Vlachogiannis G, Hedayat S, Vatsiou A,
Jamin Y, Fernández-Mateos J, Khan K, Lampis A, Eason K, Huntingford
I, Burke R, et al: Patient-derived organoids model treatment
response of metastatic gastrointestinal cancers. Science.
359:920–926. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Luo L, Ma Y, Zheng Y, Su J and Huang G:
Application progress of organoids in colorectal cancer. Front Cell
Dev Biol. 10:8150672022. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Seidlitz T and Stange DE: Gastrointestinal
cancer organoids-applications in basic and translational cancer
research. Exp Mol Med. 53:1459–1470. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Kong J, Lee H, Kim D, Han SK, Ha D, Shin K
and Kim S: Network-based machine learning in colorectal and bladder
organoid models predicts anti-cancer drug efficacy in patients. Nat
Commun. 11:54852020. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Pasch CA, Favreau PF, Yueh AE, Babiarz CP,
Gillette AA, Sharick JT, Karim MR, Nickel KP, DeZeeuw AK,
Sprackling CM, et al: Patient-Derived cancer organoid cultures to
predict sensitivity to chemotherapy and radiation. Clin Cancer Res.
25:5376–5387. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Ooft SN, Weeber F, Dijkstra KK, McLean CM,
Kaing S, van Werkhoven E, Schipper L, Hoes L, Vis DJ, van de Haar
J, et al: Patient-derived organoids can predict response to
chemotherapy in metastatic colorectal cancer patients. Sci Transl
Med. 11:eaay25742019. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Chen G, Gong T, Wang Z, Wang Z, Lin X,
Chen S, Sun C, Zhao W, Kong Y, Ai H, et al: Colorectal cancer
organoid models uncover oxaliplatin-resistant mechanisms at single
cell resolution. Cell Oncol (Dordr). 45:1155–1167. 2022.PubMed/NCBI
|
|
97
|
Lv T, Shen L, Xu X, Yao Y, Mu P, Zhang H,
Wan J, Wang Y, Guan R, Li X, et al: Patient-derived tumor organoids
predict responses to irinotecan-based neoadjuvant chemoradiotherapy
in patients with locally advanced rectal cancer. Int J Cancer.
152:524–535. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hongmao S: A Practical Guide to Rational
Drug Design. Woodhead Publishing; 2015
|
|
99
|
Pauli C, Hopkins BD, Prandi D, Shaw R,
Fedrizzi T, Sboner A, Sailer V, Augello M, Puca L, Rosati R, et al:
Personalized in vitro and in vivo cancer models to guide precision
medicine. Cancer Discov. 7:462–477. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Fielden MR and Kolaja KL: The role of
early in vivo toxicity testing in drug discovery toxicology. Expert
Opin Drug Saf. 7:107–110. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Lu W, Rettenmeier E, Paszek M, Yueh MF,
Tukey RH, Trottier J, Barbier O and Chen S: Crypt organoid culture
as an in vitro model in drug metabolism and cytotoxicity studies.
Drug Metab Dispos. 45:748–754. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Schnalzger TE, de Groot MH, Zhang C, Mosa
MH, Michels BE, Röder J, Darvishi T, Wels WS and Farin HF: 3D model
for CAR-mediated cytotoxicity using patient-derived colorectal
cancer organoids. EMBO J. 38:e1009282019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Park M, Kwon J, Shin HJ, Moon SM, Kim SB,
Shin US, Han YH and Kim Y: Butyrate enhances the efficacy of
radiotherapy via FOXO3A in colorectal cancer patient-derived
organoids. Int J Oncol. 57:1307–1318. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
De Oliveira T, Goldhardt T, Edelmann M,
Rogge T, Rauch K, Kyuchukov ND, Menck K, Bleckmann A, Kalucka J,
Khan S, et al: Effects of the Novel PFKFB3 Inhibitor KAN0438757 on
colorectal cancer cells and its systemic toxicity evaluation in
vivo. Cancers (Basel). 13:10112021. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Weeber F, Ooft SN, Dijkstra KK and Voest
EE: Tumor organoids as a pre-clinical cancer model for drug
discovery. Cell Chem Biol. 24:1092–1100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Rae C, Amato F and Braconi C:
Patient-Derived organoids as a model for cancer drug discovery. Int
J Mol Sci. 22:34832021. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Costales-Carrera A, Fernández-Barral A,
Bustamante-Madrid P, Guerra L, Cantero R, Barbáchano A and Muñoz A:
Plocabulin displays strong cytotoxic activity in a personalized
colon cancer patient-derived 3D Organoid Assay. Mar Drugs.
