Introduction
Polyamines are essential for eukaryotic cell
survival although an increase of polyamines has been shown to be
associated with survival of cancers. This indicates that the
depletion of polyamines may be a potential strategy for cancer
treatment. It has long been known that the depletion of polyamines
can induce apoptosis in several tumor types using either a
polyamine synthesis inhibitor or a polyamine analog (1,2).
However, polyamine synthesis inhibitors have only demonstrated a
moderate inhibitory effect on glioblastomas (3).
N1,N11-diethylnorspermine
(DENSPM), a well-studied polyamine analog, was shown in our
previous study to induce significant apoptosis in several
glioblastoma cell lines (3).
Although the mechanism of polyamine analog-induced cell death is
still not well understood, it has been shown that DENSPM induces
spermidine/spermine acetyltransferase (SSAT) expression, which
converts spermine and spermidine to their acetylated forms.
Acetylated spermine and spermidine can then serve as substrates for
spermine oxidase (SMO) or polyamine oxidase (PAO) in reactions that
produce H2O2 (4,5). In
most experiments, DENSPM was described to induce apoptosis in
cancer cells by damaging mitochondria, releasing cytochrome c into
cytoplasm and activating the caspase-3 and -8 signal transduction
pathways (6,7). We have also observed mTOR relocation
in U87 cell lines treated by DENSPM (8).
Unexpectedly, significant cell detachment was
observed in some glioblastoma cells but not in others. Consistent
with our results, other groups have also reported that marked cell
detachment could be induced either in CHENSPM
(N1-ethyl-N11-[(cyclopropyl)methyl]-4,8,-diazaundecane, another
polyamine analog) treated lung cancer cells (9) or in DENSPM-treated breast cancer cells
(10). Furthermore, the correlation
between increased SSAT levels and cell floating in kidney carcinoma
cells (HEK293) was also associated with morphological change in
cell shape (11). Based on these
findings we hypothesized that elevated SSAT expression plays a key
role in DENSPM-induced cell detachment in glioblastoma cells.
Materials and methods
Cells and cell culture
The human glioblastoma cell lines LN229 and U87 were
purchased from American Type Culture Collection (Manassas, VA) and
maintained in Dulbecco’s modified Eagle’s medium (DMEM)/F12 (Gibco,
USA) supplemented with 10% dialyzed serum (Hyclone, Logan, UT) at
37°C in a 5% CO2 incubator. Because fetal bovine serum
has abundant thymine and polyamines, we used the dialyzed serum to
avoid interfering with the experimental results.
Reagents
DENSPM was purchased from Tocris (Ellisville, MO)
and was dissolved in water according to the manufacturer’s
instructions. The PCMV-SSAT and PCMV empty plasmids (negative
control) were kindly provided by Professor Eugene Gerner (Arizona
Cancer Center, University of Arizona, Tucson, AZ) and were
sequenced in our laboratory before using in order to confirm their
quality and their human source.
In vitro drug treatment experiments
For the in vitro drug treatment experiments,
the cells were seeded in a 10-cm2 dish (105
cells/dish) in 10 ml of medium supplemented with 10% dialyzed fetal
bovine serum. Twenty-four hours later, 10 μM DENSPM was added.
PCMV-SSAT transfection
After LN229 cells reached 80% confluency in the
culture plates, they were collected by trypsinization and counted.
The PCMV-SSAT and negative control PCMV empty plasmids were
transfected into 1×106 LN229 cells in parallel with
Nucleofector technology according to the manufacturer’s protocol
(Amaxa Biosystems, Gaithersburg, MD). Before transfection, a GFP
plasmid was added into the target plasmids to serve as the
illumination marker to confirm successful transfection. The
transfected LN229 cells were distributed into 10-cm2
dishes and continuously cultured in the humidified incubator
containing 5% CO2 at 37°C for another 24 h.
Knockdown of SSAT expression by
siRNA
Dharmacon SMARTpool® siRNAs (Dharmacon,
Lafayette, CO) were used for silencing SSAT with Nucleofector
technology according to the manufacturer’s protocol. For the
non-specific target, nonsense siRNA (Ambion Inc., Austin, TX) was
used as a control. Briefly, 2–3×106 LN229 or U87 cells
were resuspended in 100 μl of Nucleofector solution with 100 nM of
siRNA in the electroporation cuvette. After electroporation, cells
were divided into 12-well plates and incubated in the transfection
reagent with siRNA at 37°C in a humidified incubator with 5%
CO2 for 24 h. Following the transfection procedure 10 μM
DENSPM was added into the plates.
Real-time quantitative PCR analysis
The total RNA was extracted using TRIzol
(Invitrogen, USA) according to the manufacturer’s protocol. The
mRNA level of SSAT from the PCMV-SSAT or PCMV empty plasmid
transfected LN229 cells, SSAT siRNA- or nonsense siRNA-transfected
U87 cells, DENSPM-treated and untreated U87 and LN229 cells were
quantified using the Applied Biosystems TaqMan method in
conjunction with Assays-On-Demand (ABI Prism 7900 sequence
detection system, Applied Biosystems, Foster City, CA) based on the
previous description (6). The
results of real-time PCR were analyzed by the ΔΔCT method: ΔCT =
CTselected gene - CTGAPDH, ΔΔCT =
ΔCTtherapy group - ΔCTcontrol group, RV
(relative value)therapy group = 2−ΔΔCT,
RVcontrol group = 1. The results of real-time PCR were
presented as the ratio between the selected genes and GAPDH
transcripts. The mean value of SSAT was calculated based on
triplicate experiments.
