Introduction
Lin28 is an RNA-binding protein that regulates key
cellular processes, including cell development, glucose metabolism,
differentiation, pluripotency and stem cell self-renewal (1–3). Lin28
has been identified as a central factor in reprogramming mammalian
somatic cells to a pluripotent state and maintaining pluripotency
(4,5). Additionally, Lin28 overexpression in
cancer is associated with more undifferentiated and aggressive
tumor phenotypes as well as resistance to conventional therapies
(6,7).
There are two paralogs of the Lin28 gene in
vertebrates, Lin28A and Lin28B, which share high DNA sequence
homology (8). Both paralog proteins
negatively regulate the tumor suppressor micro (mi)RNA Let-7
(3,9,10). The
present study assessed the effects of resveratrol (RSV) on Lin28A
protein stability using the NCCIT cell line, which overexpressed
the Lin28A protein (11).
The mechanisms underlying the post-translational
overexpression and stabilization of Lin28A have been previously
described. In 2017, Tsanov et al (12) demonstrated that Lin28A is
phosphorylated at serine 200 (S200) by the MAPK/ERK pathway in
pluripotent stem cells, leading to increased expression through
post-translational stabilization. Additionally, in 2019, Haq et
al (11) reported that Lin28A
undergoes ubiquitination, destabilization and degradation through
the 26S proteasome pathway in human embryonal carcinoma cells.
However, the deubiquitinase ubiquitin-specific protease 28 (USP28)
was reported to reverse Lin28A ubiquitination, preventing its
degradation and increasing stability. In contrast, several studies
have reported that therapeutic inhibition, genetic inactivation or
silencing of Lin28 reverses the invasive phenotype in cancer cells
(13,14). Together, these findings provide
insight into potential molecular mechanisms regulating Lin28A
stability at the post-translational level.
Extensive research has documented the anticancer
effects of RSV and its ability to modulate key signaling pathways
in several cancer cell types (15).
Regarding the Lin28A oncoprotein, a luciferase assay in one study
demonstrated that RSV decreased Lin28A gene expression in
colorectal cancer cells (16).
However, the impact of RSV on Lin28A protein stability is yet to be
elucidated, and understanding the regulatory mechanisms of
oncogenes such as Lin28A is essential for developing improved
cancer treatments. Furthermore, a deeper understanding of how
therapies modulate oncogenic drivers could enable more targeted and
effective combination strategies.
Natural compounds, such as polyphenols,
phytosterols, triterpenoids and saponins, also show promise when
combined with conventional cancer treatments (17). Used together, they can enhance
anticancer impacts, reduce side effects, boost immunity and promote
cancer cell death (18). RSV in
particular has demonstrated mixed benefits (19,20)
and further investigation in to the effects of RSV on the Lin28A
oncoprotein may reveal new therapeutic opportunities. For example,
downregulating critical cancer drivers like Lin28A through
pharmaceutical or dietary interventions represents an attractive
strategy, and expanding the knowledge of the molecular targets of
RSV could support its inclusion in rational, multitargeted regimens
designed to maximize clinical impact.
Materials and methods
Reagents and antibodies
Resveratrol (RSV) was purchased from Sigma-Aldrich
(Merck KGaA; cat. no. R5010) and suspended in ethanol absolute
(vehicle). MAPK/ERK inhibitor PD0325901 was purchased from
Sigma-Aldrich (Merck KGaA; cat. no. PZ0162) and suspended in DMSO.
Proteasomal inhibitor-MG132 (cat. no. M8699) and cycloheximide
(CHX; cat. no. 01810) were also purchased from Sigma-Aldrich (Merck
KGaA). Monoclonal anti-Lin28-HRP antibodies (cat. no. sc-293120)
were purchased from Santa Cruz Biotechnology, Inc. Monoclonal
anti-β-actin-peroxidase antibodies (cat. no. A3854) were purchased
from Sigma-Aldrich (Merck KGaA). Polyclonal
anti-phosphorylated-Lin28A (pLin28) (Ser200) antibodies (cat. no.
PA5-105696) were purchased from Invitrogen (Thermo Fisher
Scientific, Inc.). Mouse anti-rabbit IgG-HPR antibodies (cat. no.
sc-2357) were purchased from Santa Cruz Biotechnology, Inc.
Polyclonal anti-USP28 HRP antibodies (cat. no. orb480432) were
purchased from Biorbyt, Ltd.
Cell culture
The pluripotent embryonal carcinoma (testicular
teratocarcinoma) NCCIT cell line was purchased from the American
Type Culture Collection (cat. no. CRL-2073). This cell line was
cultured in RPMI 1640 medium (Biowest SAS) supplemented with 10%
fetal bovine serum (cat. no. S1810-500; Biowest SAS) at 37°C in a
humidified atmosphere containing 5% CO2.
Cell viability assay
Cell viability was assessed using the MTT assay,
which measures cellular metabolic activity as an indicator of
viability. Cells were seeded at a density of 8×104
cells/well in 24-well plates and cultured overnight at 37°C. The
following day, the cells were treated with several concentrations
of RSV (0, 25, 50, 100, 150 and 200 µM). After 48 h of treatment,
MTT reagent (Sigma-Aldrich; Merck KGaA) was added to each well at a
final concentration of 0.5 mg/ml and incubated at 37°C for 30 min
to allow formazan crystal formation by metabolically active cells.
The supernatant was then removed and the formazan crystals were
dissolved in 500 µl acidified isopropanol. Absorbance was measured
at 570 nm using a Tecan Sunrise™ absorbance microplate reader
(Tecan Group, Ltd.). Cell viability was expressed as a percentage
relative to vehicle-treated control cells, which were set at 100%.
Each treatment was performed in triplicate and the experiment was
repeated three times.
To assess the viability of cells treated with the
ERK inhibitor PD0325901, the MTT assay was performed following the
aforementioned procedure, with cells exposed to a range of
PD0325901 concentrations (0, 1, 2, 5, 7.5 and 10 µM).
Treatment of cells with RSV and ERK
inhibitor
For RSV treatments, the following doses were used in
the experiments: 25, 50, 100 and 150 µM. The 150 µM dose, which was
close to the calculated half-maximal inhibitory concentration
(IC50), was a particular focus of the analysis due to
the strong decrease in the Lin28A oncoprotein with this treatment
dose. For ERK inhibitor treatments, doses close to or below the
IC50 were tested, specifically 5 and 7.5 µM. The
concentration of 7.5 µM was revealed to be the most effective in
inhibiting Lin28A phosphorylation at S200.
For the vehicle controls, the amount of absolute
ethanol corresponding to the highest RSV concentration tested was
used, which was 150 µM. This ethanol vehicle was suspended in the
cell culture medium (RPMI 1640 medium supplemented with 10% fetal
bovine serum) at a final concentration of 0.18% by adding 1.8 µl of
ethanol per ml of medium.
Similarly, for the ERK inhibitor treatments, the
highest concentration used was 7.5 µM, which was suspended in DMSO.
This resulted in a final DMSO concentration of 0.05% in the medium,
achieved by adding 0.5 µl of DMSO per ml of medium.
In prior experiments, the present study assessed the
maximum tolerable concentrations of DMSO and ethanol in MTT assays
performed on untreated or vehicle-free control cells, to ensure
that these solvents did not alter the proliferation of the cells
under study.
Small interfering (si)RNA
transfection
NCCIT cells were transfected with pre-designed
siRNAs targeting USP28 (cat. nos. 4392420, s33508, s33509 and
s33510) or a non-targeting control siRNA (cat. no. 4390843); all
purchased from Ambion® (Thermo Fisher Scientific, Inc.).
The siUSP28 sequences are provided in Table I. siRNAs were transfected at a
concentration of 2 nmol/ml using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol for 24 h at 37°C. After 24 h, cells were
lysed and protein extracts were collected. Western blot analysis
was performed to assess the silencing of USP28 protein expression
after transfection with siRNAs targeting USP28 compared with the
non-targeting control. siRNA concentrations were optimized in
separate experiments to achieve maximum USP28 knockdown without
inducing cytotoxicity, as determined using previously established
criteria in the instructions provided by Ambion.
 | Table I.Sequences of select pre-designed
small-interfering ubiquitin-specific protease 28. |
Table I.
Sequences of select pre-designed
small-interfering ubiquitin-specific protease 28.
| siRNA | Strand | Sequence
(5′-3′) |
|---|
| 1 | Sense |
GAUUAUAGUUUGUUCCGAAtt |
|
| Antisense |
UUCGGAACAAACUAUAAUCtt |
| 2 | Sense |
GUGAUUGCUUUAUACCGAAtt |
|
| Antisense |
UUCGGUAUAAAGCAAUCACgg |
| 3 | Sense |
GGCCUAGAACUCUAUCAAAtt |
|
| Antisense |
UUUGAUAGAGUUCUAGGCCtg |
Reverse transcription-quantitative PCR
(RT-qPCR)
Cells were seeded at a density of 8×104
cells/dish in p60 culture dishes and incubated for 24 h at 37°C
prior to treatment. Following treatment, total RNA was extracted
from cells using TRIzol® (cat. no. 15596026; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. RNA integrity and concentration were determined by
agarose gel analysis and quantified using the Thermo Scientific™
NanoDrop™ One/OneC (Thermo Fisher Scientific, Inc.).
Complementary (c)DNA was synthesized from 1 µg total RNA using the
First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol; the incubation time was 1
h at 37°C and 5 min at 70°C. qPCR was then performed using the 7300
Real-Time PCR System (Applied Biosystems; Thermo Fisher Scientific,
Inc.) using iTaq Universal SYBR® Green Supermix (Bio-Rad
Laboratories, Inc.). PCR thermocycling conditions were as follows:
1 cycle at 94°C for 10 min for initial denaturation, 35 repetitions
of a 10-sec denaturation step at 94°C, a 30-sec annealing step at
60°C and a 30 sec extension step at 72°C. Final extension was at
72°C for 5 min. The specificity of each PCR was assessed using the
melting temperature profiles of the final products. Assays were
performed in triplicate and the relative expression of target genes
was normalized to the reference gene β-2 microglobulin (β2M) using
the 2−ΔΔCq method (21).
The pre-designed primers, Lin28A, USP28 and β2M were purchased from
Integrated DNA Technologies, Inc. and the sequences are provided in
Table II.
| Table II.Sequences of primers used for reserve
transcription-quantitative PCR. |
Table II.