17:6482019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Zerp SF, Bibi Z, Verbrugge I, Voest EE and
Verheij M: Enhancing radiation response by a second-generation
TRAIL receptor agonist using a new in vitro organoid model system.
Clin Transl Radiat Oncol. 24:1–9. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Norkin M, Ordóñez-Morán P and Huelsken J:
High-content, targeted RNA-seq screening in organoids for drug
discovery in colorectal cancer. Cell Rep. 35:1090262021. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Sailer V, Pauli C, Merzier EC, Mosquera
JM, Beltran H, Rubin MA and Rao RA: On-site cytology for
development of patient-derived three-dimensional organoid
cultures-A pilot study. Anticancer Res. 37:1569–1573. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Sholl LM, Aisner DL, Varella-Garcia M,
Berry LD, Dias-Santagata D, Wistuba II, Chen H, Fujimoto J, Kugler
K, Franklin WA, et al: Multi-institutional oncogenic driver
mutation analysis in lung adenocarcinoma: the lung cancer mutation
consortium experience. J Thorac Oncol. 10:768–777. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Le DT, Uram JN, Wang H, Bartlett BR,
Kemberling H, Eyring AD, Skora AD, Luber BS, Azad NS, Laheru D, et
al: PD-1 blockade in tumors with mismatch-repair deficiency. N Engl
J Med. 372:2509–2520. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Hsu KS, Adileh M, Martin ML, Makarov V,
Chen J, Wu C, Bodo S, Klingler S, Sauvé CG, Szeglin BC, et al:
Colorectal cancer develops inherent radiosensitivity that can be
predicted using patient-derived organoids. Cancer Res.
82:2298–2312. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Cho YW, Min DW, Kim HP, An Y, Kim S, Youk
J, Chun J, Im JP, Song SH, Ju YS, et al: Patient-derived organoids
as a preclinical platform for precision medicine in colorectal
cancer. Mol Oncol. 16:2396–2412. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Geevimaan K, Guo JY, Shen CN, Jiang JK,
Fann CSJ, Hwang MJ, Shui JW, Lin HT, Wang MJ, Shih HC, et al:
Patient-Derived organoid serves as a platform for personalized
chemotherapy in advanced colorectal cancer patients. Front Oncol.
12:8834372022. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Wang T, Pan W, Zheng H, Zheng H, Wang Z,
Li JJ, Deng C and Yan J: Accuracy of using a patient-derived tumor
organoid culture model to predict the response to chemotherapy
regimens in stage IV colorectal cancer: A blinded study. Dis Colon
Rectum. 64:833–850. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Bock C, Boutros M, Camp JG, Clarke L,
Clevers H, Knoblich JA, Liberali P, Regev A, Rios AC, Stegle O, et
al: The Organoid Cell Atlas. Nat Biotechnol. 39:13–17. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Price S, Bhosle S, Gonçalves E, Li X,
McClurg DP, Barthorpe S, Beck A, Hall C, Lightfoot H, Farrow L, et
al: A suspension technique for efficient large-scale cancer
organoid culturing and perturbation screens. Sci Rep. 12:55712022.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Ji DB and Wu AW: Organoid in colorectal
cancer: Progress and challenges. Chin Med J (Engl). 133:1971–1977.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Marinucci M, Ercan C, Taha-Mehlitz S,
Fourie L, Panebianco F, Bianco G, Gallon J, Staubli S, Soysal SD,
Zettl A, et al: Standardizing patient-derived organoid generation
workflow to avoid microbial contamination from colorectal cancer
tissues. Front Oncol. 11:7818332022. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Janda CY, Dang LT, You C, Chang J, de Lau
W, Zhong ZA, Yan KS, Marecic O, Siepe D, Li X, et al: Surrogate Wnt
agonists that phenocopy canonical Wnt and β-catenin signalling.
