Introduction
Colorectal cancer (CRC) is the third most prevalent
human cancer worldwide, with about 1 million new cases annually. It
is estimated that more than 500 thousand people will die from CRC
every year (1). Despite advances in
both earlier diagnosis through screening and better treatment
modalities which have decreased the mortality in the past decades,
there is still no effective method for increasing the overall
survival rate of affected patients (2). In order to improve the horrible
prognosis, identification of new targets of molecular therapy,
especially those that are indicative of proliferation and
invasiveness, is urgently needed.
Previously, we identified that the overexpression of
S100A4 is associated with lymph node metastasis and poor prognosis
in CRC revealed by proteome analysis (3). S100A4 belongs to the S100 superfamily,
a multi-gene family of Ca2+-binding proteins of the
EF-hand type (4). S100A4-mediated
calcium signaling is involved in a variety of cellular processes,
such as immune response, differentiation, cytoskeleton dynamics,
cell growth, and cell adhesion and motility (5–7).
However, the biological function of S100A4 involved in the
development and/or progression of cancers still remains poorly
understood.
In recent years, RNA interference has been employed
as a powerful strategy for investigating the functions of molecules
that promote the initiation, progression and metastasis of
malignancies by downregulating the expression of targeted molecules
in diverse human cancers (8–12). In
this study, we applied small interfering RNA (siRNA) technique to
specifically suppress S100A4 expression in the CRC cell line SW620,
which expresses a high level of S100A4 and possesses high
invasiveness, to investigate the phenotypic changes including the
proliferative, invasive and metastatic activities. Furthermore, we
explored the significantly dysregulated genes after S100A4
silencing using metastasis-related gene mRNA microarray
analysis.
Materials and methods
CRC cell lines and cell culture
The six CRC cell lines Caco-2, HCT116, SW480, Colo
205, Lovo and SW620 used in this study were obtained from the
American Type Culture Collection (ATCC, Rockville, MD, USA). All
cells were cultured in the corresponding medium (recommended by the
suppliers) supplemented with 10% fetal bovine serum and 1%
penicillin-streptomycin, and were maintained in a 37°C incubator
with 5% CO2.
Real-time PCR
Total-RNA was extracted using the RNeasy mini kit
(Qiagen, Valencia, CA, USA), and the concentration was detected by
a biological spectrophotometer. Real-time PCR analysis was
performed according to the manufacturer’s instructions (the Quant
SYBR-Green PCR kit, Tiangen Bioteck, Beijing, China). β-actin was
applied as an internal control. The primers for β-actin (205 bp)
were 5′-TGACGTGGACATCCGCAAAG-3′ (sense) and
5′-CTGGAAGGTGGACAGCGAGG-3′ (antisense). The primers for S100A4 (185
bp) were 5′-GCCCTGGATGTGATGGTGT-3′ (sense) and
5′-TCGTTGTCCCTGTTGCTGTC-3′ (antisense). Each assay was performed in
triplicate, and the average was calculated. For relative
quantification, 2−ΔΔCt was calculated and used as an
indication of the relative expression level.
Western blotting
A total of 1×107 cells were collected.
The cells were lysed and the protein concentrations were measured
using a BCA Protein Assay Reagent kit (Pierce, Rockford, USA). A 20
μg aliquot of the protein was subjected to 10% SDS-polyacrylamide
gel electrophoresis (PAGE), and then transferred to a
polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA,
USA). After being blocked by incubation overnight in PBST
containing 5% dry nonfat milk, the PVDF membrane was incubated with
rabbit polyclonal antibody against S100A4 (1:1,000 dilution; Abcam,
Cambridge, UK) for 2 h, and then incubated with a
horseradish-peroxidase-conjugated secondary antibody (1:100
dilution; Proteintech, Chicago, IL, USA) for 1 h. Immunoreactive
bands were visualized using an ECL detection system (Amersham,
Arlington Heights, IL, USA) and quantitated by densitometry using
an LAS-3000 imager. β-actin was detected simultaneously as a
loading control (anti-β-actin, 1:1,000 dilution; Kangchen, Beijing,
China).
siRNA transfection
The SW620 cells were divided into three groups,
namely siS100A4, siControl and mock control. The sequence used to
target S100A4 was sense, 5′-GCUGAGCA AGUUCAAUAAATT-3′ and
antisense, 5′-UUUAUUGAAC UUGCUCAGCTT-3′. An unrelated
oligonucleotide (siControl) recognizing an irrelevant transcript
was used as a negative control, and mock control was only
transfected with Oligofectamine reagent. These siRNAs were
purchased from the company GenePharma (Shanghai, China).
Transfection of siRNA, siControl and mock control was performed by
the Oligofectamine kit (Invitrogen, Carlsbad, CA, USA) according to
the manufacturer’s instructions. Briefly, SW620 cells were plated
at 50% confluency with L-15 medium (Gibco, Grand Island, NY, USA)
without antibiotics. Before transfection, culture medium was
changed to Opti-MEM Reduced Serum Medium (Gibco), and then siRNA
and non-sense siRNA were transfected into the cells at the
concentration of 100 nM. Oligofectamine reagent alone (0 nM) was
applied to the cells as a mock control. After 5 h, the medium was
replaced with L-15 medium containing serum. The effectiveness of
gene silencing was determined by real-time PCR and Western
blotting.