Sequences of primers used for reserve
transcription-quantitative PCR.
| Gene | Direction | Sequence
(5′-3′) |
|---|
| Lin28A | F |
CATCTGTAAGTGGTTCAACGTG |
|
| R |
CCCTTCCATGTGCAGCTTA |
| USP28 | F |
TGGCCAGGCTGATCTCTAACTC |
|
| R |
GAAGGCCGGGTACGATGA |
| β2M | F |
GGACTGGTCTTTCTATCTCTTGT |
|
| R |
ACCTCCATGATGCTGCTTAC |
Western blotting
Cells were seeded at a density of 8×104
cells/dish in p60 cell culture dishes and incubated for 24 h at
37°C prior the treatment. Following treatment, cells were lysed in
RIPA buffer (150 mM NaCl; 0.5% sodium deoxycholate; 0.1% SDS; 50 mM
Tris; pH 7.4) supplemented with 1X complete™, Mini, EDTA-free
Protease Inhibitor Cocktail (cat. no. 11836170001; Sigma-Aldrich;
Merck KGaA) containing phosphatase inhibitors (1 mM sodium fluoride
and 0.5 mM sodium orthovanadate; Sigma-Aldrich; Merck KGaA). Cell
lysates were sonicated for 1 min and were centrifugated at 16,800 ×
g and 4°C for 5 min to collect supernatants containing proteins of
interest. Protein integrity and concentration were determined on a
polyacrylamide gel using Coomassie staining and were quantified
using the NanoDrop One/OneC. Equal amounts of protein
(30 µg) were separated by 12% (w/v) SDS-PAGE and were transferred
onto nitrocellulose membranes (cat. no. 1620115; Bio-Rad
Laboratories, Inc.). Membranes were blocked for 1 h at room
temperature with 5% skimmed milk or 5% BSA (cat. no. 30063721;
Gibco; Thermo Fisher Scientific, Inc.) for phosphorylated protein
detection. Subsequently, the membranes were incubated overnight at
4°C with primary antibodies against β-actin (1:20,000), pLin28A
(S200; 1:1,000), Lin28A (1:750) and USP28 (1:1,000). Membranes were
washed with TBS-0.05% Tween (cat. no. P7949; Sigma-Aldrich; Merck
KGaA) and incubated with the secondary anti-rabbit antibodies
(1:20,000) for 1 h at room temperature in the case of pLin28A. For
the other antibodies, it was not necessary to use a secondary
antibody since the primary antibodies were coupled to HRP. Protein
bands were detected using the SuperSignal™ West Pico PLUS
Chemiluminescent Substrate (Thermo Fisher Scientific, Inc.) and
quantified by densitometry using Image Studio™ Lite 5.2 software
(LI-COR Biosciences). Band intensity was normalized to β-actin as a
loading control. Western blots shown are representative of ≥3
independent experiments.
Statistical analysis
All statistical analyses including correlation and
half-life tests were performed using GraphPad Prism software
(version 8; Dotmatics). Differences between experimental groups
were analyzed using one-way ANOVA, followed by Tukey's multiple
comparisons test, as appropriate. The unpaired Student's t-test was
used for comparisons between two groups. A two-way ANOVA followed
by Tukey's multiple comparisons test was used for a control
analysis of the Pearson's correlation coefficient analysis as 2
time points and 2 concentrations were evaluated. Data are presented
as the mean ± standard deviation. P<0.05 was considered to
indicate a statistically significant difference.
Results
RSV decreases the cell viability of
NCCIT cells
To assess the effect of increasing doses of RSV
(25–200 µM) on NCCIT teratocarcinoma cell proliferation, MTT assays
were performed. The results demonstrated that RSV exerted a marked
dose-dependent inhibition of NCCIT cell viability (Fig. S1). Analysis of the data revealed
that the IC50 of RSV in NCCIT cells was 148 µM. Based on
this finding, the subsequent experiments were restricted to a dose
range of 25–150 µM RSV, which encompassed concentrations below and
near the IC50 to adequately assess
concentration-dependent responses. In summary, this initial
analysis established the ability of RSV to suppress NCCIT cell
viability in a dose-dependent manner and informed the selection of
appropriate treatment doses for further mechanistic evaluation in
the present study.
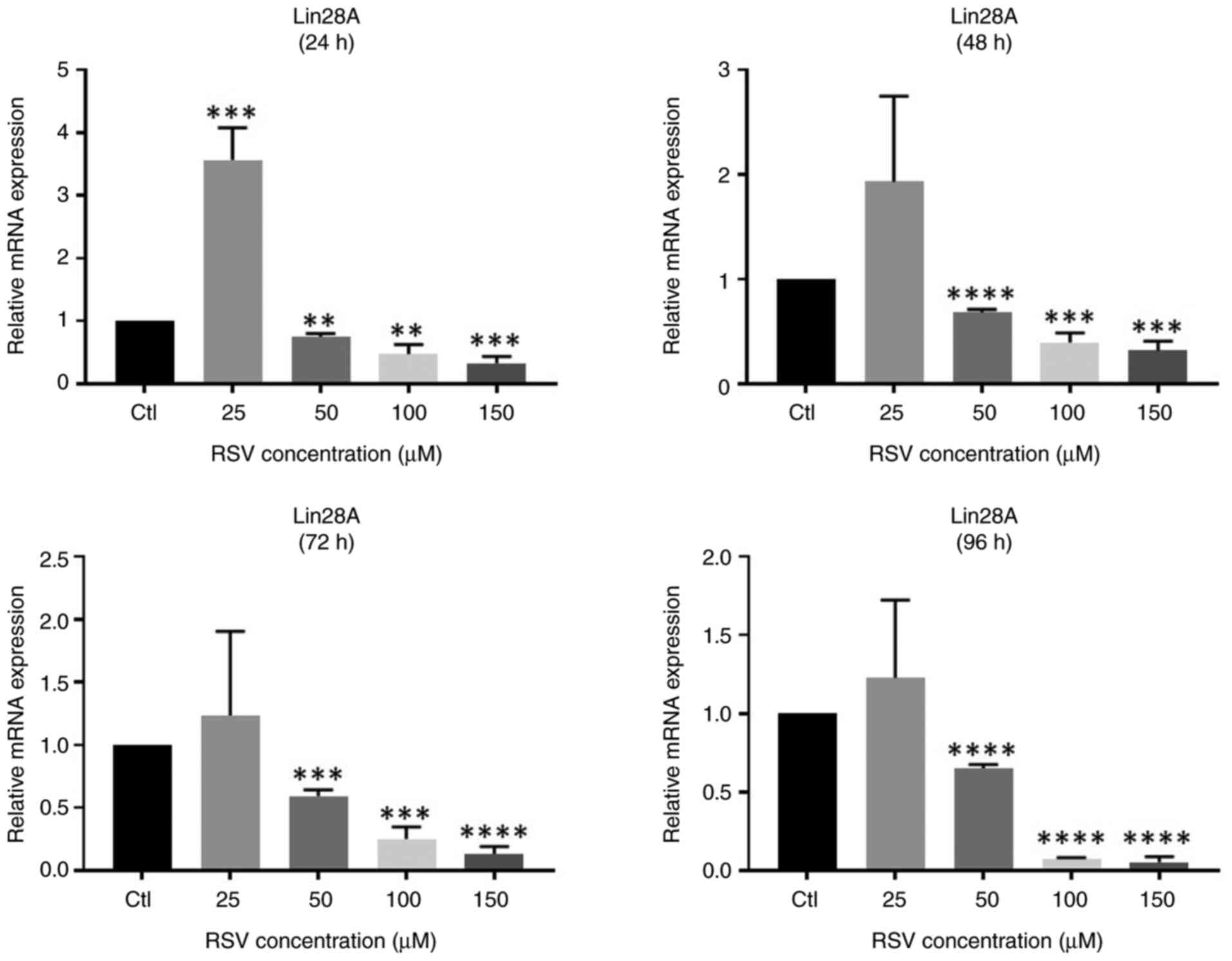
RSV decreases the mRNA expression of
Lin28A in NCCIT cells
The effect of RSV on Lin28A mRNA expression in NCCIT
was evaluated using RT-qPCR. Notably, the results revealed that
treatment with the lowest RSV concentration (25 µM) was associated
with a significant increase in Lin28A mRNA expression levels.
However, further increases (≥50 µM) in the RSV dose were associated
with significant decreases in Lin28A mRNA expression in a
concentration-dependent manner (Fig.
1). These results indicate that higher RSV doses downregulate
Lin28A expression at the transcriptional level. The present study
then sought to assess whether this inhibitory effect persisted at
the protein level.
