Introduction
At present, 14 human peroxisomal biogenesis factor
(PEX) genes that encode peroxin proteins that serve a role in
various stages of peroxisome biogenesis have been identified,
including peroxisome matrix protein input, membrane formation and
peroxisome proliferation (1,2).
Peroxisome membrane proteins are also thought to function as
signaling platforms in reactive oxygen species (ROS)-induced
autophagy and antiviral immunity (3,4).
Peroxisome is a metabolic organelle involved in cellular redox
balance and lipid metabolism (5,6). The
function of peroxisome is important for ether phospholipid
synthesis, fatty acid oxidation, bile acid synthesis and ROS
homeostasis (7). Although
peroxisomes are involved in cell metabolism, their functional
effects in cancer remain unclear compared with those of other
metabolic organelles such as mitochondria (8,9).
In the last decade, peroxisome has also been
recognized as a central regulator of immunity (9). Lipid metabolites of peroxisomes, such
as polyunsaturated fatty acids (PUFAs), are precursors of important
immune mediators, including leukotrienes (LTs), and suppressors
(10–12). Peroxisome redox metabolism regulates
cellular immune signaling, such as activation of B-cell activated
NF-κB (13). In addition, the
development and activation of innate and adaptive immune cells are
regulated by peroxisome β-oxidation and ether lipid synthesis
(12,14). These findings open up avenues for
targeting peroxisome interventions for immune disorders,
inflammation and cancer. In addition, Lee et al (2) found that PEX13 is required for
selective autophagy (virus autophagy) of Sindbis virus and damaged
mitochondria (mitochondrial autophagy), and PEX13 mutant I326T and
W313G associated with the disease are defective in mitochondria
autophagy. The mitophagy function of PEX13 is shared with another
peroxin family member, PEX3, but not with the other two peroxins
required for general autophagy, namely PEX14 and PEX19 (15). Since PEX13 gene has a significant
role in various stages of peroxisome biogenesis and autophagy, and
peroxisome or autophagy has a crucial role in tumor occurrence and
development, the present study aimed to investigate the biological
functions that PEX13 may play in multiple cancers, especially in
pancreatic adenocarcinoma (PAAD) (16,17).
In the present study, systematic bioinformatics
analysis was conducted on patient data retrieved from The Cancer
Genome Atlas (TCGA) database to verify the biological function and
prognostic significance of PEX13 in various tumors. The present
study comprehensively explored the functional significance of
alterations in PEX13 expression level in various tumors in terms of
PEX13 expression level, prognosis, genetic alterations,
PEX13-related gene enrichment analysis, weighted correlation
network analysis (WGCNA), protein interaction, long non-coding
(lnc)RNA/circular (circ)RNA-micro (mi)RNA network and tumor
immunity. As a general surgery team, the present authors are
particularly interested in PAAD and focused. Finally, the
relationships between the expression of PEX13 and the biological
functions of PAAD were verified through in vitro
experiments.
Materials and methods
PEX13 mRNA and protein expression and
prognostic analyses
The PEX13 mRNA expression data for 33 tumors
retrieved from The Cancer Genome Atlas (TCGA) and Genotype-Tissue
Expression (GTEx) database were analyzed using the SangerBox portal
(http://SangerBox.com/Tool) (18). These tumors included adrenocortical
carcinoma (ACC), bladder urothelial carcinoma (BLCA), breast
invasive carcinoma (BRCA), cervical squamous cell carcinoma (CESC),
cholangiocarcinoma (CHOL), colon adenocarcinoma (COAD), lymphoid
neoplasm diffuse large b-cell lymphoma (DLBC), esophageal carcinoma
(ESCA), glioblastoma multiforme (GBM), head and neck cancer (HNSC),
kidney chromophobe (KICH), kidney renal clear cell carcinoma
(KIRC), kidney papillary cell carcinoma (KIRP), acute myeloid
leukemia (LAML), brain lower grade glioma (LGG), liver
hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), lung
squamous cell carcinoma (LUSC), mesothelioma (MESO), ovarian cancer
(OV), pancreatic adenocarcinoma (PAAD), pheochromocytoma and
paraganglioma (PCPG), prostate adenocarcinoma (PRAD), rectum
adenocarcinoma (READ), sarcoma (SARC), skin cutaneous melanoma
(SKCM), stomach adenocarcinoma (STAD), testicular germ cell tumors
(TGCT), thyroid carcinoma (THCA), thymoma (THYM), uterine corpus
endometrial carcinoma (UCEC), uterine carcinosarcoma (UCS) and
uveal melanoma (UVM). PEX13 mRNA expression distributions were
visualized in violin plots. Subsequently, the Gene Expression
Profiling Interactive Analysis 2 (GEPIA2) portal (genepattern.org)
was applied to further explore the PEX13 mRNA expression in 33
tumors (19). The
immunohistochemical (IHC) staining data for the analysis of PEX13
expression in various tumor tissues and corresponding normal
tissues were obtained from The Human Protein Atlas (THPA) portal
(https://www.proteinatlas.org/) (20). The analyzed tumors included glioma
and breast, colorectal, liver, lung, pancreatic, skin, stomach,
thyroid and renal cancer. Survival prognosis analysis was performed
using GEPIA2 and SangerBox. Through GEPIA2, the overall survival
(OS) map data for PEX13 in multiple tumors of TCGA database was
obtained. The low- and high-expression cohorts of PEX13 were
obtained using the expression thresholds of cutoff-high (>50%)
and cutoff-low (<50%) values. Cox regression analysis was used
to analyze the OS, disease-free interval (DFI), progression-free
interval (PFI) and disease-specific survival (DSS) data in various
cancers using SangerBox portal.
Genetic alteration analysis
The characteristics of PEX13 genetic alterations
were explored in multiple tumors using the cBioPortal database
(https://www.cbioportal.org/), including
mutation, amplification and deep deletion (21). The structural variant data, mutation
data and copy number alteration (CNA) data were obtained from the
cBioPortal database. Data of somatic CNAs and somatic mutations of
PAAD were obtained from TCGA datasets (gdc.cancer.gov/). The CNAs
were related to the PEX13 expression and the threshold CNA peaks
were analyzed through GISTIC 2.0 (https://cloud.genepattern.org/) (22). According to the PEX13 expression
level, the patients with PAAD were divided into the first 25%
PEX13low (n=46) and the last 25% PEX13high (n=46) groups. To
download and visualize the somatic mutations of patients with PAAD,
the ‘maftools’ package (bioconductor.org/packages/release/bioc/vignettes/maftools/inst/doc/maftools.html#7_Visualization)
was applied in R software (4.2.1) (https://www.r-project.org/) (23).
PEX13-related genes enrichment
analysis
Through the GEPIA2 portal, the top 200 PEX13-related
genes were explored in TCGA database. Subsequently, the TIMER2
(http://timer.cistrome.org/) portal was
used to generate a heatmap of the top 10 PEX13-related genes in
various cancers, including KCMF1, KIAA1841, MPHOSPH10, MRPL19,
PPP1CB, PUS10, RAB1A, SLC30A6, SMEK2 and YIPF4 (24). The correlation coefficient (R) and
P-value were calculated. The Gene Ontology (GO)-biological process
(BP) subontology and Kyoto Encyclopedia of Genes and Genomes (KEGG)
analysis of 200 PEX13-correlated genes in pan-cancers were explored
via DAVID (https://david.ncifcrf.gov/home.jsp) and SangerBox
portal. Moreover, the top 50 PEX13 negatively related genes and top
50 PEX13 positively related genes were explored and heatmaps were
generated in PAAD using LinkedOmics (www.linkedomics.org/login.php) (25). The GO-BP and KEGG enrichment
analysis of the top 50 PEX13 negatively related genes and top 50
PEX13 positively related genes in PAAD were explored via DAVID and
SangerBox portal. Linkedomics (linkedomics.org) portal was used to
analyze the enrichment pathways of PEX13 positively and negatively
related genes in PAAD (25).
WGCNA
The hub genes affected by PEX13 expression level
were obtained through WGCNA using SangerBox portal. First, the PAAD
gene expression profile in TCGA database was used to calculate the
median absolute deviation (MAD) of each gene. The outlying genes
and samples were removed using the GoodSamplesGenes method of the
‘WGCNA’ R software package
(genetics.ucla.edu/labs/horvath/CoexpressionNetwork/Rpackages/WGCNA)
(26). WGCNA was then applied to
build a scale-free co-expression network. As a soft-thresholding
parameter, β could emphasize strong gene-gene associations and
penalize weak correlations. To ensure a scale-free network, the
power of β=7 (0.85) was selected as the soft-thresholding parameter
and the power of β=7 (16.23) was selected as the soft-thresholding
parameter in mean connectivity. Moreover, the PAAD samples were
clustered using Pearson's correlation and average linkage method.
The average linkage hierarchical clustering identified 23 modules
and the differences among these were analyzed according to the
expression level of PEX13, among which darkgrey module had the
strongest association with PEX13 expression. Consequently, the
darkgrey module was chosen as the important module for extracting
hub genes.
Protein-protein interaction and
ncRNA-miRNA-PEX13 regulatory networks analyses
Through the STRING database (https://string-db.org/), 50 PEX13 interacting proteins
were identified in pan-cancers. Using Cytoscape (3.10.1;
cytoscape.org/download.html), a PEX13 interacting protein network
map was generated. The key protein-protein interaction network of
PEX13 was extracted in this protein-protein interaction network
using Cytoscape. Molecular docking technique was applied to predict
the binding sites between PEX13 and PEX2, PEX12, PEX14 and ABCD3.
By using the HDOCK server (http://hdock.phys.hust.edu.cn/), the binding sites
between PEX13 and PEX2, PEX12, PEX14 and ABCD3 were analyzed. Using
the PyMOL tool (2.5.5; pymol.org/2/), the binding sites between
PEX13 and PEX2, PEX12, PEX14 and ABCD3 were visualized (27).