Cell proliferation assay
Proliferation was assessed by the Cell Counting
kit-8 (CCK-8; Dojin, Tokyo, Japan). Briefly, cells of mock control,
siControl and siS100A4 were incubated at a density of
1×105/well in 96-well culture plates (Costar, Cambridge,
MA, USA) at 37°C for 7 days, and every 24 h each well of the
cultured cells was incubated with 10 μl of Cell Counting kit-8
solution for 1 h at 37°C. The absorbance at 450 nm was measured
with a spectrophotometer.
Invasion and migration assay
The cell invasive potential of the three groups of
tumor cells was performed using Transwell filters (Costar).
Briefly, 32 h after RNA interference, the filters coated with
Matrigel (BD Biosciences, Bedford, MA, USA) in the upper
compartment were applied with 100 μl of 1×105 cells
seeded in serum-free medium, and the lower compartment was filled
with culture medium supplemented with 10% fetal bovine serum. The
plates were incubated in a humidified 5% CO2 incubator
at 37°C for 36 h, the cells on the top membrane surface were gently
scraped with a cotton swab, and the migrated cells on the bottom
surface were fixed with methanol and counted after staining with
toluidine blue. The cell migration assay was performed in a similar
mode, except that the cells were seeded into the uncoated filter
and incubated for 24 h.
Tumor metastasis PCR array
Total-RNA from siS100A4 and siControl was extracted
with the RNeasy mini kit (Qiagen) and further purified by the use
of a RNeasy MinElute™ Cleanup kit (Qiagen). The RT2
First Strand kit (Qiagen) was employed to produce a cDNA library
for the total-RNA extracted. Following the manufacturer’s protocol,
the cDNA was then processed to perform the Human Tumor Metastasis
RT2 Profiler™ PCR Array (Qiagen, Mississauga, ON,
Canada) containing 84 genes known to be related to tumor
metastasis, five housekeeping genes used for a genomic DNA control,
and three positive controls to ensure high quality data
normalization across samples. The results were analyzed by SA
Biosciences software. For relative quantification,
2−ΔΔCt was calculated and used as an indication of the
relative expression level.
Statistical analysis
All experiments were performed in triplicate.
Results are expressed as means ± standard deviation (SD). A
two-tailed Student’s t-test was performed to analyze the
statistical significance of differences between experimental groups
using the SPSS 11.5 software statistical package. A P-value
<0.05 was considered to denote statistical significance.
Results
S100A4 expression analysis in six human
colorectal cancer cell lines
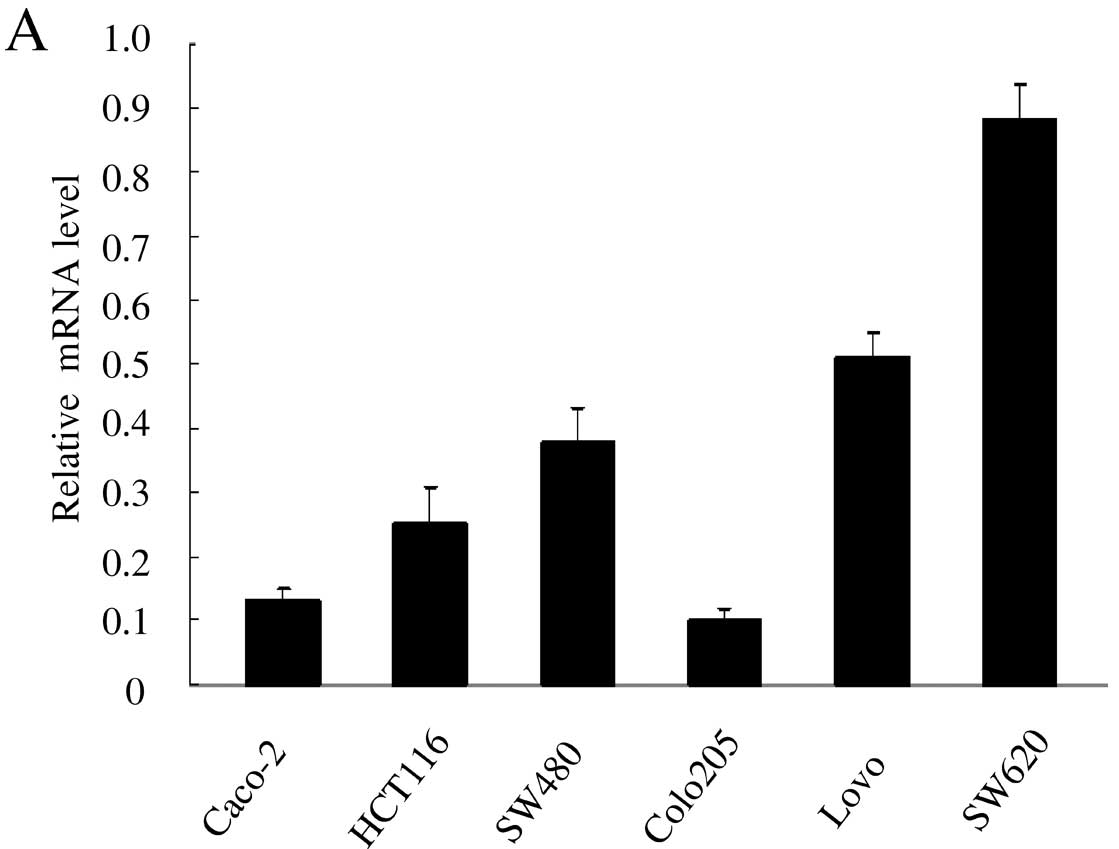
We first examined the expression of S100A4 by
quantitative real-time PCR in six colorectal cancer cell lines. The
result showed that S100A4 expression was the highest in the SW620
cell line followed by Lovo cells, and low in other cell lines,
which roughly coincided with their invasiveness (Fig. 1A). Western blotting also showed the
consistent expression of S100A4 in the six cell lines (Fig. 1B). In addition, many studies have
suggested that the SW620 cell line possesses highly invasive
potential among CRC cell lines. Taken together, we selected the
SW620 cell line for further characterization.