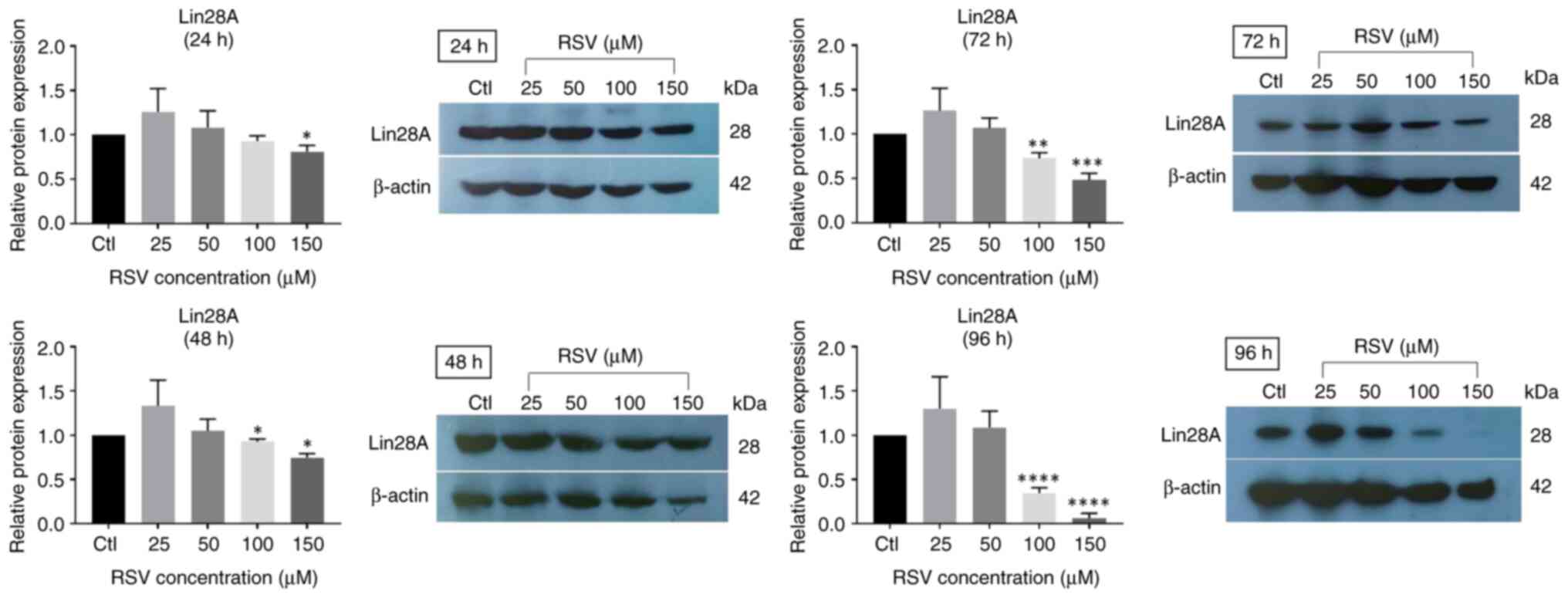
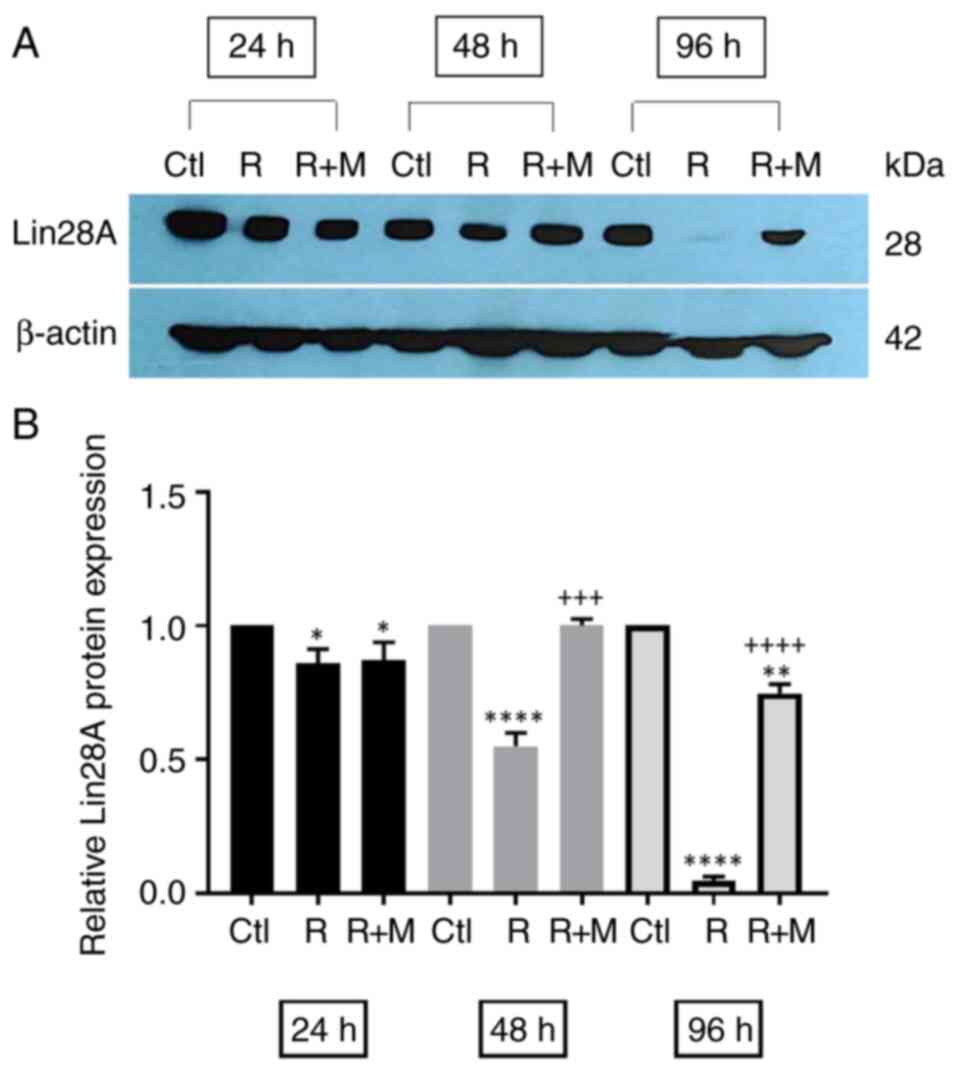
RSV at high concentrations decreases
Lin28A protein expression levels in NCCIT cells
The effect of RSV on the expression of the Lin28A
protein was evaluated in NCCIT cells using treatment with RSV at
different concentrations (25, 50, 100 and 150 µM). Western blot
analysis was performed and the results demonstrated that low
concentrations of RSV (25 and 50 µM) had no significant effect on
the Lin28A protein expression level. However, the protein
expression of Lin28A began to significantly decrease compared with
in the control group in response to 100 and 150 µM RSV; with a more
pronounced decrease in Lin28A protein detected at 72 and 96 h
compared with at earlier time points (Fig. 2).
Pearson's correlation coefficient analysis was
performed between Lin28A mRNA and protein expression. A total of
two time points and two concentrations were chosen: i) 48 h, as in
the IC50, and ii) 96 h, corresponding to the maximum
effect of RSV over Lin28A protein. Low (50 µM) and high (150 µM)
concentrations of RSV were also chosen. The results demonstrated
that there was a strong negative correlation at 48 h for 50 and 150
µM (r=−0.982 and r=−0.8841, respectively); however, this was not
significant. This indicates that, for both concentrations (50 and
150 µM), mRNA expression was downregulated, while protein was
upregulated or slightly downregulated. This demonstrated that RSV
can produce a hormetic effect (dose-response phenomenon) for both
mRNA and protein. Notably, this hormetic effect varies between
transcript and protein at the same dose. This discrepancy can be
attributed to the high stability of the Lin28A protein and the
distinct mechanisms, such as USP28 and phosphorylation by the
MAPK/ERK pathway, that protect it from degradation. At 96 h, there
was a not significant moderate negative correlation for 50 µM RSV
(r=−0.4039) and a very strong positive correlation for 150 µM
(r=0.7669). Although the results were not significant, these
results confirm that the protein expression must be being
stabilized post translationally by the aforementioned mechanisms,
and mRNA and protein expression only demonstrated a positive
correlation at the maximum time and concentration. The
concentration of 150 µM RSV proved to be the best treatment option
to reduce both Lin28A mRNA and protein levels in this cell
model.
Since RSV decreased Lin28A expression, the present
study subsequently evaluated the expression of USP28, a
deubiquitinase that has been reported to be capable of keeping
Lin28A stable, preventing its degradation (11).
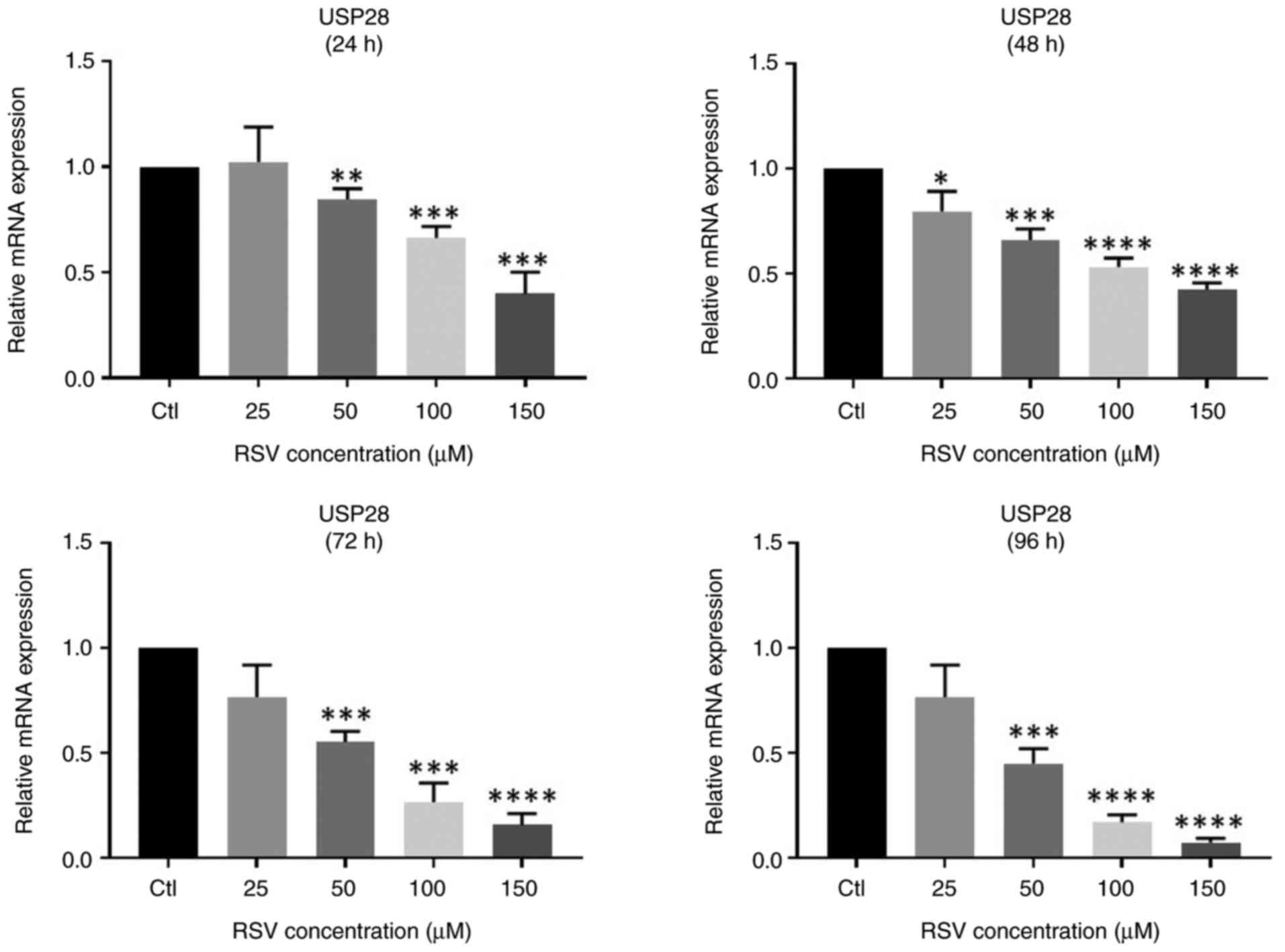
RSV decreases USP28 mRNA
expression
RT-qPCR was performed to evaluate the effect if RSV
on USP28 mRNA expression in the NCCIT cell line (Fig. 3). The results revealed that as the
concentration of RSV increased, the expression of USP28 mRNA
significantly decreased, similar to that of Lin28A. Therefore, it
was decided to also evaluate the USP28 protein.