Nature. 545:234–237. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Gjorevski N, Nikolaev M, Brown TE,
Mitrofanova O, Brandenberg N, DelRio FW, Yavitt FM, Liberali P,
Anseth KS and Lutolf MP: Tissue geometry drives deterministic
organoid patterning. Science. 375:eaaw90212022. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Nikolaev M, Mitrofanova O, Broguiere N,
Geraldo S, Dutta D, Tabata Y, Elci B, Brandenberg N, Kolotuev I,
Gjorevski N, et al: Homeostatic mini-intestines through
scaffold-guided organoid morphogenesis. Nature. 585:574–578. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Kleinman HK and Martin GR: Matrigel:
Basement membrane matrix with biological activity. Semin Cancer
Biol. 15:378–386. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Heo JH, Kang D, Seo SJ and Jin Y:
Engineering the extracellular matrix for organoid culture. Int J
Stem Cells. 15:60–69. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Rathje F, Klingler S and Aberger F:
Organoids for Modeling (Colorectal) Cancer in a Dish. Cancers
(Basel). 14:54162022. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Ng S, Tan WJ, Pek MMX, Tan MH and Kurisawa
M: Mechanically and chemically defined hydrogel matrices for
patient-derived colorectal tumor organoid culture. Biomaterials.
219:1194002019. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Luo X, Fong ELS, Zhu C, Lin QXX, Xiong M,
Li A, Li T, Benoukraf T, Yu H and Liu S: Hydrogel-based colorectal
cancer organoid co-culture models. Acta Biomater. 132:461–472.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Tayler IM and Stowers RS: Engineering
hydrogels for personalized disease modeling and regenerative
medicine. Acta Biomater. 132:4–22. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Yang H, Zhang N and Liu YC: An organoids
biobank for recapitulating tumor heterogeneity and personalized
medicine. Chin J Cancer Res. 32:408–413. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Boers SN, van Delden JJM and Bredenoord
AL: Organoids as hybrids: Ethical implications for the exchange of
human tissues. J Med Ethics. 45:131–139. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Botti G, Di Bonito M and Cantile M:
Organoid biobanks as a new tool for pre-clinical validation of
candidate drug efficacy and safety. Int J Physiol Pathophysiol
Pharmacol. 13:17–21. 2021.PubMed/NCBI
|
|
133
|
Wallaschek N, Niklas C, Pompaiah M,
Wiegering A, Germer CT, Kircher S, Brändlein S, Maurus K, Rosenwald
A, Yan HHN, et al: Establishing pure cancer organoid cultures:
Identification, selection and verification of cancer phenotypes and
genotypes. J Mol Biol. 431:2884–2893. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Driehuis E, Kretzschmar K and Clevers H:
Establishment of patient-derived cancer organoids for
drug-screening applications. Nat Protoc. 15:3380–3409. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Grigorian A and O'Brien CB: Hepatotoxicity
secondary to chemotherapy. J Clin Transl Hepatol. 2:95–102.
2014.PubMed/NCBI
|
|
136
|
Fiore D, Ramesh P, Proto MC, Piscopo C,
Franceschelli S, Anzelmo S, Medema JP, Bifulco M and Gazzerro P:
Rimonabant kills colon cancer stem cells without inducing toxicity
in normal colon organoids. Front Pharmacol. 8:9492018. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Idris M, Alves MM, Hofstra RMW, Mahe MM
and Melotte V: Intestinal multicellular organoids to study
colorectal cancer. Biochim Biophys Acta Rev Cancer.
1876:1885862021. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Zhang J, Tavakoli H, Ma L and Li X, Han L
and Li X: Immunotherapy discovery on tumor organoid-on-a-chip
platforms that recapitulate the tumor microenvironment. Adv Drug
Deliv Rev. 187:1143652022. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Junttila MR and de Sauvage FJ: Influence
of tumour micro-environment heterogeneity on therapeutic response.
Nature. 501:346–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
LeSavage BL, Suhar RA, Broguiere N, Lutolf
MP and Heilshorn SC: Next-generation cancer organoids. Nat Mater.
21:143–159. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Bulin AL and Hasan T: Spatiotemporal
tracking of different cell populations in cancer organoid models
for investigations on photodynamic therapy. Methods Mol Biol.