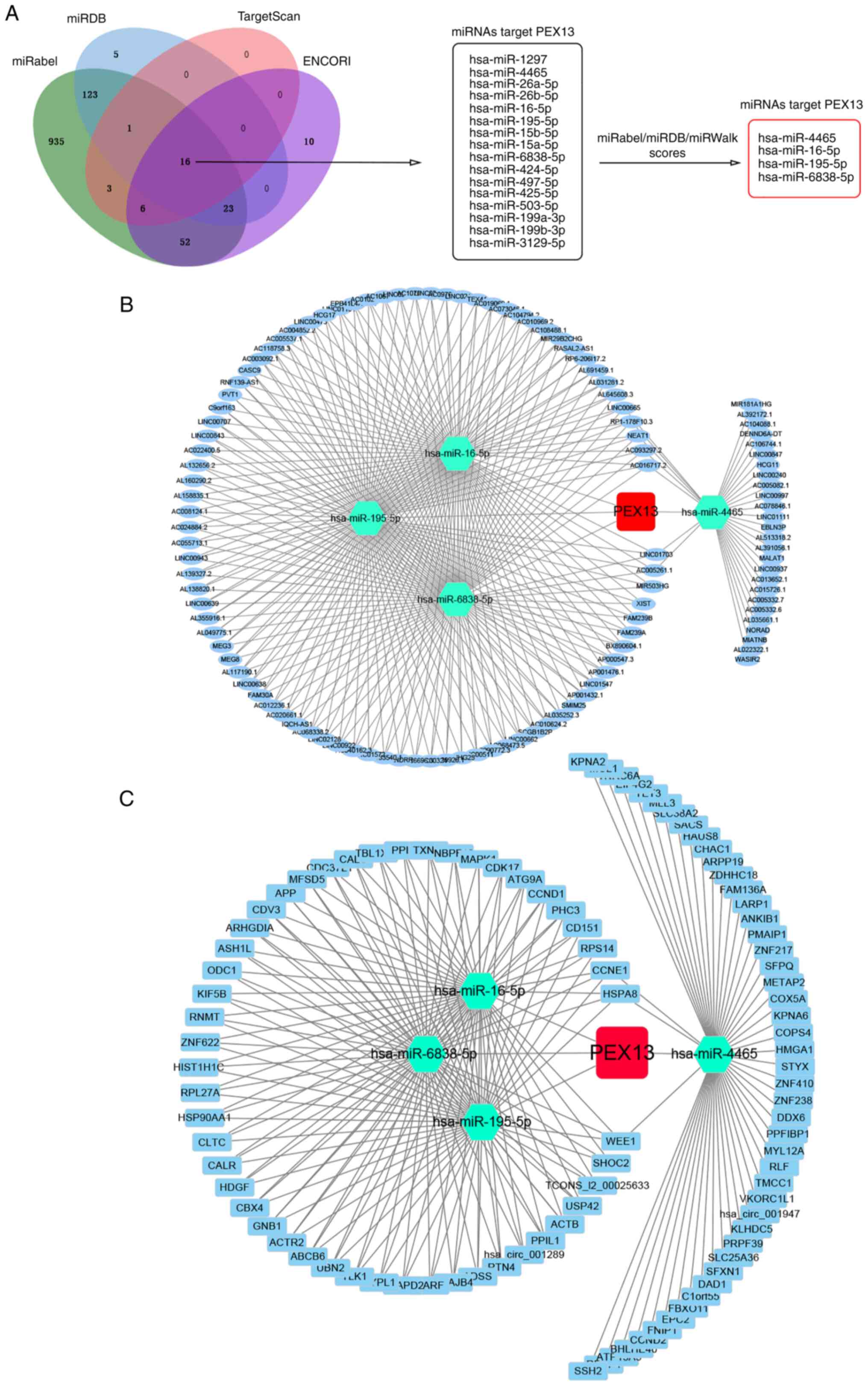
In addition, lncRNA/circRNA-miRNA-PEX13 networks
were investigated. Through the niRabel (http://bioinfo.univ-rouen.fr/mirabel/index.php),
miRDB (http://mirdb.org/), TargetScan (http://www.targetscan.org/vert_72/) and ENCORI
(https://starbase.sysu.edu.cn/)
databases, 16 miRNAs that might target PEX13 mRNA were identified.
Subsequently, four miRNAs were further screened and selected
through miRabel, miRDB and miRWalk scores, including hsa-miR-4465,
hsa-miR-16-5p, hsa-miR-195-5p and hsa-miR-6838-5p. Using the ENCORI
online tool, 105 lncRNAs and 96 circRNAs that targeted
hsa-miR-4465, hsa-miR-16-5p, hsa-miR-195-5p and hsa-miR-6838-5p
were predicted, and the lncRNA-miRNA-PEX13 and circRNA-miRNA-PEX13
regulatory networks were visualized via Cytoscape tool.
Immune-related analysis
The SangerBox portal was used to investigate the
relationships between PEX13 mRNA expression and tumor infiltration
immune cells (TIICs) in various cancers, including T_cells_CD8,
T_cells_CD4_memory_resting, B_cell_memory, Tregs,
NK_cells_activated, macrophages and dendritic_cells_activated.
Subsequently, the correlations between PEX13 mRNA expression and
immune checkpoint (ICP) genes, microsatellite instability (MSI),
tumor mutation burden (TMB) and tumor purity in multiple TCGA
tumors we investigated using SangerBox. Furthermore, TIMER2 portal
was applied to analyze the associations between PEX13 and the
infiltration levels of B cell and macrophage in pan-cancers,
especially in PAAD via different algorithms, such as XCELL,
CIBERSORT and EPIC. Moreover, the relationships between PEX13 and
stromal score, immune score and ESTIMATE score in multiple cancers
were explored using SangerBox, especially in lung adenocarcinoma
(LUAD), stomach and esophageal carcinoma (STES), sarcoma (SARC) and
lung squamous cell carcinoma (LUSC).
Cell culture
The human PAAD cell lines of PANC1 and BxPC3 were
obtained from the Shanghai Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). Comprehensive cell line authentication
was performed and cells were periodically checked for mycoplasma
contamination. The DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (FBS; Satorius AG) was used to culture
the PAAD cell lines. The PAAD cells were incubated in a cell
incubator at 37°C with 5% CO2.
Small-interfering (si)RNAs delivery,
RNA isolation and reverse transcription-quantitative polymerase
chain reaction (RT-qPCR)
siRNAs used to knock down the expression of PEX13
were obtained from Shanghai GenePharma Co., Ltd. A total of
2×105 PAAD cells were seeded in 6-well plates, 4.5 µl
Lipofectamine™ RNAiMAX reagent (Thermo Fisher Scientific, Inc.) and
40 pmol siRNA were mixed for 10 min in each well at room
temperature. Subsequently, the mixture was added to the cells
cultured with FBS-free medium and the cells were replaced with full
medium at 24 h after transfection. The PAAD cells were collected
after 48 h with TRIzol® reagent (Takara Bio, Inc.). The
GoScript reverse transcription system (Promega Corporation) was
applied to generate PEX13 cDNA and the GoTaq® qPCR
Master Mix (Promega Corporation) was used to detect the knockdown
efficiency of PEX13 siRNAs through qPCR using the ABI QuantStudio 3
system. The siRNA sequences were as follows: si-PEX13-1 sense,
5′-ACUUGAUUCCACCGUUUUCCUTT-3′ and antisense,
5′-GAAAACGGUGGAAUCAAGUAATT-3′; si-PEX13-2 sense,
5′-UUAUUCUAGUCCUGAAUUCCUTT-3′ and antisense,
5′-GAAUUCAGGACUAGAAUAAAGTT-3′; and siRNA negative control (si-NC)
sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. The thermocycling conditions used for
qPCR were: Initial denaturation at 95°C for 10 min; 40 cycles of
amplification for 15 sec at 95°C and 45 sec at 60°C, and an
extension step at 72°C for 2 min. The relative expression levels of
the PEX13 mRNA were calculated using the 2−ΔΔCq method
and normalized to the internal control GAPDH (28). The primer sequences were as follows:
GAPDH forward, 5′-GGTGGTCTCCTCTGACTTCAACA-3′ and reverse,
5′-GTTGCTGTAGCCAAATTCGTTGT-3′ reverse; PEX13 forward,
5′-GCCCCACTTTCCAATCTGCT-3′ and reverse,
5′-AGAATAGGTGGGGGCACTCT-3′.
Cell Counting Kit-8 (CCK-8) assay
A total of 2×103 PANC1 or BxPC3 cells
were plated into 96-well plates. PEX13 expression was knocked down
using the PEX13 siRNAs. Subsequently, the optical density (OD) at
450 wavelengths was measured for 5 consecutive days (at days 0, 1,
2, 3, 4 and 5) after incubation for 1 h at 37°C with the CCK-8
reagent (ApexBio Technology) using a multiplate reader.
Wound healing and Transwell
assays
A total of 2.5×105 PANC1 or BxPC3 cells
were plated in 6-well plates. A 200 µl pipette tip was used to
scratch the cell monolayer. After 0, 12 and 24 h incubation in an
FBS-free medium, the images of wound closure were captured and
further explored via an inverted microscope (Olympus Corporation)
and Image J software (1.8.0; National Institutes of Health). For
the Transwell assay, 8-µm pore filters coated with Matrigel matrix
glue (Corning, Inc.) were used. Matrigel was added into the upper
chamber for 40 min at 37°C. A total of 5×104 PANC1 and
BxPC3 cells in 300 µl serum-free DMEM were seeded into the upper
chamber and 600 µl DMEM with 10% FBS medium was added to the lower
chamber. After 24 h, cells were fixed with 4% paraformaldehyde at
room temperature and stained with 1% crystal violet at room
temperature. The images were captured using a microscope (BX53;
Olympus Corporation).
Colony formation assay
In 6-well plates, 1×103 PANC1 and BxPC3
cells were seeded in each well with 2.5 ml DMEM containing 10% FBS.
The cells were incubated for 2 weeks at 37°C in a humidified
chamber with 5% CO2. The colonies were fixed with 4%
paraformaldehyde for 30 min at room temperature before being
stained with crystal violet for 10 min at room temperature. A cell
mass of >50 cells was considered a colony, and Image J (1.8.0)
was used for counting colonies.
Statistical analysis
All of the experimental data were statistically
analyzed using GraphPad Prism 8.0 (Dotmatics). Comparisons between
two groups were made using an unpaired t-test and the statistical
method used for survival analysis was Kaplan-Meier. Cox regression
analysis was performed using logrank test to obtain prognostic
significance. The correlation analyses in the manuscript were
applied using Pearson's correlation analysis. One-way ANOVA with
Bonferroni's post-hoc test was used to compare the differences
between multiple groups. Two-way ANOVA with Bonferroni's post-hoc
test was used to compare the differences between CCK-8 groups
(si-NC and si-PEX13-1/si-PEX13-2). The data are presented as mean ±
standard deviation from three individual experiments and each
experiment was repeated at least three times. P<0.05 was
considered to indicate a statistically significant difference.