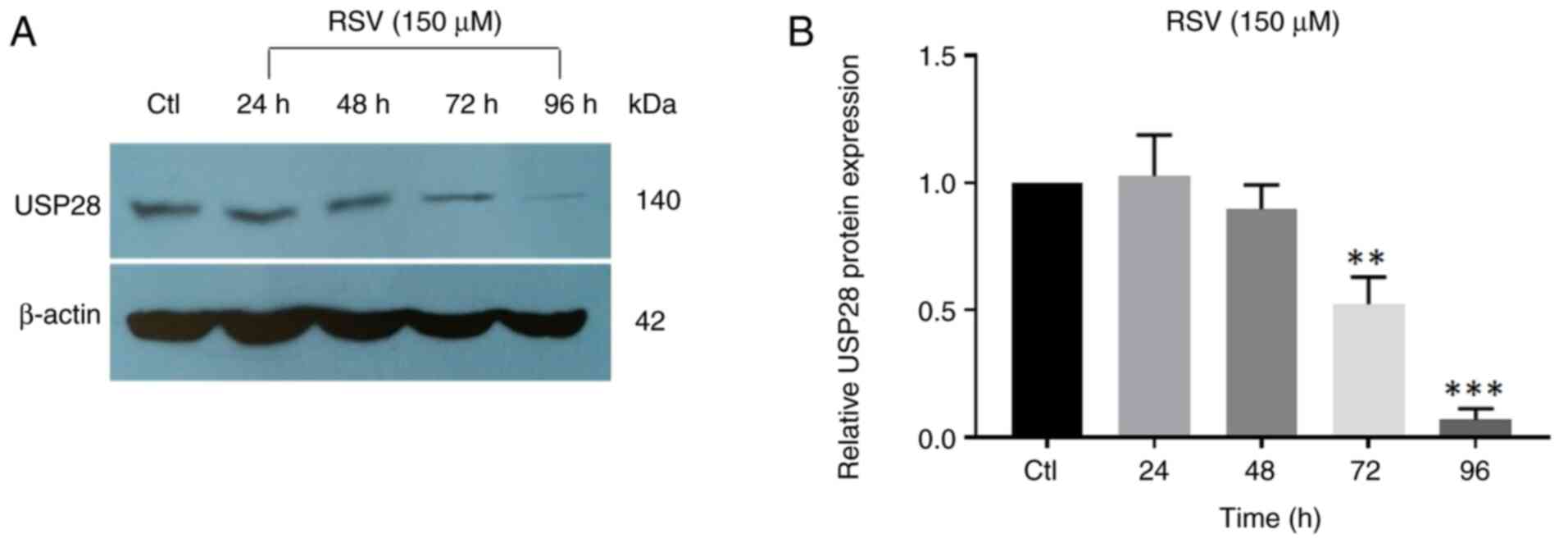
RSV decreases the expression level of
the USP28 protein
The IC50 concentration at which there was
a greatest decrease in Lin28A protein expression was 150 µM RSV
(Fig. S1). To assess the effect of
150 µM RSV on USP28 protein expression, western blot analysis was
performed (Fig. 4). The
administration of 150 µM RSV significantly reduced the protein
level of USP28 deubiquitinase, with the greatest effect observed at
72 and 96 h of treatment, similar to that of the Lin28A protein.
Once the decrease in USP28 was demonstrated with RSV treatment, the
present study continued to evaluate whether the degradation of
Lin28A by RSV occurred via the proteasomal pathway.
RSV degrades the Lin28A protein via
the proteasomal pathway
To assess if RSV treatment could induce the Lin28A
protein degradation by the proteasomal pathway, the protein level
of Lin28A was evaluated in the presence of the proteasomal
inhibitor MG132 (M) in NCCIT cells treated with 150 µM RSV (R) at
different time points (Fig. 5). The
results demonstrated that the presence of MG132 in the RSV
treatment significantly inhibited the degradation of Lin28A. Given
that the degradation of Lin28A was by the proteasomal pathway, this
could explain the stability of Lin28A protein as USP28
deubiquitinase was also downregulated at high RSV concentrations,
and at 72 and 96 h.
Inhibition of USP28 using siUSP28
decreases the levels of Lin28A, but the reduction is less
pronounced compared to the decrease observed after RSV
treatment
To evaluate the expression of Lin28A when USP28 is
silenced, NCCIT cells were transfected with different specific
siUSP28s using lipofectamine (Fig.
S2). The results were evaluated using western blotting, noting
that for each of the siRNAs used, silencing of the expression of
USP28 was observed. However, the present study used a combination
of the three siRNAs to obtain a better result for the following
experiments. Fig. 6A shows the
expression of USP28 mRNA in cells transfected with siUSP28 at
different times, and how expression significantly decreased
compared with in the group without siRNAs. Concordantly, a
significant decrease in USP28 protein expression was observed when
NCCIT cells were transfected with siUSP28 (Fig. 6B). Notably, when evaluating the mRNA
expression of Lin28A in NCCIT cells, it decreased significantly
after 48 h of transfection with siUSP28 (Fig. 6C). By contrast, Lin28A protein
expression remained stable at the same time (48 h) in response to
siUSP28 transfection in NCCIT cells (Fig. 6D); however, a decrease in Lin28A
protein levels was observed after 72 and 96 h of siUSP28
transfection, although this reduction was smaller compared with in
response to RSV treatment (150 µM) alone (Fig. S3). This result is one of the key
findings that confirms the importance of the USP28 mechanism in
stabilizing the Lin28A protein. However, when one mechanism was
knocked down (e.g., using siRNA against USP28), the other mechanism
appeared to contribute to the stabilization of the protein by
increasing its half-life, as seen with phosphorylation at S200 by
the MAPK pathway. Fig. S3 shows
the effect of 150 µM RSV on decreasing protein levels of Lin28A at
different treatment times; this demonstrates that RSV at that
concentration substantially reduces Lin28A protein levels, even
more so than siUSP28 treatment. The minor reduction in Lin28A
protein expression in response to siUSP28 transfection (Fig. 6D) could be explained by the fact
that siUSP28 did not completely ablate USP28 mRNA and protein
expression. Therefore, the remaining expression of USP28 may have
been sufficient to protect the Lin28A protein. Alternatively, an
additional mechanism for Lin28A stability is in place. Given that
Lin28A phosphorylation is a key protective mechanism, the present
study examined the expression of pLin28A in cells transfected with
siUSP28. As expected, no downregulation of pLin28A was
demonstrated, given that USP28 has no direct phosphorylating
activity (12). Notably, the
results demonstrated a marked increase in pLin28A expression upon
knockdown of USP28 with siRNAs (Fig.
6D). This increase may also have helped in the protection of
Lin28A and may be due to a negative feedback loop between USP28 and
ERK signaling.
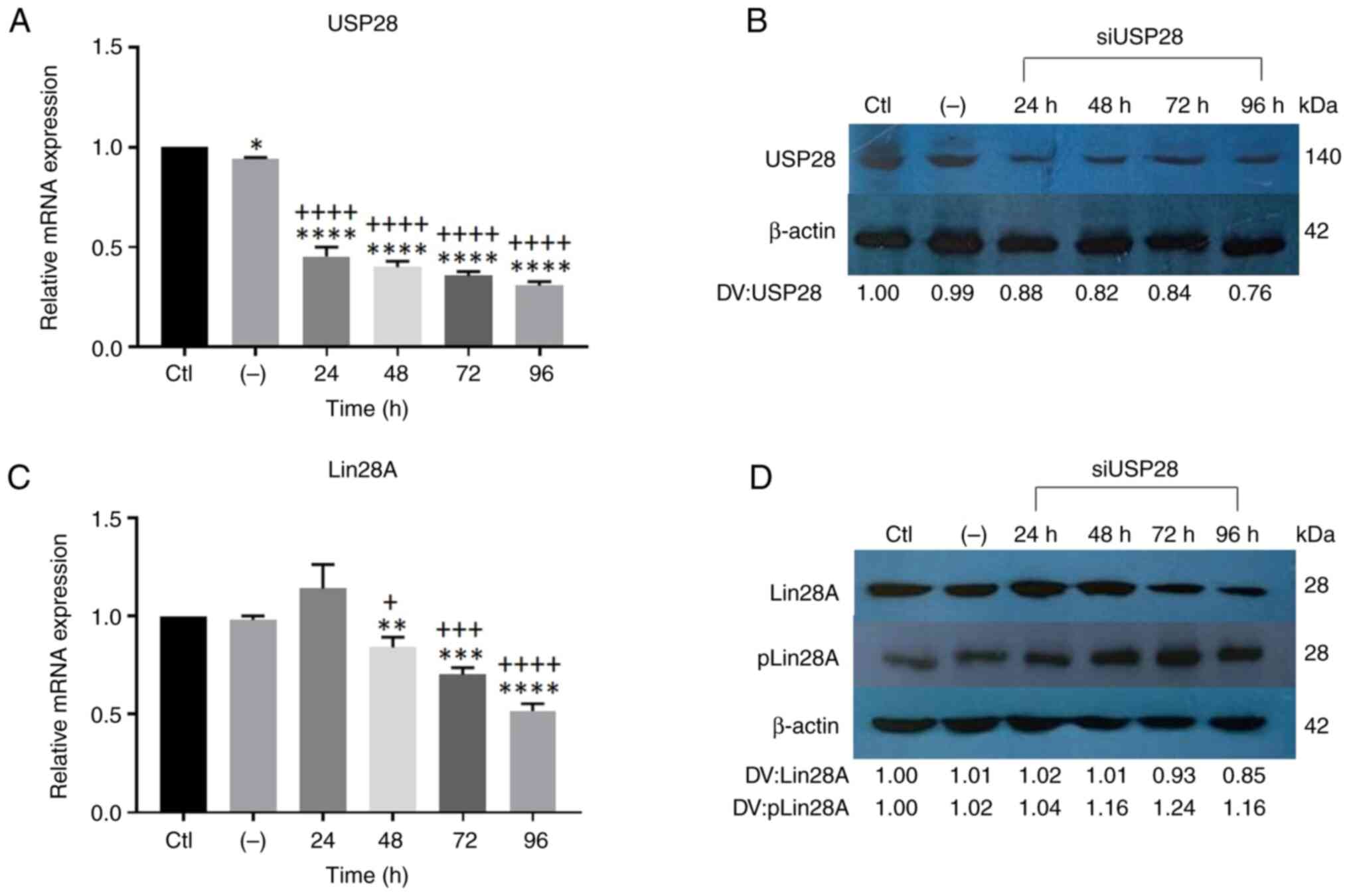 | Figure 6.Inhibition of USP28 with specific
siRNAs is not sufficient to totally silence Lin28A protein
expression. (A) Expression of USP28 mRNA in cells transfected with
siUSP28, quantified using RT-qPCR. The controls were transfected
NCCIT cells and the transfection of a nonspecific siRNA that did
not affect the gene of interest (USP28). (B) Representative western
blot of USP28 protein expression in cells transfected with siUSP28
at different times (24, 48, 72 and 96 h). (C) Expression of Lin28A
mRNA in cells transfected with siUSP28, quantified by stem-loop
RT-qPCR. (D) Representative western blot of the expression of
Lin28A protein and pLin28A by MAPK/ERK (serine 200) in cells
transfected with siUSP28 at different times (24, 48, 72 and 96 h).
