Introduction
Colorectal cancer is one of the leading causes of
cancer-related death worldwide and is therefore a major public
health concern (1). Although the
causes are not completely understood, dietary factors appear to be
important (2). In a large number of
epidemiological studies, increased consumption of vegetables and
fruits have been consistently associated with a low risk of
colorectal cancer (3–5).
For example, it was reported that cruciferous
vegetables, and particularly broccoli consumption, are linked to a
lowered risk of colon cancer, and the protective effects are
particularly evident in individuals with the glutathione
transferase M1 null genotype (6).
Much attention has been focused on which dietary constituents play
critical roles in cancer preventive actions, and with cruciferous
vegetables the beneficial effects may be attributed, at least in
part, to the high content of isothiocyanates (ITCs). ITCs are
abundant in cruciferous vegetables such as broccoli, horseradish,
mustard, watercress, cabbage or cauliflower. Interestingly, some of
the purified products have been shown to possess potent
anticarcinogenic properties in cell culture models as well as in
experimental animal models. Inhibitory influence has been
documented for rat lung, esophagus, mammary gland, liver, small
intestine, colon and bladder tumorigenesis (7,8).
Although the ultimate causes of the preventive effects have yet to
be defined in detail, enhanced detoxification of carcinogens (phase
II enzyme activation) as well as blocking carcinogen activation
(phase I enzyme inhibition) are presumably involved (9).
Various ITCs have been shown to inhibit cytochrome
P450 (CYP) and increase the carcinogen excretion or detoxification
by phase II detoxification enzymes such as glutathione
S-transferase (GST) and NAD(P)H: quinone reductase (QR) (10–13). A
typical Japanese condiment, wasabi (Wasabia japonica) was
considered as a possible source of antimutagenic and anticancer
agents (14,15). An extract was found to reduce the
growth of the human monoblastic leukemia cells by inducing
apoptosis (16). One particular
ingredient, 6-methylsulfinylhexyl isothiocyanate (6-MSITC), was
found to inhibit cell proliferation in human breast cancer and
melanoma cell lines (17).
Morimitsu et al reported that 6-MSITC activates the
ARE/Nrf2-dependent detoxification pathway, and oral administration
resulted in the induction of hepatic GST and QR in mouse liver
(18). Nrf-2 also regulated the
detoxification enzyme, UDP-glucuronyltransferase (UDPGT), in the
mouse (19).
The present bioassay, using rats, was conducted to
determine whether dietary 6-MSITC can modulate carcinogen-induced
colon carcinogenesis. Thus, the incidences of two different mucosal
lesions, aberrant crypt foci (ACF) and β-catenin-accumulated crypts
(BCAC) as surrogate endpoints in the place of tumors were
evaluated. In addition, to assess whether 6-MSITC affects cell
proliferation activity in colonic preneoplastic lesions, the
proliferating cell nuclear antigen (PCNA) index was evaluated.
Protein levels of CYP enzymes and the activity of UDPGT in the
liver were also measured.
Materials and methods
Animals, chemicals and diets
All animal studies were carried out under the
Regulations for Animal Experiments at Gifu University. Male F344
rats (Shizuoka Laboratory Animal Center, Shizuoka, Japan) aged 4
weeks were housed in wire cages (three or four rats per cage). The
rats had free access to drinking water and diets, under controlled
conditions of humidity (50±10%), lighting (12-h light/dark cycle)
and temperature (23±2°C). The rats were quarantined for 7 days and
then randomized into experimental and control groups. Powdered CE-2
(Clea Japan, Tokyo, Japan) diet was used as the basal diet
throughout the study. 6-MSITC (purity >99%) was kindly donated
by Kinjirushi Co., Ltd. (Nagoya, Japan) and incorporated into the
experimental diet at concentrations of 200 and 400 ppm. The diets
were stored in a cold room (4°C) until use. 1,2-Dimethylhydrazine
(DMH) was obtained from Tokyo Kasei Kogyo Co., Ltd. (Tokyo, Japan)
and administered by subcutaneous (s.c.) injection (40 mg/kg)
between 10:00 and 11:00 am to induce colorectal preneoplastic
lesions.