Results
PEX13 expression patterns in various
cancers
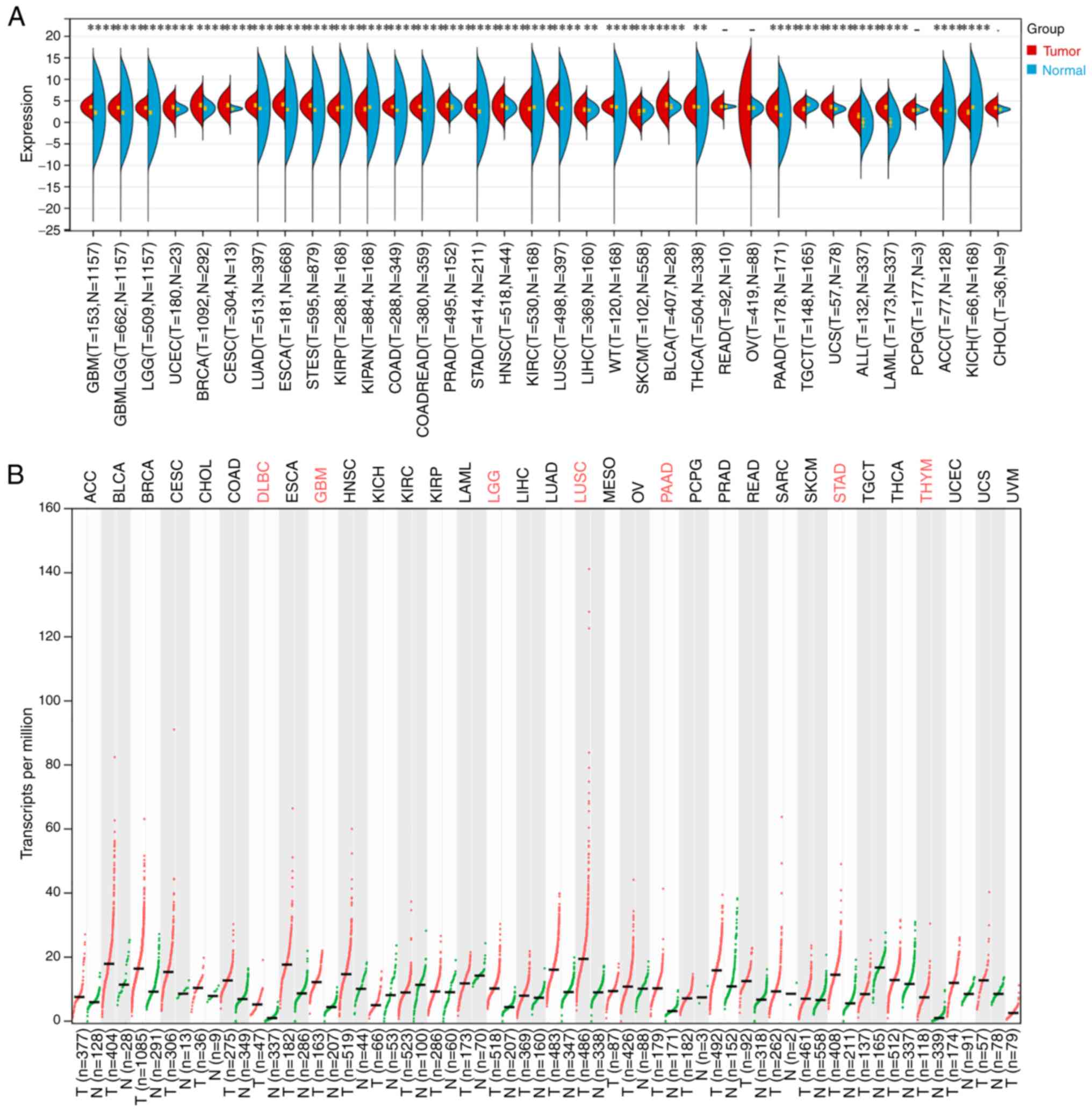
Through TCGA and GTEx databases analysis, PEX13 was
revealed to be highly expressed in multiple tumors, including GBM,
LGG, GBMLGG, BRCA, UCEC, LUAD, CESC, ESCA, COAD, STES, READ, STAD,
PRAD, HNSC, LUSC, LIHC, high-risk wilms tumor (WT), BLCA, THCA,
PAAD, UCS, LAML, acute lymphoblastic leukemia (ALL) and ACC. In
addition, the expression level of PEX13 was downregulated in KIRP,
pan-kidney cohort (KIPAN), KIRC, SKCM, TGCT and KICH compared with
the corresponding normal tissues (Fig.
1A). In GEPIA2 portal, it was discovered that PEX13 was
remarkable upregulated in DLBC, GBM, LGG, LUSC, PAAD, STAD and THYM
(Fig. 1B). In addition, the results
of IHC showed that the expression of PEX13 protein in breast
cancer, colorectal cancer, glioma, liver cancer, lung cancer,
pancreatic cancer, skin cancer, stomach cancer, thyroid cancer and
renal cancer was higher than that in normal tissues (Fig. S1). The present findings indicated
that PEX13 mRNA and protein expression levels are increased in a
variety of tumors, suggesting that PEX13 may function as an
oncogene in various cancers, including PAAD.
PEX13 expression and prognosis across
multiple types of cancer
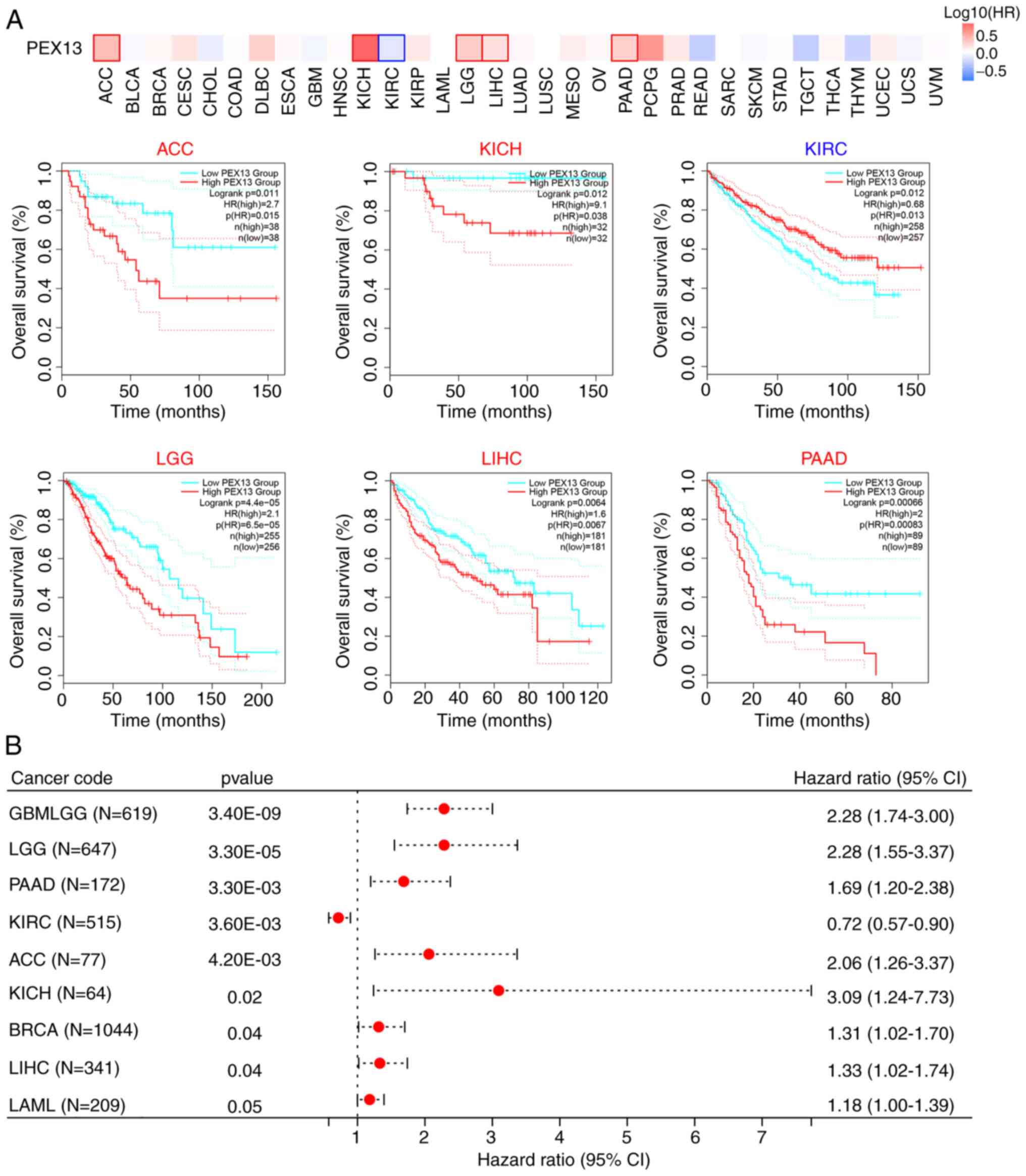
The prognostic value of PEX13 in various tumors was
analyzed by comparing the OS of patients with PEX13 expression.
Analysis of the pan-cancer cohort using the GEPIA2 database
demonstrated that high PEX13 mRNA expression levels was associated
with a significantly lower OS in patients with ACC, KICH, LGG, LIHC
and PAAD, and high PEX13 mRNA expression levels were associated
with a significantly higher OS in patients with KIRC (Fig. 2A). Cox regression analysis in
SangerBox database suggested that PEX13 mRNA expression level was
related to OS, DFI, DSS and PFI in patients with various tumors.
Regarding the OS, the findings revealed that high PEX13 mRNA
expression was linked to a shorter OS in GBMLGG, LGG, PAAD, ACC,
KICH, BRCA, LIHC and LAML (Fig.
2B). Regarding the DFI, high PEX13 mRNA expression was linked
to a shorter DFI in PAAD, KIPAN, KIRP and ACC (Fig. S2A). In PFI, the high PEX13
expression was related to shorter PFI in GNMLGG, ACC, LGG, PAAD,
LIHC, KICH and UVM (Fig. S2B).
Furthermore, a high PEX13 mRNA expression was correlated with a
shorter DSS in GBMLGG, LGG, PAAD, ACC, KICH and LIHC (Fig. S2C). It was also observed that high
PEX13 mRNA expression was significantly correlated with shorter OS,
DFI, PFI, and DSS in PAAD and ACC. The present results revealed
that the level of PEX13 mRNA expression was related to the
prognosis of various tumors. Furthermore, high PEX13 mRNA
expression was linked to a worse prognosis in patients with
PAAD.
PEX13 genetic alterations in various
cancers
Mutations, deletions or amplification of oncogenes
or tumor suppressor genes have been linked to the growth and
progression of several tumors (29). Hence, the present study analyzed
diverse types of genetic alterations in the PEX13 gene using the
cBioPortal portal, including amplification, mutation and deep
deletion. The most common genetic alteration of PEX13 gene in DLBC
(18.75%), LUSC (5.75%), BLCA (2.68%), UCS (1.75%), ovarian serous
cystadenocarcinoma (1.54%) and ESCA (1.1%) was amplification. In
UCEC (2.46%), STAD (1.36%), COADREAD (1.35%) and KIRP (0.71%), the
most common genetic alteration of PEX13 gene was mutation (Fig. 3A). Subsequently, the TCGA-PAAD
dataset was interrogated to investigate the correlation between
PEX13 and specific genomic features such as copy number variations
(CNVs) and somatic mutations. Fig.