*P<0.05; **P<0.01; ***P<0.001; ****P<0.0001 vs. Ctl.
+P<0.05; +++P<0.001;
++++P<0.0001 vs. (−). DV, densitometry value; USP28,
ubiquitin-specific protease 28; si, small-interfering; RT-qPCR,
reverse transcription-quantitative PCR; Ctl, control; (−),
nonspecific siRNA; pLin28A, phosphorylated Lin28A. |
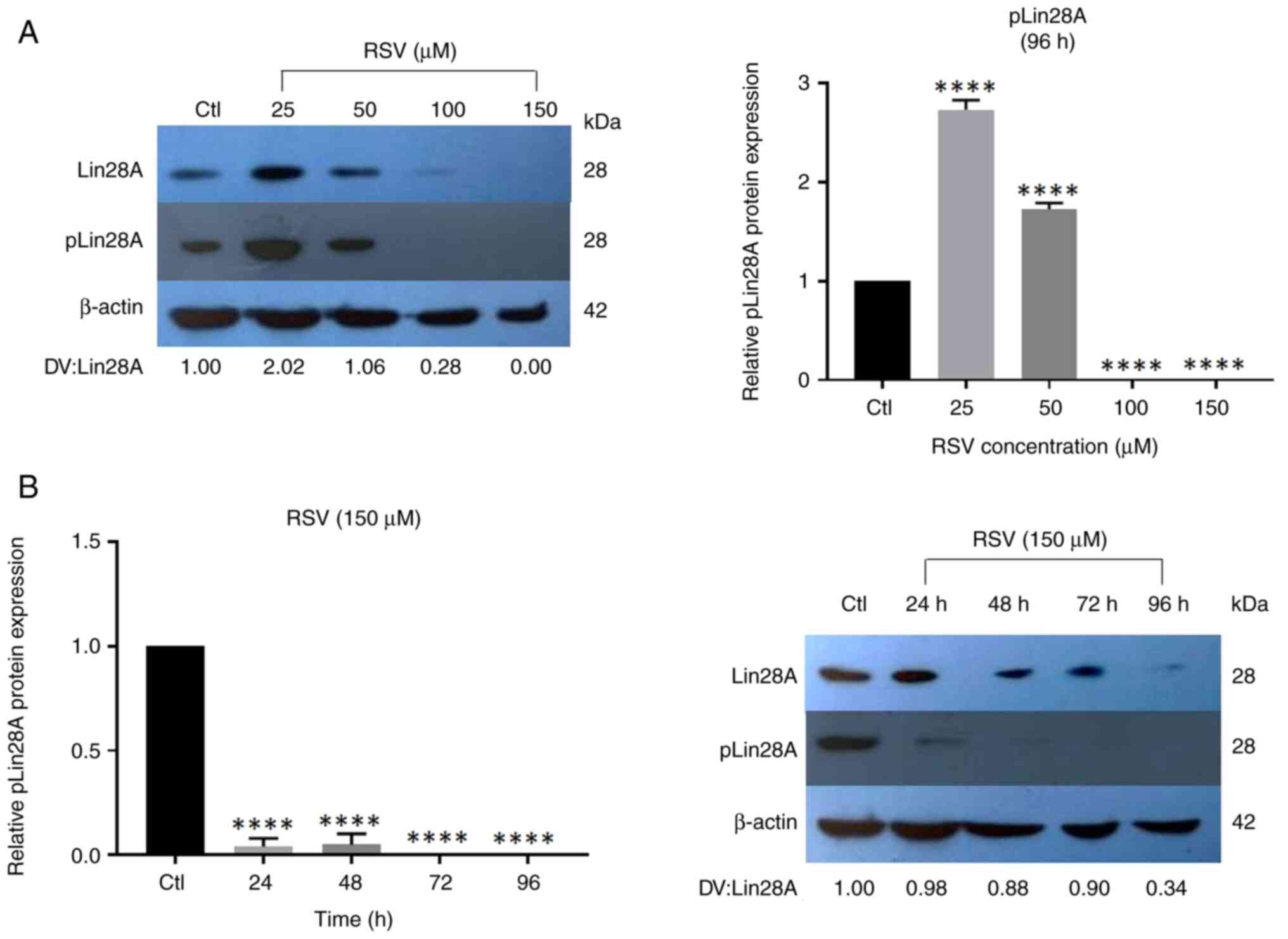
RSV decreases the expression of Lin28A
S200 phosphorylation
The effect of RSV on the phosphorylation of S200 in
the Lin28A protein was evaluated. NCCIT cells were treated with
different concentrations of RSV for 96 h (a time for which a
greater effect on the reduction of Lin28A protein was
demonstrated). The results revealed that RSV at low concentrations
(25 and 50 µM) significantly increased pLin28A expression, whereas
concentrations of 100 and 150 µM RSV significantly decreased
pLin28A expression compared with those in the control group
(Fig. 7A). The effect of 150 µM RSV
was also evaluated at different time periods. The findings
demonstrated that treatment with 150 µM RSV significantly decreased
pLin28A expression from 24 h compared with that in the control
group (Fig. 7B). This suggests that
RSV can reduce Lin28A phosphorylation via the MAPK/ERK pathway,
which was reported by Tsanov et al (12). The observed decrease in Lin28A
phosphorylation following RSV exposure provides experimental
evidence explaining why siUSP28 alone did not impact Lin28A protein
stability. By contrast, RSV, by simultaneously reducing USP28
levels and affecting the MAPK/ERK pathway (both mechanisms involved
in Lin28A stabilization), was able to destabilize Lin28A.
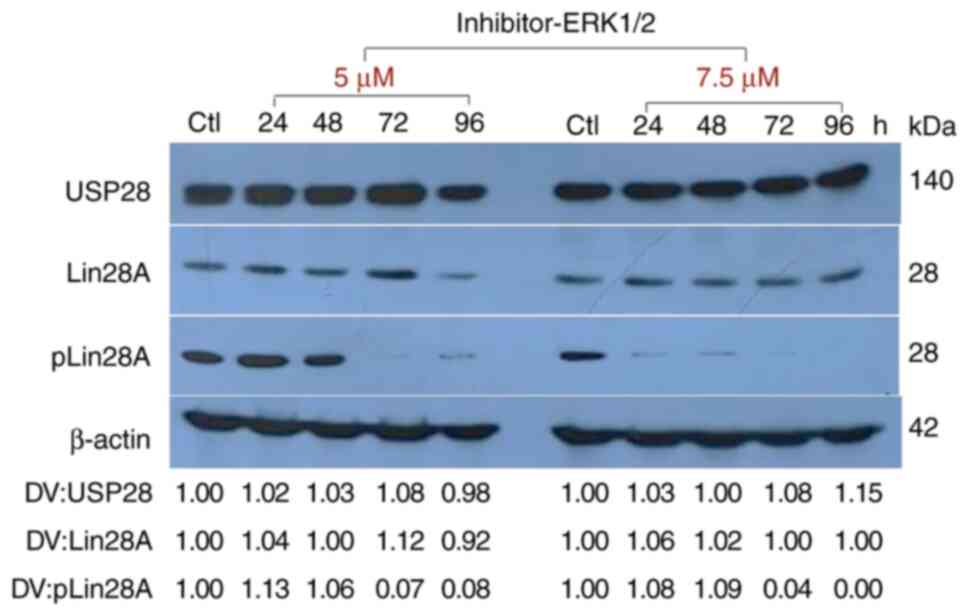
Inhibition of the MAPK/ERK pathway
reduces the level of pLin28A protein
The effect of inhibiting only the activation of
ERK1/2 on the phosphorylation of Lin28A at S200 (pLin28A) was
evaluated, as well as if the stability of total Lin28A and USP28
expression in the NCCIT cell line was affected. The cells were
treated with a specific inhibitor of the activation of ERK1/2, a
member of the MAPK pathway responsible for the phosphorylation of
Lin28A at S200 (12). First, an MTT
assay was performed to assess the safe dose of inhibitor that could
be used to treat the cells; in this experiment, an IC50
of 9 µM was obtained (Fig. S4).
Subsequently, the following experiments were performed with 5 or
7.5 µM of the inhibitor, which were concentrations that had
minimally effects on cell viability. It was demonstrated that both
concentrations were sufficient for a marked decrease in pLin28A
level. Furthermore, neither concentration notably affected the
expression of the Lin28A and USP28 proteins (Fig. 8). This indicates that both
stabilization pathways (USP28 and Lin28A phosphorylation by
MAPK/ERK) can be blocked by RSV to reduce Lin28A protein
expression.
RSV strongly decreases Lin28A protein
expression
Finally, the present study evaluated the kinetics of
Lin28A protein degradation in NCCIT cells using different
treatments (RSV, siUSP28 and inhibitor/ERK) in a CHX condition
assay (Figs. S5 and 9). In this experiment, RSV treatment
significantly reduced Lin28A protein expression compared with that
in the other treatment groups, such as cells treated with siUSP28,
the ERK inhibitor, or a combination of siUSP28 and ERK inhibitor.
Furthermore, the half-life of the Lin28A protein was 12.4 h with
siUSP28 treatment, 17.9 h with inhibitor/ERK treatment, 14.9 h with
the combined treatment of siUSP28 + inhibitor/ERK, and 10.8 h with
RSV treatment. This suggests that RSV may regulate other factors
that could be involved in Lin28A stability.
Discussion
The present study aimed to elucidate the potential
mechanisms by which RSV treatment downregulates the oncoprotein
Lin28A in NCCIT cells. The NCCIT cell line was used, which is
classified as a germ cell tumor and served as a convenient in
vitro model of cancer stem-like cells (21–23).
