Introduction
Globally, cancer-associated mortality is dominated
by liver cancer, the burden of which is continually rising. For
instance, liver cancer accounted for ~758,000 deaths (7.8% of all
cancer deaths) in 2022, ranking as the third leading cause of
cancer death worldwide (1).
Moreover, between 2020 and 2040, new liver cancer cases and deaths
are projected to increase by >55% (2). The majority of patients with
hepatocellular carcinoma (HCC) are diagnosed at an advanced stage,
thereby may not get the opportunity to undergo curative treatments
(3). Molecular targeted drugs,
including lenvatinib, may bring hope to patients with advanced HCC.
However, lenvatinib resistance has been widely reported,
representing a new challenge in HCC clinical management (4,5).
As a broad-spectrum tyrosine kinase inhibitor,
lenvatinib antagonizes VEGFR, fibroblast growth factor receptor
(FGFR) and platelet-derived growth factor receptor families and
associated kinases (rearranged during transfection and c-Kit, among
others), which curtails neovascularization and cancer cell
expansion (6). Its resistance
mechanisms involve the synergistic action of numerous pathways,
including c-Myc, Wnt/β-catenin and PI3K/AKT signaling (7–9).
Novel observations have revealed the pivotal role of
circular RNAs (circRNAs) in mediating lenvatinib refractoriness as
they can indirectly regulate gene expression as ‘micro (mi)-RNA
sponges’ or directly bind to and modify key signaling proteins
(such as GSK3β and PSIP1) as ‘protein function modulators’,
enhancing cancer stem cell stemness and driving resistance by
activating core pathways such as c-Myc or Wnt/β-catenin (7–9).
Androgen receptor (AR) expression has shown marked
heterogeneity in HCC. High-metastasis cell lines such as MHCC97L
and HCCLM3 exhibit high AR expression, whereas epithelial cell
lines including HepG2 and Huh7 show nearly undetectable AR protein
levels. This expression pattern implies a potential association
between elevated AR levels and HCC invasion and metastasis
(10–12). A recent study further indicated that
high AR expression is also associated with lenvatinib resistance,
whereby in AR-high, α-fetoprotein (AFP)-negative HCC models, AR can
promote angiogenesis, leading to the failure of lenvatinib
monotherapy, while combination therapy with the AR antagonist
bicalutamide has been shown to notably restore its antitumor and
anti-angiogenic effects (11).
However, this mechanism primarily focuses on vascular
microenvironment remodeling. Whether AR drives lenvatinib
resistance through other pathways remains to be elucidated.
Therefore, the present study investigated, from the perspective of
circRNA, the transcriptional regulatory role of AR on circRNAs,
with the aim of revealing a novel molecular mechanism underlying
AR-high-mediated lenvatinib resistance.
Bioinformatic analysis has suggested that AR may be
a potential transcriptional regulator of host genes for a number of
differentially expressed circRNAs in HCC. Among them,
hsa_circ_0011385 and hsa_circ_0001955 have been demonstrated to be
highly expressed in HCC, promoting HCC cell proliferation and
metastasis (13–16). miRNA-212-5p (miR-212-5p), a
potential target of hsa_circ_0011385 downregulated in hepatitis B
virus-associated HCC, is considered an effective biomarker for
early HCC diagnosis and has also been shown to inhibit HCC
progression. Akt is at the hub of the PI3K/Akt/mTOR network and is
hyperactivated by receptor tyrosine kinases, including EGFR, MET
and FGFR, thereby accelerating hepatocellular carcinoma
progression. Furthermore, sustained Akt activation is a key driver
of acquired lenvatinib resistance and Akt inhibitors can reverse
this resistance (17–19). Akt3, a key Akt isoform potentially
controlled by miR-212-5p, has been repeatedly implicated in HCC
progression (20,21).
Based on these findings, the present study
hypothesized that AR, through its transcription factor function,
may regulate the transcription of EIF3I, which generates
hsa_circ_0011385 and thus alter the level of this circRNA.
Hsa_circ_0011385 sequesters miR-212-5p, thereby easing the
repression of the downstream oncogene Akt3 and ultimately promoting
lenvatinib resistance in HCC. The present study aimed to validate
this molecular mechanism using the AR-high cell line MHCC97H as the
research model.
Materials and methods
Bioinformatic analysis
Raw circRNA expression matrices for HCC were
downloaded from the Gene Expression Omnibus (GEO) database
(https://www.ncbi.nlm.nih.gov/geo) by
using ‘circRNA’ and ‘hepatocellular carcinoma’ as search terms. Two
GEO datasets, GSE97332 (circRNA expression in HCC and matched
non-tumor tissues) (22) and
GSE242797 (construct a ceRNA regulatory network to explore
potential pathogenesis and therapy options of human hepatocellular
carcinoma) (23), were used for
differential expression analysis. The names of the circRNAs were
standardized according to the circBase database (http://www.circbase.org/). The differential expression
of circRNAs was analysed by the R package ‘limma’ (version 4.2.2;
Posit Software, PBC) with log2 (fold-change) >1.5 and P<0.05
(24). Host gene promoter sequences
were obtained from the University of California, Santa Cruz
database (https://genome.ucsc.edu/) and
subsequently submitted to the JASPAR database (https://jaspar.elixir.no/) to predict potential AR
binding sites. The Cancer Genome Atlas (TCGA) database (https://www.cancer.gov/tcga) data were analyzed using
the University of Alabama at Birmingham Cancer data analysis portal
online tool (http://ualcan.path.uab.edu) to assess differential
gene expression. The median expression value was used as the cutoff
to define high- and low-expression groups.
Cell culture and transfection
THLE-2 normal hepatocytes and liver cancer cell
lines MHCC97H, HCCLM3 and HepG2 were obtained from iCell Bioscience
Inc. Short tandem repeat profiling was performed externally by the
cell supplier to confirm the identity of each line, and all tested
negative for Mycoplasma. Using DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.), cells were kept at 37°C with 5% CO2
in a humidified incubator. For overexpression, the hsa_circ_0011385
sequence was cloned into the pCD2.1-ciR circRNA expression vector
(Guangzhou Geneseed Biotech. Co., Ltd.) using 50 ng of vector per
ligation reaction and a 3:1 molar ratio of insert to vector, as
recommended by standard protocols. Cells transfected with the
corresponding empty vector were used as the negative control in
these experiments. Small interfering RNA (siRNA), overexpression
plasmids and corresponding control vectors were manufactured by
Wuhan GeneCreate Biological Engineering Co., Ltd. Lipofectamine™
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) was used for cell
transfection in complete compliance with the manufacturer's
instructions. The sequences used were as follows: AR-si1 sense:
5′-GGUUCAACAGUCCACAATT-3′, antisense: 5′-UUGGUGAGUGGUAGAAGCTT-3′;
AR-si2 sense: 5′-CUGAUCUGUGGAGAUGAATT-3′, antisense:
5′-UCAUCUCACAGAUCAGTT-3′; AR-si3 sense: 5′-GCAAGGUUGUGCUAGUATT-3′,
antisense: 5′-UACUAGCACAAGCUUGCTT-3′; si-negative control sense:
5′-UUCUCCGAACGUGUCACGUTT-3′. Among these, si-AR-3 exhibited the
most potent AR knock-down and was therefore selected for all
subsequent functional experiments, which were performed at 48 h
post-transfection.
Establishment of lenvatinib-resistant
cell line
Lenvatinib was purchased from Beyotime
Biotechnology. A lenvatinib-resistant cell line was generated using
a stepwise concentration-increasing method. MHCC97H cells at
logarithmic growth phase were treated with a starting concentration
of ~10 µM (half of the 72-h IC50 value). The
concentration was doubled every 2–3 weeks until reaching 80 µM and
cells were then stably passaged at this concentration for 4 weeks.
Throughout the procedure, cells were maintained in a 5%
CO2 humidified incubator at 37°C, with drug-containing
medium refreshed every 48 h. Cells were passaged upon reaching ~90%
confluence. The entire induction process lasted ~4 months. The
final resistant cell line obtained was named MHCC97H-LR
(lenvatinib-resistant).