3B shows the comparison of the CNV profiles in the PEX13high
(n=46) and PEX13low (n=43) groups. In the PEX13high group, deletion
peaks were found in the 5p12.3, 6p25.3, 9p21.3, 10p15.3, 10q23.2,
18q12.1, 18q21.2 and 19p13.2 chromosomal locations, while
amplification peaks were found in the 1p12, 6p21.31, 7q21.13,
8q24.21, 9p13.3, 12p13.33, 18q11.2 and 19q13.2 chromosomal regions
(Fig. 3C-D). In the PEX13low group,
deletion peaks were found in the 9p23 and 9p21.3 chromosomal
locations, while frequent amplification peaks were found in the
8p11.23 and 19q13.2 chromosomal regions (Fig. 3C and D). These findings suggested
that high PEX13 expression led to more deletions and amplifications
of multiple sites in the genome of patients with PAAD. The PEX13low
group indicated that high frequency of somatic mutations in the
FRG1B (72%), KRAS (49%), TP53 (47%), UBBP4 (44%) and TTN (40%)
genes and the PEX13high group suggested that high frequency of
mutations in the KRAS (89%), FRG1B (76%), TP53 (72%), MAMLD1 (35%),
MED12L (35%) and UBBP4 (30%) genes (Fig. 3E and F). These findings revealed
that high PEX13 expression may lead to the increased mutation
probability of KRAS, FRG1B, TP53 and other genes, and to the
decreased mutation probability of UBBP4 gene in PAAD, which may be
a potential mechanism leading to the occurrence and development of
tumors.
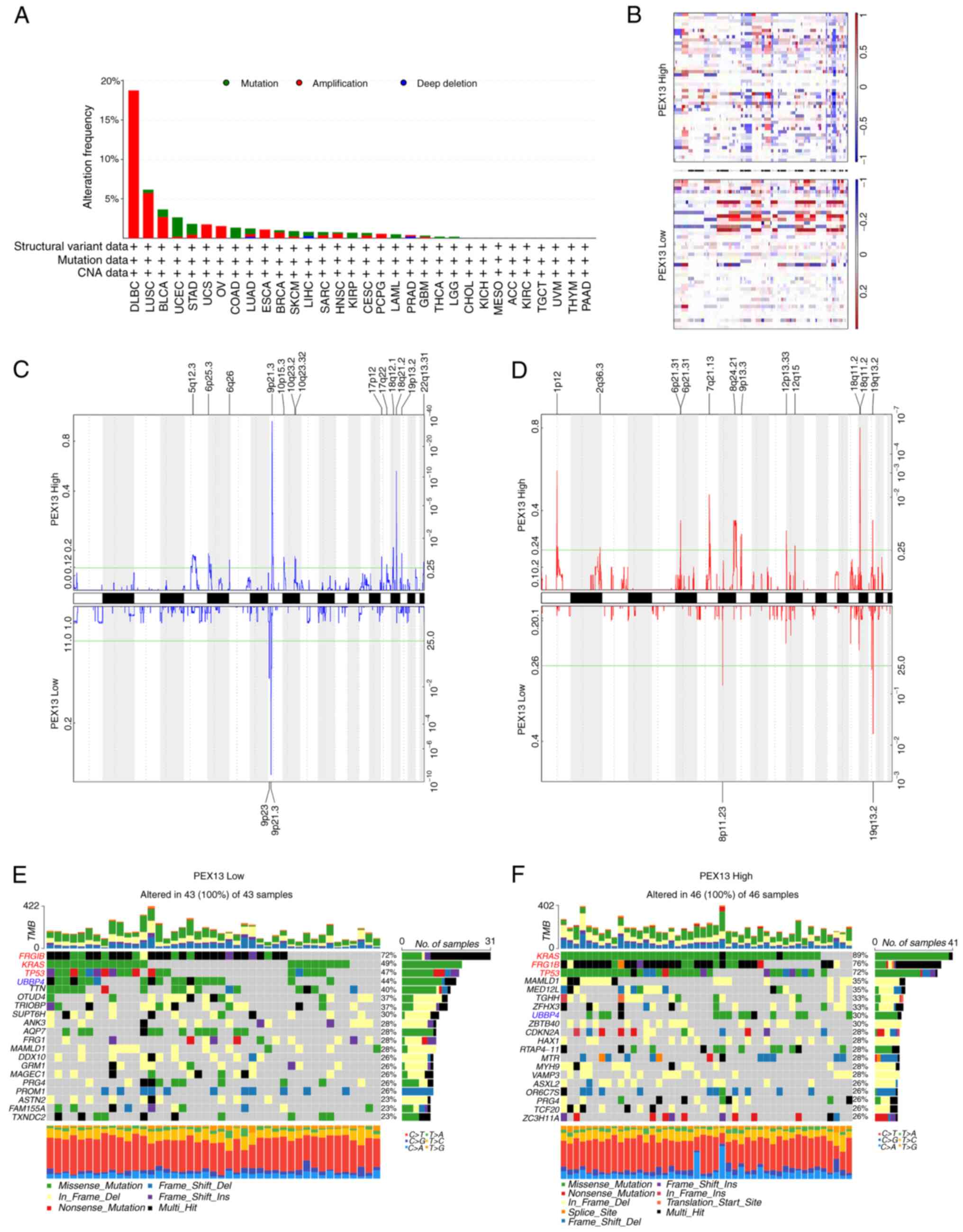 | Figure 3.Distinct genomic profiles associated
with PEX13 expression in pan-cancers, including PAAD. (A) Genetic
alterations of PEX13 in different cancers in TCGA database were
discovered using the cBioPortal tool, including mutation,
structural variant, amplification and deep deletion. (B) The copy
number alteration profile analysis between 25% PEX13low and 25%
PEX13high groups in a PAAD cohort was preformed using GISTIC2.0.
Frequencies of (C) deletion and (D) amplification of PAAD
associated with PEX13 expression (blue, deletion; red,
amplification). Detection of somatic mutations in PAAD, including
(E) 25% PEX13low and (F) 25% PEX13high group. PEX, peroxisomal
biogenesis factor. TCGA, The Cancer Genome Atlas; PAAD, pancreatic
adenocarcinoma. |
These findings suggested the presence of PEX13 gene
mutations, amplification and deletion in multiple tumors. The PAAD
tissues revealed different somatic mutations and CNVs depending on
the level of PEX13 expression. This suggested that alterations in
the PEX13 gene could influence the occurrence and progression of
multiple cancers, particularly PAAD.
Pathway enrichment analysis of
PEX13-associated genes
The PEX13-related genes were analyzed in 33 cancers
through GEPIA2 database and 200 PEX13-related genes were obtained.
The top 10 genes with the highest correlation among 200
PEX13-related genes were generated correlation heat maps in 33
cancers via TIMER2, including KCMF1, KIAA1841, MPHOSPH10, MRPL19,
PPP1CB, PUS10, RAB1A, SLC30A6, SMEK2 and YIPF4 (Fig. 4A). Fig.
4B shows that PEX13 expression was positively correlated with
PUS10 (R=0.61), KIAA1841 (R=0.61), RAB1A (R=0.59), SMEK2 (R=0.58),
YIPF4 (R=0.57), PPP1CB (R=0.57), KCMF1 (R=0.57), SLC30A6 (R=0.57),
MPHOSPH10 (R=0.56) and MRPL19 (R=0.56) in GEPIA2. The GO-BP
analysis of 200 PEX13-related genes in 33 tumors showed that
PEX13-related genes were mainly enriched in mRNA metabolic process,
cellular component biogenesis, cell division, intracellular
transport, cytokinesis, mRNA processing, cell cycle and RNA
splicing (Fig. 4C). PEX13-related
genes were mainly enriched in Endocytosis, mRNA surveillance
pathway, nucleocytoplasmic transport, T cell receptor, signaling
pathway and hippo signaling pathway based on the results of the
KEGG enrichment analysis (Fig.
4D).
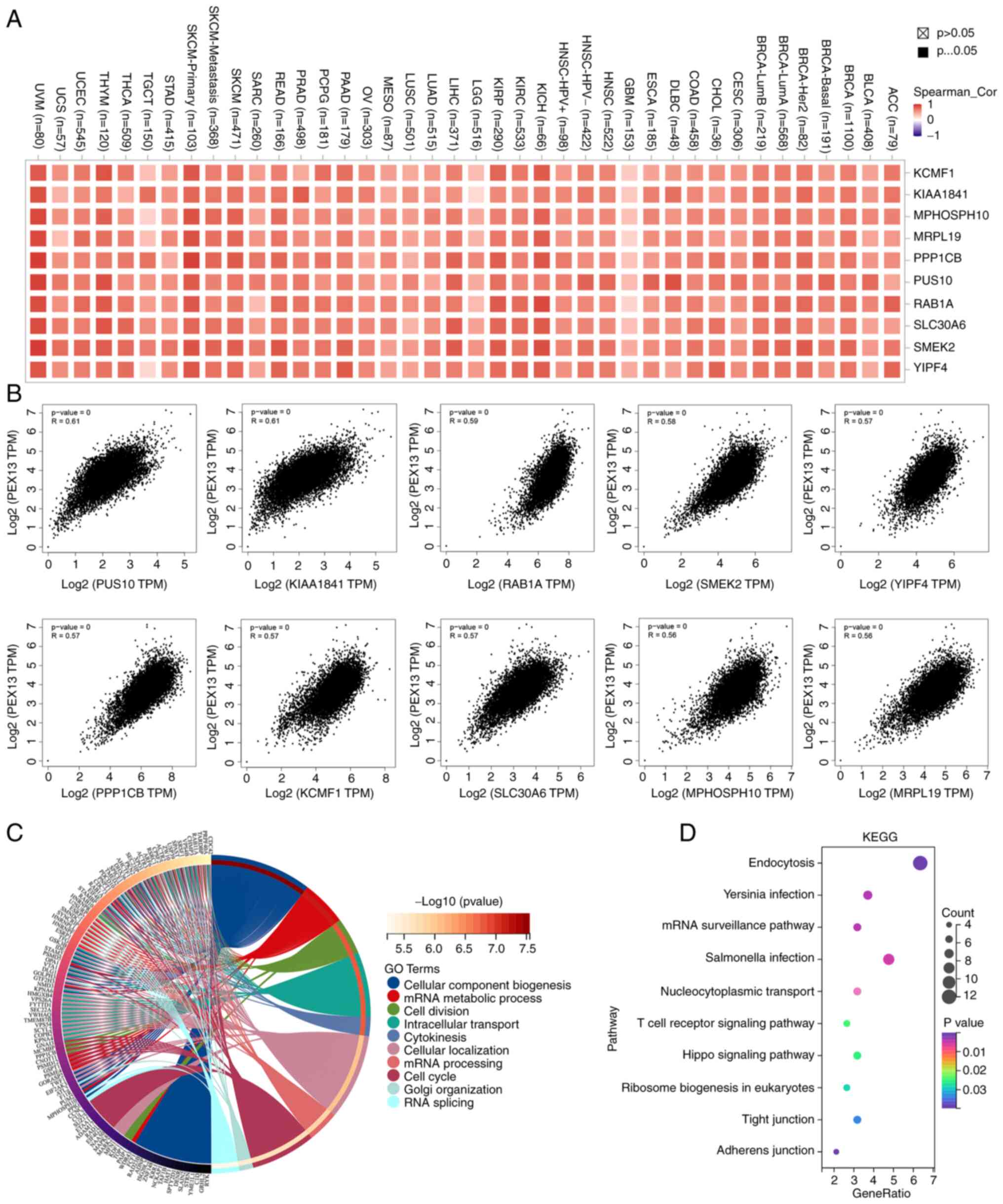 | Figure 4.The enrichment analysis of
PEX13-related genes in pan-cancers. (A) Correlation heat map
displayed the top 10 PEX13-related genes in various tumors via the
TIMER2.0 portal. (B) Correlations between PEX13 expression and top
10 PEX13-related genes in pan-cancers via GEPIA2. The (C) Gene
Ontology-Biological Process and (D) KEGG enrichment analyses of 200
PEX13-related genes in various cancers. PEX, peroxisomal biogenesis
factor; TPM, transcripts per million; KEGG, Kyoto Encyclopedia of
Genes and Genomes; UVM, uveal melanoma; UCS, uterine