Furthermore, the NCCIT line is known to have high expression of
both Lin28A and USP28 (11). Given
the high basal expression of Lin28A and USP28 in the NCCIT cells,
this made the NCCIT line the most appropriate choice to assess
whether the natural compound RSV could effectively reduce Lin28A
expression, and to elucidate the specific mechanisms by which it
does so, including the role of USP28.
The results of the present study indicate that RSV
inhibits Lin28A mRNA expression in a dose-dependent manner between
concentrations 50–150 µM. Notably, at the lowest dose of RSV tested
(25 µM), there was an increase in Lin28A mRNA expression levels.
This biphasic effect is likely due to the hormesis phenomenon
exhibited by certain compounds, in which low doses stimulate
activity but higher doses reverse this effect (24). Whilst the exact reason for hormesis
remains unclear, evaluating this dose-response behavior is
important to determine the most effective concentration for a given
positive or negative outcome, regardless of the substance. The
RSV-mediated decrease in Lin28A expression may be linked to reduced
levels of transcription factors such as SP1 and STAT3, both of
which have been shown to bind the Lin28A promoter and promote its
transcription (14,25,26).
In this regard, the present study assessed SP1 and STAT3 expression
following RSV treatment (50–150 µM) and demonstrated that both were
downregulated (data not shown).
Whilst the present study provides useful insights,
further research is needed to validate certain findings. For
example, experiments assessing whether the aforementioned
transcription factors directly bind to the Lin28A promoter in this
model, and whether RSV specifically impacts SP1 and STAT3 protein
levels, would strengthen the conclusions. It would also be valuable
to investigate whether RSV uniformly affects long noncoding RNAs
known to promote Lin28A transcription, such as H19, ZNFX1 antisense
RNA 1 and long intergenic non-protein coding RNA 1451 (27–29).
Additionally, exploring the expression of several oncoproteins
previously reported to form positive feedback loops with Lin28A,
such as c-MYC, octamer-binding transcription factor 4 and NANOG,
could further elucidate the molecular mechanisms of RSV (29,30).
Moreover, further targeted validation studies are required to fully
characterize the multi-level effects of RSV on Lin28A regulation
and contextualize the findings within existing knowledge of
transcriptional and epigenetic control of this critical
cancer-related gene. Overall, whilst promising, additional
experimentation would help substantiate certain aspects of the
present work.
When assessing Lin28A protein expression in NCCIT
cells, the results of the present study revealed that its stability
was not directly associated with mRNA downregulation, as protein
levels only decreased at the highest RSV concentrations tested (100
and 150 µM). These results indicate that the Lin28A protein is
highly stable, likely due to post-translational regulatory
mechanisms previously described. For example, Haq et al
(11) reported that Lin28A
interacts with USP28 and deubiquitinates it, maintaining the
protein stability and prolonging its half-life. Conversely, Tsanov
et al (12) reported that
Lin28A is phosphorylated by a MAPK (ERK1/2) at S200, allowing
post-translational stabilization. Considering these studies, the
present study evaluated whether RSV treatment could interfere with
these post-translational regulatory pathways. Specifically, the
present study assessed whether RSV would disrupt the interactions
between Lin28A and USP28 or prevents ERK1/2-mediated
phosphorylation at S200.
The MTT assay results in the present study agreed
with previous reports (31,32), indicating that RSV inhibited NCCIT
cell viability in a dose-dependent manner with an IC50
of 148 µM. All experiments were performed using the same
concentrations (25–150 µM). Notably, only treatments with 100 or
150 µM RSV resulted in a significant decrease in Lin28A protein
expression levels over time, with the protein expression almost
completely absent after 96 h at 150 µM. The present study also
assessed USP28 protein expression following treatment with 150 µM
RSV. The findings demonstrated a significant reduction in USP28
protein levels similar to the decrease observed for Lin28A protein
expression. These findings support the hypothesis that disruption
of the Lin28A-USP28 interaction, which stabilizes Lin28A, could
underlie the mechanism of RSV of reducing Lin28A protein expression
levels at higher doses.
USP28 is an important regulator of proteins involved
in proliferation and metastasis, such as c-MYC, lysine-specific
histone demethylase 1, hypoxia-inducible factor 1α, c-JUN and
mediator of DNA damage checkpoint protein 1; moreover, USP28 is
usually upregulated in several cancers including pancreatic,
squamous, gastric and colorectal (33–37),
marking it as an important therapeutic target (33,37).
The results of the present study demonstrated that RSV
downregulated USP28 at the protein level. Furthermore, analysis
revealed that RSV also reduced USP28 mRNA expression in a dose- and
time-dependent manner, suggesting transcriptional regulation. This
effect may occur through downregulation of transcription factors
that bind the USP28 promoter or upregulation of inhibitory micro
(mi)RNAs. Potential candidates affected by RSV include oncogenic
regulators c-JUN and c-MYC, which activate USP28 transcription
(38–40). Previous studies have reported RSV
inhibits these factors (40–42).
Additionally, RSV may increase miR92b-3p expression, reported to
repress USP28 by binding its 3′-untraslated region (43). RSV could also boost miR-216b
expression, which inhibits USP28 mRNA in hepatocellular carcinoma
cells (44). Increased miR-622
expression is also possible as it acts as a tumor suppressor
against KRAS, an oncogene that upregulates USP28 (38). Whilst speculative, future
experiments could assess whether RSV modulation of these
transcriptional and post-transcriptional regulators underlies USP28
mRNA reduction. Overall, downregulating both USP28 protein and mRNA
expression may contribute to the anticancer effects of RSV in the
model in the present study. In addition, the present study
demonstrated that RSV induces Lin28A degradation by the proteasomal
pathway, consistent with the findings of Haq et al (11), who reported that Lin28A is degraded
by this route. On the other hand, comparing the expression of
Lin28A and pLin28A after USP28 inhibition using siRNAs revealed a
small decrease in Lin28A protein expression, while pLin28A levels
increased. We hypothesize that constant Lin28A mRNA expression may
compensate for loss of USP28-mediated protection, whilst decreased
USP28 may elevate ERK activation and consequently pLin28A levels
(12,45). Notably, USP28 knockout alone in the
NCCIT cells was insufficient to completely reduce Lin28A protein
expression, indicating additional regulatory mechanisms.
Phosphorylation of Lin28A at S200 is another critical
post-translational modification that enhances stability (12); therefore, the effects on RSV on the
phosphorylation of Lin28A mediated by the MAPK/ERK pathway was
analyzed. A biphasic effect was observed, with low RSV
concentrations (25–50 µM) increasing pLin28A expression, and higher
doses (100–150 µM) significantly decreasing it. Low doses of RSV
(25 µM) appeared to stimulate the MAPK/ERK pathway, as demonstrated
by the increased Lin28A phosphorylation. This likely increased
Lin28A mRNA and protein synthesis at early time points under normal
conditions and it could be possible that the stability of the
Lin28A protein influences in the autoregulation of Lin28 mRNA.
Previous studies have reported that phosphorylation can enhance the
activity of Lin28 on its mRNA targets (12,46).
Moreover, there is evidence that Lin28 binds to the mRNA of several
targets, including its own mRNA, increasing the steady-state levels
of its own transcript and consequently enhancing its expression
(47,48). Additionally, increased Lin28
expression has been reported to boost the translation of its
targeted mRNAs by recruiting RNA helicase A and LINE-1 type
transposase domain containing 1 to polysomes (49,50).
Therefore, it seems plausible that the interaction of Lin28 with
its own mRNA could also promote its own translation.
On the other hand, higher doses of RSV (100–150 µM)
inhibited the MAPK/ERK pathway and decreased Lin28A synthesis. As
RSV can inhibit the MAPK/ERK signaling cascade (51–53),
we hypothesize that this polyphenol can reduces phosphorylation at
S200, favoring Lin28A degradation. Notably, a recent study by Tan
et al (54) described the
significance of phosphorylation at S200 for Lin28A stability and
function, reporting that mutation of this region reduced stability
and impaired function. The results of the present study
demonstrated that whilst RSV at 150 µM decreases S200
phosphorylation as early as 24 h, total Lin28A protein levels
remain stable through 96 h of treatment. This indicates that
additional protective mechanisms may sustain Lin28A stability over
prolonged periods, despite diminished S200 phosphorylation. It
would be worthwhile to elucidate these stabilizing mechanisms more
precisely using the model in the present study. Specifically
assessing how Lin28A stability is maintained for lengthy durations
when S200 phosphorylation is suppressed by RSV could yield valuable
insights into the complex regulation of this important oncoprotein.
Overall, the present study provides new context regarding Lin28A
phosphorylation and complements prior findings on phosphorylation's
role in Lin28A turnover.
Subsequently, the present study assessed whether
inhibiting the MAPK/ERK pathway was sufficient to decrease both
pLin28A and total Lin28A protein levels. pLin28A and total Lin28A
were analyzed in NCCIT cells treated with only the ERK inhibitor
(PD0325901), which was also used in the study by Tsanov et
al (12). These experiments
demonstrated that whilst phosphorylation of Lin28A was reduced,
total Lin28A protein remained stable. USP28 protein expression with
the ERK inhibitor treatment was also analyzed and it was
demonstrated that USP28 expression was maintained, suggesting that
Lin28A protein stability depends on both pathways (USP28 and
MAPK/ERK). Inhibiting only one pathway may not be sufficient for
complete Lin28A degradation, at least in a model with continuous
Lin28A mRNA expression. Furthermore, using CHX, the production of
new proteins was inhibited. Therefore, at the highest RSV
concentration of 150 µM, the degradation of Lin28A was most
effectively observed under these assay conditions.
In contrast, the experiments presented in Figs. 1 and 2 measured steady-state protein and mRNA
levels for 24 h, allowing both the synthesis of the new Lin28A
protein and the production of other proteins to affect stability
and degradation to influence protein levels. It was demonstrated
that inhibiting USP28 was more important for Lin28A degradation
than inhibiting ERK activation. The most significant finding was
that RSV was the most effective treatment at inhibiting Lin28A
protein levels. Notably, the degradation kinetics observed with
combined USP28 siRNA and ERK inhibitor treatments mimicked those
seen with RSV alone, suggesting these pathways are key mediators of
the negative regulation of the Lin28A protein by RSV. However, it
is possible RSV may also impact other pathways. For example, RSV
could affect Kruppel-Associated Box Domain-Associated Protein 1
(KAP1), a protein reported to bind Lin28A and inhibit its
ubiquitination similar to USP28 (55). Additionally, RSV may directly
upregulate miRNA Let-7 expression, indirectly downregulating Lin28A
in turn (9,56). However, further study is needed to
confirm whether RSV does indeed act through these or other
potential mechanisms. Specifically, targeted experiments
investigating the direct effects of RSV on KAP1 and Let-7 levels
could provide insight into additional pathways of Lin28A regulation
affected by this polyphenol. Whilst the findings of the present
study implicate USP28 and ERK signaling, more research is warranted
to fully elucidate the multidimensional impact of RSV on Lin28A
expression and turnover at the post-transcriptional and
translational levels.