2451:81–90. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Mosa MH, Michels BE, Menche C, Nicolas AM,
Darvishi T, Greten FR and Farin HF: A Wnt-Induced phenotypic switch
in cancer-associated fibroblasts inhibits EMT in colorectal cancer.
Cancer Res. 80:5569–5582. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Naruse M, Ochiai M, Sekine S, Taniguchi H,
Yoshida T, Ichikawa H, Sakamoto H, Kubo T, Matsumoto K, Ochiai A
and Imai T: Re-expression of REG family and DUOXs genes in CRC
organoids by co-culturing with CAFs. Sci Rep. 11:20772021.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Noel G, Baetz NW, Staab JF, Donowitz M,
Kovbasnjuk O, Pasetti MF and Zachos NC: A primary human
macrophage-enteroid co-culture model to investigate mucosal gut
physiology and host-pathogen interactions. Sci Rep. 7:452702017.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Neal JT, Li X, Zhu J, Giangarra V,
Grzeskowiak CL, Ju J, Liu IH, Chiou SH, Salahudeen AA, Smith AR, et
al: Organoid modeling of the tumor immune microenvironment. Cell.
175:1972–1988.e16. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Dijkstra KK, Cattaneo CM, Weeber F,
Chalabi M, van de Haar J, Fanchi LF, Slagter M, van der Velden DL,
Kaing S, Kelderman S, et al: Generation of tumor-reactive T cells
by co-culture of peripheral blood lymphocytes and tumor organoids.
Cell. 174:1586–1598.e12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Cattaneo CM, Dijkstra KK, Fanchi LF,
Kelderman S, Kaing S, van Rooij N, van den Brink S, Schumacher TN
and Voest EE: Tumor organoid-T-cell coculture systems. Nat Protoc.
15:15–39. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Frenkel N, Poghosyan S, Alarcón CR, García
SB, Queiroz K, van den Bent L, Laoukili J, Rinkes IB, Vulto P,
Kranenburg O and Hagendoorn J: Long-Lived human lymphatic
endothelial cells to study lymphatic biology and lymphatic
vessel/tumor coculture in a 3D microfluidic model. ACS Biomater Sci
Eng. 7:3030–3042. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Sontheimer-Phelps A, Hassell BA and Ingber
DE: Modelling cancer in microfluidic human organs-on-chips. Nat Rev
Cancer. 19:65–81. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Bein A, Shin W, Jalili-Firoozinezhad S,
Park MH, Sontheimer-Phelps A, Tovaglieri A, Chalkiadaki A, Kim HJ
and Ingber DE: Microfluidic Organ-on-a-Chip models of human
intestine. Cell Mol Gastroenterol Hepatol. 5:659–668. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Yan J, Li Z and Guo J, Liu S and Guo J:
Organ-on-a-chip: A new tool for in vitro research. Biosens
Bioelectron. 216:1146262022. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Park SE, Georgescu A and Huh D:
Organoids-on-a-chip. Science. 364:960–965. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Takebe T, Zhang B and Radisic M:
Synergistic engineering: Organoids Meet Organs-on-a-Chip. Cell Stem
Cell. 21:297–300. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Karzbrun E, Kshirsagar A, Cohen SR, Hanna
JH and Reiner O: Human brain organoids on a chip reveal the physics
of folding. Nat Phys. 14:515–522. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Zhang YS, Aleman J, Shin SR, Kilic T, Kim
D, Mousavi Shaegh SA, Massa S, Riahi R, Chae S, Hu N, et al:
Multisensor-integrated organs-on-chips platform for automated and
continual in situ monitoring of organoid behaviors. Proc Natl Acad
Sci USA. 114:E2293–E2302. 2017.PubMed/NCBI
|
|
156
|
Skardal A, Shupe T and Atala A:
Organoid-on-a-chip and body-on-a-chip systems for drug screening
and disease modeling. Drug Discov Today. 21:1399–1411. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Rajasekar S, Lin DSY, Abdul L, Liu A,
Sotra A, Zhang F and Zhang B: IFlowPlate-A Customized 384-Well
Plate for the culture of perfusable vascularized colon organoids.
Adv Mater. 32:e20029742020. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Pinho D, Santos D, Vila A and Carvalho S:
Establishment of colorectal cancer organoids in microfluidic-based
system. Micromachines (Basel). 12:4972021. View Article : Google Scholar : PubMed/NCBI
|