Experiment 1
A total of 66 male F344 rats were divided into seven
groups. Groups 1–5 were initiated with DMH by four weekly s.c.
injections (40 mg/kg body weight). Rats in groups 2 and 3 were fed
the diet containing 200 and 400 ppm of 6-MSITC, respectively, for 5
weeks, starting 1 week before the first dosing of DMH until the end
of week 5. The rats were then switched to and maintained on the
basal diet until the termination. Starting 1 week after the
cessation of DMH treatment, rats in groups 4 and 5 were fed the
diet mixed with 200 and 400 ppm 6-MSITC, respectively, and this was
continued until the termination. Rats in group 6 were fed the diet
containing 400 ppm 6-MSITC throughout the experiment and group 7
served as the untreated control. At termination, all animals were
sacrificed to assess the incidences of ACF and BCAC in the large
bowel. Tissues were embedded in paraffin blocks and processed for
routine histological observation with hematoxylin and eosin
(H&E) staining.
Identification of ACF
After fixation of resected colons in 10% buffered
formalin for at least 24 h at room temperature, the colons were
stained using 0.5% methylene blue (in saline) for 1–3 min. After
staining, ACF were counted and recorded according to the procedure
described by Bird (20). The size
of the lesions was scored as crypt multiplicity (ACF/lesion). In
this study, we divided the colon into three portions after excision
of the caecum (proximal, middle and distal) and used all of the
segments for the analysis (21).
Identification of BCAC
After ACF counting, colon tissues were embedded in
paraffin and processed for immunohistochemistry of BCAC. Briefly,
serial sections (4-μm thick) were prepared to include the middle
portion between the surface and the bottom of the crypt. Some of
these sections were used for immunohistochemistry of β-catenin and
routine H&E staining. Immunohistochemistry was performed after
exposure to 3% H2O2 for 20 min to block the
endogenous peroxidase activity using a primary antibody to the
β-catenin protein (1:1000 dilution; Transduction Laboratories,
Lexington, KY, USA) at room temperature for 60 min. Then, the
sections were stained with an LSAB kit (Dako, Glostrup,
Denmark).
PCNA immunohistochemistry
To assess the proliferative activity of cells in ACF
and BCAC, immunohistochemical staining for PCNA was performed by
using anti-PCNA (1:200 dilution) antibody and an LSAB kit (both
from Dako). The nuclei that densely immunoreacted with PCNA were
regarded as PCNA-positive. PCNA labeling indices were determined by
counting the number of positive cells among at least 200 epithelial
cells in ACF and BCAC, and were indicated as percentages.
Experiment 2
Nine male F344 rats were divided into three groups.
At 6 weeks of age, rats in group 1 were given corn oil through an
intragastric tube and served as a control. Animals in groups 2 and
3 were given 6-MSITC at a dose of 20 and 40 mg/kg body weight in
corn oil using a gastric tube, respectively (equivalent to daily
consumption of 6-MSITC at 200 or 400 ppm). The animals were
decapitated 24 h after the last dose of the vehicle or 6-MSITC.
Livers were perfused in situ with ice-cold sterile 1.15%
KCl, and 25% homogenates in 1.15% KCl were prepared. Microsomal
fractions from these tissues were prepared using established
procedures (22).
Western blot analysis
Gel electrophoresis and blot analysis were carried
out as described in detail previously, according to the established
methods of Laemmli (23) and Towbin
et al (24), respectively.
Separated proteins were transferred by semi-dry electroblotting
from sodium dodecylsulfate-polyacrylamide gels to polyvinylidene
difluoride membranes (Immobilon-P; Millipore, Bedford, MA, USA) in
25 mM Tris buffer (pH 8.3) containing 0.19 M glycine and 20% (v/v)
methanol. The membranes were blocked by incubation with
phosphate-buffered saline containing 5% skim milk for 1 h before
incubation with goat anti-rat polyclonal antibodies for CYP1A1/2,
CYP2B1/2, CYP2E1 and CYP3A2 (Daiichi Pure Chemicals, Tokyo, Japan),
and then stained using biotinylated anti-goat immunoglobulin G
(Vector Laboratories, Burlingame, CA, USA) using a Wako ABC-POD kit
(Wako Pure Chemicals, Osaka, Japan).
Assay of UDPGT activity
UDPGT activity towards bilirubin and 4-nitrophenol
in liver microsomes was assayed according to the methods of
Heirwegh et al (25) and
Isselbacher et al (26),
respectively, and towards testosterone with
UDP-[14C(U)]glucuronic acid, as described by Matern
et al (27). Each test was
carried out with liver microsomes pooled from three rats.
Experiments were performed six to eight times, and then the average
value was calculated.
