Introduction
Extensive research has established that
neoangiogenesis plays a pivotal role in solid malignant tumour
growth and metastasis (1–3). Tumours cannot exceed a few millimetres
in diameter without the development of a neovasculature supply.
Thus, antiangiogenesis therapy is a potentially promising
tumouristatic approach (4–6). However, evidence has emerged that
angiogenesis is tightly regulated by a balance of activating and
inhibiting factors (7,8). Therefore, long-term overexpression of
angiogenesis suppressors may be required for effectively
controlling tumour proliferation through counteracting
tumour-induced angiogenesis.
Pigment epithelium-derived factor (PEDF) is a 50-kDa
secreted glycoprotein that belongs to the serine protease inhibitor
superfamily but lacks protease inhibitor activity (9–11).
PEDF was first identified and purified from the conditioned medium
of cultured human neonatal retinal pigment epithelial cells
(9,12). This factor is involved in multiple
and varied biological activities (13), which makes it an appealing potential
treatment for Lewis lung carcinoma (LCC). With regard to its
antiangiogenic activity, PEDF is more potent than any other
endogenous inhibitors of neovascularisation (14,15);
this property makes PEDF an excellent candidate for LLC treatment
as a form of targeted gene therapy. The antiangiogenic potency of
PEDF has been shown to inhibit tumour angiogenesis in several
preclinical cancer models (16–27).
In addition, PEDF is thought to exert its antiangiogenic activity
through two major pathways, namely, endothelial cell apoptosis via
activation of the Fas/Fas-L death pathway (28), and disruption of the crucial balance
between pro- and anti-angiogenic factors, via downregulation of
vascular endothelial growth factor (VEGF) expression (29–31).
Furthermore, a few recent reports indicate that PEDF not only acts
to halt angiogenesis, but also has the ability to increase
apoptosis in tumours (16–18,26).
This apoptotic activity is likely due to a distinct functional
epitope on the PEDF protein (20).
Successful antiangiogenic therapy requires efficient
and continuous secretion of the candidate protein for long periods
of time. Gene transfer is usually utilized as an effective strategy
for chronic delivery of antiangiogenic factors. Adeno-associated
virus (AAV) vectors represent a very promising tool for cancer gene
therapy because they are capable of sustained, long-term gene
expression, non-pathogenicity, low immunogenicity, and they lack
cytotoxicity (32–34).
In this study, we constructed AAV vectors that
express PEDF in order to investigate the effect of AAV-mediated
intratumoural PEDF expression on LLC tumour suppression.
Materials and methods
Cell lines and culture
HUVECs were isolated and cultured in DMEM medium
(Gibco-BRL, NY, USA) supplemented with 20% FBS and 100 g/ml bovine
fibroblast growth factor (BFGF). LLC cells were obtained from the
American Type Culture Collection (ATCC, Rockville, MD, USA) and
cultured in DMEM medium (Gibco-BRL) supplemented with 10% FBS and
100 μg/ml amikacin.
AAV-PEDF preparation
AAV-PEDF was constructed using CMV as the promoter.
cDNAs containing full-length human PEDF sequences under the CMV
promoter were cloned. The construct sequence was confirmed via DNA
sequencing (Invitrogen Inc., Shanghai). A control virus containing
a green fluorescent protein cDNA under the same promoter (AAV-EGFP)
was also detected by DNA sequencing (Invitrogen Inc.). Packaging
and purification of rAAV particles were performed as previously
described (35).
Cell infection with AAV-PEDF
LLC cells were seeded in 6-well plates. After an
overnight culture, the cells were infected with AAV-PEDF and
AAV-EGFP viruses at a multiplicity of infection (MOI) of
1×105 infectious particles/cell. The cells and
supernatants were collected after 72 h.
Western blot analysis
LLC cells seeded in the 6-well plates were harvested
and resuspended in lysis buffer while the LLC tumours were grinded
and then lysed with RIPA solution, respectively. Equal amounts of
protein were separated by SDS-polyacrylamide gel electrophoresis
(PAGE) and then electrotransferred onto a polyvinylidene difluoride
membrane (PVDF). Blots were probed with a goat anti-human PEDF
monoclonal antibody (1:1000, mAb; R&D Systems, Boston, MA, USA)
plus a secondary biotinylated antibody against goat IgG (1:10,000,
ZSGB-BIO, Beijing, China). Immunoreactivity was detected using an
enhanced chemiluminescence (ECL) detection system (Pierce,
Rockford, IL, USA).