The primary limitation of the present study is that
human cancer samples were not assessed as obtaining human cancer
samples that include treatment with RSV to observe the relationship
between the decrease in Lin28A expression and patient survival is
not feasible in Mexico. However, the findings of the present study
could be validated through further investigations using other types
of cancer cells, such as ovarian, gastric, prostate and breast
cancer, where previous research has established that both Lin28A
and USP28 are overexpressed. Although the results of the present
study contribute to the general knowledge of RSV pathways and
mechanisms to regulate Lin28A and USP28 that may help to develop
new strategies to combat cancer, it is acknowledged that the study
lacks data on human tissue experimentation and in vivo
models, which are the most important counterpart of in vitro
first approach experiments.
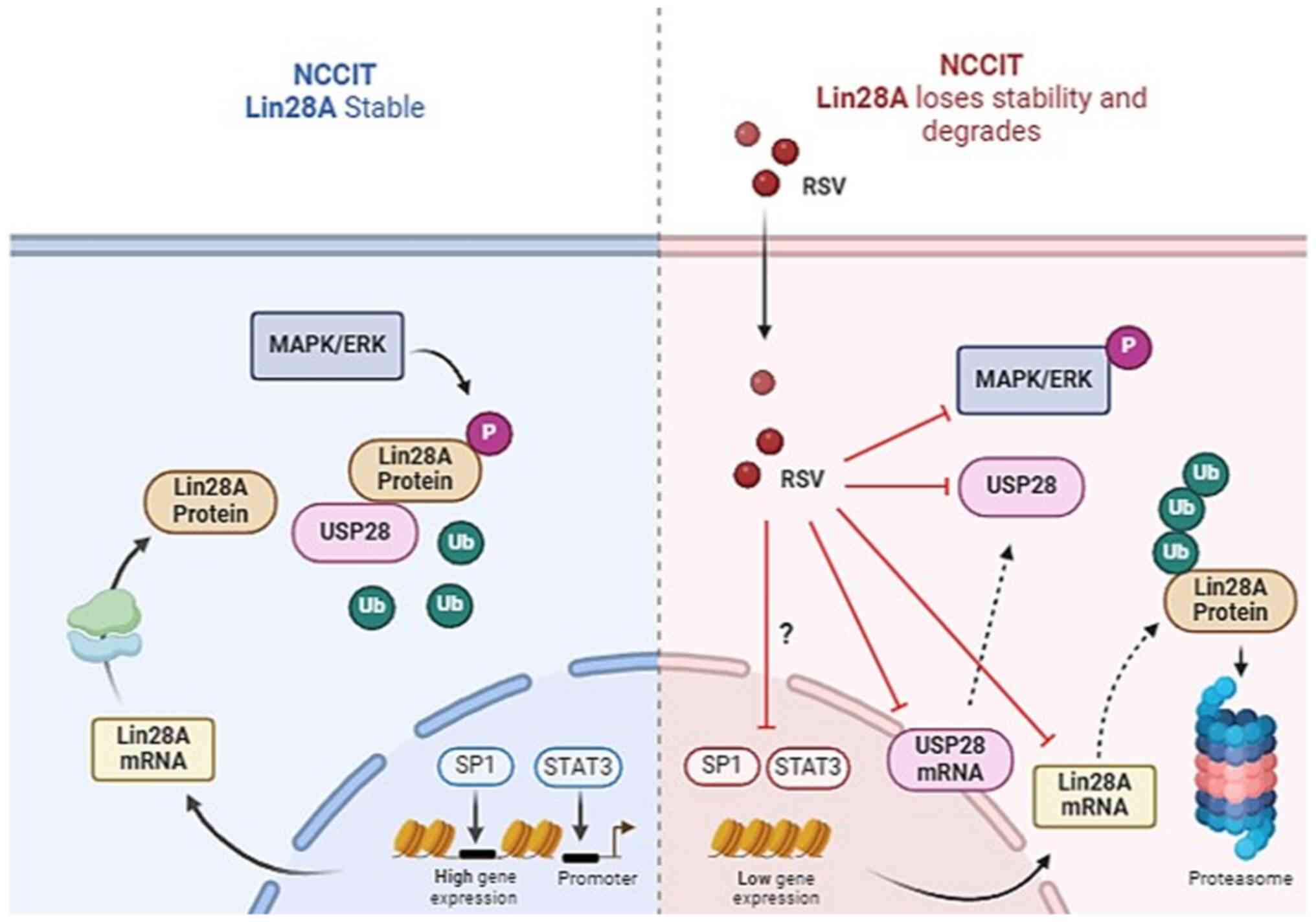
In conclusion, the present study demonstrated that
RSV inhibits Lin28A expression through multiple mechanisms. RSV
targets both USP28-mediated post-translational regulation of Lin28A
as well as MAPK/ERK phosphorylation of Lin28A, promoting Lin28A
degradation through the proteasomal pathway (Fig. 10). Notably, the finding that RSV
inhibited USP28 is significant, as USP28 stabilizes several
oncoproteins and facilitates EMT and metastasis (57). The ability of RSV to modulate the
USP28 pathway implies it may have broader anticancer effects beyond
Lin28A suppression. Collectively, RSV exerts multidirectional
inhibitory effects on Lin28A expression and stabilization,
supporting its potential as an attractive adjunctive therapy.
Moreover, RSV simultaneously targets both Lin28A and USP28, which
likely enhances its antitumor efficacy. This multifaceted mechanism
supports further development of RSV as part of comprehensive cancer
treatment regimens.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Ms. Elizabeth
Álvarez-Ríos, Dr Rodolfo Ocádiz-Delgado and Mr. Lauro
Macias-González for technical support (all affiliated with the
Department of Genetics and Molecular Biology, Center for Research
and Advanced Studies of the National Polytechnic Institute, Mexico
City, Mexico).
Funding
Funding: No funding was received.
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
SCN, SCR and EGV performed all experiments. SCN and
EGV performed the data interpretation and analysis of the results.
SCN, EGV, PG and JDC conceptualized and proposed the study. SCN
wrote the manuscript and generated the figures. SCN, EGV, SCR, PG
and JDC confirm the authenticity of all raw data. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mayr F and Heinemann U: Mechanisms of
Lin28-mediated miRNA and mRNA regulation-a structural and
functional perspective. Int J Mol Sci. 14:16532–16553. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang J, Ratanasirintrawoot S,
Chandrasekaran S, Wu Z, Ficarro SB, Yu C, Ross CA, Cacchiarelli D,
Xia Q, Seligson M, et al: Lin28 regulates stem cell metabolism and
conversion to primed pluripotency. Cell Stem Cell. 19:66–80. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin Z, Radaeva M, Cherkasov A and Dong X:
Lin28 regulates cancer cell stemness for tumour progression.
Cancers (Basel). 14:46402022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hanna J, Saha K, Pando B, van Zon J,
Lengner CJ, Creyghton MP, van Oudenaarden A and Jaenisch R: Direct
cell reprogramming is a stochastic process amenable to
acceleration. Nature. 462:595–601. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu J, Vodyanik MA, Smuga-Otto K,
Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA,
Ruotti V, Stewart R, et al: Induced pluripotent stem cell lines
derived from human somatic cells. Science. 318:1917–1920. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hamano R, Miyata H, Yamasaki M, Sugimura
K, Tanaka K, Kurokawa Y, Nakajima K, Takiguchi S, Fujiwara Y, Mori
M and Doki Y: High expression of Lin28 is associated with tumour
aggressiveness and poor prognosis of patients in oesophagus cancer.
Br J Cancer. 106:1415–1423. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li M, Chen H and Wu T: LIN28: A cancer
stem cell promoter for immunotherapy in head and neck squamous cell
carcinoma. Oral Oncol. 98:92–95. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsialikas J and Romer-Seibert J: LIN28:
Roles and regulation in development and beyond. Development.
142:2397–2404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Balzeau J, Menezes MR, Cao S and Hagan JP:
The LIN28/let-7 pathway in cancer. Front Genet. 8:312017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Piskounova E, Polytarchou C, Thornton JE,
LaPierre RJ, Pothoulakis C, Hagan JP, Iliopoulos D and Gregory RI:
Lin28A and Lin28B inhibit let-7 MicroRNA biogenesis by distinct
mechanisms. Cell. 147:1066–1079. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haq S, Das S, Kim DH, Chandrasekaran AP,
Hong SH, Kim KS and Ramakrishna S: The stability and oncogenic
function of LIN28A are regulated by USP28. Biochim Biophys Acta Mol
Basis Dis. 1865:599–610. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tsanov KM, Pearson DS, Wu Z, Han A,
Triboulet R, Seligson MT, Powers JT, Osborne JK, Kane S, Gygi SP,
et al: LIN28 phosphorylation by MAPK/ERK couples signalling to the
post-transcriptional control of pluripotency. Nat Cell Biol.
19:60–67. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Y, Xie C, Zheng X, Nie X, Wang Z, Liu
H and Zhao Y: LIN28/ let-7/PD-L1 pathway as a target for
cancer immunotherapy. Cancer Immunol Res. 7:487–497. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu J, Zhang Z, Huang L, Xiong J, Zhou Z,
Yu H, Wu L, Liu Z and Cao K: Let-7a suppresses Ewing sarcoma CSCs'
malignant phenotype via forming a positive feedback circuit with
STAT3 and lin28. J Bone Oncol. 31:1004062021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang R, Dong H, Jia S and Yang Z:
Resveratrol as a modulatory of apoptosis and autophagy in cancer
therapy. Clin Transl Oncol. 24:1219–1230. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee SR, Jin H, Kim WT, Kim WJ, Kim SZ,
Leem SH and Kim SM: Tristetraprolin activation by resveratrol
inhibits the proliferation and metastasis of colorectal cancer
cells. Int J Oncol. 53:1269–1278. 2018.PubMed/NCBI
|
|
17
|
Sauter ER: Cancer prevention and treatment
using combination therapy with natural compounds. Expert Rev Clin
Pharmacol. 13:265–285. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Naeem A, Hu P, Yang M, Zhang J, Liu Y, Zhu
W and Zheng Q: Natural products as anticancer agents: Current
status and future perspectives. Molecules. 27:83672022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Farooqi A, Khalid S and Ahmad A:
Regulation of cell signaling pathways and miRNAs by resveratrol in
different cancers. Int J Mol Sci. 19:6522018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ren B, Kwah MXY, Liu C, Ma Z, Shanmugam
MK, Ding L, Xiang X, Ho PC, Wang L, Ong PS and Goh BC: Resveratrol
for cancer therapy: Challenges and future perspectives. Cancer
Lett. 515:63–72. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tykwinska K, Lauster R, Knaus P and
Rosowski M: Growth and differentiation factor 3 induces expression
of genes related to differentiation in a model of cancer stem cells
and protects them from retinoic acid-induced apoptosis. PLoS One.