Statistical analysis
Data are expressed as the mean ± SD. The statistical
significance of the difference in mean values was analyzed using
one-way analysis of variance (ANOVA) and the unpaired t-test.
Significance was defined as P<0.05.
Results
General observation
The rats tolerated s.c. injection of DMH and/or
6-MSITC feeding well. During the study, no clinical signs of
toxicity were present in any group. The mean body, liver and kidney
weights did not significantly differ among the groups (Table I). Histologically, there were no
pathological alterations indicative of toxicity of 6-MSITC in the
major organs, such as the liver or kidney.
 | Table IBody, liver and kidney weights. |
Table I
Body, liver and kidney weights.
| Group | Treatment (no. of
rats examined) | Body weight
(g) | Liver weight
(g) | Kidney weight
(g) |
|---|
| 1 | DMH + basal diet
(10) | 319.4±14.9 | 10.55±0.87 | 2.13±0.13 |
| 2 | DMH + 200 ppm
6-MSITC (10) | 316.8±20.4 | 11.04±1.13 | 2.20±0.19 |
| 3 | DMH + 400 ppm
6-MSITC (10) | 311.3±17.9 | 11.24±0.81 | 2.21±0.17 |
| 4 | DMH→200 ppm 6-MSITC
(10) | 309.4±17.8 | 10.99±1.45 | 2.13±0.13 |
| 5 | DMH→400 ppm 6-MSITC
(10) | 307.5±22.6 | 11.37±1.19 | 2.14±0.16 |
| 6 | 400 ppm 6-MSITC
(8) | 299.5±14.7 | 11.37±0.79 | 2.16±0.18 |
| 7 | Basal diet (8) | 315.7±24.4 | 11.94±1.30 | 2.26±0.11 |
Inhibitory effects of 6-MSITC on ACF and
BCAC
The rats in groups 1–5 developed ACF and BCAC in the
colonic mucosa. No such lesions were noted in any of the rats in
groups 6 and 7. In group 1, the frequency of ACF and BCAC were
323.8±69.7/colon and 3.80±1.05/cm2 colonic mucosa,
respectively. The number of ACF/colon was significantly lower in
group 3 DMH-treated rats fed a 400 ppm 6-MSITC diet during the
initiation phase than that in rats treated with DMH alone (group 1)
(153.9±62.4, 52% reduction, P<0.0001; Table II). The reduction was also
significant for the larger ACF with ≥4 crypts: from 155.1 to 65.4
(58% reduction, P<0.001; Table
II, group 3 vs. group 1). In group 3, 400 ppm 6-MSITC also
caused a significant decrease in the mean number of
BCAC/cm2 colon when compared with the control rats in
group 1 (0.91±0.58, 76% reduction, P<0.00001; Table III). Thus, both ACF and BCAC were
reduced to approximately the same extent. Regarding crypt
multiplicity, a significant decrease in the number of crypts/lesion
in group 3 was found in both ACF and BCAC when compared with that
in group 1 (P<0.05 and P<0.001, respectively). The number of
crypts/BCAC in groups 2 (DMH + 200 ppm 6-MSITC) and 5 (DMH→400 ppm
6-MSITC) were smaller than those in group 1, with a statistical
significance of P<0.05 and P<0.01, respectively.
| Table IIEffect of 6-MSITC on DMH-induced ACF
formation in male F344 rats. |
Table II
Effect of 6-MSITC on DMH-induced ACF
formation in male F344 rats.
| Group | Treatment | No. of
ACF/colon | No. of ACF ≥4
crypts | No. of
crypts/ACF |
|---|
| 1 | DMH + basal
diet | 323.8±69.7 | 155.1±45.4 | 3.67±0.23 |
| 2 | DMH + 200 ppm
6-MSITC | 351.6±32.8 | 162.1±18.7 | 3.67±0.17 |
| 3 | DMH + 400 ppm
6-MSITC | 153.9±62.4a | 65.4±36.6b | 3.38±0.24c |
| 4 | DMH→200 ppm
6-MSITC | 339.3±80.4 | 164.1±45.7 | 3.70±0.31 |
| 5 | DMH→400 ppm
6-MSITC | 310.0±55.6 | 140.3±18.6 | 3.61±0.21 |
| 6 | 400 ppm
6-MSITC | 0 | 0 | 0 |
| 7 | Basal diet | 0 | 0 | 0 |
| Table IIIEffect of 6-MSITC on DMH-induced BCAC
formation in male F344 rats. |
Table III
Effect of 6-MSITC on DMH-induced BCAC
formation in male F344 rats.