carcinosarcoma; UCEC, uterine corpus endometrial carcinoma; TGCT,
testicular germ cell tumors; THCA, thyroid carcinoma; THYM,
thymoma; STAD, stomach adenocarcinoma; SKCM, skin cutaneous
melanoma; SARC, sarcoma; READ, rectum adenocarcinoma; PRAD,
prostate adenocarcinoma; PCPG, pheochromocytoma and paraganglioma;
PAAD, pancreatic adenocarcinoma; OV, ovarian serous
cystadenocarcinoma; MESO, mesothelioma; LUSC, lung squamous cell
carcinoma; LUAD, lung adenocarcinoma; LIHC, liver hepatocellular
carcinoma; LGG, brain lower grade glioma; KIRC, kidney renal clear
cell carcinoma; KICH, kidney chromophobe; HNSC, head and neck
cancer; HPV, human papillomavirus; GBM, glioblastoma multiforme;
ESCA, esophageal carcinoma; DLBC, lymphoid neoplasm diffuse large
b-cell lymphoma; COAD, colon adenocarcinoma; CHOL,
cholangiocarcinoma; CESC, cervical squamous cell carcinoma and
endocervical adenocarcinoma; BRCA, breast invasive carcinoma; BLCA,
bladder urothelial carcinoma; ACC, adrenocortical carcinoma. |
To further explore the biological functions of PEX13
in PAAD, the enrichment of PEX13 positively and negatively related
genes and their enrichment pathways in PAAD were analyzed using the
Linkedomics portal. Fig. S3A shows
the heat maps of 50 positively and 50 negatively related genes of
PEX13 in PAAD. Fig. S3B-E showed
the GO-BP and KEGG enrichment analysis of 100 positively and 100
negatively related genes of PEX13, respectively. The results showed
that GO-BP was mainly enriched in positive regulation of drug
response, small GTPase mediated signal transduction, regulation of
epidermal growth factor-activated receptor activity and ERBB
signaling pathway among the positively related genes of PEX13 and
in the regulation of glycogen (starch) synthase activity,
intracellular protein transport, intracellular protein transport
and autophagosome maturation among the negatively related genes of
PEX13 (Fig. S3B and C). For the
KEGG enrichment analysis, PEX13-positive related genes were mainly
enriched in Rap1 signaling pathway, ErbB signaling pathway, EGFR
tyrosine kinase inhibitor resistance, Neurotrophin signaling
pathway and AMPK signaling pathway and PEX13-negatively related
genes were mainly enriched in Vitamin B6 metabolism, Apelin and
Insulin signaling pathway (Fig.
S3D-E). These findings show that PEX13 could regulate mRNA
processing, RNA splicing, cell cycle and the response to drugs in
cancers. In PAAD, PEX13 expression was related to protein
transport, tumor cell response to drugs, multiple cancer pathways
and immune pathways.
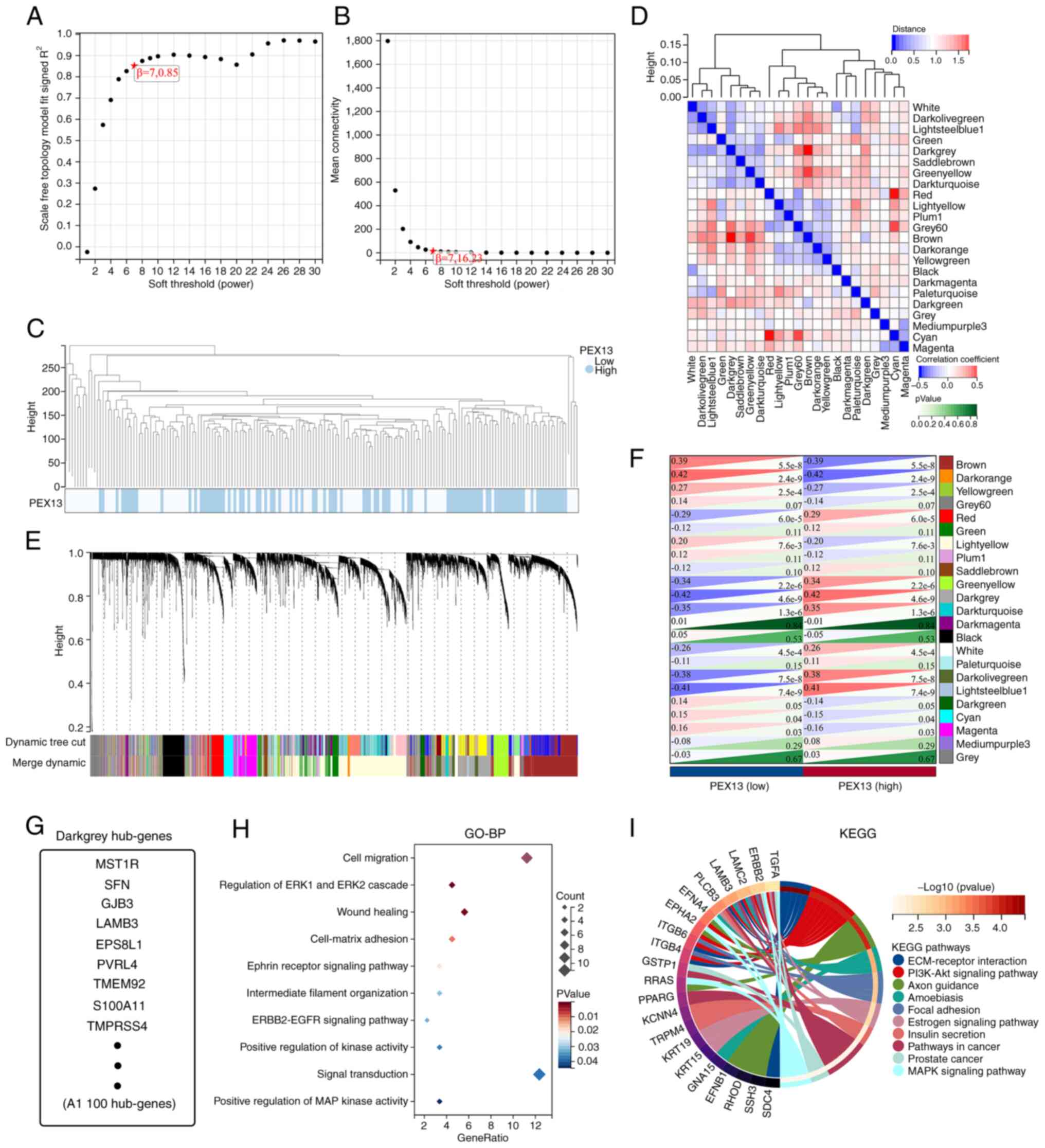
Analysis of WGCNA in PAAD
The TCGA-PAAD cohort was analyzed via WGCNA to
identify hub genes strongly associated with PEX13 expression in
PAAD. Using the average linkage approach and Pearson's correlation
method, the samples of TCGA-PAAD data were first clustered
(Fig. 5A-C). With the help of the
average linkage hierarchical clustering, a total of 23 modules were
obtained (Fig. 5D). According to
the expression level of PEX13, we analyzed the differences among 23
modules, among which darkgrey module had the strongest association
with PEX13 expression (Fig. 5E-F).
The darkgrey module was selected as the significant module for
further analysis. Fig. 5G shows the
hub genes of darkgrey module and GO-BP and KEGG enrichment analyses
were used to analyze these hub genes. In GO-BP analysis, hub genes
mainly enriched in cell migration, regulation of ERK1 and ERK2
cascade, wound healing, ephrin receptor signaling pathway,
ERBB2-EGFR signaling pathway, positive regulation of MAP kinase
activity and signal transduction (Fig.
5H). In KEGG enrichment analysis, hub genes mainly enriched in
PI3K-Akt signaling pathway, extracellular matrix (ECM)-receptor
interaction, focal adhesion, pathways in cancer and MAPK signaling
pathway (Fig. 5I). These results
suggested that the hub genes of darkgrey module with different
PEX13 expression were mainly involved in cell migration, wound
healing, PI3K-Akt and MAPK signaling pathway.
Construction of protein interaction
and ncRNA-miRNA-PEX13 regulatory network
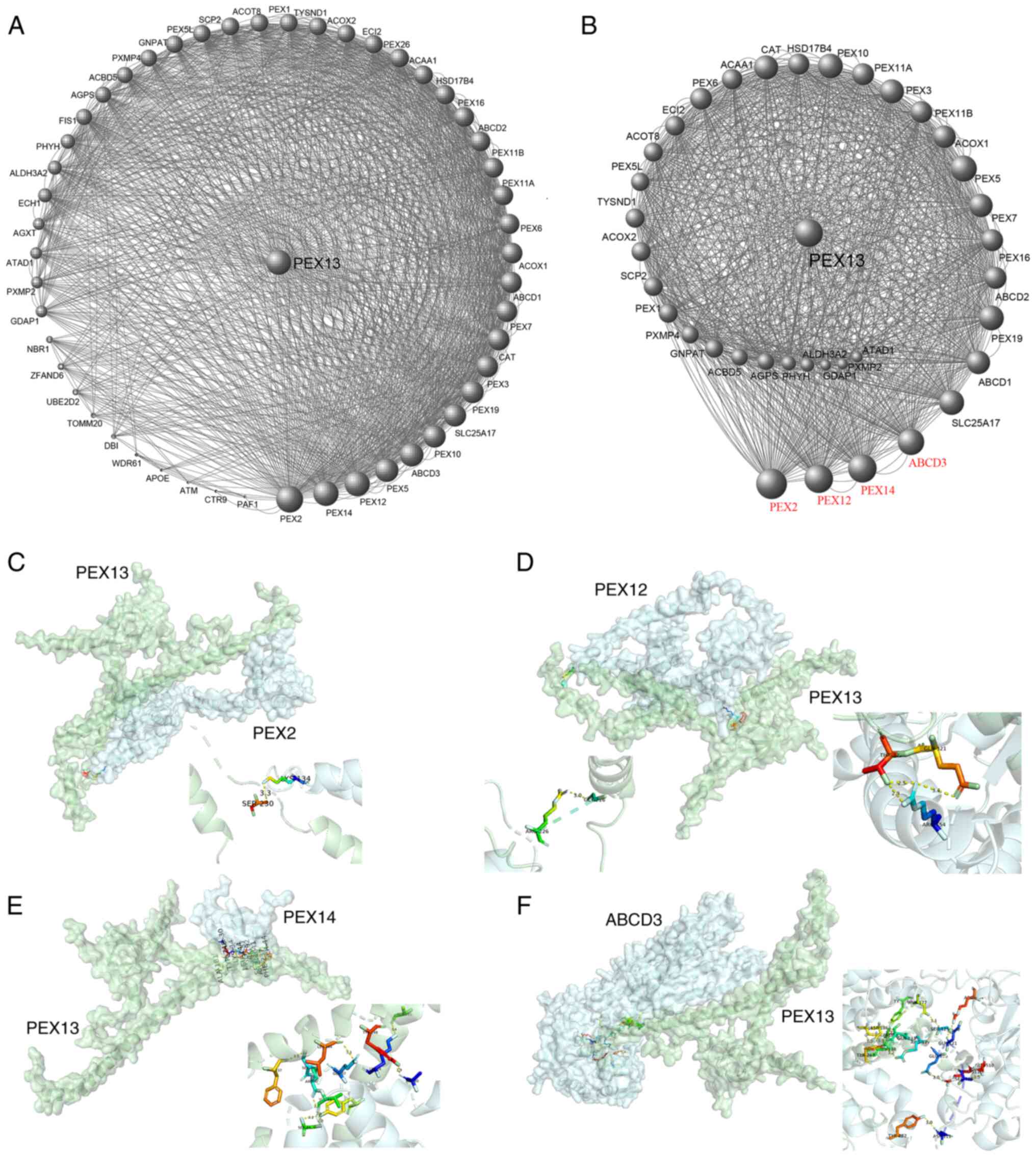
The 50 PEX13 interacting proteins were identified
using STRING database and the PEX13 interacting protein network map
was generated in Cytoscape (Fig.