8:e706122013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yun JH, Kim KA, Yoo G, Kim SY, Shin JM,
Kim JH, Jung SH, Kim J and Nho CW: Phenethyl isothiocyanate
suppresses cancer stem cell properties in vitro and in a xenograft
model. Phytomedicine. 30:42–49. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
You JS, Kang JK, Seo DW, Park JH, Park JW,
Lee JC, Jeon YJ, Cho EJ and Han JW: Depletion of embryonic stem
cell signature by histone deacetylase inhibitor in NCCIT cells:
Involvement of nanog suppression. Cancer Res. 69:5716–5725. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jodynis-Liebert J and Kujawska M: Biphasic
dose-response induced by phytochemicals: Experimental evidence. J
Clin Med. 9:7182020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
You X, Liu F, Zhang T, Lv N, Liu Q, Shan
C, Du Y, Kong G, Wang T, Ye L and Zhang X: Hepatitis B virus X
protein upregulates Lin28A/Lin28B through Sp-1/c-Myc to enhance the
proliferation of hepatoma cells. Oncogene. 33:449–460. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo L, Chen C, Shi M, Wang F, Chen X, Diao
D, Hu M, Yu M, Qian L and Guo N: Stat3-coordinated
Lin-28-let-7-HMGA2 and miR-200-ZEB1 circuits initiate and maintain
oncostatin M-driven epithelial-mesenchymal transition. Oncogene.
32:5272–5282. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhuo ZL, Xian HP, Sun YJ, Long Y, Liu C,
Liang B and Zhao XT: Long noncoding RNA ZNFX1-AS1 promotes the
invasion and proliferation of gastric cancer cells by regulating
LIN28 and CAPR1N1. World J Gastroenterol. 28:4973–4992. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi H, Xie J, Wang K, Li W, Yin L, Wang G,
Wu Z, Ni J, Mao W, Guo C and Peng B: LINC01451 drives
epithelial-mesenchymal transition and progression in bladder cancer
cells via LIN28/TGF-β/Smad pathway. Cell Signal. 81:1099322021.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Krsnik D, Marić T, Bulić-Jakuš F, Sinčić N
and Bojanac AK: LIN28 family in testis: Control of cell renewal,
maturation, fertility and aging. Int J Mol Sci. 23:72452022.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee S, Wottrich S and Bonavida B:
Crosstalks between Raf-kinase inhibitor protein and cancer stem
cell transcription factors (Oct4, KLF4, Sox2, Nanog). Tumour Biol.
39:1010428317692252017. View Article : Google Scholar
|
|
31
|
Wu H, Chen L, Zhu F, Han X, Sun L and Chen
K: The cytotoxicity effect of resveratrol: Cell cycle arrest and
induced apoptosis of breast cancer 4T1 cells. Toxins (Basel).
11:7312019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fukuda M, Ogasawara Y, Hayashi H, Inoue K
and Sakashita H: Resveratrol inhibits proliferation and induces
autophagy by blocking SREBP1 expression in oral cancer cells.
Molecules. 27:82502022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Liu Z, Zhang L, Yang Z, Chen X,
Luo J, Zhou Z, Mei X, Yu X, Shao Z, et al: Targeting deubiquitinase
USP28 for cancer therapy. Cell Death Dis. 9:1862018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao L, Zhang T, Feng X, Chang J, Suo FZ,
Ma JL, Liu YJ, Liu Y, Zheng YC and Liu HM: USP28 contributes to the
proliferation and metastasis of gastric cancer. J Cell Biochem.
120:7657–7666. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Maier CR, Hartmann O, Prieto-Garcia C,
Al-Shami KM, Schlicker L, Vogel FCE, Haid S, Klann K, Buck V, Münch
C, et al: USP28 controls SREBP2 and the mevalonate pathway to drive
tumour growth in squamous cancer. Cell Death Differ. 30:1710–1725.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen L, Xu Z, Li Q, Feng Q, Zheng C, Du Y,
Yuan R and Peng X: USP28 facilitates pancreatic cancer progression
through activation of Wnt/β-catenin pathway via stabilising FOXM1.
Cell Death Dis. 12:8872021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Prieto-Garcia C, Tomašković I, Shah VJ,
Dikic I and Diefenbacher M: USP28: Oncogene or tumor suppressor? A
unifying paradigm for squamous cell carcinoma. Cells. 10:26522021.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Serra RW, Fang M, Park SM, Hutchinson L
and Green MR: A KRAS-directed transcriptional silencing pathway
that mediates the CpG island methylator phenotype. Elife.
3:e023132014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Diefenbacher ME, Popov N, Blake SM,
Schülein-Völk C, Nye E, Spencer-Dene B, Jaenicke LA, Eilers M and
Behrens A: The deubiquitinase USP28 controls intestinal homeostasis
and promotes colorectal cancer. J Clin Invest. 124:3407–3418. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Faid I, Al-Hussaini H and Kilarkaje N:
Resveratrol alleviates diabetes-induced testicular dysfunction by
inhibiting oxidative stress and c-Jun N-terminal kinase signaling
in rats. Toxicol Appl Pharmacol. 289:482–494. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang M, Jiang S, Yu F, Zhou L and Wang K:
Noncoding RNAs as molecular targets of resveratrol underlying its
anticancer effects. J Agric Food Chem. 67:4709–4719. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fouad M, Agha A, Merzabani MA and Shouman
S: Resveratrol inhibits proliferation, angiogenesis and induces
apoptosis in colon cancer cells: Calorie restriction is the force
to the cytotoxicity. Hum Exp Toxicol. 32:1067–1080. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hao X, Ma C, Chen S, Dang J, Cheng X and
Zhu D: Reverse the down regulation of miR-92b-3p by hypoxia can
suppress the proliferation of pulmonary artery smooth muscle cells
by targeting USP28. Biochem Biophys Res Commun. 503:3064–3077.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang JF: MicroRNA-216b suppresses the
cell growth of hepatocellular carcinoma by inhibiting
Ubiquitin-specific peptidase 28 expression. Kaohsiung J Med Sci.
36:423–428. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Saei A, Palafox M, Benoukraf T, Kumari N,
Jaynes PW, Iyengar PV, Muñoz-Couselo E, Nuciforo P, Cortés J,
Nötzel C, et al: Loss of USP28-mediated BRAF degradation drives
resistance to RAF cancer therapies. J Exp Med. 215:1913–1928. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu X, Chen M, Li L, Gong L, Zhou H and
Gao D: Extracellular signal-regulated kinases (ERKs) phosphorylate
lin28a protein to modulate P19 cell proliferation and
differentiation. J Biol Chem. 292:3970–3976. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Polesskaya A, Cuvellier S, Naguibneva I,
Duquet A, Moss EG and Harel-Bellan A: Lin-28 binds IGF-2 mRNA and
participates in skeletal myogenesis by increasing translation
efficiency. Genes Dev. 21:1125–1138. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wilbert ML, Huelga SC, Kapeli K, Stark TJ,
Liang TY, Chen SX, Yan BY, Nathanson JL, Hutt KR, Lovci MT, et al:
LIN28 binds messenger RNAs at GGAGA motifs and regulates splicing
factor abundance. Mol Cell. 48:195–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Närvä E, Rahkonen N, Emani MR, Lund R,
Pursiheimo JP, Nästi J, Autio R, Rasool O, Denessiouk K, Lähdesmäki
H, et al: RNA-binding protein L1TD1 interacts with LIN28 via RNA
and is required for human embryonic stem cell self-renewal and
cancer cell proliferation. Stem Cells. 30:452–460. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jin J, Jing W, Lei XX, Feng C, Peng S,
Boris-Lawrie K and Huang Y: Evidence that Lin28 stimulates
translation by recruiting RNA helicase A to polysomes. Nucleic
Acids Res. 39:3724–3734. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kortam MA, Ali BM and Fathy N: The
deleterious effect of stress-induced depression on rat liver:
Protective role of resveratrol and dimethyl fumarate via inhibiting
the MAPK/ERK/JNK pathway. J Biochem Mol Toxicol. 35:e226272021.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yu X, Sun Z, Nie S, Zhang T and Lu H:
Effects of resveratrol on mouse B16 melanoma cell proliferation
through the SHCBP1-ERK1/2 signaling pathway. Molecules.
28:76142023. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu S, Ren J, Liu S, Zhao X, Liu H, Zhou
T, Wang X, Liu H, Tang L and Chen H: Resveratrol inhibits autophagy
against myocardial ischemia-reperfusion injury through the
DJ-1/MEKK1/JNK pathway. Eur J Pharmacol. 951:1757482023. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tan T, Gao B, Yu H, Pan H, Sun Z, Lei A,
Zhang L, Lu H, Wu H, Daley GQ, et al: Dynamic nucleolar phase
separation influenced by non-canonical function of LIN28A instructs
pluripotent stem cell fate decisions. Nat Commun. 15:12562024.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Moon HJ, Lee NY, Do EK, Lee SY, Park GT,
Lim JK, Seo JK and Kim JH: Kap1 regulates the stability of Lin28A
in embryonic stem cells. Stem Cells. 40:385–396. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Peng F, Li TT, Wang KL, Xiao GQ, Wang JH,
Zhao HD, Kang ZJ, Fan WJ, Zhu LL, Li M, et al: H19/let-7/LIN28
reciprocal negative regulatory circuit promotes breast cancer stem
cell maintenance. Cell Death Dis. 8:e2569. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ren X, Jiang M, Ding P, Zhang X, Zhou X,
Shen J, Liu D, Yan X and Ma Z: Ubiquitin-specific protease 28: The
decipherment of its dual roles in cancer development. Exp Hematol
Oncol. 12:272023. View Article : Google Scholar : PubMed/NCBI
|