Cell detachment examination
To evaluate the detachment status of cells treated
with 10 μM DENSPM or after PCMV-SSAT transfection or knockdown of
SSAT, floating cells in the medium were collected first and then
the adherent cells were collected by trypsinization. The percentage
of detached cells was calculated by dividing the amount of the
total floating cells and the trypsinized adherent cells by the
number of the floating cells in the medium. The mean percentage of
the detached cells was calculated based on triplicate
experiments.
Cell viability assay
Cell viability was evaluated using the MTS assay
(Promega Corporation, Madison, WI). For the MTS assay, we seeded
3,000 LN229 cells transfected with PCMV-SSAT or transfected SSAT
siRNA per well in 100 μl of medium in a 96-well plate. On the
second day, varying concentrations of DENSPM were added to the
wells. After 20 μl of MTS solution had been added to each well and
mixed, the cells were incubated at 37°C in the 5% CO2
incubator. Absorbance at 490 nm was measured with a microplate
reader (MRX, Danatech Laboratory, Houston, TX). All MTS assays were
performed in triplicate for each treatment condition and
experiments were repeated at least twice to confirm the consistency
of results.
Western blotting
LN229 cells transfected with PCMV-SSAT were lysed
and homogenates were clarified by centrifugation at 12,000 × g for
15 min at 4°C. Supernatant samples were electrophoresed on 7.5%
sodium dodecyl sulfate (SDS)-polyacrylamide gels followed by
transfer to polyvinylidene difluoride (PVDF) membranes (Millipore,
USA). Incubation with primary polyclonal antibodies against