Enzalutamide treatment
MHCC97H cells were seeded in 6-well plates at a
density of 3×105 cells per well and cultured overnight. Cells were
then treated with 10 µM enzalutamide (SC0074-10 mM; Beyotime
Biotechnology) at 37°C for 48 h. Control cells were treated with
DMSO (cat. no. ST038; Beyotime Biotechnology) at a final
concentration of 0.1% at 37°C. After 48 h, cells were harvested for
RNA extraction.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Cells from all experimental groups (including normal
hepatocytes, HCC cell lines, lenvatinib-resistant cells and
transfected cells) were lysed for total RNA or miRNA isolation with
the Animal Total RNA Isolation Kit (cat. no. RE-03011; Chengdu Fuji
Biotechnology Co., Ltd.) according to the manufacturer's
instructions and the yield stored at −80°C. miRNA was
reverse-transcribed with the SeqHunt® First Strand cDNA
Synthesis Kit (cat. no. CA01; Seq-Hunt Biotechnology Co., Ltd.).
mRNA and microRNA levels were then quantified by RT-qPCR using 2×
Blue Universal SYBR qPCR Master Mix (cat. no. AF07; Seq-Hunt
Biotechnology Co., Ltd.) with GAPDH and U6 as the respective
internal controls. The thermocycling conditions were as follows:
95°C for 5 min, followed by 40 cycles of 95°C for 15 sec and 60°C
for 40 sec. Expression ratios were calculated using the
2−ΔΔCq method (25) and
primer sequences are provided in Table
I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Primer | Sequence
(5′-3′) |
|---|
| GAPDH-F |
GGAGCGAGATCCCTCCAAAAT |
| GAPDH-R |
GGCTGTTGTCATACTTCTCATGG |
| AR-F |
CCAGGGACCATGTTTTGCC |
| AR-R |
CGAAGACGACAAGATGGACAA |
|
hsa_circ_0011385-F |
ATAGTGCCAAGGAAAGC |
|
hsa_circ_0011385-R |
TGTCCGTGGAGAACAT |
|
hsa_circ_0001955-F |
AAATCAGGTGAAGGTC |
|
hsa_circ_0001955-R |
CACATGGTCCAAAGTA |
|
hsa-miR-212-5p-F |
CGCGACCTTGGCTCTAGACTG |
|
hsa-miR-212-5p-R |
AGTGCAGGGTCCGAGGTATT |
| hsa-miR-212-5p
RT |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGTAAG |
| U6-F |
CTCGCTTCGGCAGCACA |
| U6-R |
AACGCTTCACGAATTTGCGT |
| Akt3-F |
AATGGACAGAAGCTATCCAGGC |
| Akt3-R |
TGATGGGTTGTAGAGGCATCC |
| EIF3I-F |
CTCAAGACCAATTCGGCTGTC |
| EIF3I-R |
CTGGTAGCCCATCTGCTTGTC |
| CSNK1G1-F |
CCATCACAACAGCAGCCTCTTC |
| CSNK1G1-R |
CTTCCTCCACTACCTCCACCTC |
Cell Counting Kit-8 (CCK-8)
proliferation assay
To perform this assay, an optimal density of cells
was seeded into 96-well plates and incubated overnight at 37°C
under 5% CO2 to ensure adherence. Following a 24-h
exposure to 100 µl serially diluted drug (under identical culture
conditions), cell viability was assessed. Specifically, 10 µl CCK-8
reagent (cat. no. CK04; Dojindo Laboratories, Inc.) was introduced
per well and the plates were returned to the 37°C incubator for a 1
to 2-h development period. The reaction was then stopped by
immediate mixing after adding 10 µl stop solution to each well.
Finally, absorbance at 450 nm was recorded using a microplate
reader (TMR-100; Tuoh Electromechanical Technology Co., Ltd.).
Transwell migration assay
Following digestion and harvesting, cells were
resuspended in serum-free DMEM (Gibco; Thermo Fisher Scientific,
Inc.) for subsequent counting. A cellular suspension containing
1×104 cells in 100 µl was applied to the upper chamber
of a Transwell insert. Meanwhile, the lower compartment received
600 µl of complete medium supplemented with 10% FBS. After a 24-h
incubation at 37°C and 5% CO2, the insert was carefully
withdrawn. Stationary cells remaining on the upper membrane surface
were then cleared away. Subsequent steps for the migrated cells on
the lower membrane involved sequential processing: First, fixation
in 4% paraformaldehyde (15 min), followed by staining with crystal
violet (20 min), both at room temperature, followed by two washes
with PBS (cat. no. BL302; Biosharp Life Sciences). Finally, the
membranes were examined and photographed using an inverted MI40
microscope (Guangzhou Micro-Shot Technology Co., Ltd).
Flow cytometric analysis of
apoptosis
Apoptosis was detected by flow cytometry using the
Annexin V-FITC/PI double-staining method, with experimental
procedures strictly following the kit instructions (cat. no.
KTA0004; Abbkine, Inc.). Briefly, after discarding the cell culture
medium and collecting the supernatant, cells were gently washed
twice with pre-chilled PBS. Digestion was performed using 0.25%
EDTA-free trypsin, which was then terminated by adding complete
medium containing 10% FBS, followed by gentle pipetting to obtain a
single-cell suspension. The suspension was centrifuged at 120 × g
for 5 min at room temperature. After discarding the supernatant,
the cell pellet was resuspended in pre-chilled 1× PBS and washed
twice again. Subsequently, cells were resuspended in pre-chilled 1×
Annexin V binding buffer and adjusted to a density of
1×106 cells/ml. A 100 µl aliquot of the cell suspension
was taken and 5 µl Annexin V-FITC and 10 µl PI staining solution
were added sequentially, followed by incubation at room temperature
in the dark for 15 min. Finally, 400 µl binding buffer was added to
each tube and samples were immediately analyzed using a flow
cytometer (CytoFLEX; Beckman Coulter, Inc.). Data analysis was
performed using CytExpert software (version 2.5; Beckman Coulter,
Inc.).
Dual-luciferase reporter assay
For hsa_circ_0011385 and the Akt3 3′-untranslated
region (UTR), luciferase reporters carrying either wild-type (WT)
or mutant (MT) fragments were generated and named circ_0011385-WT,
circ_0011385-MT, Akt3-WT and Akt3-MT, respectively. All constructs
were sequence-verified prior to use. Subsequently, 293T cells were
co-transfected with these reporters together with hsa-miR-212-5p
mimics using the same transfection protocol as aforementioned
(Lipofectamine 2000; Invitrogen; Thermo Fisher Scientific, Inc.);
48 h later, firefly and Renilla activities were quantified
with the Dual-Luciferase Reporter Assay Kit (cat. no. JKR23008;
Wuhan GeneCreate Biological Engineering Co., Ltd.). The miR-212-5p
mimics (sense: 5′-ACCUUGGCUCUAGACUGCUUACU-3′, antisense:
5′-AGUAAGCAGUCUAGAGCCAAGGU-3′) and NC mimics (sense:
5-UCACAACCUCCUAGAAAGAGUAGA-3′, antisense:
5′-UCUACUCUUUCUAGGAGGUUGUGA-3′) were utilized in the present
study.
Chromatin immunoprecipitation-qPCR
(ChIP-qPCR)
Chromatin immunoprecipitation was carried out using
the ChIP Kit (cat. no. JKR23002A; Wuhan GeneCreate Biological
Engineering Co., Ltd.). After 10 min cross-linking with 1%
formaldehyde, the cross-linking reaction was quenched through a
5-min incubation with 0.125 M glycine at room temperature.
Subsequent to two washes with PBS, cells were pelleted by
centrifugation at 1,000 × g for 5 min at 4°C and rapidly frozen at
−80°C. For chromatin lysis, cells were resuspended in Lysis Buffer
provided in the ChIP Kit (cat. no. JKR23002A; Wuhan GeneCreate
Biological Engineering Co., Ltd.), supplemented with 1X Protease
Inhibitor and 5 µl DTT according to the manufacturer's protocol. A
10-min lysis step was performed on ice. After centrifugation at
12,000 × g for 10 min at 4°C to pellet debris and remove the
supernatant, the pellet was subjected to a second lysis and
sonication. Chromatin fragments were then incubated with either AR
antibody (cat. no. 22089-1-AP; Proteintech Group, Inc.) or control
immunoglobulin G (normal rabbit IgG; cat. no. F040301; Beyotime
Biotechnology), both used at a final concentration of 1 µg/ml,
overnight at 4°C. After elution and reversal of cross-links (65°C,
overnight), DNA was purified according to the kit protocol and
analysed by RT-qPCR with primers specific for the EIF3I promoter
region. The primer sequences used were as follows: Primer 1:
forward 5′-TGCCAGGCACTCTCCTAGAT-3′ and reverse
5′-GTTGGTTTGAAGCCTGGCAG-3′; primer 2: forward
5′-CTGCCAGGCTTCAAACCAAC-3′ and reverse 5′-GTTTGAGGCCACCTGGAAGA-3′;
and primer 3: forward 5′-CACAGAGACGCACCTCAGTAT-3′ and reverse
5′-TGACTCACTCGTCTGCATTC-3′.