Downregulation of S100A4 expression by
siRNA silencing in SW620 cells
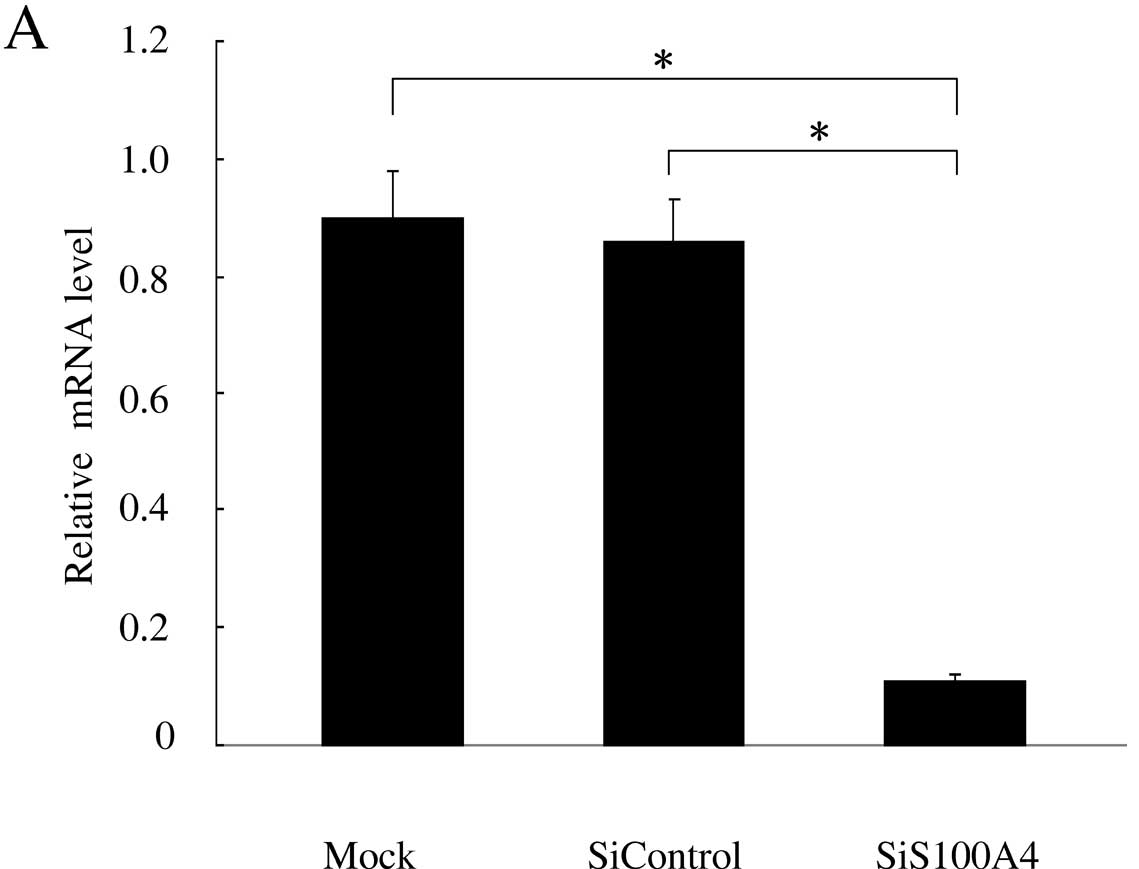
The effect of transfection on the silencing of the
S100A4 gene expression was evaluated using both real-time PCR and
Western blotting (Fig. 2). After 36
h following transfection, real-time PCR analysis showed that the
mRNA expression levels of S100A4 in mock control and siControl of
SW620 cells were 0.9±0.08 and 0.86±0.07 respectively, and there was
no statistical difference between them (P>0.05). The relative
expression of S100A4 was 0.11±0.01 in siS100A4-transfected SW620
cells, which was significantly decreased compared with those in
mock control and siControl (P<0.05) (Fig. 2A). After 48 h following siS100A4
transfection in the SW620 cell lines, Western blot analysis also
revealed that the S100A4 protein expression was significantly
reduced compared with those in the mock control and the siControl,
and the reduction persisted for 6 days (Fig. 2B). The data indicate that RNAi
effectively suppresses S100A4 expression in SW620 colorectal cancer
cells.