6A). In this protein interaction network, key protein
interaction network of PEX13 was extracted using Cytoscape, as
shown in Fig. 6B. The PEX13
interacting proteins were scored in Cytoscape and a larger volume
indicated higher interacting score of these proteins in the network
diagram. Subsequently, four proteins had the highest interaction
scores with PEX13, including PEX2, PEX12, PEX14 and ABCD3 (Fig. 6B). Molecular docking technique was
applied to forecast the binding sites between PEX13 and PEX2,
PEX12, PEX14 and ABCD3 (Fig. 6C-F).
The results showed that PEX13 may interact with PEX2, PEX12, PEX14
and ABCD3 to influence the biological functions and acted as a
significant role in various tumors.
In addition, numerous studies found that lncRNA and
circRNA regulated the expression of downstream mRNA by adsorbing
target miRNAs, which acted as a crucial factor in tumorigenesis and
the progression of tumors (30,31).
Therefore, the present study explored the lncRNA/circRNA-miRNA
network that may regulate mRNA expression in multiple cancers.
LncRNA-miRNA regulatory networks of PEX13 were constructed to
investigate the underlying molecular mechanisms of PEX13 in tumors.
Through the miRabel, miRDB, TargetScan and ENCORI databases, 16
miRNAs that might target PEX13 mRNA were identified (Fig. 7A). Subsequently, four miRNAs,
including hsa-miR-4465, hsa-miR-16-5p, hsa-miR-195-5p and
hsa-miR-6838-5p, were screened and selected using miRabel, miRDB
and miRWalk. Using the ENCORI online tool, the lncRNAs and circRNAs
that targeted their corresponding miRNAs were predicted and
Cytoscape was used to visualize the lncRNA-miRNA and circRNA-miRNA
regulatory networks (Fig. 7B and
C). These findings suggested the upstream ncRNA-miRNA
regulatory networks that may control the aberrant expression of
PEX13 in cancers.
PEX13 regulated tumor immunity in
multiple tumors
The relationships between PEX13 and the tumor immune
microenvironment (TIM) were further analyzed in a variety of
tumors, and the correlation between PEX13 and the composition of
tumor infiltrating immune cells (TIICs) was investigated in
multiple cancers. Studies showed that TIICs were a significant
component of the tumor microenvironment and regulated the
occurrence, development and metastasis of tumors (32,33).
The Sangerbox database was applied to investigate the correlation
between PEX13 and TIIC levels in diverse tumors. The expression of
PEX13 in LAML, PRAD, BRCA, PAAD, STES, HNSC, LIHC, KIRC, THCA,
STAD, SKCM, THYM and CESC was remarkably related to various immune
cells, including T_cells_CD4_memory_resting, B_cell_memory,
T_cells_CD8, macrophages_M1, macrophages_M2, Tregs,
NK_cells_activated, dendritic_cells_activated and neutrophils
(Fig. 8A). Fig. S4A demonstrated the relationship
between PEX13 expression and B cell infiltration levels in
different tumors. In PAAD, the expression level of PEX13 under
different algorithms was mainly negatively related to the
infiltration level of B cells (Fig.
S4B). Fig. S4C presents a heat
map of the relationship between PEX13 expression levels and
macrophage infiltration levels in various tumors. PEX13 expression
level was also negatively related to the infiltration level of
macrophages under different algorithms in PAAD (Fig. S4D).
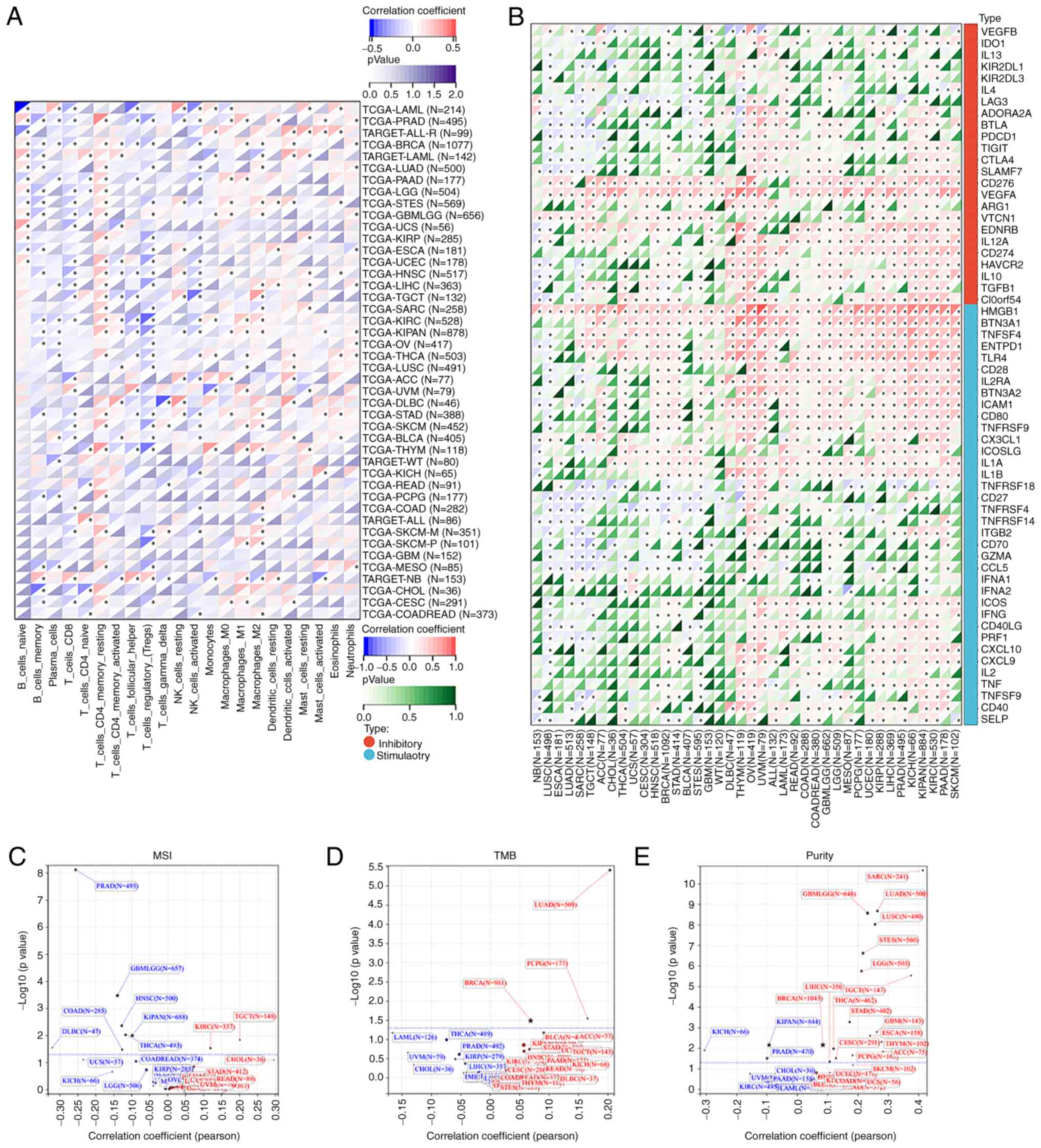
Furthermore, the correlation between PEX13
expression level and ICP genes, MSI, TMB and tumor purity were
analyzed via SangerBox. The occurrence of MSI is caused by the
functional defect of DNA mismatch repair in tumor tissues (34). The presence of MSI and ineffective
DNA mismatch repair is a crucial clinical tumor marker (35). TMB has a strong relationship to the
effectiveness of PD-1/PD-L1 inhibitors and can forecast their
benefits in terms of efficacy (36). The present study discovered that
PEX13 expression was positively linked with ICP genes in several
tumor types (Fig. 8B). PEX13
expression was positively correlated with MSI in TGCT and KIRC and
negatively correlated with MSI in GBMLGG, HNSC, PRAD, KIPAN, COAD,
DLBC and THCA (Fig. 8C). In LUAD
and BRCA and pheochromocytoma and paraganglioma, PEX13 expression
was linked favorably with TMB (Fig.
8D). In tumor purity, PEX13 expression was negatively related
to purity in KICH, KIPAN and PRAD and positively related to purity
in GBMLGG, SARC, LUAD, LUSC, STES, LGG, STAD, BRCA, ESCA, etc.
(Fig. 8E). Moreover, it was
evaluated the relationship between the ESTIMATE score and PEX13
expression level in various cancers. ImmuneScore reflects the
proportion of infiltrating immune cells in cancer tissues. The
proportion of stromal cells in tumor tissue was reflected by the
Stromal score (StromalScore). The ESTIMATE score (ESTIMATEScore) is
the sum of immune and stromal scores and reflects the state of the
TIM and tumor purity (18,37). The present study found significant
relationships between PEX13 levels and StromalScore, ImmuneScore
and ESTIMATEScore across multiple tumors, especially in LUAD, STES,
SARC and LUSC (Fig. S5A-C). These
findings indicated that PEX13 expression regulated the sensitivity
and resistance of certain cancers to immunotherapy. Thus, PEX13
emerged from this analysis as a potential immunotherapy biomarker
and predictor of tumor immunotherapy response.