Western blotting
Western blotting was performed as follows: Proteins
were extracted from cells using RIPA buffer (cat. no. BL504A;
Biosharp Life Sciences), quantified by BCA (BL521A; Biosharp Life
Sciences) and 20 µg was denatured and separated by 10% SDS-PAGE
(80/120 V). Proteins were wet-transferred (200 mA; 90 min; ice
bath) to PVDF membranes (0.22 µm). After blocking (5% skimmed milk
in TBST with 0.1% Tween-20; 1 h; room temperature), membranes were
probed with primary antibodies (1:1,000-1:50,000 in 1% BSA-PBST;
4°C; overnight) and HRP-secondary antibodies (1:10,000 in 5% skim
milk-PBST; 1 h; room temperature), with PBST washes between steps.
Detection was conducted with the SuperKine™ West Pico PLUS
Chemiluminescent Substrate (cat. no. BMU101-CN; Abbkine, Inc.; 1.5
min) and a ChemiScope 6100T imager (Clinx Science Instruments Co.,
Ltd.). Detailed information on the antibodies used is provided in
Table II.
| Table II.Antibodies used for western blotting
analysis. |
Table II.
Antibodies used for western blotting
analysis.
| Target/purpose | Cat. no. | Supplier | Dilution |
|---|
| AR | 22089-1-AP | Proteintech Group,
Inc. | 1:5,000 |
| Akt3 | 21641-1-AP | Proteintech Group,
Inc. | 1:1,000 |
| EIF3I | 11287-1-AP | Proteintech Group,
Inc. | 1:1,000 |
| GAPDH (loading
control) | 60004-1-Ig | Proteintech Group,
Inc. | 1:50,000 |
| HRP-conjugated Goat
Anti-Rabbit IgG | A21020 | Abbkine, Inc. | 1:10,000 |
| HRP-conjugated Goat
Anti-Mouse IgG | A21010 | Abbkine, Inc. | 1:10,000 |
Statistical analysis
After determining parametric assumptions (normality
by Shapiro-Wilk test and homogeneity of variances by Levene's
test), pairwise comparisons were analyzed using unpaired Student's
t tests. Multi-factor datasets were analyzed using one-way ANOVAs
followed by Tukey's multiple-comparison tests, except for CCK-8
data which were analyzed by two-way ANOVA followed by Tukey's test.
Data are presented as the mean ± SD from at least three independent
experiments. P<0.05 was considered to indicate a statistically
significant difference. All statistical analyses were performed
using GraphPad Prism (version 7.0; Dotmatics), R software (version
4.2.2; Posit Software, PBC) or SPSS Statistics (version 24; IBM,
Corp.).
Results
Bioinformatic screening identifies
AR-regulated candidate circRNAs
Preliminary bioinformatic analysis of GEO microarray
datasets (GSE97332 and GSE242797) identified 16 significantly
upregulated and 16 significantly downregulated circRNAs in HCC
tissues compared with paired normal tissues (log2 FC
>1.5; P<0.05), visualized as a heatmap and volcano plot
(Fig. 1A). Analysis of TCGA data
revealed high expression of two circRNAs previously implicated in
HCC (13–16) along with their corresponding host
genes: hsa_circ_0001955 [casein kinase 1 γ-1 (CSNK1G1)] and
hsa_circ_0011385 (EIF3I; Fig. 1B).
RT-qPCR analysis of AR expression in the same cell lines showed
that, compared with THLE-2 normal hepatocytes, MHCC97H and HCCLM3
cell lines expressed significantly higher AR mRNA levels, whereas
HepG2 cells exhibited the lowest AR expression (Fig. 1C). RT-qPCR determined that, relative
to normal THLE-2 hepatocytes, hsa_circ_0001955 (CSNK1G1) and
hsa_circ_0011385 (EIF3I) exhibited markedly elevated levels in
MHCC97H and HCCLM3 cells, whereas HepG2 cells exhibited only faint
expression (Fig. 1D).
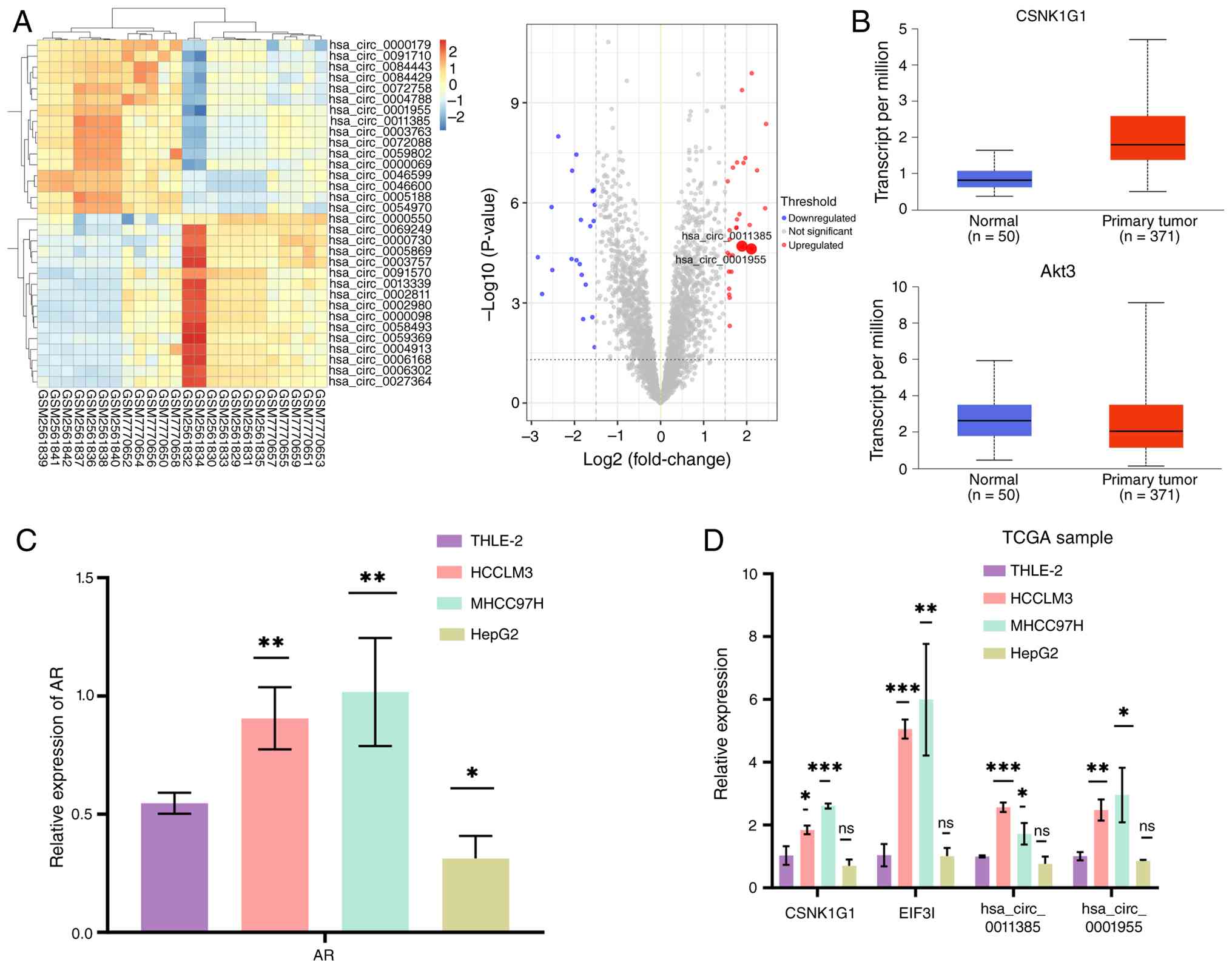 | Figure 1.Identification of AR-regulated
candidate circRNAs in HCC. (A) Heatmap and volcano plot
illustrating circRNA expression changes in HCC relative to matched
non-tumour specimens deposited under Gene Expression Omnibus:
GSE242797 and GSE97332. The volcano plot highlights key circRNAs
(hsa_circ_0011385 and hsa_circ_0001955) with larger circles.