integrin α5 (BD Biosciences, San Jose, CA), integrin β1 (BD
Biosciences), AKT, mTOR (Cell Signaling Technology, Danvers, MA),
SSAT (Santa Cruz Biotechnology, Santa Cruz, CA) and actin (Sigma,
St. Louis, MO) were performed at a dilution of 1:1,000 overnight at
4°C. After washing, the membranes were incubated with secondary
antibodies conjugated to biotin (Amersham Pharmacia Biotech,
Piscataway, NJ) at a dilution of 1:3,000 for 30 min at room
temperature. Reactions were developed with ECL or ECL plus (GE
Healthcare, Buckinghamshire, UK). All Western blotting assays were
performed in triplicate for each probed protein.
Statistical analysis
The SPSS16.0 software (SPSS, Chicago, IL) was used
for statistical analysis. Differences in means were analyzed by the
two-tailed t-test, assuming unequal variances. All data are
expressed as mean ± SD. A P-value <0.05 was considered
significant.
Results
Cell detachment in DENSPM-treated
glioblastoma cells is dependent on SSAT expression levels
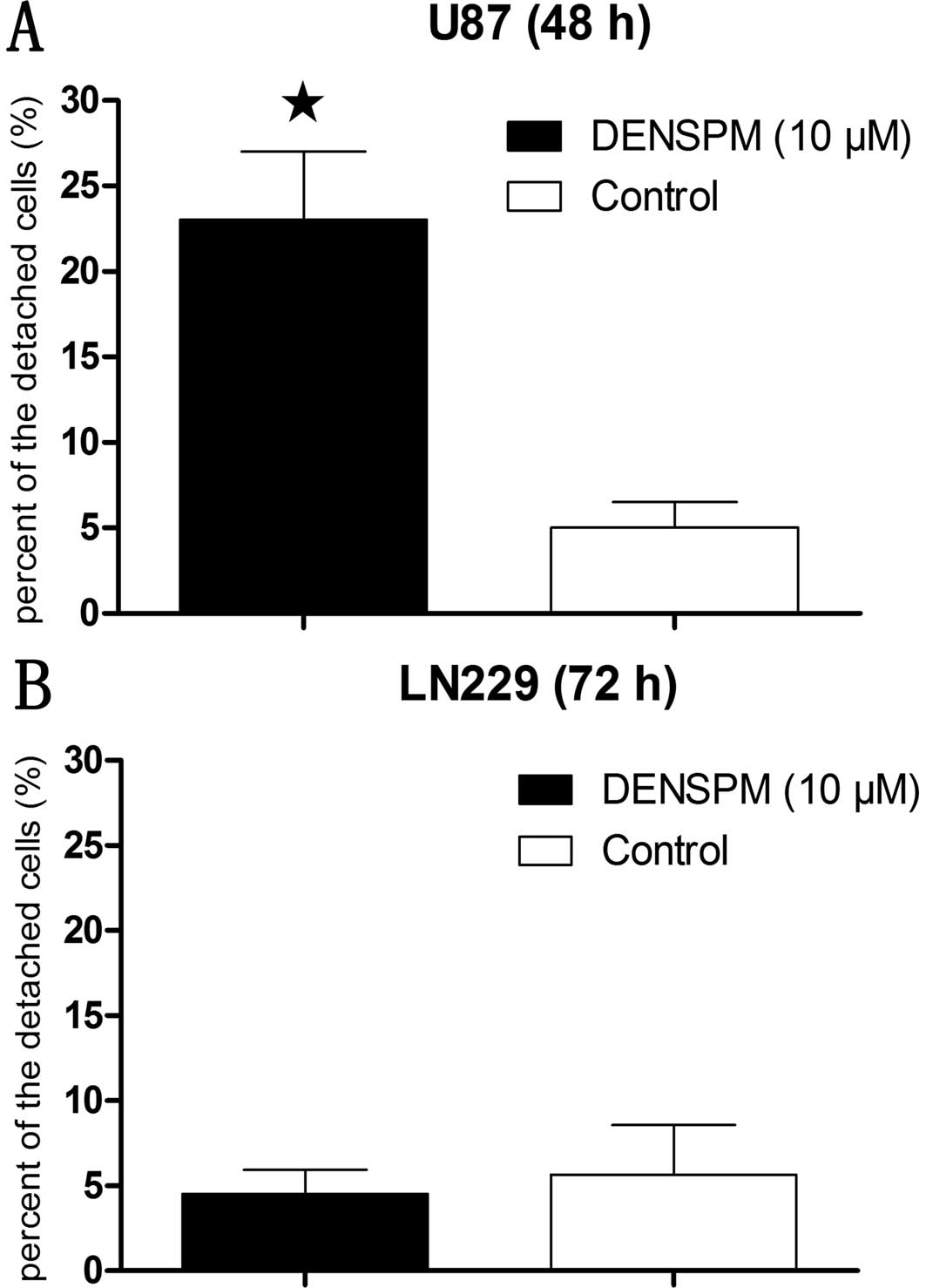
After incubation with DENSPM (10 μM) for 48 h, the
percentage of floating cells in U87 cells was ~25% (Fig. 1A). While the incubation time of the
same concentration of DENSPM in LN229 cells was extended up to 72
h, this did not result in any significant cell detachment relative
to control (Fig. 1B). The
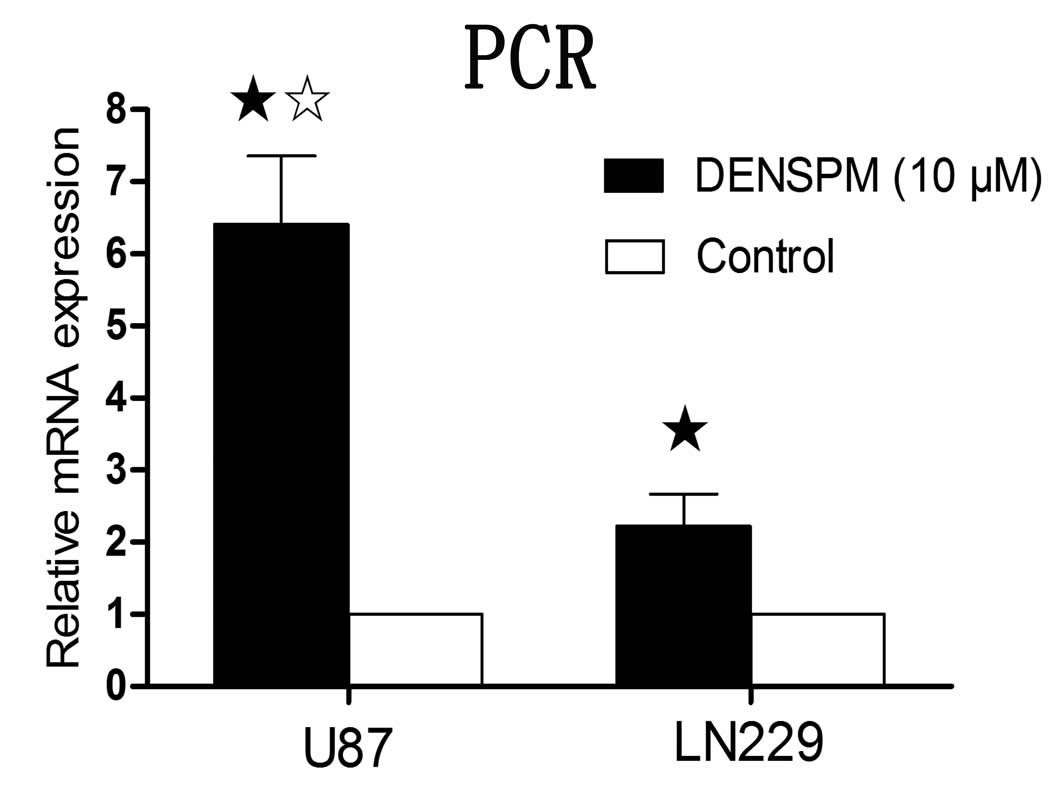
measurement of SSAT mRNA level with the real-time quantitative PCR
revealed that 10 μM of DENSPM induced significantly increased
expression of SSAT in both U87 and LN229 cells. The induction of
SSAT mRNA level was significantly higher in U87 cells than in LN229
when incubated for the same period of 24 h (Fig. 2).