Effect of S100A4 knockdown on cell
proliferation
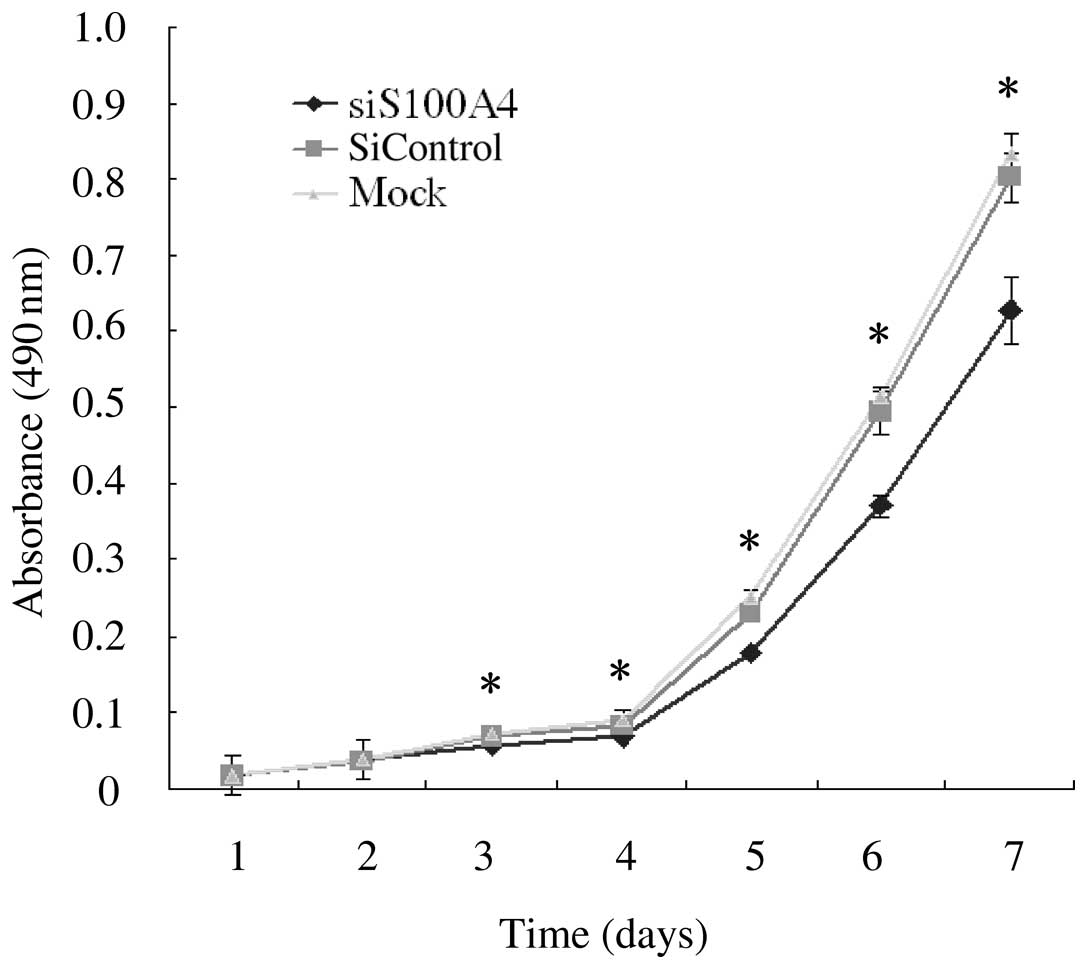
To investigate the effect of S100A4 knockdown on
functional alteration, growth of cells was detected by a cell
proliferation assay. SW620 cells were transfected with siS100A4,
siControl, and mock control, and cell growth was assessed daily for
7 days in all. The growth rate in siS100A4 transfected cells was
significantly slower than those in siControl transfected cells, and
mock-transfected cells (P<0.05 at Days 3, 4, 5, 6 and 7)
(Fig. 3). This result indicates
that S100A4 mediates cell proliferation in SW620 cells and its
suppression leads to inhibition of cell growth in vitro.