| Group | Treatment | No. of
BCAC/cm2 | No. of
crypts/BCAC |
|---|
| 1 | DMH + basal
diet | 3.80±1.05 | 8.75±2.04 |
| 2 | DMH + 200 ppm
6-MSITC | 3.47±1.34 | 6.87±1.30a |
| 3 | DMH + 400 ppm
6-MSITC | 0.91±0.58b | 5.01±1.48c |
| 4 | DMH→200 ppm
6-MSITC | 3.56±1.00 | 8.38±2.86 |
| 5 | DMH→400 ppm
6-MSITC | 3.20±1.40 | 6.43±1.34d |
| 6 | 400 ppm
6-MSITC | 0 | 0 |
| 7 | Basal diet | 0 | 0 |
Inhibition of the PCNA labeling index in
ACF and BCAC
In group 3, treatment of rats with 400 ppm levels of
6-MSITC resulted in a significant decrease in the labeling index of
PCNA in BCAC when compared to group 1 treated with DMH alone
(P<0.01; Table IV). The mean
PCNA labeling indices of groups 4 and 5 in ACF were lower than
group 1, but the differences did not reach statistical
significance.
| Table IVPCNA-labeling index in ACF and
BCAC. |
Table IV
PCNA-labeling index in ACF and
BCAC.
| Group | Treatment (no. of
rats examined) | ACF | BCAC |
|---|
| 1 | DMH + basal diet
(5) | 26.8±9.5 | 50.6±10.1 |
| 2 | DMH + 200 ppm
6-MSITC (5) | 21.4±15.3 | 39.9±10.9 |
| 3 | DMH + 400 ppm
6-MSITC (5) | 21.2±19.1 | 28.4±8.4a |
| 4 | DMH→200 ppm 6-MSITC
(5) | 11.8±8.6 | 38.1±5.1 |
| 5 | DMH→400 ppm 6-MSITC
(5) | 11.6±9.6 | 36.2±12.1 |
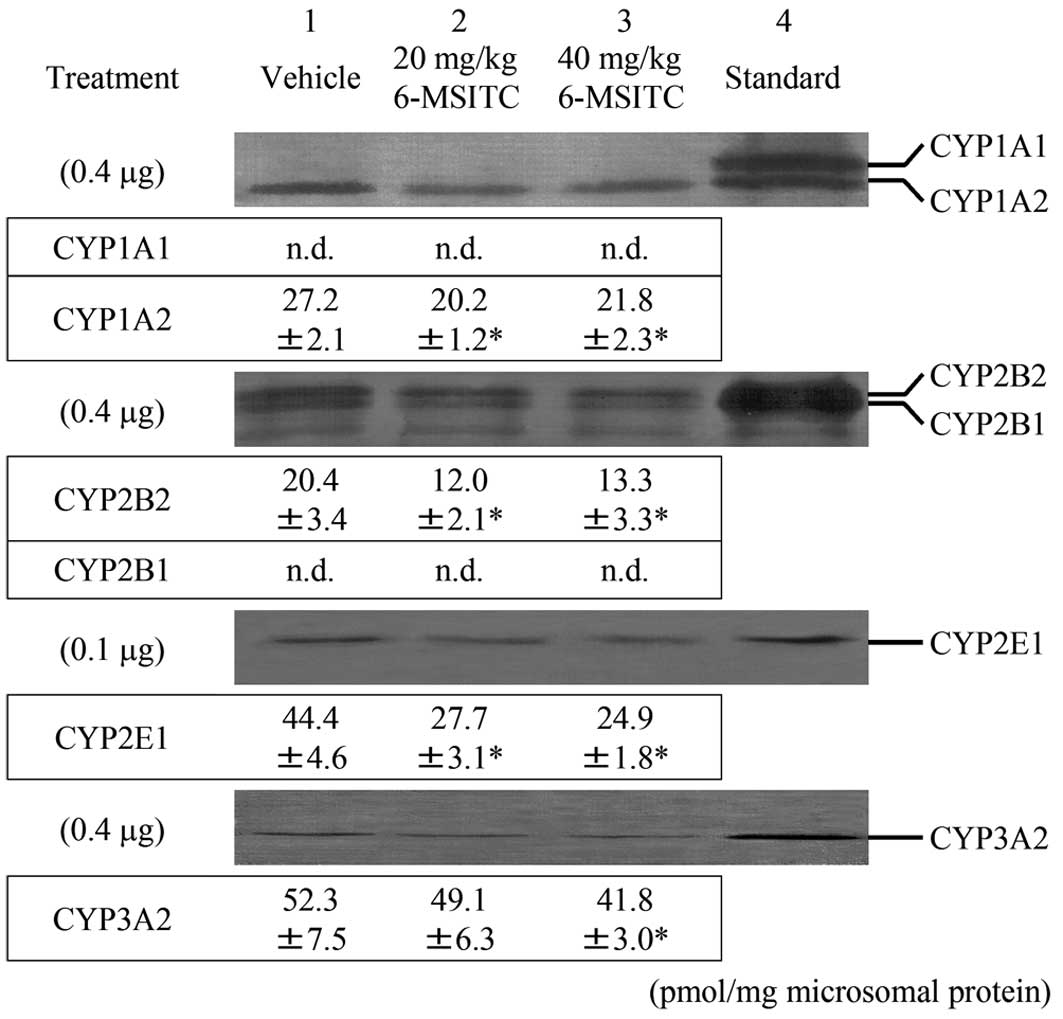
Determination of CYP isoforms in the
liver
Fig. 1 shows
immunoblots and levels of liver microsomal CYP proteins in male
F344 rats 24 h after 6-MSITC treatment. Neither hepatic CYP2B1 nor
CYP1A1 were constitutively expressed. Treatment with 40 mg/kg b.w.
6-MSITC significantly decreased CYP1A2, -2B2, -2E1 and -3A2
proteins by 20, 35, 44 and 20%, respectively, relative to the
vehicle group. Exposure to 20 mg/kg b.w. 6-MSITC also significantly
decreased hepatic CYP1A2, -2B2 and -2E1 protein levels when
compared to group 1.
Hepatic UDPGT activity
Table V summarizes
the effects of 6-MSITC treatment on hepatic UDPGT activity towards
4-nitrophenol, bilirubin and testosterone in liver microsomes. No
significant differences were noted among the three groups.
| Table VUDPGT activities in liver microsomes
from male F344 rats. |
Table V
UDPGT activities in liver microsomes
from male F344 rats.
| | UDPGT activity
(nmol/min/mg protein) |