Overexpression of SSAT in LN229 cells
induced by transfection of the PCMV-SSAT results in increased cell
detachment
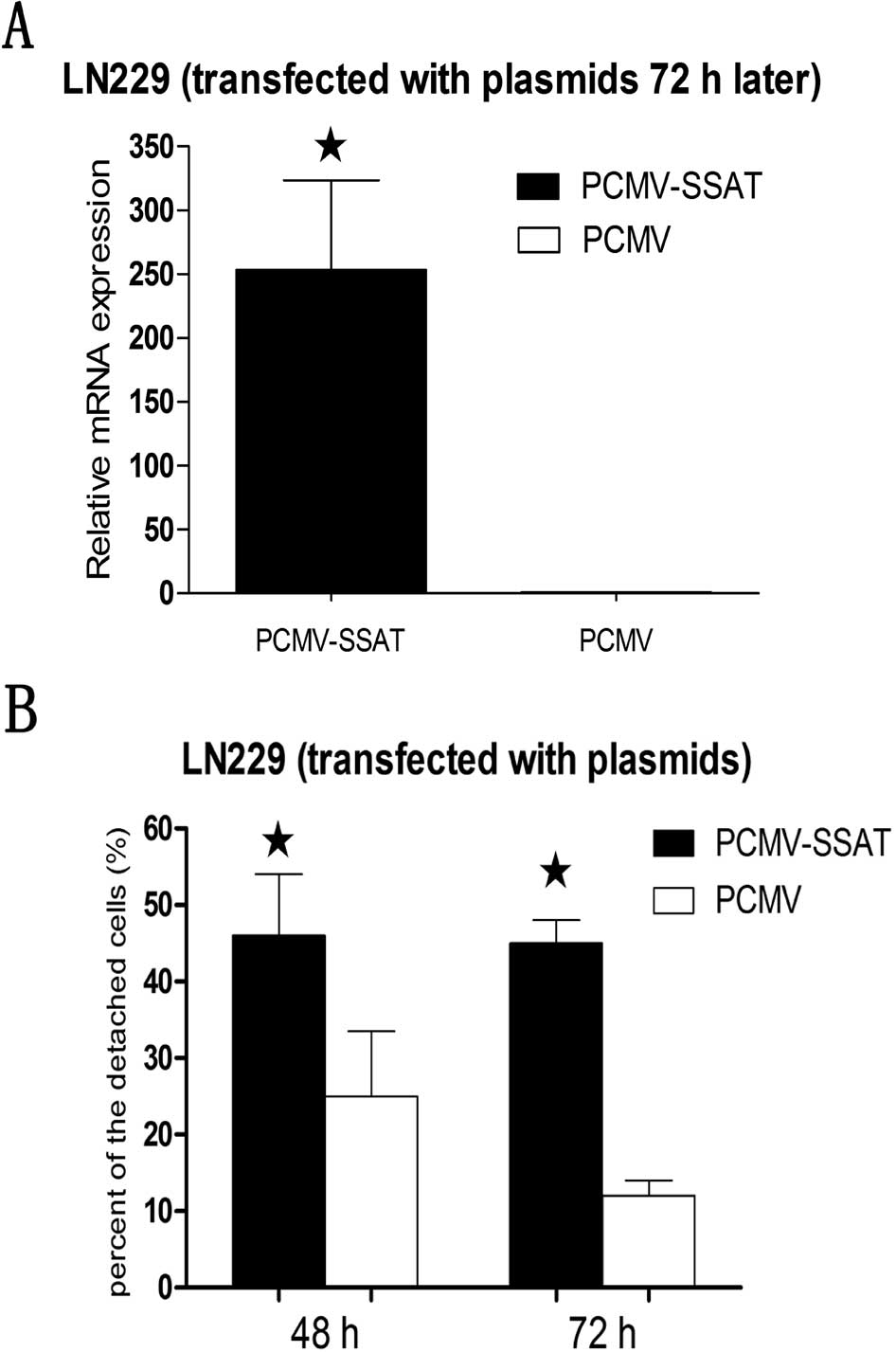
The plasmids of PCMV-SSAT were successfully
transfected into LN229 cells. Real-time quantitative PCR was
performed to confirm that SSAT mRNA was increased in the
PCMV-SSAT-LN229 cells after 72 h (Fig.
3A). Then the percentage of the floating cells in the
PCMV-SSAT-LN229 cells increased significantly after both 48 h and
72 h compared with cells transfected by PCMV empty plasmids. There
was no significant difference in SSAT levels between 48 h and 72 h
in transfected cells (Fig. 3B).
SSAT knockdown inhibits cell detachment
in DENSPM-treated U87 cells
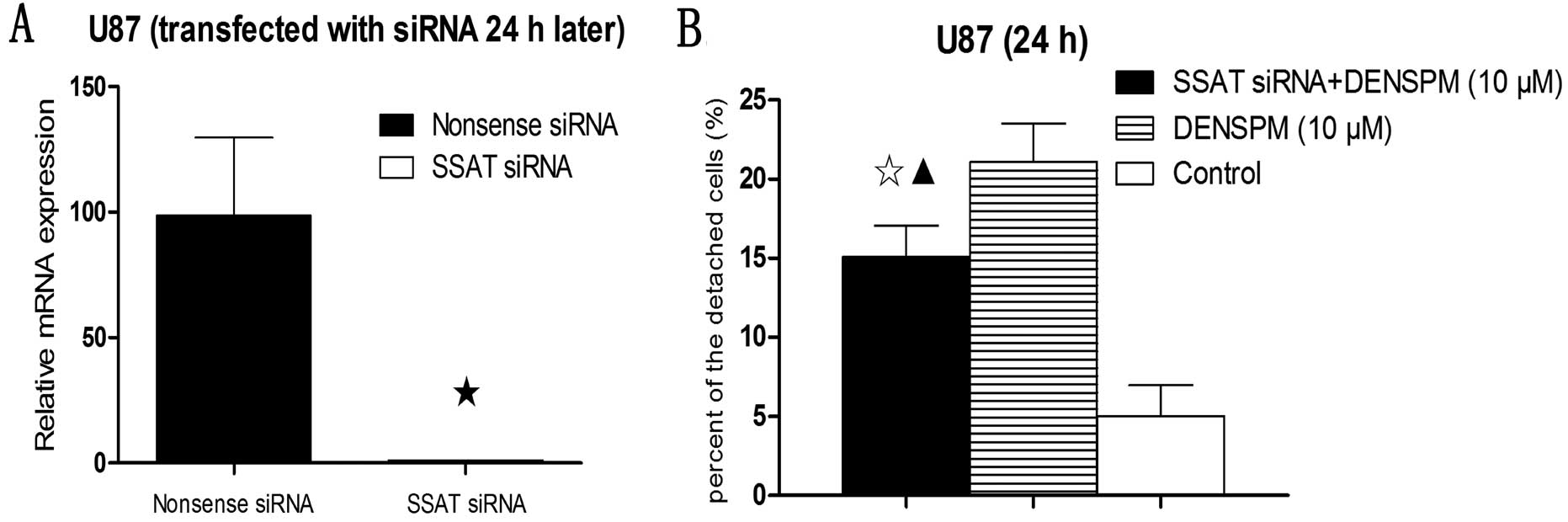
Knockdown of SSAT expression with siRNA was
confirmed with real-time quantitative PCR (Fig. 4A). Detached cells were counted in
DENSPM-treated and untreated U87 cells. A significant reduction in
cell detachment was observed in SSAT siRNA-transfected U87 cells
treated with DENSPM compared with the DENSPM alone treatment but
was still significantly higher than in the control group (Fig. 4B).