Animal experiments
Male C57BL/6 mice were purchased from the
Experimental Animal Centre at Sichuan University. All animal
experimental procedures were approved by the West China Hospital
Cancer Centre’s Animal Care and Use Committee. Aliquots of LLC
cells (5×105) were subcutaneously inoculated into the
mice. When the average tumour volume reached 90–100 mm3
in size, the mice were randomly divided into 3 groups. Each mouse
in the AAV-PEDF group was treated with an intratumoural injection
of 2×1010 AAV-PEDF virus particles. The mice in the
control groups received 2×1010 AAV-EGFP particles or
normal saline (NS). Tumour sizes and animal weights were measured
every three days. The tumour volumes (mm3) were
calculated according to the following formula: (length ×
width2 × 0.52). Mouse sera were collected for liver and
kidney function analyses on the fifteenth day post-treatment using
an AU7020 Automatic Biochemical Analyzer (Hitachi).
TUNEL assay, immunohistochemistry
analysis and H&E staining
Apoptotic tumour cells were determined by the
DeadEnd colorimetric terminal deoxynucleotidyl transferase-mediated
dUTP nick-end labelling (TUNEL) System (Promega Corp., Madison,
USA) and the caspase-3 immunohistochemical assay. Prepared tumour
cryosections were incubated with primary anti-human PEDF antibody
(1:200, mAb; R&D Systems) overnight, then with biotinylated
anti-goat IgG secondary antibody (ZSJQ Biotechnology) and finally
with diaminobenzidine (DAB; ZSJQ Biotechnology) as a substrate for
visualization of the antigen-antibody complex. Frozen tumour
specimens were analyzed by CD31 immunohistochemistry. The
microvessel density was quantified using the reported method of
Weidner et al (36).
Paraffin sections were also stained with haematoxylin and eosin
(H&E) to observe the structure and histological morphology of
the tumours and basic organs.
Alginate-encapsulated tumour cell and
tube formation assay
LLC cells were resuspended in alginate solution. LLC
cells in alginate solution were dropped into a swirling 0.25 M
CaCl2 solution to prepare alginate beads
(1×105 cells/bead). Male C57Bl/6 mice were implanted
s.c. with alginate beads into the back (1 bead/side). The next day,
the mice were treated with i.v. administration of 2×1010
particles AAV-PEDF per mouse, or 2×1010 particles
AAV-EGFP per mouse or with NS.
On Day 11 after treatment, 0.1 ml of 1% FITC-dextran
solution (100 mg/kg) was injected i.v. into the tail vein of the
mice. Alginate beads were photographed and rapidly removed 20 min
after FITC-dextran injection. The beads were then vortexed and
centrifuged in tubes containing 2 ml NS and the supernatant
fluorescence was measured.
The tube formation assay was performed using
Matrigel (BD Biosciences, San Jose, CA) that was thawed overnight
at 4°C. Pre-chilled 24-well plates were coated with 300 μl/well of
Matrigel (BD Biosciences). HUVECs were seeded in each well at a
concentration of 1×105 cells, and then treated with
conditioned medium (CM) from LLC cells infected in vitro
with AAV-PEDF, AAV-EGFP or NS. Tubule branches were photographed 6
h after incubation.
Statistical analysis
Values are presented as means ± SD. Statistical
analysis was conducted using the SPSS program (version 18.0, SPSS
Inc., Chicago, IL, USA). The statistical significances were
calculated by one-way ANOVA. The Kaplan-Meier method was used to
evaluate survival curves and survival rate among groups. A P-value
<0.05 was considered significant.