|---|
| |
|
|---|
| Group | Treatment | 4-Nitrophenol | Bilirubin | Testosterone |
|---|
| 1 | Vehicle | 31.2±5.2 | 0.18±0.01 | 0.12±0.04 |
| 2 | 20 mg/kg
6-MSITC | 35.8±4.7 | 0.17±0.02 | 0.12±0.02 |
| 3 | 40 mg/kg
6-MSITC | 31.4±2.7 | 0.17±0.01 | 0.13±0.05 |
Discussion
In this study, the modulating effects of 6-MSITC on
colonic ACF and BCAC formation were proven by their exposure at the
initiation and promotion phases in rats. ACF have been widely used
as intermediate biomarkers of colon carcinogenesis in experimental
animal models, and a positive correlation has been described
between the effects of chemopreventive agents on ACF and tumor
development (28). It is likely
that larger ACF (≥4 crypts/lesion) are histogenetically close to
dysplastic adenomas in order for adenomas to be more appropriate
biomarkers (29,30). BCAC, another mucosal lesion, was
evaluated in the present bioassay as a surrogate biomarker of colon
carcinogenesis. Our previous results indicated that BCAC are more
sensitive and more reliable than ACF as intermediate biomarkers of
colon carcinogenesis (31,32). Significantly, ACF and BCAC are
considered to be independent and distinct, as they differ in
biology, genetics and morphology (31,33,34).
In this context, the present result that 6-MSITC inhibited the
formation of two different lesions can be regarded as convincing.
Furthermore, rats fed the diets containing 6-MSITC showed no
adverse effects and no clinical signs of toxicity. Collectively,
these findings suggest that 6-MSITC is a new chemopreventive agent
against colon cancer development.
Several explanations for the inhibitory effects of
6-MSITC on ACF and BCAC formation by DMH were considered. Cell
proliferation has long been suspected to play a significant role in
the initiation step as well as the promotion of carcinogenesis
(35). In the current study, we
found that 6-MSITC inhibited proliferation of epithelial cells in
ACF and BCAC. A previous study showed that 6-MSITC inhibited the
growth of MCF-7 human breast cancer and LOX-IMVI human melanoma
cell lines (17). Thus, the
inhibitory effect of 6-MSITC may be, in part, due to the
modification of cell proliferation activity in cryptal cells.