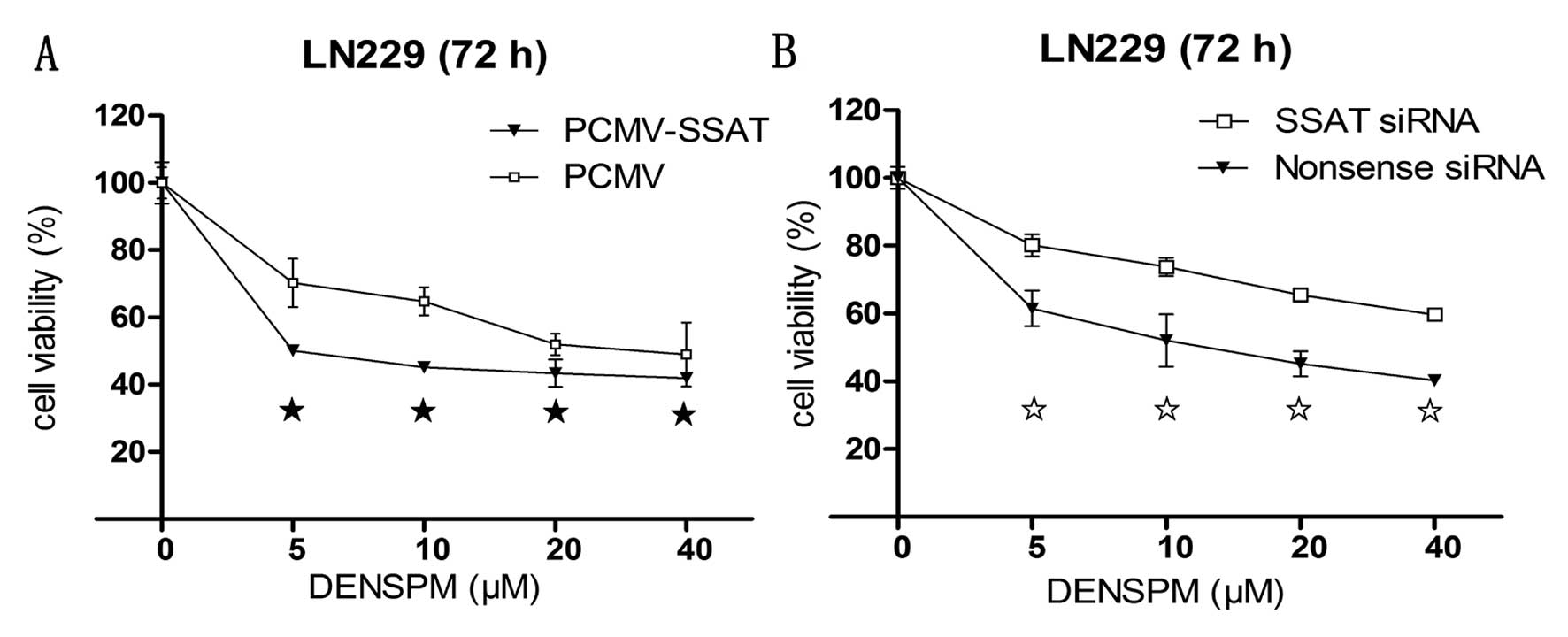
Levels of SSAT affect DENSPM-mediated
cytotoxicity in LN229 cells
Following successful SSAT upregulation by PCMV-SSAT
transfection or knockdown by SSAT siRNA, an MTS assay was performed
to determine the cytotoxicity of DENSPM in the LN229 cells with
serial concentrations of DENSPM. An elevation of SSAT resulted in
enhanced cell killing in DENSPM-treated PCMV-SSAT-LN229 cells
(Fig. 5A), while a mild attenuation
of the DENSPM killing result was observed after the SSAT was turned
down (Fig. 5B).