Migration and invasion assay
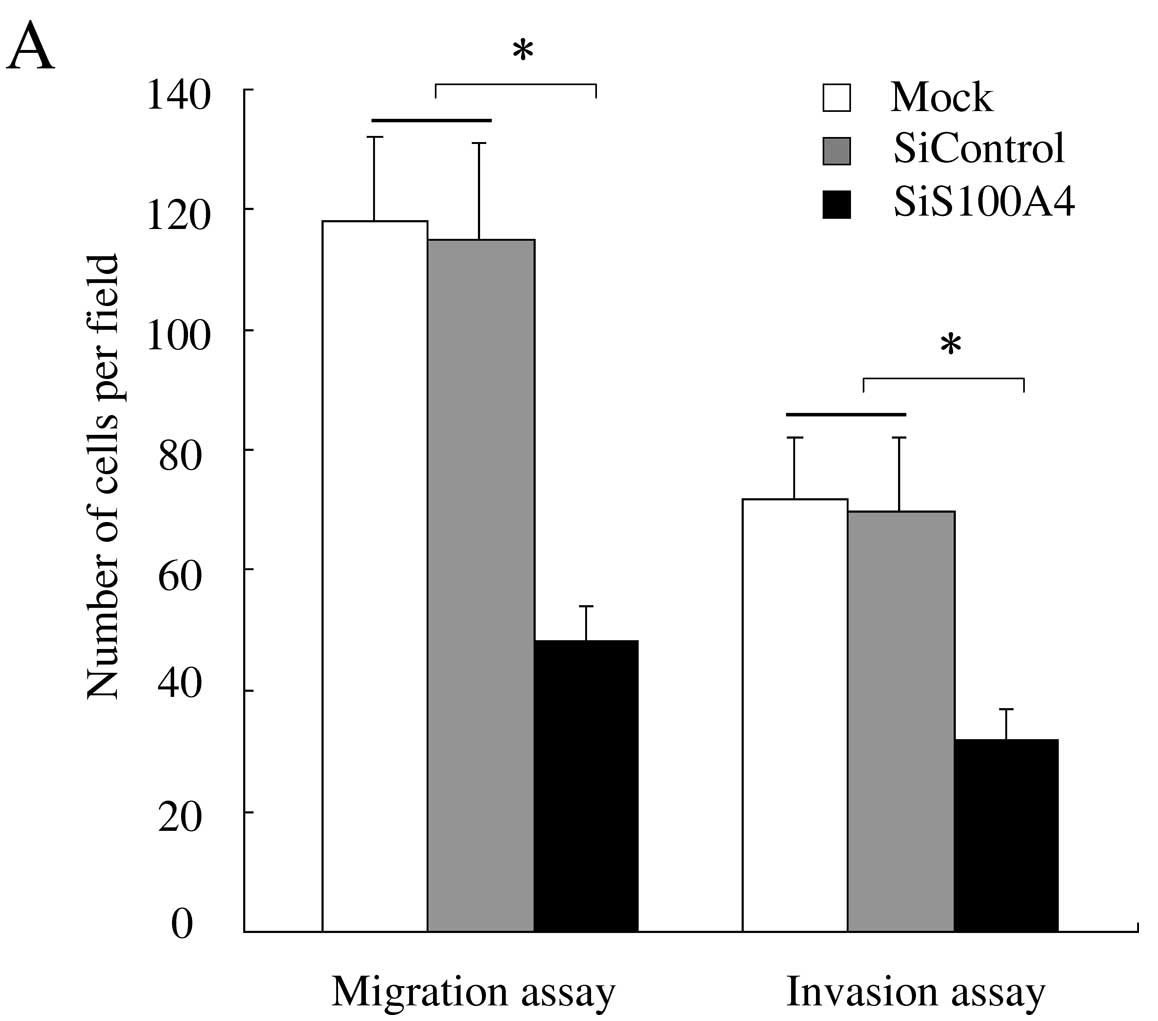
We further studied the effect of S100A4 suppression
on the migration and invasion of SW620 by mobility assays. The cell
migration assay showed that the number of cells that had moved to
the bottom chamber was much smaller in the siS100A4-transfected
cells than in the siControl and mock-transfected cells (P<0.01)
(Fig. 4A). The result of the
Matrigel invasion assay also revealed that the number of invaded
cells transfected with S100A4 siRNA significantly decreased in
comparison with those in the siControl and mock-transfected cells
(P<0.05) (Fig. 4A). Fewer
siS100A4-transfected cells than siControl and mock-transfected
cells were observed when the polycarbonate filters were stained
with toluidine blue (Fig. 4B). Our
data suggested that gene silencing by S100A4-siRNA can inhibit the
invasive potential of SW620.
Expression profiles after knockdown of
S100A4
To further study the possible mechanisms by which
S100A4 modulated the growth, invasion and metastasis potential of
SW620 cells, we purified total-RNA from the SW620-siS100A4 and
SW620-siControl and used PCR array analysis to profile the
expression of tumor metastasis-related genes in response to the
knockdown of S100A4 expression in SW620 cells. A careful analysis
of the results showed that four genes were markedly dysregulated
(>2-fold) after S100A4 silencing in SW620 cells, including three
downregulated genes (MMP10, MMP9 and CDH11) and one upregulated
gene (TIMP4). Based on the attributes and molecular events involved
in the metastasis-related procession, the encoded proteins of these
dysregulated genes could be categorized as extracellular matrix
proteins (MMP9, MMP10, TIMP4) and cell adhesion genes (CDH11)
(Table I).
 | Table IGenes dysregulated by >2-fold after
S100A4 silencing in SW620 cells identified by human tumor
metastasis-related gene mRNA microarray analyses. |
Table I
Genes dysregulated by >2-fold after
S100A4 silencing in SW620 cells identified by human tumor
metastasis-related gene mRNA microarray analyses.