Data on the incidence and crypt multiplicity of
colonic preneoplastic lesions indicated that a 6-MSITC feeding
together with or after DMH exposure was able to inhibit colonic
tumorigenesis, the suppression of dietary 6-MSITC during the
initiation phase being more effective. This is in accordance with
the previous findings for a 6-MSITC inhibition of chemically
induced carcinogenesis (36), and
the theory that the compound should be viewed as a ‘blocking’ agent
against colon carcinogenesis (37).
Regarding mechanisms, ‘blocking’ compounds generally have a
capacity to induce phase II detoxification enzymes and/or inhibit
phase I enzymes that are required for the bioactivation of
carcinogens (38). It has also been
reported that ITCs function by blocking the initiation phase in
part via inhibition of phase I enzymes and/or induction of phase II
enzymes (10–13). This may be the case with 6-MSITC,
since it decreased hepatic CYP1A2, -2B2, -2E1 and -3A2 by 7–44%,
compared with the vehicle control. It is generally accepted that
CYPs are known to play a prominent role in the biotransformation of
carcinogenic xenobiotics. In the rat, hepatic CYP1A1/2, CYP2B1/2
and CYP3A2 were reported to contribute to the mutagenic activation
of heterocyclic amines, benzo[a]pyrene and aflatoxins
(39,40). CYP2B1/2 and -2E1 are also
specifically involved in the metabolic activation of environmental
N-nitrosamines to ultimate carcinogens (41,42).
Furthermore, CYP2E1 is one of the enzymes catalyzing the conversion
of azoxymethane and methylazoxymethanol, which are metabolites of
DMH, to DNA alkylating species capable of causing initiation events
(43,44). In this context, decreased levels of
CYPs by 6-MSITC were expected to result in the reduction and/or
slowing of the transformation of metabolites of DMH to proximate or
ultimate carcinogens. On the other hand, 6-MSITC did not
significantly alter the UDPGT activity towards bilirubin,
4-nitrophenol and testosterone in hepatic microsomes.
Glucuronidation is generally considered to be one of the
detoxification reactions in the metabolism of various chemicals
including carcinogens. UDPGT1A1 (for bilirubin), UDPGT1A6 (for
4-nitrophenol) and UDPGT2B1 (for testosterone) are found as major
enzymes in the liver, and are responsible for the glucuronidation
reaction. In a previous study, we showed that certain carcinogenic
compounds are substrates for the enzymes (45). However, the present findings showed
that hepatic UDPGT activity was not involved in 6-MSITC
modification.
In conclusion, the results described herein
demonstrate that the dietary administration of 6-MSITC can
significantly inhibit the induction of colonic ACF and BCAC by DMH,
by reducing cell proliferative activity and the protein levels of
phase I enzymes.
Acknowledgements
We appreciate the technical assistance of Ms. K.
Takahashi, the secretarial assistance of Ms. S. Gotou and the
animal care given by Mr. Y. Kinjou. This work was supported by
Grants-in-Aid from the Ministry of Education, Culture, Sports,
Science and Technology and the Ministry of Health, Labor and
Welfare of Japan.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
2
|
Willett W: The search for the causes of
breast and colon cancer. Nature. 338:389–394. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hebert JR, Landon J and Miller DR:
Consumption of meat and fruit in relation to oral and esophageal
cancer: a cross-national study. Nutr Cancer. 19:169–179. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Block G, Patterson B and Subar A: Fruit,
vegetables and cancer prevention: a review of the epidemiological
evidence. Nutr Cancer. 18:1–29. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boyle P, Zaridze DG and Smans M:
Descriptive epidemiology of colorectal cancer. Int J Cancer.
36:9–18. 1985. View Article : Google Scholar
|
|
6
|
Lin HJ, Probst-Hensch NM, Louie AD, et al:
Glutathione transferase null genotype, broccoli and lower
prevalence of colorectal adenomas. Cancer Epidemiol Biomarkers
Prev. 7:647–652. 1998.
|
|
7
|
Hecht SS: Chemoprevention by
isothiocyanates. J Cell Biochem Suppl. 22:195–209. 1995. View Article : Google Scholar
|
|
8
|
Munday R, Mhawech-Fauceglia P, Munday CM,
et al: Inhibition of urinary bladder carcinogenesis by broccoli
sprouts. Cancer Res. 68:1593–1600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Higdon JV, Delage B, Williams DE and
Dashwood RH: Cruciferous vegetables and human cancer risk:
epidemiologic evidence and mechanistic basis. Pharmacol Res.