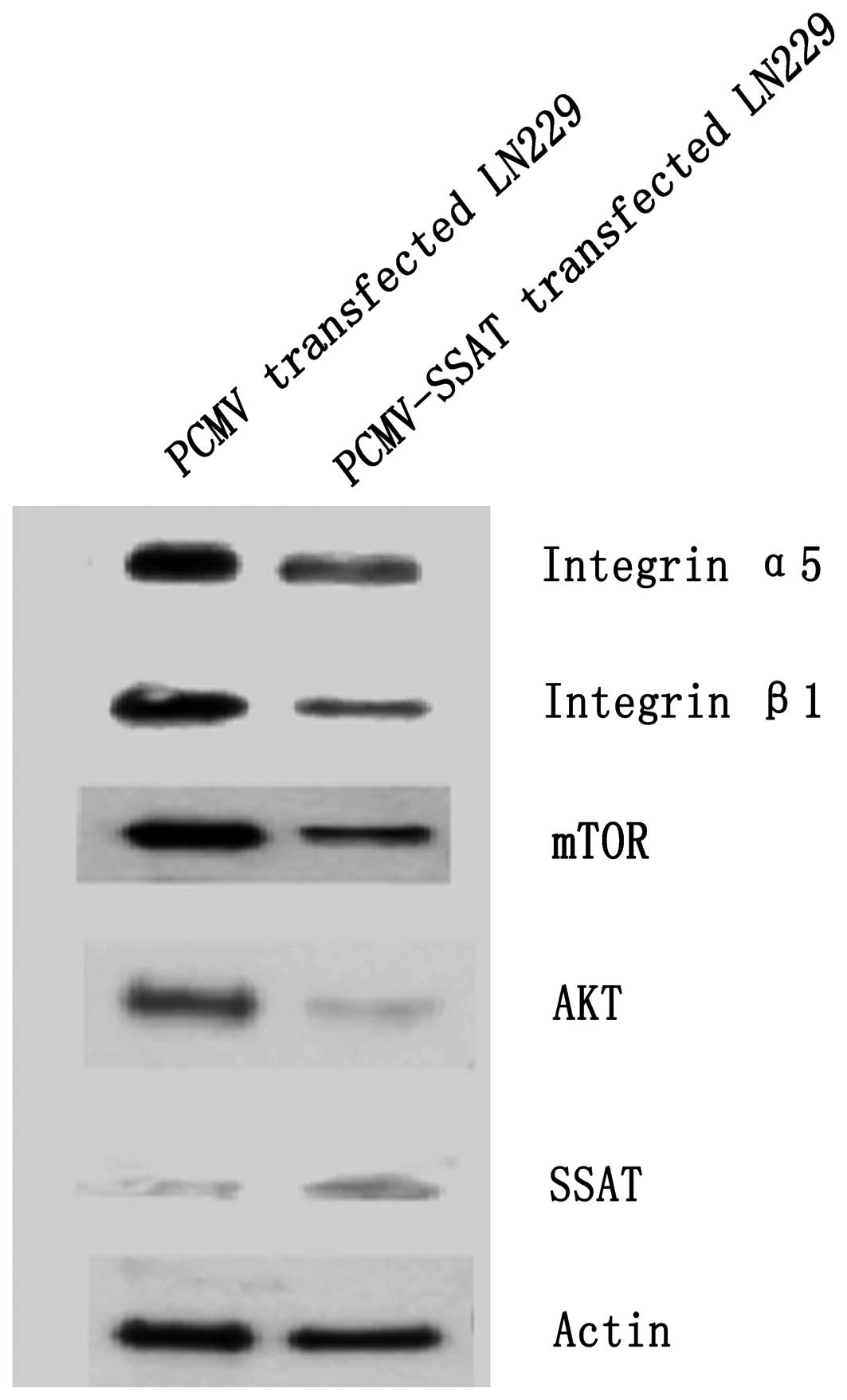
SSAT transfection induces degradation of
anti-apoptosis and adhesion-related proteins
PCMV-SSAT was transfected in LN229 cells and
confirmed by the real-time quantitative PCR. Expression of SSAT
protein was found to be increased by 256.52% after transfection.
The cell lysates were extracted after 48 h and Western blotting run
for expression of anti-apoptosis-related proteins AKT, mTOR and the
adhesion related proteins integrin α5, integrin β1. SSAT
transfection reduced AKT, mTOR, integrin α5 and integrin β1
expression by 35.90, 77.06, 79.91 and 47.33%, respectively
(Fig. 6).
Discussion
Polyamines are essential requirements for eukaryotic
cell growth. The metabolism of polyamines are frequently
dysregulated in cancer, and the polyamine pathway is a main target
for inhibiting the proliferation of carcinoma (2). Polyamine analogues resulting in
polyamine depletion in cells are highly attractive chemotherapy
targets for cancer. Polyamine analogues can enter into cells,
compete with the natural polyamines for uptake but do not
substitute for the natural polyamines in growth-related functions.
This translates to polyamine analogue uptake in cancer cells
resulting in inhibition of polyamine biosynthesis and polyamine
catabolism (12). This is a key
reason why polyamine analogues are more effective in cancer
treatment than inhibitors of the polyamine biosynthesis enzymes.
Significant induction of apoptosis by DENSPM was shown to occur in
glioblastoma cell lines in our previous report (3). A preliminary exploration on the
apoptosis-related signals induced by the polyamine analogue,
DENSPM, has revealed that the AKT and the mTOR pathway play an
important role in this process. SSAT expression has been noted to
be induced after DENSPM application. However, its role in the
apoptosis induction by a polyamine analog is far from clear
(3,8).