| Abbreviations | mRNA ratioa | GenBank ID | Gene description | Character and
function |
|---|
| TIMP4 | 2.18 | 7079 | Tissue inhibitors of
metalloproteinase 4 | Inhibitors of matrix
metalloproteinases |
| CDH11 | 0.39 | 1009 | Cadherin 11, type 2,
OB-cadherin | Integral membrane
proteins that mediate calcium-dependent cell-cell adhesion |
| MMP9 | 0.46 | 4318 | Matrix
metallo-peptidase 9 | Involved in the
breakdown of the extra-cellular matrix in normal physiological
processes as well as in disease processes |
| MMP10 | 0.40 | 4319 | Matrix
metallo-peptidase 10 | Involved in the
breakdown of the extra-cellular matrix in normal physiological
processes as well as in disease processes |
Discussion
S100A4, also known as 18A2/mts1, CAPL, PEL-98, 42A,
p9Ka, and metastasin, belongs to the S100 superfamily of
calcium-binding proteins (4). It is
located in a 2.05 Mbp segment of the genomic DNA of chromosome 1q21
region (13). The molecule occurs
as non-covalently bound homodimers that can interact with an array
of target proteins in a calcium-dependent manner (14).
Recently, many studies have shown that S100A4 is an
important factor relevant to progression and prognosis in various
human cancers, such as thyroid tumors (15), breast (16), pancreatic (17,18),
lung (19), gastric (20) and colorectal cancers (21–24).
Our previous study found that the overexpression of S100A4 existed
more frequently in colorectal cancer patients with advanced stage,
lymph node metastasis and was associated with poor prognosis
(3). However, the functional
significance of S100A4 in colorectal cancer is still not clear.
Understanding its functional roles and molecular mechanisms
involved in the tumorigenesis and progression of cancers might
provide us the opportunity for the early diagnosis and development
of novel management for malignancies.
In this study, we first examined the expression of
S100A4 in six colorectal cancer cell lines with various invasive
potentials at the mRNA level by real-time PCR and at the protein
level by Western blotting. The level of S100A4 expression among the
six colorectal cancer cell lines roughly coincided with their
invasiveness, which was consistent with our previous conclusion
that increased expression of S100A4 correlated with increased
invasiveness in tumor tissues (3).
To demonstrate the role of S100A4 in tumor development and
progression, we silenced the expression of S100A4 using RNAi in the
colorectal cancer cell line SW620 with highly invasive potential
and S100A4 high expression. In order to validate the effect of
siRNA targeting on the S100A4 gene, we measured the expression of
S100A4 at both the mRNA and protein levels. The results
demonstrated that the RNAi against S100A4 can downregulate the
expression of S100A4 expression effectively at both the mRNA and
protein levels in SW620 cells after transfection. The CCK-8 assay
was used to detect the change of cell proliferation in a
time-dependent manner daily for a total of 7 days. The result
confirmed that the proliferation in siS100A4 cells was reduced
significantly compared with those in the siControl and mock control
(P<0.05). In addition, assessment of the invasive potential,
after transfection with S100A4-siRNA, demonstrated that the rate of
cell migration was significantly reduced compared with those in
siControl and mock control, suggesting that S100A4 may be an
important contributor to the invasion of tumor cells and the
expression level of S100A4 can influence the metastatic behavior of
SW620 cells.
Tumor metastasis is a complicated process, with a
variety of dysregulated molecules playing a significant role
(25,26). We used metastasis-related gene mRNA
microarrays to analyze the dynamic change of genes before and after
S100A4 silencing, which may provide a helpful insight into the
potential mechanism of tumor growth and metastasis inhibition due
to S100A4 silencing. As a result, we identified that four
metastasis-related genes were significantly dysregulated (more than
2-fold) after S100A4 silencing, including three downregulated genes
[matrix metalloproteinase 9 (MMP9), MMP10, and cadherin 11 (CDH11)]
and one upregulated gene [tissue inhibitors of metalloproteinase 4
(TIMP4)] (Table I), suggesting that
S100A4 functions in association with a series of important genes
involved in tumor growth and metastasis, instead of the alteration
of a single molecule.
MMPs belong to a family of zinc- and
calcium-dependent endopeptidases which intrinsically are
responsible for the degradation of the components of the
extracellular matrix (27). In
particular, MMP9 has been identified as a significant molecule
contributing to tumor cell metastasis (28). In a human prostate cell line, the
expression of MMP9 was significantly reduced after S100A4
suppression by RNA interference (29), which was in agreement with our
study. Another MMP, MMP10 was also notably reduced after S100A4
silencing. It has been demonstrated that MMP10 may serve as a
diagnostic and prognostic marker in patients with gastric cancer
and those with oral cancer, suggesting an important role of MMP10
in cancer progression (30,31). Tissue inhibitors of
metalloproteinases (TIMPs) may play an important role in the
metastatic process of cancer cells by metalloproteinase inhibition
and in other pathways independent of MMP inhibition (32). The expression of TIMP4 was revealed
to be a protective factor for prostate cancer progression (33). The mechanisms by which the decreased
expression of MMP9 and MMP10 and the increased of TIMP4 are
responsible for the depressed growth and invasion of tumor cells
due to S100A4 silencing, are currently being investigated in our
laboratory.