55:224–236. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Leonard TB, Popp JA, Graichen ME and Dent
JG: alpha-Naphthylisothiocyanate induced alterations in hepatic
drug metabolizing enzymes and liver morphology: implications
concerning anticarcinogenesis. Carcinogenesis. 2:473–482. 1981.
View Article : Google Scholar
|
|
11
|
Guo Z, Smith TJ, Wang E, Eklind KI, Chung
FL and Yang CS: Structure-activity relationships of arylalkyl
isothiocyanates for the inhibition of
4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone metabolism and the
modulation of xenobiotic-metabolizing enzymes in rats and mice.
Carcinogenesis. 14:1167–1173. 1993. View Article : Google Scholar
|
|
12
|
Zhang Y and Talalay P: Anticarcinogenic
activities of organic isothiocyanates: chemistry and mechanisms.
Cancer Res. 54:S1976–S1981. 1994.PubMed/NCBI
|
|
13
|
Guo Z, Smith TJ, Wang E, et al: Effects of
phenethyl isothiocyanate, a carcinogenesis inhibitor, on
xenobiotic-metabolizing enzymes and nitrosamine metabolism in rats.
Carcinogenesis. 13:2205–2210. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kinae N, Masuda H, Shin IS, Furugori M and
Shimoi K: Functional properties of wasabi and horseradish.
Biofactors. 13:265–269. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Morimitsu Y, Hayashi K, Nakagawa Y, Horio
F, Uchida K and Osawa T: Antiplatelet and anticancer
isothiocyanates in Japanese domestic horseradish, wasabi.
Biofactors. 13:271–276. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Watanabe M, Ohata M, Hayakawa S, et al:
Identification of 6-methylsulfinylhexyl isothiocyanate as an
apoptosis-inducing component in wasabi. Phytochemistry. 62:733–739.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nomura T, Shinoda S, Yamori T, et al:
Selective sensitivity to wasabi-derived 6-(methylsulfinyl)hexyl
isothiocyanate of human breast cancer and melanoma cell lines
studied in vitro. Cancer Detect Prev. 29:155–160. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Morimitsu Y, Nakagawa Y, Hayashi K, et al:
A sulforaphane analogue that potently activates the Nrf2-dependent
detoxification pathway. J Biol Chem. 277:3456–3463. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thimmulappa RK, Mai KH, Srisuma S, Kensler
TW, Yamamoto M and Biswal S: Identification of Nrf2-regulated genes
induced by the chemopreventive agent sulforaphane by
oligonucleotide microarray. Cancer Res. 62:5196–5203.
2002.PubMed/NCBI
|
|
20
|
Bird RP: Observation and quantification of
aberrant crypts in the murine colon treated with a colon
carcinogen: preliminary findings. Cancer Lett. 37:147–151. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamada Y, Yoshimi N, Hirose Y, et al:
Suppression of occurrence and advancement of
beta-catenin-accumulated crypts, possible premalignant lesions of
colon cancer, by selective cyclooxygenase-2 inhibitor, celecoxib.
Jpn J Cancer Res. 92:617–623. 2001. View Article : Google Scholar
|
|
22
|
Mori Y, Yamazaki H, Toyoshi K, et al:
Mutagenic activation of carcinogenic N-nitrosopropylamines by rat
liver: evidence for a cytochrome P-450-dependent reaction.
Carcinogenesis. 6:415–420. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Towbin H, Staehelin T and Gordon J:
Electrophoretic transfer of proteins from polyacrylamide gels to
nitrocellulose sheets: procedure and some applications. Proc Natl
Acad Sci USA. 76:4350–4354. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Heirwegh KP, van de Vijver M and Fevery J:
Assay and properties of dititonin-activated bilirubin uridine
diphosphate glucuronyltransferase from rat liver. Biochem J.
129:605–618. 1972.PubMed/NCBI
|
|
26
|
Isselbacher KJ, Chrabas MF and Quinn RC:
The solubilization and partial purification of a glucuronyl
transferase from rabbit liver microsomes. J Biol Chem.
237:3033–3036. 1962.PubMed/NCBI
|
|
27
|
Matern H, Heinemann H and Matern S:
Radioassay of UDP-glucuronosyltransferase activities toward
endogenous substrates using labeled UDP-glucuronic acid and an
organic solvent extraction procedure. Anal Biochem. 219:182–188.