Thresholds for significance were set at log2 (fold-change) >1.5
and adjusted P<0.05. Red, green and gray dots represent
significantly upregulated, downregulated and non-significant
circRNAs, respectively. (B) Analysis of TCGA data shows high
expression of circRNAs hsa_circ_0001955 (host gene: CSNK1G1) and
hsa_circ_0011385 (host gene: EIF3I) in HCC. (C) RT-qPCR analysis of
AR mRNA expression in THLE-2 normal hepatocytes and HCC cell lines
(HepG2, MHCC97H and HCCLM3). (D) RT-qPCR determined the abundance
of hsa_circ_0001955 and hsa_circ_0011385 in THLE-2 normal
hepatocytes compared with the HCC models MHCC97H, HCCLM3 and HepG2.
Data are presented as the mean ± SD from three independent
experiments. *P<0.05, **P<0.01 and ***P<0.001 vs. THLE-2.
AR, androgen receptor; circRNA, circular RNA; HCC, hepatocellular
carcinoma; TCGA, The Cancer Genome Atlas; RT-qPCR, reverse
transcription-quantitative PCR; CSNK1G1, casein kinase 1 γ-1; ns,
not significant; LIHC, liver hepatocellular carcinoma; EIF3I,
eukaryotic translation initiation factor 3 subunit I. |
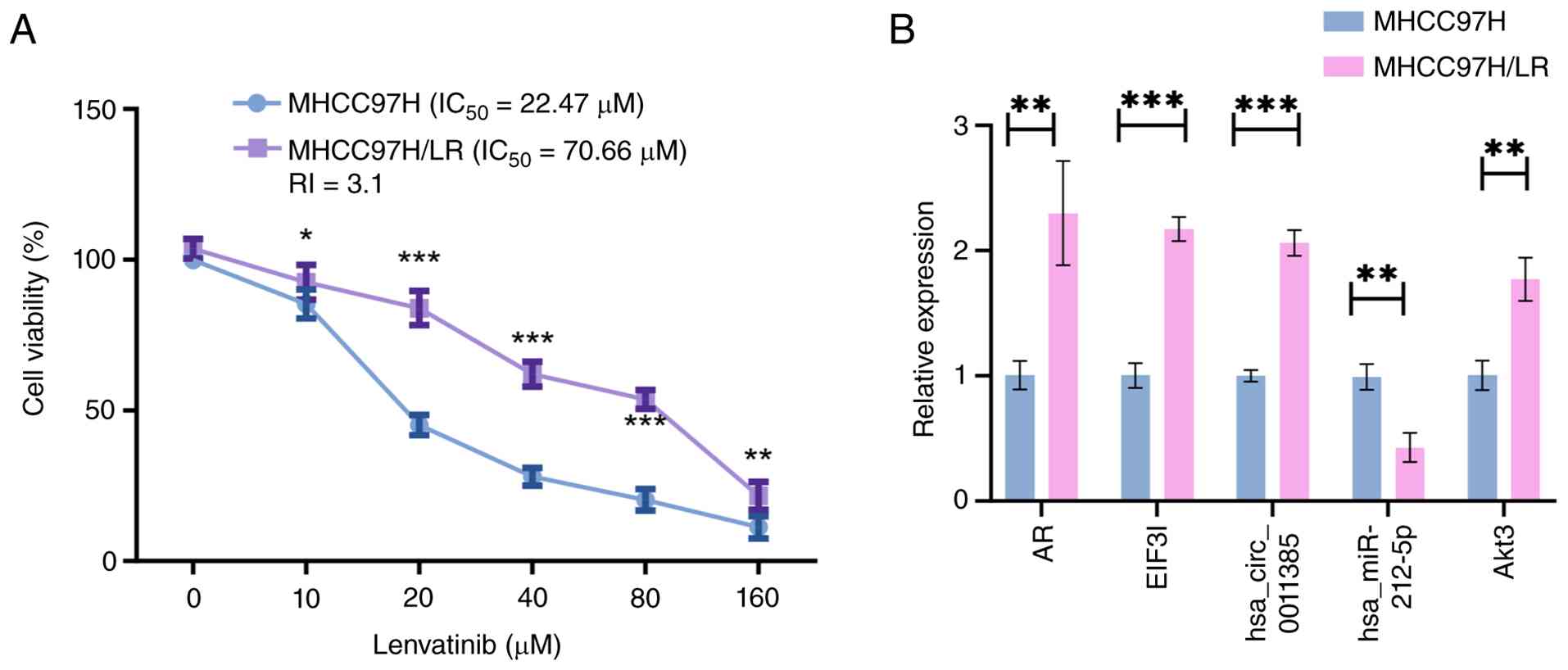
Establishment of lenvatinib-resistant
cells and expression changes in the
AR/hsa_circ_0011385/miR-212-5p/Akt3 axis
MHCC97H-LR, a lenvatinib-resistant HCC derivative,
was obtained by stepwise escalation of drug concentration. Relative
to the parental population, the selected line exhibited markedly
reduced lenvatinib sensitivity: IC50 increased from
19.7–25.7 to 63.0–79.6 µM, giving a 3.1-fold resistance index
(Fig. 2A). In these resistant
cells, AR, Akt3 and hsa_circ_0011385 (EIF3I) were upregulated,
while hsa-miR-212-5p expression was downregulated (Fig. 2B).
Expression changes in the
hsa_circ_0011385/miR-212- 5p/Akt3 axis following AR inhibition
RT-qPCR showed that compared with the THLE-2 cell
line, MHCC97H cells exhibited significantly higher expression of
AR, hsa_circ_0011385, its host gene EIF3I and the downstream target
gene Akt3, while hsa-miR-212-5p expression was significantly lower
(Fig. 3A). RT-qPCR demonstrated an
inhibitory effect after AR siRNA silencing (Fig. 3B) and showed that AR knockdown in
MHCC97H cells led to significant downregulation of Akt3 and
hsa_circ_0011385 (EIF3I) expression and significant upregulation of
hsa-miR-212-5p expression (Fig.
3C). Western blotting analysis determined significantly higher
protein expression of AR, EIF3I and Akt3 in MHCC97H cells compared
with THLE-2 cells and demonstrated that AR knockdown relatively
reduced EIF3I and Akt3 protein levels (Fig. 3D).