We first demonstrated that the rapid and significant
increase in SSAT mRNA was associated with marked cell detachment
and cell death in 2 glioblastoma cell lines. Porter and Casero have
reported that the cell growth inhibition was caused by the excess
induction of SSAT (13,14). The SSAT activity in response to
polyamine analogues has been shown to increase in many cell types
(15,16), but cannot be overexpressed in some
cancer cell types (9,17). DENSPM caused marked cell detachment
in U87 cells and to a lesser extent in LN229 cells (Fig. 1). The mechanism responsible for the
observed results of SSAT in different cell lines is not well
understood. Our results support that the specificity of cell types
determines the effect of SSAT cell adherence in response to
DENSPM.
To evaluate whether elevated SSAT expression plays a
key role in DENSPM-induced cell detachment and cell death in
glioblastoma cells, the plasmid construct, PCMV-SSAT was developed
to regulate the expression of SSAT in the tumor cells. Our results
confirmed that the expression of SSAT was involved in determining
the degree of the cell detachment and death consistent with
findings by other groups. Hegardt and colleagues have reported that
the elevation of SSAT activity in DENSPM-treated human breast
cancer cells (L56Br-C1) induces whole cell detachment after 48 h
incubation and a total cell death 72 h later (10). An overexpression of SSAT in kidney
HEK 293 cells was also found to cause morphological shape change,
the loss of cell anchorage, and the significant alteration of
actin-containing filopodia which strongly suggested that the
adhesion defect was associated with the high SSAT expression
(11).
To further confirm that the induction of SSAT is
associated with the cell adhesion, several adhesion-related
proteins were examined. Integrins, control cell migration and
proliferation through the interaction between neighboring cells and
the surrounding extracellular matrix (ECM). Although several
integrins have been recognized as key regulators in glioblastoma
growth and angiogenesis (18),
antagonism of the nonpeptidic α5β1 integrins has demonstrated
anti-proliferative activity in glioblastoma cells and inhibition of
cell adhesion in vitro and in vivo. Antagonism of
α5β1 integrins are believed to inhibit angiogenesis impairing
adhesion and migration of endothelial cells (19,20).
In our study, the expression of integrin α5 and integrin β1 was
down-regulated in PCMV-SSAT transfected LN229 cells detected by
Western blotting. The low expression of both integrin α5 and
integrin β1 following SSAT induction correlates with increased cell
detachment.
Increased SSAT expression was associated with
apoptosis in the glioblastoma cells as demonstrated by decreased
expression of anti-apoptosis-related signaling proteins AKT and
mTOR. Both were deactivated in SSAT up-regulated glioblastomas and
have been recognized as promising targets for therapeutic
interventions (21–23). Moreover, simultaneous blockage of
AKT and mTOR has been shown to markedly affect proliferative arrest
in xenografted tumors (24).
Interestingly, the overexpression of SSAT down-regulated AKT and
mTOR expression in glioblastoma cells suggesting an effect on cell
survival, since the DENSPM-inducing AKT and mTOR down-regulation
has been proven to lead glioma cells to apoptosis (8).
Chen et al have reported that SSAT binds to
integrin α9 and overexpression of SSAT increases integrin
α9β1-mediated migration without decreasing cell viability.
Furthermore, siRNA knockdown of SSAT inhibited this migration
without affecting cell adhesion (25), which is in contrast to our findings.
However, the association of SSAT with α9β1-mediated migration
supports the notion of SSAT’s role in cell adhesion and adhesion
related apoptosis (26,27).
In conclusion, our findings suggest that SSAT plays
a partial role in adhesion related to cancer cell survival as
evidenced by the subpopulation of cancer cells that responded to
SSAT up-regulation. Further studies are warranted to confirm the
function of SSAT in the treatment of glioblastoma.
Acknowledgements
We are grateful to Dr Eugen Park from Department of
Critical Care in St. Michael’s Hospital in Toronto for his
invaluable suggestion and expert editorial review of our
manuscript. This study was supported by the China National Natural
Scientific Fund (30772230).
References
|
1
|
Elmore E, Stringer DE, Steele VE, Gerner
EW and Redpath JL: Chemoprevention by difluoromethylornithine:
correlation of an in vitro human cell assay with human clinical
data for biomarker modulation. Anticancer Res. 21:1163–1165.
2001.PubMed/NCBI
|
|
2
|
Gerner EW and Meyskens FL Jr: Polyamines
and cancer: old molecules, new understanding. Nat Rev Cancer.
4:781–792. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang R, Choi W, Khan A, et al: Activation
of polyamine catabolism by
N1,N11-diethylnorspermine leads to cell death
in glioblastoma. Int J Oncol. 31:431–440. 2007.PubMed/NCBI
|
|
4
|
Pledgie A, Huang Y, Hacker A, et al:
Spermine oxidase SMO(PAOh1), not N1-acetylpolyamine oxidase PAO, is
the primary source of cytotoxic H2O2 in
polyamine analogue-treated human breast cancer cell lines. J Biol
Chem. 280:39843–39851. 2005.PubMed/NCBI
|
|
5
|
Xu H, Chaturvedi R, Cheng Y, et al:
Spermine oxidation induced by Helicobacter pylori results in
apoptosis and DNA damage: implications for gastric carcinogenesis.
Cancer Res. 64:8521–8525. 2004.PubMed/NCBI
|
|
6
|
Choi W, Gerner EW, Ramdas L, et al:
Combination of 5-fluorouracil and N1,N11-diethylnorspermine
markedly activates spermidine/spermine N1-acetyltransferase
expression, depletes polyamines, and synergistically induces
apoptosis in colon carcinoma cells. J Biol Chem. 280:3295–3304.