Results
PEDF gene expression in vivo and in
vitro
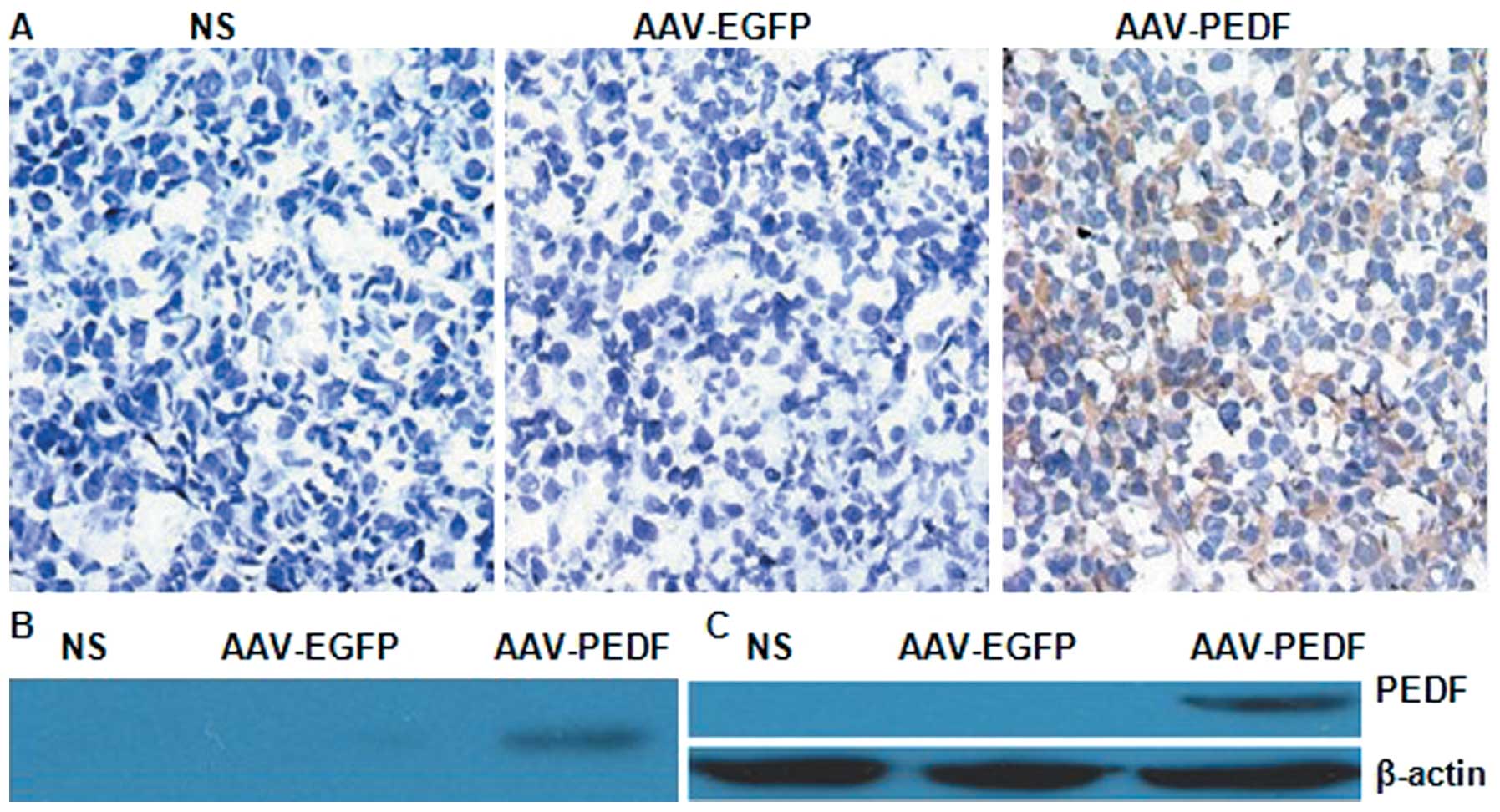
To confirm PEDF gene expression in vivo and
in vitro, western blotting was performed. PEDF protein was
detected only in LLC tumours from AAV-PEDF treated mice (Fig. 2B). Human PEDF was detected as a
single 50 kDa band in AAV-PEDF transduced cells (Fig. 2C). No PEDF was found in the lysates
from AAV-EGFP-transduced or NS-treated cells. Seven days after
inoculation, tumours were injected with AAV-EGFP, AAV-PEDF, or NS,
and only the cytoplasm of LLC cells treated with AAV-PEDF clearly
was stained for intratumoural PEDF (Fig. 2A). These results demonstrate
successful PEDF gene expression in vivo and in
vitro.