Additionally, CDH11, type 2, OB-cadherin also showed
much lower expression after S100A4 silencing. CDH11 is an integral
membrane protein and mediates cell adhesion in a calcium-dependant
manner (34). Overexpression of
this molecule is strongly associated with tumor metastasis and
recurrence in pleomorphic adenomas (35), breast cancer (36,37)
and prostate cancer (38). In
prostate cancer, it has been suggested that downregulation of CDH11
leads to inhibition of tumor cell migration in vivo and
in vitro (38). CDH11
exhibited lower expression after S100A4 silencing in colorectal
cancer cells, suggesting that it may correlate with S100A4. Further
study of this hypothesis is under way.
In conclusion, our present study is the first to
show that knockdown of S100A4 using RNA interference could
effectively inhibit proliferation, invasion and metastasis in SW620
colorectal cancer cells. The function of S100A4 as an oncoprotein
may be associated with several important molecules involved in
growth, invasion and metastasis of cancer cells. These results
further indicate that S100A4 may serve as a potential target for
the development of therapies for colorectal cancer, although
additional studies in vivo are necessary.
Acknowledgements
We thank Zhen Wang for her assistance with the cell
culture experiments, Yue Cao for her comments regarding this study
and Mengyun Wang for kindly providing siRNA transfection
construction. This study was supported by the Shanghai Rising-star
Program from the Science and Technology Commission of Shanghai
Municipality (no. 10QA1401400, China), Fudan University Shanghai
Cancer Center Grant (no. YJ201005).
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
2
|
Huerta S: Recent advances in the molecular
diagnosis and prognosis of colorectal cancer. Expert Rev Mol Diagn.
8:277–288. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang LY, Xu Y, Cai GX, Guan ZQ, Sheng WQ,
Lu HF, Xie LQ, Lu HJ and Cai SJ: S100A4 overexpression underlies
lymph node metastasis and poor prognosis in colorectal cancer.
World J Gastroenterol. 17:69–78. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marenholz I, Volz A, Ziegler A, Davies A,
Ragoussis I, Korge BP and Mischke D: Genetic analysis of the
epidermal differentiation complex (EDC) on human chromosome 1q21:
chromosomal orientation, new markers, and a 6-Mb YAC contig.
Genomics. 37:295–302. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mazzucchelli L: Protein S100A4: too long
overlooked by pathologists? Am J Pathol. 160:7–13. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marenholz I, Heizmann CW and Fritz G: S100
proteins in mouse and man: from evolution to function and pathology
(including an update of the nomenclature). Biochem Biophys Res
Commun. 322:1111–1122. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barraclough R: Calcium-binding protein
S100A4 in health and disease. Biochim Biophys Acta. 1448:190–199.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Micklem DR and Lorens JB: RNAi screening
for therapeutic targets in human malignancies. Curr Pharm
Biotechnol. 8:337–343. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tuschl T and Borkhardt A: Small
interfering RNAs: a revolutionary tool for the analysis of gene
function and gene therapy. Mol Interv. 2:158–167. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gentile A, Lazzari L, Benvenuti S,
Trusolino L and Comoglio PM: Ror1 is a pseudokinase that is crucial
for Met-driven tumorigenesis. Cancer Res. 71:3132–3141. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang Z, Huang S, Wang Q, Liang L, Ni S,
Wang L, Sheng W, He X and Du X: MicroRNA-95 promotes cell
proliferation and targets sorting Nexin 1 in human colorectal
carcinoma. Cancer Res. 71:2582–2589. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mortazavi F, Dubinett S and Rettig M:
c-Crk proto-oncogene contributes to transcriptional repression of
p120-catenin in non-small cell lung cancer cells. Clin Exp
Metastasis. 28:391–404. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mischke D, Korge BP, Marenholz I, Volz A
and Ziegler A: Genes encoding structural proteins of epidermal
cornification and S100 calcium-binding proteins form a gene complex
(‘epidermal differentiation complex’) on human chromosome 1q21. J
Invest Dermatol. 106:989–992. 1996.PubMed/NCBI
|
|
14
|
Vallely KM, Rustandi RR, Ellis KC,
Varlamova O, Bresnick AR and Weber DJ: Solution structure of human
Mts1 (S100A4) as determined by NMR spectroscopy. Biochemistry.
41:12670–12680. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zou M, Al-Baradie RS, Al-Hindi H, Farid NR
and Shi Y: S100A4 (Mts1) gene overexpression is associated with
invasion and metastasis of papillary thyroid carcinoma. Br J
Cancer. 93:1277–1284. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rudland PS, Platt-Higgins A, Renshaw C,
West CR, Winstanley JH, Robertson L and Barraclough R: Prognostic
significance of the metastasis-inducing protein S100A4 (p9Ka) in
human breast cancer. Cancer Res. 60:1595–1603. 2000.PubMed/NCBI
|
|
17
|
Ai KX, Lu LY, Huang XY, Chen W and Zhang
HZ: Prognostic significance of S100A4 and vascular endothelial
growth factor expression in pancreatic cancer. World J
Gastroenterol. 14:1931–1935. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ikenaga N, Ohuchida K, Mizumoto K, Yu J,
Fujita H, Nakata K, Ueda J, Sato N, Nagai E and Tanaka M: S100A4
mRNA is a diagnostic and prognostic marker in pancreatic carcinoma.