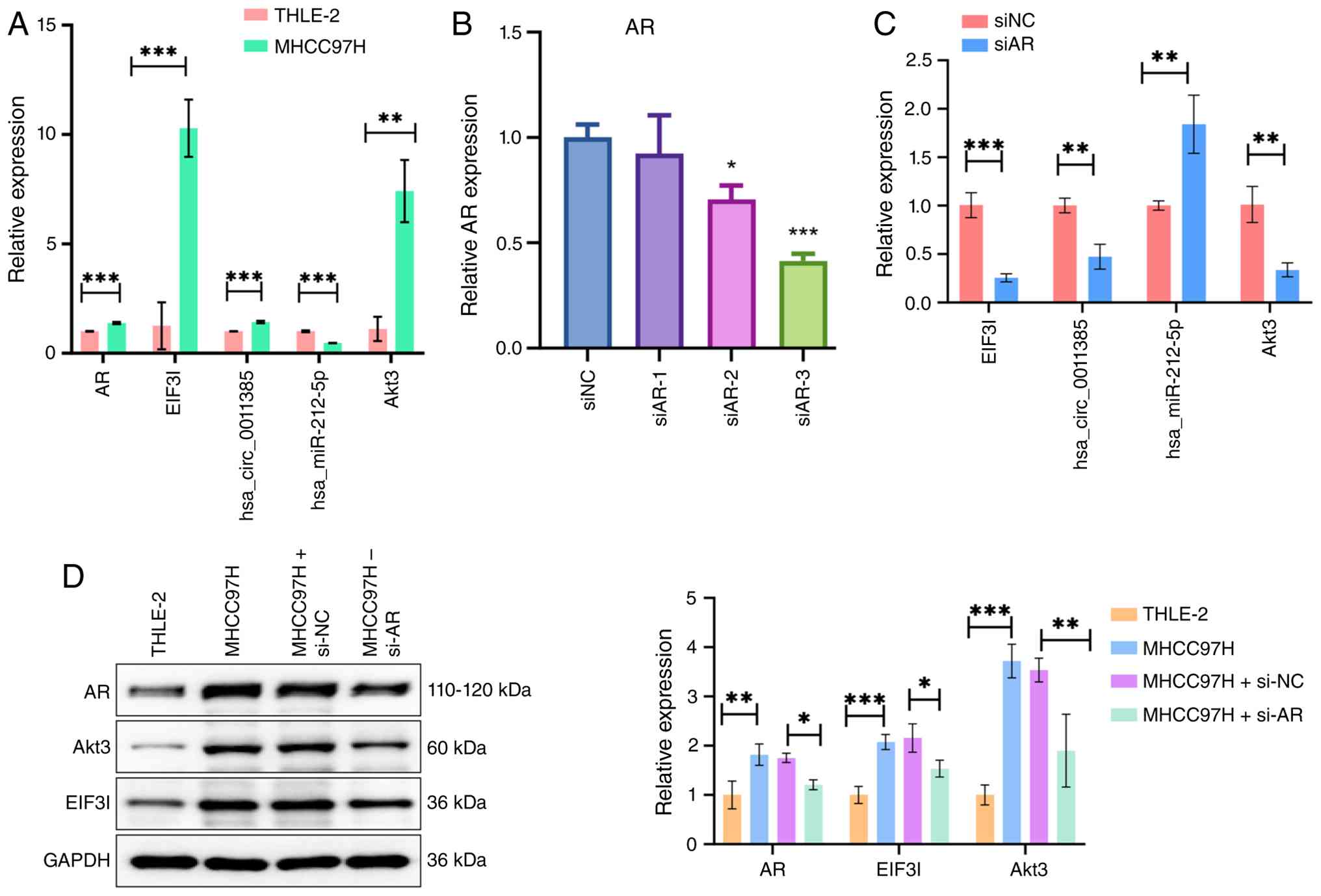 | Figure 3.AR modulates the expression of the
hsa_circ_0011385/miR-212-5p/Akt3 axis. (A) RT-qPCR analysis of AR,
EIF3I, hsa_circ_0011385, miR-212-5p and Akt3 expression in THLE-2
and MHCC97H cells. (B) RT-qPCR determining efficient knockdown of
AR by siRNA in MHCC97H cells. (C) RT-qPCR demonstrating the effects
of AR knockdown on the expression of hsa_circ_0011385 (EIF3I),
miR-212-5p and Akt3. (D) Western blotting analysis of AR, EIF3I and
Akt3 protein levels in THLE-2, MHCC97H and AR-knockdown MHCC97H
cells. GAPDH served as the loading control. Data are presented as
the mean ± SD from three independent experiments. *P<0.05,
**P<0.01 and ***P<0.001. AR, androgen receptor; EIF3I,
eukaryotic translation initiation factor 3 subunit I; RT-qPCR,
reverse transcription-quantitative PCR; miR, microRNA; si/siRNA,
small interfering RNA; NC, negative control. |
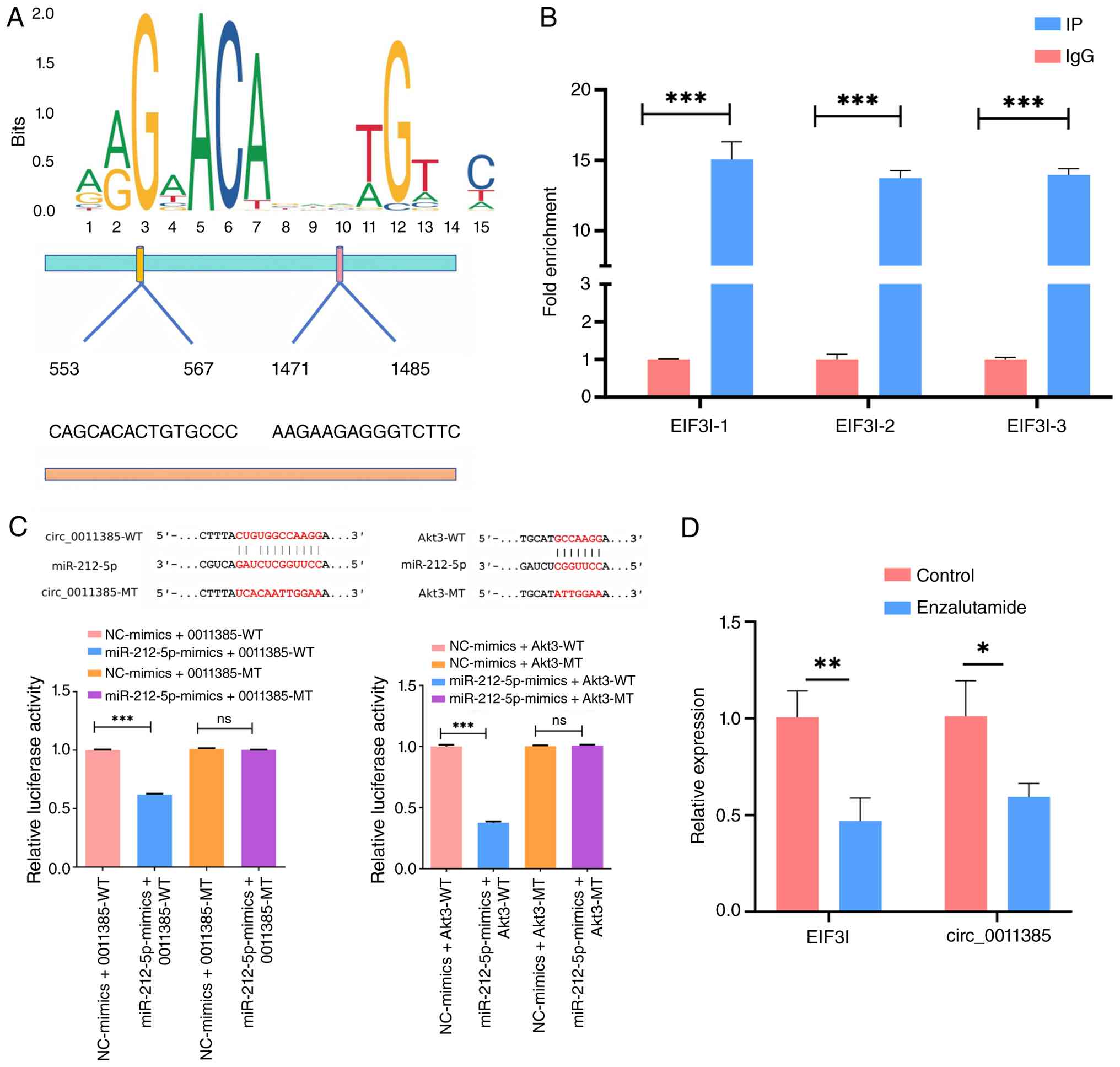
Validation of the targeting regulatory
association within the AR/hsa_circ_0011385/miR-212-5p/Akt3
axis
To define how AR regulates hsa_circ_0011385, the
EIF3I promoter (source of circ_0011385) was scanned in JASPAR for
AR motifs (Fig. 4A). ChIP-qPCR with
three primer pairs verified strong AR enrichment at the predicted
sites (Fig. 4B). Dual-luciferase
assays showed that miR-212-5p directly bound circ_0011385-WT and
Akt3-WT, reducing their reporter signals (Fig. 4C). To validate AR transcriptional
regulation, MHCC97H cells were treated with the AR antagonist
enzalutamide. Enzalutamide significantly reduced EIF3I mRNA and
hsa_circ_0011385 levels compared with DMSO control (Fig. 4D), consistent with the ChIP-qPCR
results. These findings determined a targeted regulatory cascade
within the AR/hsa_circ_0011385/miR-212-5p/Akt3 axis.
Low AR expression attenuates the
biological activity of lenvatinib-resistant MHCC97H cells and
hsa_circ_0011385 overexpression reverses the effects of AR
knockdown on lenvatinib resistance
RT-qPCR validated the overexpression effect of
hsa_circ_0011385 (Fig. 5A). CCK-8
assay results showed that compared with parental MHCC97H cells,
lenvatinib-resistant cells exhibited significantly enhanced
proliferation capacity, which was significantly weakened upon AR
interference (Fig. 5B). Transwell
assays exhibited a similar trend with lenvatinib-resistant cells
displaying strong migration capacity, which was reduced after AR
interference (Fig. 5C). Flow
cytometric apoptosis assays indicated that compared with MHCC97H,
resistant cells exhibited a decreased apoptosis rate, which
significantly increased after AR interference (Fig. 5D). Overexpression of
hsa_circ_0011385 significantly reversed the inhibitory effects
caused by AR knockdown.