PEX13 promoted cell proliferation,
migration, invasion and colony formation in PAAD
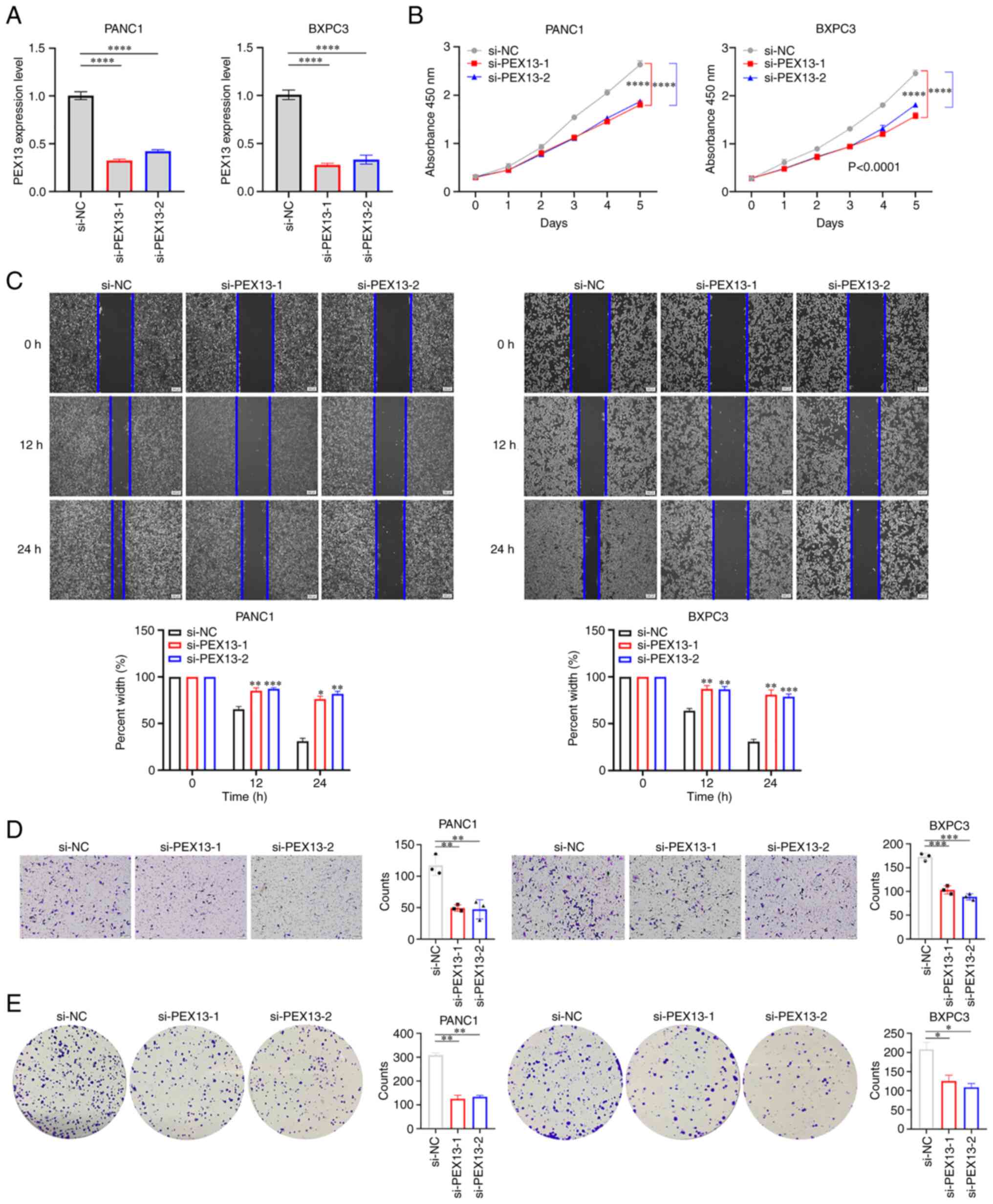
PANC1 and BxPC3 cell lines were used to verify the
biological functions of PEX13. The knockdown of PEX13 expression in
PANC1 and BxPC3 cells transfected with PEX13 siRNAs was confirmed
using RT-qPCR (Fig. 9A). The CCK-8
assay suggested that PEX13 knockdown inhibited the cell
proliferation of PANC1 and BxPC3 cells in vitro (Fig. 9B). In addition, wound healing and
Transwell experiments demonstrated that PEX13 knockdown suppressed
the cell migration and invasion of PANC1 and BxPC3 cells in
vitro (Fig. 9C-D). The colony
formation assay suggested that PEX13 knockdown could inhibit the
cell colony formation ability of PANC1 and BxPC3 cells in
vitro (Fig. 9E). These findings
showed that PEX13 could increase cell proliferation, migration,
invasion and colony formation abilities of PANC1 and BxPC3 cell
lines in vitro.
Discussion
Metabolic reprogramming is one of the hallmarks of
cancer and has a selective advantage in the occurrence and
progression of cancer cells (38).
Metabolites ingested through cell membranes are mostly initially
processed in the cytoplasm and then typically transported to
intracellular organelles for further processing, such as peroxisome
and mitochondria (39). Although
mitochondria are key metabolic hubs of cancer cells, the metabolic
functions of other organelles in cancer cells are rarely studied
and need further exploration.
In mammals, the peroxisome, a single membrane
organelle, participates in >50 separate enzymatic functions
(40). Defects in genes encoding
peroxisome proteins are associated with peroxisome disease. Lipid
metabolism is also critical to tumorigenicity (8). Lipid synthesis pathways (such as the
production of lipogenic triglycerides and the synthesis of
phospholipids and cholesterol) are critical for energy storage,
cell membrane structure and cell signaling regulation. For lipid
synthesis and activation of the corresponding signaling network to
be coordinated in rapidly growing cancer cells, a significant
volume of cell membrane formation is required (41). High levels of ether phospholipid in
certain cancers suggest that elevated lipid synthesis of peroxisome
is related to tumor progression (42). PEX13 is an indispensable key
molecule in the occurrence of peroxisome, but, to the best of our
knowledge, its functions and effects in tumors have not been
reported in the literature. Therefore, the present study aimed to
investigate the biological functions and clinical significance of
PEX13 in various tumors, especially in PAAD.
The present study first revealed that PEX13 was
upregulated in several types of tumors compared with the
corresponding normal tissues, including GBM, LGG, GBMLGG, UCEC,
CESC, BRCA, ESCA, LUAD, STES, COADREAD, COAD, PRAD, STAD, LUSC,
HNSC, WT, LIHC, THCA, BLCA, PAAD, UCS, ALL, LAML and ACC. In ACC,
KICH, LGG, LIHC and PAAD, patients with high PEX13 mRNA expression
had significantly lower OS, and patients with high PEX13 mRNA
expression had significantly higher OS in KIRC. Cox regression
analysis indicated that PEX13 mRNA was related to OS, DFI, PFI and
DSS in various tumors. In addition, high expression of PEX13 was
associated with poorer prognosis in patients with PAAD.
Subsequently, genetic alteration analysis of PEX13 showed the
presence of PEX13 gene mutations, amplification and deletion in
multiple tumors. The PAAD tissues displayed different somatic
mutations and CNVs depending on the level of PEX13 expression,
indicating that the PEX13 gene alteration may mediate the
occurrence and progression of multiple tumors, especially in PAAD.
The GO-BP and KEGG enrichment analyses of PEX13-related genes found
that PEX13 expression was significantly correlated with
endocytosis, mRNA surveillance pathway, nucleocytoplasmic
transport, T cell receptor signaling pathway and hippo signaling
pathway in cancers. In PAAD, GO-BP and KEGG enrichment analyses of
PEX13-related genes revealed that PEX13 expression was
significantly associated with Rap1, ErbB, Neurotrophin, AMPK,
Apelin and Insulin signaling pathways. WGCNA analysis suggested
that PEX13-related hub genes were mainly enriched in PI3K-Akt
signaling pathway, ECM-receptor interaction, focal adhesion and
MAPK signaling pathway via KEGG enrichment analysis. Moreover, the
present study discovered that PEX13 may interact with PEX2, PEX12,
PEX14 and ABCD3 to influence the biological functions and serve as
a significant factor in pan-cancers and the upstream
lncRNA/circRNA-miRNA regulatory networks may regulate the aberrant
expression of PEX13 in various cancers.
Furthermore, PEX13-related genes were mainly
enriched in immune and cancer-related pathways, and the correlation
between PEX13 and tumor immunity was further analyzed. As an
important component of the immune microenvironment, TIICs act as a
key factor in the occurrence, progression and treatment of tumors.
PEX13 mRNA levels were associated with TIICs in multiple tumors,
including PAAD. The present study evaluated the effect of PEX13 on
immunotherapy sensitivity and resistance in patients with cancer.
The ICP, MSI, TMB and tumor purity analyses suggested that PEX13
may be a promising target for the treatment of patients with
certain tumors, especially in immunotherapy. In vitro assays
also demonstrated that PEX13 knockdown could significantly inhibit
the proliferation, migration, invasion and colony formation of PAAD
cells.
In conclusion, the present results demonstrated that
PEX13 may be an invaluable prognostic indicator and a possible
predictor of immunotherapy sensitivity and resistance in patients
with malignant tumors, especially in PAAD. However, the present
study has limitations because it only uses TCGA database, while
further databases should be used for corroboration, and only
verifies the relationship between PEX13 and PAAD. The possible
mechanisms by which PEX13 functions in PAAD were not further
explored.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of Shanxi Province (grant no. 202103021223011).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JS designed the study. PD and XD performed the
bioinformatics analyses and experiments. TY and DL analyzed the
results. YW and YD performed data analysis and wrote the
manuscript. JS and PD confirm the authenticity of all the raw data.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ACC
|
adrenocortical carcinoma
|
|
BLCA
|
bladder urothelial carcinoma
|
|
BRCA
|
breast invasive carcinoma
|
|
CESC
|
cervical squamous cell carcinoma and
endocervical adenocarcinoma
|
|
CHOL
|
cholangiocarcinoma
|
|
CNA
|
copy number alteration
|
|
COAD
|
colon adenocarcinoma
|
|
DLBC
|
lymphoid neoplasm diffuse large b-cell
lymphoma
|
|
ESCA
|
esophageal carcinoma
|
|
GTEx
|
Genotype-Tissue Expression
|
|
GBM
|
glioblastoma multiforme
|
|
GEPIA2
|
Gene Expression Profiling Interactive
Analysis 2
|
|
HGG
|
brain higher grade glioma
|
|
HNSC
|
head and neck cancer
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
KICH
|
kidney chromophobe
|
|
KIRC
|
kidney renal clear cell carcinoma
|
|
KIPAN
|
Pan-kidney cohort
|
|
ICP
|
immune checkpoint
|
|
LAML
|
acute myeloid leukemia
|
|
LGG
|
brain lower grade glioma
|
|
LIHC
|
liver hepatocellular carcinoma
|
|
LUAD
|
lung adenocarcinoma
|
|
LUSC
|
lung squamous cell carcinoma
|
|
MESO
|
mesothelioma
|
|
MSI
|
microsatellite instability
|
|
OS
|
overall survival
|
|
OV
|
ovarian serous cystadenocarcinoma
|
|
PAAD
|
pancreatic adenocarcinoma
|
|
PCPG
|
pheochromocytoma and paraganglioma
|
|
PEX
|
peroxisomal biogenesis factor
|
|
PRAD
|
prostate adenocarcinoma
|
|
READ
|
rectum adenocarcinoma
|
|
SARC
|
sarcoma
|
|
SKCM
|
skin cutaneous melanoma
|
|
STAD
|
stomach adenocarcinoma
|
|
TCGA
|
The Cancer Genome Atlas
|
|
TGCT
|
testicular germ cell tumors
|
|
THCA
|
thyroid carcinoma
|
|
THYM
|
thymoma
|
|
TMB
|
tumor mutation burden
|
|
UCEC
|
uterine corpus endometrial
carcinoma
|
|
UCS
|
uterine carcinosarcoma
|
|
UVM
|
uveal melanoma
|
|
WGCNA
|
weighted gene co-expression network
analysis
|
References
|
1
|
Fujiki Y, Okumoto K, Mukai S, Honsho M and
Tamura S: Peroxisome biogenesis in mammalian cells. Front Physiol.