J Gastrointest Surg. 13:1852–1858. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsuna M, Kageyama S, Fukuoka J, Kitano H,
Doki Y, Tezuka H and Yasuda H: Significance of S100A4 as a
prognostic marker of lung squamous cell carcinoma. Anticancer Res.
29:2547–1554. 2009.PubMed/NCBI
|
|
20
|
Wang YY, Ye ZY, Zhao ZS, Tao HQ and Chu
YQ: High-level expression of S100A4 correlates with lymph node
metastasis and poor prognosis in patients with gastric cancer. Ann
Surg Oncol. 17:89–97. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gongoll S, Peters G, Mengel M, Piso P,
Klempnauer J, Kreipe H and von Wasielewski R: Prognostic
significance of calcium-binding protein S100A4 in colorectal
cancer. Gastroenterology. 123:1478–1484. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Boye K, Nesland JM, Sandstad B, Maelandsmo
GM and Flatmark K: Nuclear S100A4 is a novel prognostic marker in
colorectal cancer. Eur J Cancer. 46:2919–2925. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kwak JM, Lee HJ, Kim SH, Kim HK, Mok YJ,
Park YT, Choi JS and Moon HY: Expression of protein S100A4 is a
predictor of recurrence in colorectal cancer. World J
Gastroenterol. 16:3897–3904. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang HY, Zhang JY, Cui JT, Tan XH, Li WM,
Gu J and Lu YY: Expression status of S100A14 and S100A4 correlates
with metastatic potential and clinical outcome in colorectal cancer
after surgery. Oncol Rep. 23:45–52. 2010.PubMed/NCBI
|
|
25
|
Royston D and Jackson DG: Mechanisms of
lymphatic metastasis in human colorectal adenocarcinoma. J Pathol.
217:608–619. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sundar SS and Ganesan TS: Role of
lymphangiogenesis in cancer. J Clin Oncol. 25:4298–4307. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brinckerhoff CE and Matrisian LM: Matrix
metalloproteinases: a tail of a frog that became a prince. Nat Rev
Mol Cell Biol. 3:207–214. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saleem M, Kweon MH, Johnson JJ, Adhami VM,
Elcheva I, Khan N, Bin HB, Bhat KM, Sarfaraz S, Reagan-Shaw S,
Spiegelman VS, Setaluri V and Mukhtar H: S100A4 accelerates
tumorigenesis and invasion of human prostate cancer through the
transcriptional regulation of matrix metalloproteinase 9. Proc Natl
Acad Sci USA. 103:14825–14830. 2006. View Article : Google Scholar
|
|
30
|
Aung PP, Oue N, Mitani Y, Nakayama H,
Yoshida K, Noguchi T, Bosserhoff AK and Yasui W: Systematic search
for gastric cancer-specific genes based on SAGE data: melanoma
inhibitory activity and matrix metalloproteinase-10 are novel
prognostic factors in patients with gastric cancer. Oncogene.
25:2546–2547. 2006. View Article : Google Scholar
|
|
31
|
Yen CY, Chen CH, Chang CH, Tseng HF, Liu
SY, Chuang LY, Wen CH and Chang HW: Matrix metalloproteinases (MMP)
1 and MMP10 but not MMP12 are potential oral cancer markers.
Biomarkers. 14:244–249. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Brew K and Nagase H: The tissue inhibitors
of metalloproteinases (TIMPs): an ancient family with structural
and functional diversity. Biochim Biophys Acta. 1803:55–71. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Riddick AC, Shukla CJ, Pennington CJ, Bass
R, Nuttall RK, Hogan A, Sethia KK, Ellis V, Collins AT, Maitland
NJ, Ball RY and Edwards DR: Identification of degradome components
associated with prostate cancer progression by expression analysis
of human prostatic tissues. Br J Cancer. 92:2171–2180. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Okazaki M, Takeshita S, Kawai S, Kikuno R,
Tsujimura A, Kudo A and Amann E: Molecular cloning and
characterization of OB-cadherin, a new member of cadherin family
expressed in osteoblasts. J Biol Chem. 269:12092–12098.
1994.PubMed/NCBI
|
|
35
|
Brieger J, Duesterhoeft A, Brochhausen C,
Gosepath J, Kirkpatrick CJ and Mann WJ: Recurrence of pleomorphic
adenoma of the parotid gland - predictive value of cadherin-11 and
fascin. APMIS. 116:1050–1057. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li Z, Zhou Z and Donahue HJ: Alterations
in Cx43 and OB-cadherin affect breast cancer cell metastatic
potential. Clin Exp Metastasis. 25:265–272. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tamura D, Hiraga T, Myoui A, Yoshikawa H
and Yoneda T: Cadherin-11-mediated interactions with bone marrow
stromal/osteoblastic cells support selective colonization of breast
cancer cells in bone. Int J Oncol. 33:17–24. 2008.PubMed/NCBI
|
|
38
|
Chu K, Cheng CJ, Ye X, Lee YC, Zurita AJ,
Chen DT, Yu-Lee LY, Zhang S, Yeh ET, Hu MC, Logothetis CJ and Lin
SH: Cadherin-11 promotes the metastasis of prostate cancer cells to
bone. Mol Cancer Res. 6:1259–1267. 2008. View Article : Google Scholar : PubMed/NCBI
|