Intratumoural treatment of AAV-PEDF
suppresses tumour growth and prolongs mouse survival rate
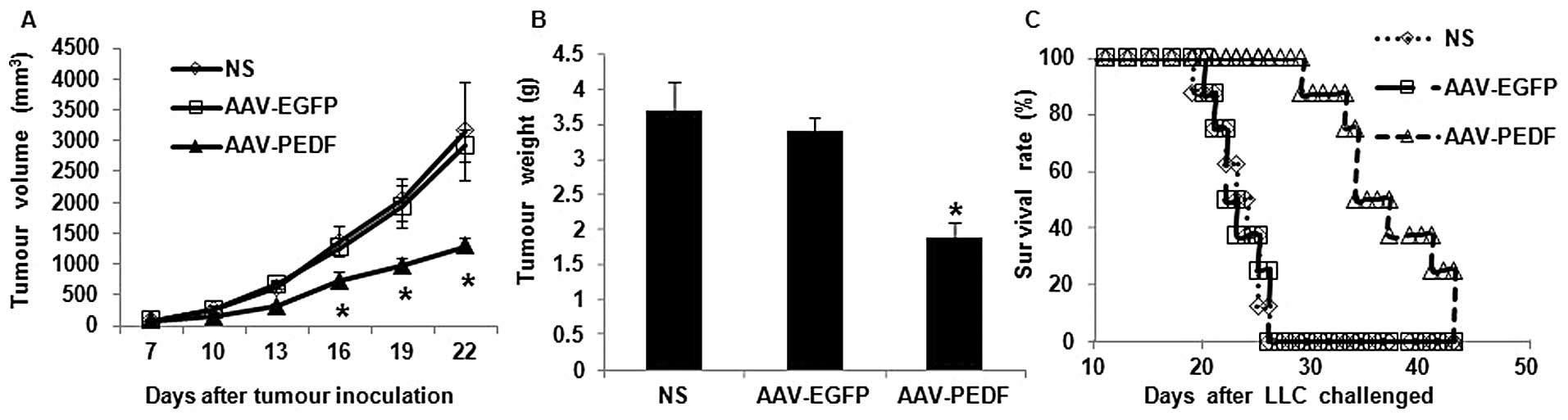
Established subcutaneous LLC tumours in the C57BL/6
mouse model were used to investigate the antitumoural efficacy of
AAV-PEDF by intratumoural injection. There were no significant
differences in size between those tumours receiving NS vs. AAV-EGFP
(Fig. 1A). In contrast, the growth
of AAV-PEDF-treated LLC tumours was significantly inhibited
compared with either AAV-EGFP or NS (Fig. 1A, P<0.05). The maximum tumour
growth inhibition was observed on day 15 after treatment (56 and
58% inhibition, respectively, compared with mice treated with
AAV-EGFP or NS, Fig. 1A). Tumour
weights in the AAV-PEDF group were significantly lower than either
the AAV-EGFP or NS groups (Fig. 1B,
P<0.05). In addition, intratumoural treatment of AAV-PEDF
resulted in prolonged animal survival (Fig. 1C, P<0.05).