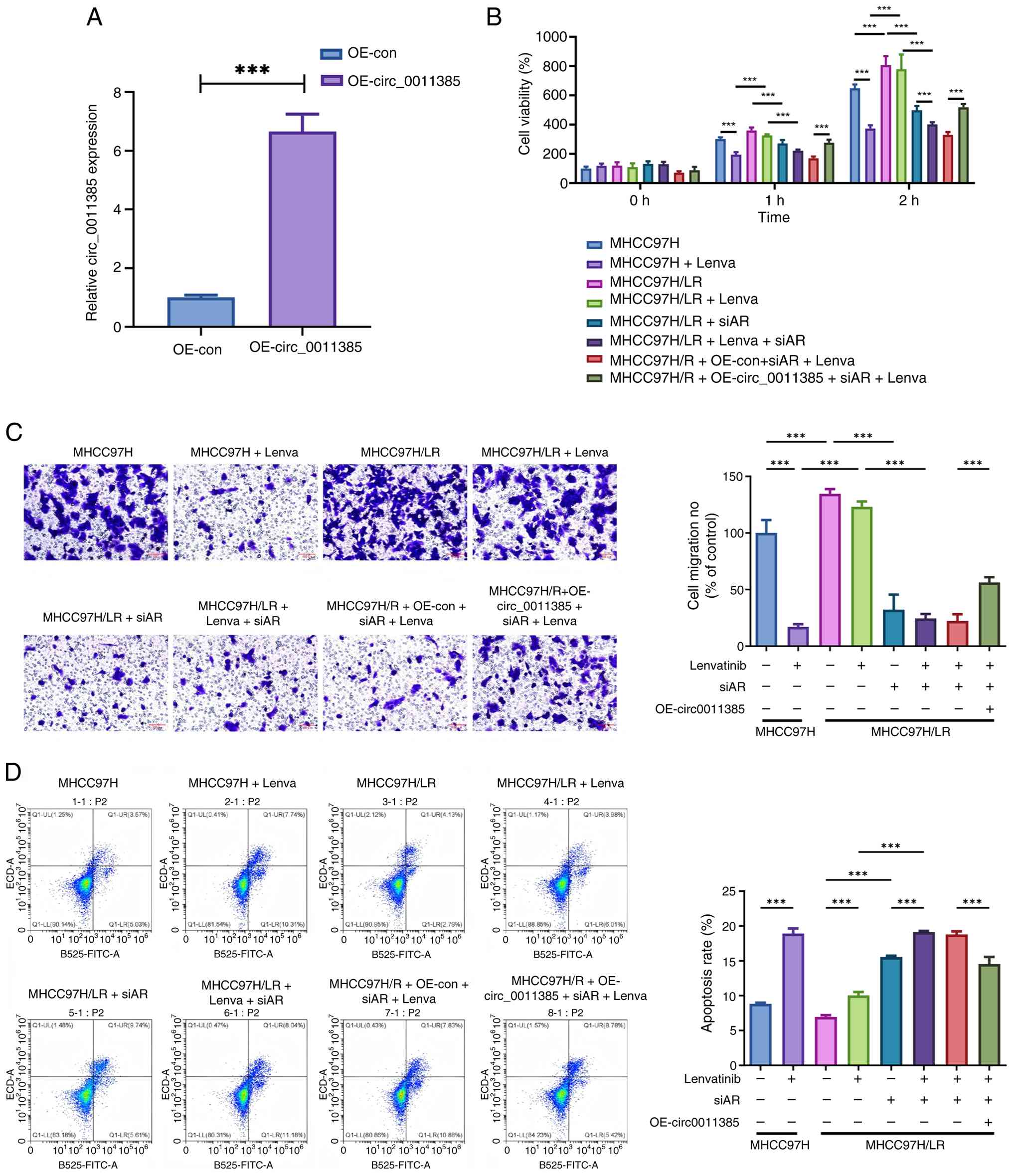 | Figure 5.Functional role of AR and
hsa_circ_0011385 in lenvatinib resistance. (A) Reverse
transcription-quantitative PCR validation of hsa_circ_0011385
overexpression efficiency. Functional assays were performed in the
following groups: MHCC97H parental cells with or without
lenvatinib; MHCC97H-LR cells with or without lenvatinib; MHCC97H-LR
cells transfected with si-AR, with or without lenvatinib; and
MHCC97H-LR cells co-transfected with si-AR and oe-circ (or empty
vector) plus lenvatinib. (B) Cell proliferation assessed by Cell
Counting Kit-8 assay after lenvatinib treatment. (C) Cell mobility
evaluated with Transwell chambers; representative fields (left) and
pooled counts (right) are shown (scale bar, 100 µm). (D)
Flow-cytometric apoptosis profiles (left, scatter plots; right,
summary data). All values are presented as the mean ± SD from three
independent experiments. ***P<0.001. AR, androgen receptor; si,
small interfering; ns, not significant; oe, overexpression; con,
control; circ; circular RNA; Lenva, lenvatinib; LR,
lenvatinib-resistant. |
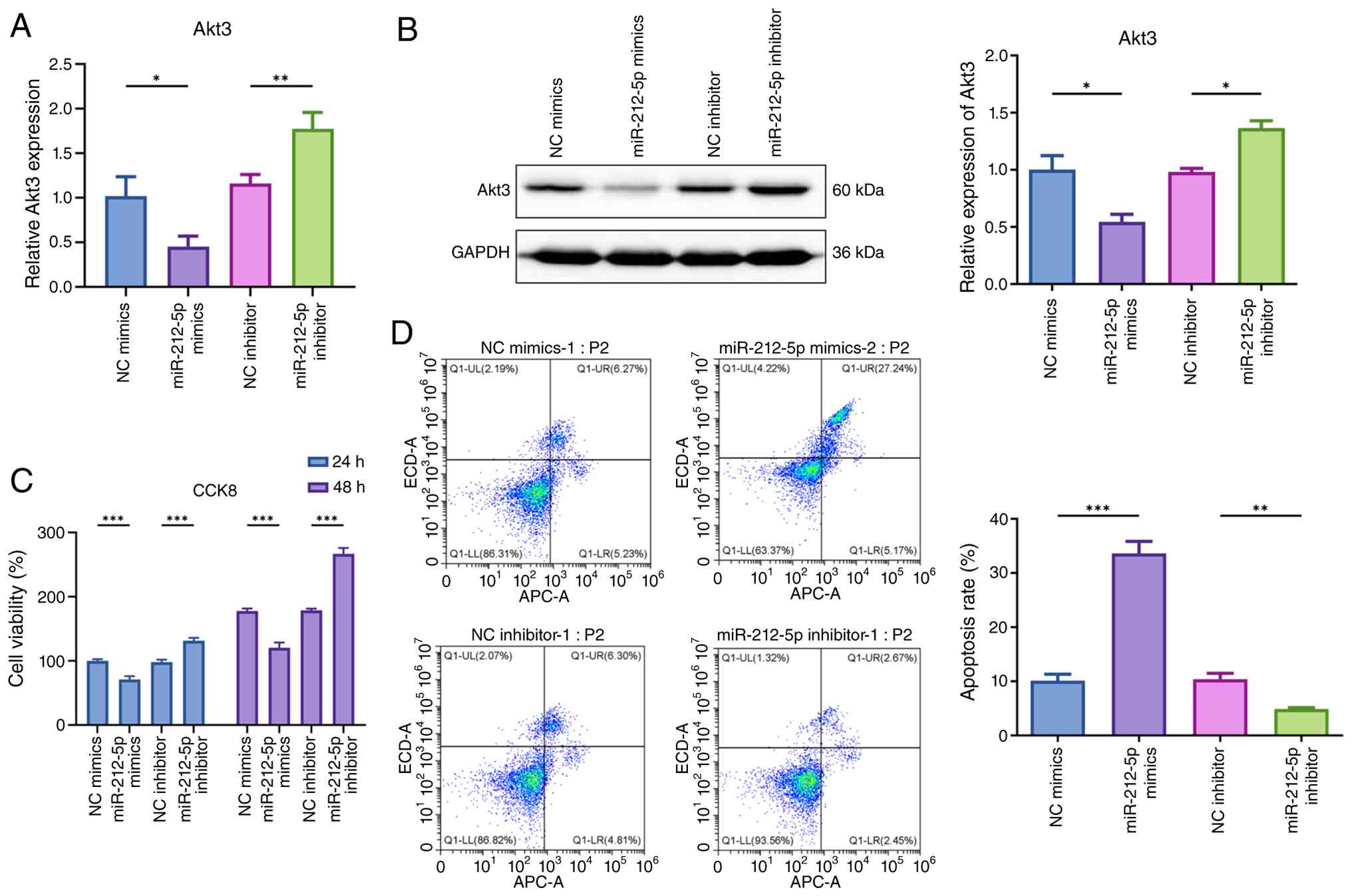
MiR-212-5p directly modulates Akt3
expression and lenvatinib sensitivity in resistant cells
To provide direct functional evidence for the role
of miR-212-5p in lenvatinib resistance, MHCC97H-LR cells were
transfected with miR-212-5p mimic or inhibitor. RT-qPCR and western
blotting analyses showed that miR-212-5p mimic significantly
downregulated Akt3 expression at both mRNA and protein levels,
whereas miR-212-5p inhibitor upregulated Akt3 expression (Fig. 6A and B). The CCK-8 assay
demonstrated that the miR-212-5p mimic suppressed the proliferation
of lenvatinib-resistant MHCC97H-LR cells, while miR-212-5p
inhibitor further enhanced proliferation (Fig. 6C). Flow cytometric apoptosis
analysis revealed that miR-212-5p mimic increased the apoptosis
rate, whereas miR-212-5p inhibitor decreased apoptosis in resistant
cells (Fig. 6D). These results
determined that miR-212-5p directly regulated Akt3 expression and
lenvatinib sensitivity, supporting the proposed competing
endogenous RNA (ceRNA) mechanism.