5:3072014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee MY, Sumpter RJ, Zou Z, Sirasanagandla
S, Wei Y, Mishra P, Rosewich H, Crane DI and Levine B: Peroxisomal
protein PEX13 functions in selective autophagy. EMBO Rep. 18:48–60.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang J, Kim J, Alexander A, Cai S,
Tripathi DN, Dere R, Tee AR, Tait-Mulder J, Di Nardo A, Han JM, et
al: A tuberous sclerosis complex signalling node at the peroxisome
regulates mTORC1 and autophagy in response to ROS. Nat Cell Biol.
15:1186–1196. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dixit E, Boulant S, Zhang Y, Lee AS,
Odendall C, Shum B, Hacohen N, Chen ZJ, Whelan SP, Fransen M, et
al: Peroxisomes are signaling platforms for antiviral innate
immunity. Cell. 141:668–681. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sugiura A, Mattie S, Prudent J and Mcbride
HM: Newly born peroxisomes are a hybrid of mitochondrial and
ER-derived pre-peroxisomes. Nature. 542:251–254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Smith JJ and Aitchison JD: Peroxisomes
take shape. Nat Rev Mol Cell Biol. 14:803–817. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jain IH, Calvo SE, Markhard AL, Skinner
OS, To TL, Ast T and Mootha VK: Genetic screen for cell fitness in
high or low oxygen highlights mitochondrial and lipid metabolism.
Cell. 181:716–727. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Q, Luo Q, Halim A and Song G:
Targeting lipid metabolism of cancer cells: A promising therapeutic
strategy for cancer. Cancer Lett. 401:39–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Di Cara F, Savary S, Kovacs WJ, Kim P and
Rachubinski RA: The peroxisome: An up-and-coming organelle in
immunometabolism. Trends Cell Biol. 33:70–86. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Diskin C, Zotta A, Corcoran SE, Tyrrell
VJ, Zaslona Z, O'Donnell VB and O'Neill L: 4-Octyl-Itaconate and
Dimethyl Fumarate inhibit COX2 expression and prostaglandin
production in macrophages. J Immunol. 207:2561–2569. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wanders RJ and Waterham HR: Biochemistry
of mammalian peroxisomes revisited. Annu Rev Biochem. 75:295–332.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ding L, Sun W, Balaz M, He A, Klug M,
Wieland S, Caiazzo R, Raverdy V, Pattou F, Lefebvre P, et al:
Peroxisomal beta-oxidation acts as a sensor for intracellular fatty
acids and regulates lipolysis. Nat Metab. 3:1648–1661. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee JY, Plakidas A, Lee WH, Heikkinen A,
Chanmugam P, Bray G and Hwang DH: Differential modulation of
Toll-like receptors by fatty acids: Preferential inhibition by n-3
polyunsaturated fatty acids. J Lipid Res. 44:479–486. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moreno-Fernandez ME, Giles DA, Stankiewicz
TE, Sheridan R, Karns R, Cappelletti M, Lampe K, Mukherjee R, Sina
C, Sallese A, et al: Peroxisomal beta-oxidation regulates whole
body metabolism, inflammatory vigor, and pathogenesis of
nonalcoholic fatty liver disease. JCI Insight. 3:e936262018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Demers ND, Riccio V, Jo DS, Bhandari S,
Law KB, Liao W, Kim C, Mcquibban GA, Choe SK, Cho DH, et al: PEX13
prevents pexophagy by regulating ubiquitinated PEX5 and peroxisomal
ROS. Autophagy. 19:1781–1802. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen XF, Tian MX, Sun RQ, Zhang ML, Zhou
LS, Jin L, Chen LL, Zhou WJ, Duan KL, Chen YJ, et al: SIRT5
inhibits peroxisomal ACOX1 to prevent oxidative damage and is
downregulated in liver cancer. Embo Rep. 19:e451242018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ahn YH, Yang Y, Gibbons DL, Creighton CJ,
Yang F, Wistuba II, Lin W, Thilaganathan N, Alvarez CA, Roybal J,
et al: Map2k4 functions as a tumor suppressor in lung
adenocarcinoma and inhibits tumor cell invasion by decreasing
peroxisome proliferator-activated receptor gamma2 expression. Mol
Cell Biol. 31:4270–4285. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wei C, Wang B, Peng D, Zhang X, Li Z, Luo
L, He Y, Liang H, Du X, Li S, et al: Pan-cancer analysis shows that
ALKBH5 is a potential prognostic and immunotherapeutic biomarker
for multiple cancer types including gliomas. Front Immunol.
13:8495922022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang Z, Kang B, Li C, Chen T and Zhang Z:
GEPIA2: An enhanced web server for large-scale expression profiling
and interactive analysis. Nucleic Acids Res. 47:W556–W560. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Navani S: Manual evaluation of tissue
microarrays in a high-throughput research project: The contribution
of Indian surgical pathology to the Human Protein Atlas (HPA)
project. Proteomics. 16:1266–1270. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu P, Heins ZJ, Muller JT, Katsnelson L,
de Bruijn I, Abeshouse AA, Schultz N, Fenyo D and Gao J:
Integration and analysis of CPTAC proteomics data in the context of
cancer genomics in the cBioPortal. Mol Cell Proteomics.
18:1893–1898. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liefeld T, Reich M, Gould J, Zhang P,
Tamayo P and Mesirov JP: GeneCruiser: A web service for the
annotation of microarray data. Bioinformatics. 21:3681–3682. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mayakonda A, Lin DC, Assenov Y, Plass C
and Koeffler HP: Maftools: Efficient and comprehensive analysis of
somatic variants in cancer. Genome Res. 28:1747–1756. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li T, Fu J, Zeng Z, Cohen D, Li J, Chen Q,
Li B and Liu XS: TIMER2.0 for analysis of tumor-infiltrating immune
cells. Nucleic Acids Res. 48:W509–W514. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vasaikar SV, Straub P, Wang J and Zhang B:
LinkedOmics: Analyzing multi-omics data within and across 32 cancer
types. Nucleic Acids Res. 46:D956–D963. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yan Y, Zhang D, Zhou P, Li B and Huang SY:
HDOCK: A web server for protein-protein and protein-DNA/RNA docking
based on a hybrid strategy. Nucleic Acids Res. 45:W365–W373. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei C, Zhang X, Peng D, Zhang X, Guo H, Lu
Y, Luo L, Wang B, Li Z, He Y, et al: LncRNA HOXA11-AS promotes
glioma malignant phenotypes and reduces its sensitivity to ROS via
Tpl2-MEK1/2-ERK1/2 pathway. Cell Death Dis. 13:9422022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martincorena I, Raine KM, Gerstung M,
Dawson KJ, Haase K, Van Loo P, Davies H, Stratton MR and Campbell
PJ: Universal patterns of selection in cancer and somatic tissues.
Cell. 171:1029–1041. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Luo Y, Zheng S, Wu Q, Wu J, Zhou R, Wang
C, Wu Z, Rong X, Huang N, Sun L, et al: Long noncoding RNA (lncRNA)
EIF3J-DT induces chemoresistance of gastric cancer via autophagy
activation. Autophagy. 17:4083–4101. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wei Y, Lu C, Zhou P, Zhao L, Lyu X, Yin J,
Shi Z and You Y: EIF4A3-induced circular RNA ASAP1 promotes
tumorigenesis and temozolomide resistance of glioblastoma via
NRAS/MEK1/ERK1-2 signaling. Neuro Oncol. 23:611–624. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Baxevanis CN, Fortis SP and Perez SA: The
balance between breast cancer and the immune system: Challenges for
prognosis and clinical benefit from immunotherapies. Semin Cancer
Biol. 72:76–89. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen Y, Jia K, Sun Y, Zhang C, Li Y, Zhang
L, Chen Z, Zhang J, Hu Y, Yuan J, et al: Predicting response to
immunotherapy in gastric cancer via multi-dimensional analyses of
the tumour immune microenvironment. Nat Commun. 13:48512022.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Baretti M and Le DT: DNA mismatch repair
in cancer. Pharmacol Ther. 189:45–62. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin A, Zhang J and Luo P: Crosstalk
between the MSI status and tumor microenvironment in colorectal
cancer. Front Immunol. 11:20392020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luchini C, Bibeau F, Ligtenberg M, Singh
N, Nottegar A, Bosse T, Miller R, Riaz N, Douillard JY, Andre F, et
al: ESMO recommendations on microsatellite instability testing for
immunotherapy in cancer, and its relationship with PD-1/PD-L1
expression and tumour mutational burden: A systematic review-based
approach. Ann Oncol. 30:1232–1243. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu W, Wang X, Le W, Lu C, Li H, Zhu Y,
Chen X, An W, Xu C, Wu Q, et al: Immune microenvironment
infiltration landscape and immune-related subtypes in prostate
cancer. Front Immunol. 13:10012972022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Deberardinis RJ and Chandel NS:
Fundamentals of cancer metabolism. Sci Adv. 2:e16002002016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Porporato PE, Filigheddu N, Pedro J,
Kroemer G and Galluzzi L: Mitochondrial metabolism and cancer. Cell
Res. 28:265–280. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wanders RJ and Waterham HR: Biochemistry
of mammalian peroxisomes revisited. Annu Rev Biochem. 75:295–332.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Beloribi-Djefaflia S, Vasseur S and
Guillaumond F: Lipid metabolic reprogramming in cancer cells.
Oncogenesis. 5:e1892016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dean JM and Lodhi IJ: Structural and
functional roles of ether lipids. Protein Cell. 9:196–206. 2018.
View Article : Google Scholar : PubMed/NCBI
|