1994. View Article : Google Scholar
|
|
28
|
Corpet DE and Tache S: Most effective
colon cancer chemopreventive agents in rats: a systematic review of
aberrant crypt foci and tumor data, ranked by potency. Nutr Cancer.
43:1–21. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Corpet DE, Stamp D, Medline A, Minkin S,
Archer MC and Bruce WR: Promotion of colonic microadenoma growth in
mice and rats fed cooked sugar or cooked casein and fat. Cancer
Res. 50:6955–6958. 1990.PubMed/NCBI
|
|
30
|
Pretlow TP, O’Riordan MA, Somich GA, Amini
SB and Pretlow TG: Aberrant crypts correlate with tumor incidence
in F344 rats treated with azoxymethane and phytate. Carcinogenesis.
13:1509–1512. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yamada Y, Yoshimi N, Hirose Y, et al:
Sequential analysis of morphological and biological properties of
beta-catenin-accumulated crypts, provable premalignant lesions
independent of aberrant crypt foci in rat colon carcinogenesis.
Cancer Res. 61:1874–1878. 2001.
|
|
32
|
Hirose Y, Kuno T, Yamada Y, et al:
Azoxymethane-induced beta-catenin-accumulated crypts in colonic
mucosa of rodents as an intermediate biomarker for colon
carcinogenesis. Carcinogenesis. 24:107–111. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yamada Y, Yoshimi N, Hirose Y, et al:
Frequent beta-catenin gene mutations and accumulations of the
protein in the putative preneoplastic lesions lacking macroscopic
aberrant crypt foci appearance, in rat colon carcinogenesis. Cancer
Res. 60:3323–3327. 2000.
|
|
34
|
Yamada Y, Oyama T, Hirose Y, et al:
beta-Catenin mutation is selected during malignant transformation
in colon carcinogenesis. Carcinogenesis. 24:91–97. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cohen SM and Ellwein LB: Cell
proliferation in carcinogenesis. Science. 249:1007–1011. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yano T, Yajima S, Virgona N, et al: The
effect of 6-methylthiohexyl isothiocyanate isolated from Wasabia
japonica (wasabi) on
4-(methylnitrosamino)-1-(3-pyridyl)-1-buatnone-induced lung
tumorigenesis in mice. Cancer Lett. 155:115–120. 2000.PubMed/NCBI
|
|
37
|
Surh YJ: Cancer chemoprevention with
dietary phytochemicals. Nat Rev Cancer. 3:768–780. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Myzak MC and Dashwood RH: Chemoprotection
by sulforaphane: keep one eye beyond Keap1. Cancer Lett.
233:208–218. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mori Y, Koide A, Fuwa K and Kobayashi Y:
N-benzylimidazole for preparation of S9 fraction with
multi-induction of metabolizing enzymes in short-term genotoxicity
assays. Mutagenesis. 16:479–486. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Degawa M, Ueno H, Miura S, Ohta A and
Namiki M: A simple method for assessment of rat cytochrome P-448
isozymes responsible for the mutagenic activation of carcinogenic
chemicals. Mutat Res. 203:333–338. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mori Y, Tatematsu K, Koide A, Sugie S,
Tanaka T and Mori H: Modification by curcumin of mutagenic
activation of carcinogenic N-nitrosamines by extrahepatic
cytochromes P-450 2B1 and 2E1 in rats. Cancer Sci. 97:896–904.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mori Y, Koide A, Kobayashi Y, Morimura K,
Kaneko M and Fukushima S: Effect of ethanol treatment on metabolic
activation and detoxification of esophagus carcinogenic
N-nitrosamines in rat liver. Mutagenesis. 17:251–256. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sohn OS, Ishizaki H, Yang CS and Fiala ES:
Metabolism of azoxymethane, methylazoxymethanol and
N-nitrosodimethylamine by cytochrome P450IIE1. Carcinogenesis.
12:127–131. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sohn OS, Fiala ES, Requeijo SP, Weisburger
JH and Gonzalez FJ: Differential effects of CYP2E1 status on the
metabolic activation of the colon carcinogens azoxymethane and
methylazoxymethanol. Cancer Res. 61:8435–8440. 2001.PubMed/NCBI
|
|
45
|
Mori Y, Takahashi H, Yamazaki H, et al:
Distribution, metabolism and excretion of
N-nitrosobis(2-hydroxypropyl)amine in Wistar rats. Carcinogenesis.
5:1443–1447. 1984. View Article : Google Scholar : PubMed/NCBI
|