2005. View Article : Google Scholar
|
|
7
|
Chen Y, Kramer DL, Diegelman P, Vujcic S
and Porter CW: Apoptotic signaling in polyamine analogue-treated
SK-MEL-28 human melanoma cells. Cancer Res. 61:6437–6444.
2001.PubMed/NCBI
|
|
8
|
Jiang R, Choi W, Hu L, Gerner EW, Hamilton
SR and Zhang W: Activation of polyamine catabolism by
N1,N11-diethylnorspermine alters the cellular
localization of mTOR and downregulates mTOR protein level in
glioblastoma cells. Cancer Biol Ther. 6:1644–1648. 2007.PubMed/NCBI
|
|
9
|
Ha HC, Woster PM, Yager JD and Casero RA
Jr: The role of polyamine catabolism in polyamine analogue-induced
programmed cell death. Proc Natl Acad Sci USA. 94:11557–11562.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hegardt C, Johannsson OT and Oredsson SM:
Rapid caspase-dependent cell death in cultured human breast cancer
cells induced by the polyamine analogue
N(1),N(11)-diethylnorspermine. Eur J Biochem. 269:1033–1039. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Z, Zahedi K, Barone S, et al:
Overexpression of SSAT in kidney cells recapitulates various
phenotypic aspects of kidney ischemia-reperfusion injury. J Am Soc
Nephrol. 15:1844–1852. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Casero RA Jr and Marton LJ: Targeting
polyamine metabolism and function in cancer and other
hyperproliferative diseases. Nat Rev Drug Discov. 6:373–390. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Porter CW, Ganis B, Libby PR and Bergeron
RJ: Correlations between polyamine analogue-induced increases in
spermidine/spermine N1-acetyltransferase activity, polyamine pool
depletion, and growth inhibition in human melanoma cell lines.
Cancer Res. 51:3715–3720. 1991.
|
|
14
|
Casero RA Jr, Celano P, Ervin SJ, Wiest L
and Pegg AE: High specific induction of spermidine/spermine
N1-acetyltransferase in a human large cell lung
carcinoma. Biochem J. 270:615–620. 1990.PubMed/NCBI
|
|
15
|
Allen WL, McLean EG, Boyer J, et al: The
role of spermidine/spermine N1-acetyltransferase in
determining response to chemotherapeutic agents in colorectal
cancer cells. Mol Cancer Ther. 6:128–137. 2007.
|
|
16
|
Oredsson SM, Alm K, Dahlberg E, et al:
Inhibition of cell proliferation and induction of apoptosis by
N(1),N(11)-diethylnorspermine-induced polyamine pool reduction.
Biochem Soc Trans. 35:405–409. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Casero RA Jr, Mank AR, Xiao L, Smith J,
Bergeron RJ and Celano P: Steady-state messenger RNA and activity
correlates with sensitivity to
N1,N12-bis(ethyl)spermine in human cell lines
representing the major forms of lung cancer. Cancer Res.
52:5359–5363. 1992.PubMed/NCBI
|
|
18
|
Uhm JH, Gladson CL and Rao JS: The role of
integrins in the malignant phenotype of gliomas. Front Biosci.
4:D188–D199. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maglott A, Bartik P, Cosgun S, et al: The
small alpha5beta1 integrin antagonist, SJ749, reduces proliferation
and clonogenicity of human astrocytoma cells. Cancer Res.
66:6002–6007. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Farber K, Synowitz M, Zahn G, et al: An
alpha5beta1 integrin inhibitor attenuates glioma growth. Mol Cell
Neurosci. 39:579–585. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kesari S, Ramakrishna N, Sauvageot C,
Stiles CD and Wen PY: Targeted molecular therapy of malignant
gliomas. Curr Neurol Neurosci Rep. 5:186–197. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Capdevila J, Salazar R, Halperin I, Abad A
and Yao JC: Innovations therapy: mammalian target of rapamycin
(mTOR) inhibitors for the treatment of neuroendocrine tumors.
Cancer Metastasis Rev. 30:27–34. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Martelli AM, Evangelisti C, Follo MY, et
al: Targeting the phosphatidylinositol 3-kinase/akt/mammalian
target of rapamycin signaling network in cancer stem cells. Curr
Med Chem. 18:2715–2726. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan QW, Knight ZA, Goldenberg DD, et al: A
dual PI3-kinase/mTOR inhibitor reveals emergent efficacy in glioma.
Cancer Cell. 9:341–349. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen C, Young BA, Coleman CS, Pegg AE and
Sheppard D: Spermidine/spermine N1-acetyltransferase
specifically binds to the integrin alpha9 subunit cytoplasmic
domain and enhances cell migration. J Cell Biol. 167:161–170.
2004.
|
|
26
|
Ross EA, Douglas MR, Wong SH, et al:
Interaction between integrin alpha9beta1 and vascular cell adhesion
molecule-1 (VCAM-1) inhibits neutrophil apoptosis. Blood.
107:1178–1183. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Taooka Y, Chen J, Yednock T and Sheppard
D: The integrin alpha9beta1 mediates adhesion to activated
endothelial cells and transendothelial neutrophil migration through
interaction with vascular cell adhesion molecule-1. J Cell Biol.
145:413–420. 1999. View Article : Google Scholar
|