Discussion
Acquired resistance to lenvatinib is a marked
clinical challenge in the treatment of advanced HCC. Recent studies
have identified AR as a pro-oncogenic factor in HCC, whose
activation contributes to cell proliferation and angiogenesis and
may be associated with lenvatinib resistance (11,26,27).
The present study revealed that in the AR-high HCC subset, AR
drives lenvatinib resistance by directly transcriptionally
upregulating the circRNA hsa_circ_0011385, which then acts as a
‘molecular sponge’ for miR-212-5p, relieving its inhibition of the
downstream oncogene Akt3. This finding provides a new molecular
perspective for understanding the poor prognosis of patients with
high AR expression.
In recent years, the role of circRNAs in HCC drug
resistance has been increasingly uncovered. A number of studies
have determined that circRNAs confer acquired lenvatinib resistance
in HCC by sponging tumor-suppressive miRNAs or acting as scaffolds
to activate pathways such as c-Myc/β-catenin/EGFR, rapidly
rebuilding VEGF-independent angiogenesis and tumor stemness
(7–9,28,29).
These key discoveries collectively established the research
foundation for circRNAs in the field of drug resistance. The
importance of the present study lies not only in identifying
another functionally unknown resistance-associated circRNA, but in
associating the transcriptional regulatory function of AR with
circRNA dysregulation in a specific cellular context, thereby
revealing a previously unclear molecular pathway through which high
AR expression leads to lenvatinib resistance.
In the successfully established lenvatinib-resistant
cell model, the present study demonstrated coordinated
dysregulation of the AR/hsa_circ_0011385/miR-212-5p/Akt3 axis in
resistant cells. At the mechanistic level, the present study
clarified the causal associations within this axis. Upstream,
ChIP-qPCR experiments provided key evidence for the direct binding
of AR to the EIF3I promoter. To the best of our knowledge, the
present study is the first to propose an explanation of the
upregulation mechanism of hsa_circ_0011385 at the transcriptional
initiation level: AR promotes the transcription of its host gene
EIF3I, providing a richer pool of precursor mRNA templates for the
back-splicing of hsa_circ_0011385, which is the primary reason for
the increased expression of this circRNA. The regulation of tumor
progression through the transcription factor activity of AR has
been frequently reported in prostate cancer but is less documented
in HCC (30–32). A study by Zhang et al
(12) involving 142 patients with
HCC found that 37% exhibited nuclear AR upregulation, which was
clearly associated with poor patient prognosis, suggesting that the
transcription factor activity of AR may be an important factor in
HCC progression. Recent research has also indicated that lenvatinib
efficacy is suboptimal in patients with high AR levels but are
AFP-negative, while combination therapy with the AR antagonist
bicalutamide and lenvatinib can notably improve resistance
(11). These conclusions support
the present findings that the transcription factor activity of AR
is closely associated with lenvatinib resistance.
Downstream, dual-luciferase reporter assays
determined the direct targeting associations between
hsa_circ_0011385 and miR-212-5p and between miR-212-5p and the Akt3
3′-UTR, forming a ceRNA regulatory network extending from
transcriptional to post-transcriptional regulation. Functional
experiments with the miR-212-5p mimic and inhibitor further
demonstrated that miR-212-5p suppressed Akt3 expression, inhibited
proliferation and promoted apoptosis in resistant cells, consistent
with its role in the ceRNA axis. The present study indicated that
Akt3 was the key downstream effector molecule in this AR-driven
pathway. Extensive prior research has established the important
role of Akt3, a key component of the PI3K/Akt signaling pathway, in
HCC progression. Its activation has been shown to drive lenvatinib
resistance by regulating a number of downstream substrates such as
mTOR and GSK-3β (19–21,33–35).
The present study did not aim to redundantly validate the detailed
pathways downstream of Akt3 but focused on revealing a novel axis
upstream of Akt3, where AR regulates circRNA through its host gene
EIF3I.
Functionally, the present study found that AR
knockdown significantly reversed the malignant phenotypes of
resistant cells, including inhibiting proliferation and migration
and promoting apoptosis. This beneficial effect was successfully
reversed by exogenous overexpression of hsa_circ_0011385. This
suggested that hsa_circ_0011385, driven by AR and produced through
EIF3I transcription, is a key downstream effector in mediating
lenvatinib resistance and suggests that targeting this specific
node may have therapeutic potential.
The present study exhibited a number of limitations,
the most notable being that the conclusions have not yet been
validated in animal models or larger clinical cohorts. Furthermore,
the mechanisms of lenvatinib resistance are highly heterogeneous
and the AR/circ_0011385 axis may represent only one component. It
is particularly important to note that the mechanism established in
the present study was based on AR-high cell models; therefore, the
present conclusions may primarily apply to the subset of patients
with HCC and high AR expression. Whether this axis functions in
AR-low or AR-negative HCC remains unclear and its interaction with
other known resistance pathways warrants further exploration.
Prospective clinical studies in lenvatinib-treated patients with
HCC are needed to evaluate whether baseline expression levels of
AR, hsa_circ_0011385 or Akt3 associate with treatment response and
survival outcomes.
Despite these limitations, the present findings have
important potential clinical implications. First, they suggest that
AR expression levels and its regulated circRNA could serve as
potential predictive biomarkers for lenvatinib efficacy in AR-high
patients with HCC. Second, interventions targeting this pathway,
such as using AR antagonists or specific oligonucleotides targeting
hsa_circ_0011385, may represent novel strategies to overcome
lenvatinib resistance in this specific patient subset in the
future.
In summary, to the best of our knowledge, the
present study revealed for the first time that in AR-high HCC, the
AR drives the expression of its circRNA product hsa_circ_0011385 by
directly transcriptionally activating its host gene, EIF3I. The
highly expressed hsa_circ_0011385 functions by sequestering
miR-212-5p, thereby alleviating its inhibitory effect on the
downstream target gene Akt3, which ultimately activates
pro-survival signaling pathways and confers lenvatinib resistance.
These findings not only elucidate a complete AR-dominated ceRNA
axis, spanning from transcriptional to post-transcriptional
regulation, but importantly, establish a robust conceptual
framework and highlight promising therapeutic targets. The present
study therefore aids in identifying the subset of AR-high patients
with HCC, predicting their lenvatinib response and devising
combined treatment strategies (such as AR antagonists or
circRNA-targeting therapies) against this pathway.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the Medical
Science and Technology Project of Zhejiang Province (grant no.
2022KY699) and the Chinese Medicine Research Program of Zhejiang
Province (grant no. GZY-ZJ-KJ-24060).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
CL and BZ conceived and designed the present study.
MW, ZW and ZC performed the experiments. CL and YJ conducted the
statistical analyses. BZ critically reviewed and revised the
manuscript. CL, BZ and ZW prepared the figures. CL and BZ confirm
the authenticity of all the raw data. All authors have read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
2
|
Rumgay H, Arnold M, Ferlay J, Lesi O,
Cabasag CJ, Vignat J, Laversanne M, McGlynn KA and Soerjomataram I:
Global burden of primary liver cancer in 2020 and predictions to
2040. J Hepatol. 77:1598–1606. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Park JW, Chen M, Colombo M, Roberts LR,
Schwartz M, Chen PJ, Kudo M, Johnson P, Wagner S, Orsini LS and
Sherman M: Global patterns of hepatocellular carcinoma management
from diagnosis to death: The BRIDGE study. Liver Int. 35:2155–2166.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen R, Hu X, Huang Y, Jiang Y, Chen G,
Shan Q, Xu X and Zheng S: Regulated cell death in lenvatinib
resistance of hepatocellular carcinoma: From molecular mechanisms
to therapeutic strategies. Int J Biol Sci. 21:2012–2026. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ye G, Ye M and Jin X: Roles of clinical
application of lenvatinib and its resistance mechanism in advanced
hepatocellular carcinoma (Review). Am J Cancer Res. 14:4113–4171.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Qin Y, Han S, Yu Y, Qi D, Ran M, Yang M,
Liu Y and Li Y, Lu L, Liu Y and Li Y: Lenvatinib in hepatocellular
carcinoma: Resistance mechanisms and strategies for improved
efficacy. Liver Int. 44:1808–1831. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang L, Ji Y, Ni C, Xu Z, Shen Y, Lu H,
Zhang C and Yang S: EIF4A3-mediated biogenesis of CircFADS1
promotes the progression of hepatocellular carcinoma via
Wnt/β-catenin pathway. Adv Sci (Weinh). 12:e24118692025. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tang Y, Yuan F, Cao M, Ren Y, Li Y, Yang
G, Zhong Z, Liang H, Xiong Z, He Z, et al: CircRNA-mTOR promotes
hepatocellular carcinoma progression and lenvatinib resistance
through the PSIP1/c-Myc Axis. Adv Sci (Weinh). 12:e24105912025.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yuan F, Tang Y, Liang H, Cao M, Ren Y, Li
Y, Yang G, Zhong Z, Xiong Z, He Z, et al: CircPIK3C3 inhibits
hepatocellular carcinoma progression and lenvatinib resistance by
suppressing the Wnt/β-catenin pathway via the miR-452-5p/SOX15
axis. Genomics. 117:1109992025. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dauki AM, Blachly JS, Kautto EA, Ezzat S,
Abdel-Rahman MH and Coss CC: Transcriptionally active androgen
receptor splice variants promote hepatocellular carcinoma
progression. Cancer Res. 80:561–575. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin Z, Liu X, Wang H, Li S, Miao Z, Yang
J, Zhang Y, Lei K, Wu Y, Kang Y, et al: Androgen receptor promotes
arachidonic acid metabolism and angiogenic microenvironment in
AFP-negative hepatocellular carcinoma. Nat Commun. 16:64512025.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H, Li XX, Yang Y, Zhang Y, Wang HY
and Zheng XFS: Significance and mechanism of androgen receptor
overexpression and androgen receptor/mechanistic target of
rapamycin cross-talk in hepatocellular carcinoma. Hepatology.
67:2271–2286. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li X, Lv J, Hou L and Guo X: Circ_0001955
acts as a miR-646 sponge to promote the proliferation, metastasis
and angiogenesis of hepatocellular carcinoma. Dig Dis Sci.
67:2257–2268. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bai K, Ma Y and Li J: Circular RNA
circ_0001955 promotes hepatocellular carcinoma tumorigenesis by
up-regulating alkaline ceramidase 3 expression through
microRNA-655-3p. Bioengineered. 13:2099–2113. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fu D, Ji Q, Wang C, Yu L and Yu R: Aloin
decelerates the progression of hepatocellular carcinoma through
circ_0011385/miR-149-5p/WT1 axis. Cell Cycle. 20:2476–2493. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ni C, Yang S, Ji Y, Duan Y, Yang W, Yang
X, Li M, Xie J, Zhang C, Lu Y and Lu H: Hsa_circ_0011385 knockdown
represses cell proliferation in hepatocellular carcinoma. Cell
Death Discov. 7:2702021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hou W, Bridgeman B, Malnassy G, Ding X,
Cotler SJ, Dhanarajan A and Qiu W: Integrin subunit beta 8
contributes to lenvatinib resistance in HCC. Hepatol Commun.
6:1786–1802. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tian LY, Smit DJ and Jücker M: The role of
PI3K/AKT/mTOR signaling in hepatocellular carcinoma metabolism. Int
J Mol Sci. 24:26522023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Z, Dou X, Shan Q, Ning Y, Wang J,
Wang T, Cheng T, Shi K, Li S, Han X and Cao G: Targeting AKT to
treat liver disease: Opportunities and challenges. Biochem
Pharmacol. 242:1172082025. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shu G, Su H, Wang Z, Lai S, Wang Y, Liu X,
Dai L, Bi Y, Chen W, Huang W, et al: LINC00680 enhances
hepatocellular carcinoma stemness behavior and chemoresistance by
sponging miR-568 to upregulate AKT3. J Exp Clin Cancer Res.
40:452021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang F, Lin Y, Du J, Qiu S, Luo G and
Zhang Z: The transcription factor TBP promotes hepatocellular
carcinoma progression by activating AKT3. Am J Cancer Res.
13:5656–5666. 2023.PubMed/NCBI
|
|
22
|
Han D, Li J, Wang H, Su X, Hou J, Gu Y,
Qian C, Lin Y, Liu X, Huang M, et al: Circular RNA circMTO1 acts as
the sponge of microRNA-9 to suppress hepatocellular carcinoma
progression. Hepatology. 66:1151–1164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Dong Z, Chen W, Chen L, Ju L, Cai
W, Luo X and Bian Z: Construction of a ceRNA regulatory network to
explore potential pathogenesis mechanisms involved in human
hepatocellular carcinoma. Sci Rep. 13:220582023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth G: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao J, Fang L, Pu R, Liu W, Cai S, Wang
R, Shi Y, Li Z, Zhang Z, Li Z and Cao G: Androgen receptor-induced
molecules and androgen contribute synergistically to
male-predominance of hepatocellular carcinoma. iScience.
27:1105192024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yavuz M, Takanlou LS, Avcı ÇB and Demircan
T: A selective androgen receptor modulator, S4, displays robust
anti-cancer activity on hepatocellular cancer cells by negatively
regulating PI3K/AKT/mTOR signalling pathway. Gene. 869:1473902023.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei X, Si A, Zhao S, Fu Y, Li J,
Aishanjiang K, Ma Y, Yu C, Yu B, Cui C, et al: CircUCK2(2,3)
promotes cancer progression and enhances synergistic cytotoxicity
of lenvatinib with EGFR inhibitors via activating CNIH4-TGFα-EGFR
signaling. Cell Mol Biol Lett. 30:152025. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Abudoureyimu M, Sun N, Chen W, Lin X, Pan
F and Wang R: Aurora-A promotes lenvatinib resistance
experimentally through hsa-circ-0058046/miR-424-5p/FGFR1 axis in
hepatocellular carcinoma. Int J Immunopathol Pharmacol.
39:39463202513166922025. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Q, Wu Y, Long Y, Li R, Shi Y, Zheng
Y, Chen X, Li X, Zhou Y, Huang X and Jiang G: AR+TREM2+ macrophage
induced pathogenic immunosuppression promotes prostate cancer
progression. Nat Commun. 16:69642025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu P, Yang JC, Chen B, Nip C, Van Dyke JE,
Zhang X, Chen HW, Evans CP, Murphy WJ and Liu C: Androgen receptor
blockade resistance with enzalutamide in prostate cancer results in
immunosuppressive alterations in the tumor immune microenvironment.
J Immunother Cancer. 11:e0065812023. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chukhu M, Dahiya UR and Heemers HV:
Evolving roles for the androgen receptor and its protein
interactome in castration-resistant prostate cancer. Oncogene.
44:3883–3894. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tang Y, Fan S, Peng R, Liu H, Su B, Tu D,
Wang S, Jin X, Jiang G, Jin S, et al: TRIM29 reverses lenvatinib
resistance in liver cancer cells by ubiquitinating and degrading
YBX1 to inhibit the PI3K/AKT pathway. Transl Oncol. 53:1022942025.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang J, Shi J, Mi L, Zhao M, Han G and Yin
F: Aberrant activation of the PI3K/AKT/HIF-1α pathway promotes
glycolysis and lenvatinib resistance in liver cancer. Mol Med Rep.
32:3012025. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tian LY, Smit DJ, Popova NV, Horn S,
Velasquez LN, Huber S and Jücker M: All three AKT isoforms can
upregulate oxygen metabolism and lactate production in human
hepatocellular carcinoma cell lines. Int J Mol Sci. 25:21682024.
View Article : Google Scholar : PubMed/NCBI
|