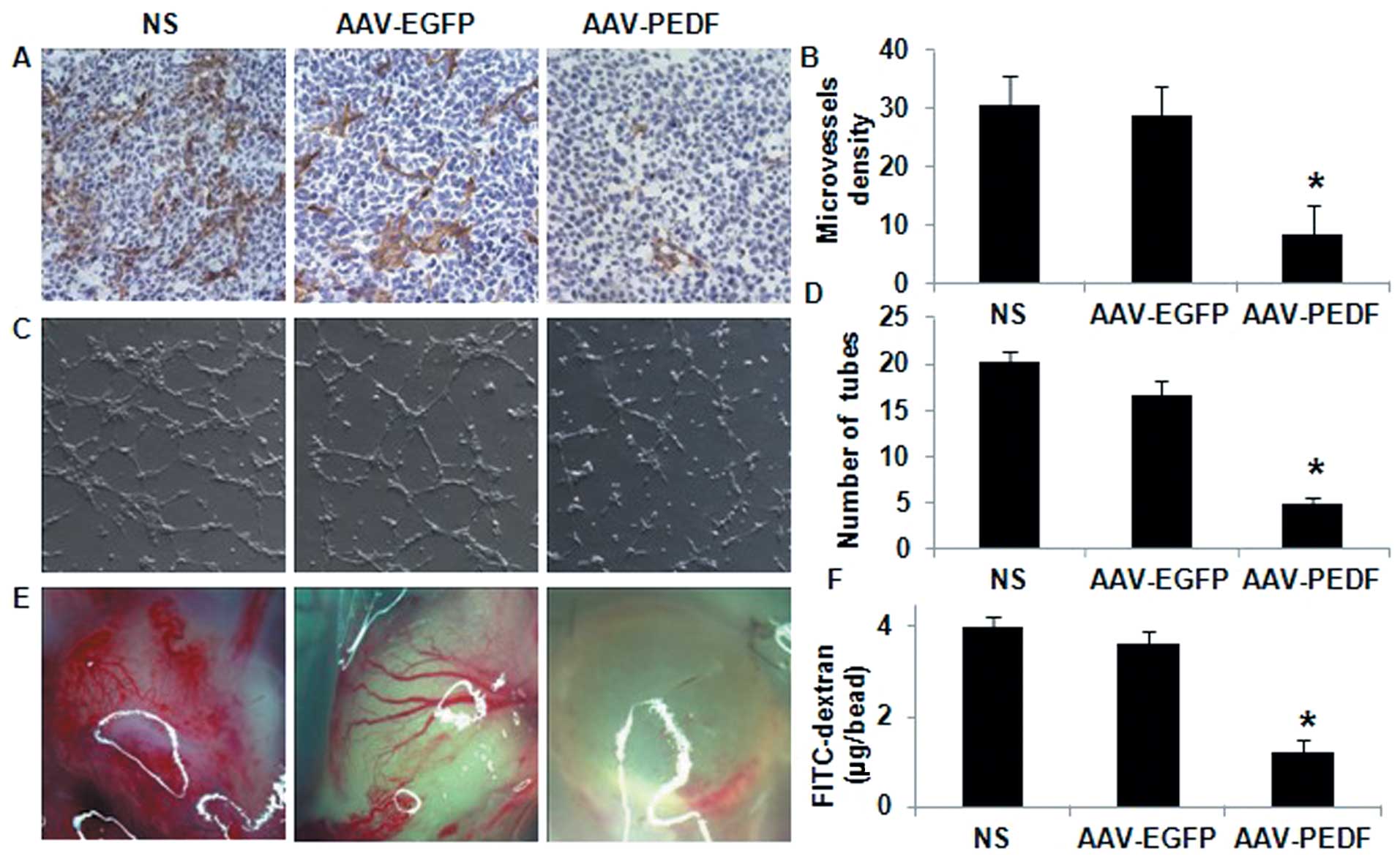
AAV-PEDF treatment decreases microvessel
density and increases apoptosis in tumour tissue
Tumours from AAV-PEDF-treated animals showed marked
reduction in microvessel density (MVD) (Fig. 3A). Quantitative analysis showed a 71
and 73% reduction in intratumoural MVD in AAV-PEDF treated animals
compared with control animals receiving AAV-EGFP and NS,
respectively (Fig. 3B). There was
no significant difference in vessel density between tumours that
received NS or AAV-EGFP (Fig. 3B;
NS vs. AAV-EGFP, P>0.1).
More apoptotic cells in tumour tissue were observed
in AAV-PEDF-treated animals than in AAV-EGFP- or NS-treated animals
(Fig. 4A). The average percentage
of apoptosis in the AAV-PEDF group was significantly increased
compared with that in the AAV-EGFP and NS groups (Fig. 4B; P<0.05). Furthermore, there
were more cell nuclei stained in the AAV-PEDF group compared with
the other groups (Fig. 4C and D;
P<0.05). As shown in Fig. 4E,
histological analysis revealed that AAV-PEDF-treated tumours had
increased cell death necrosis.
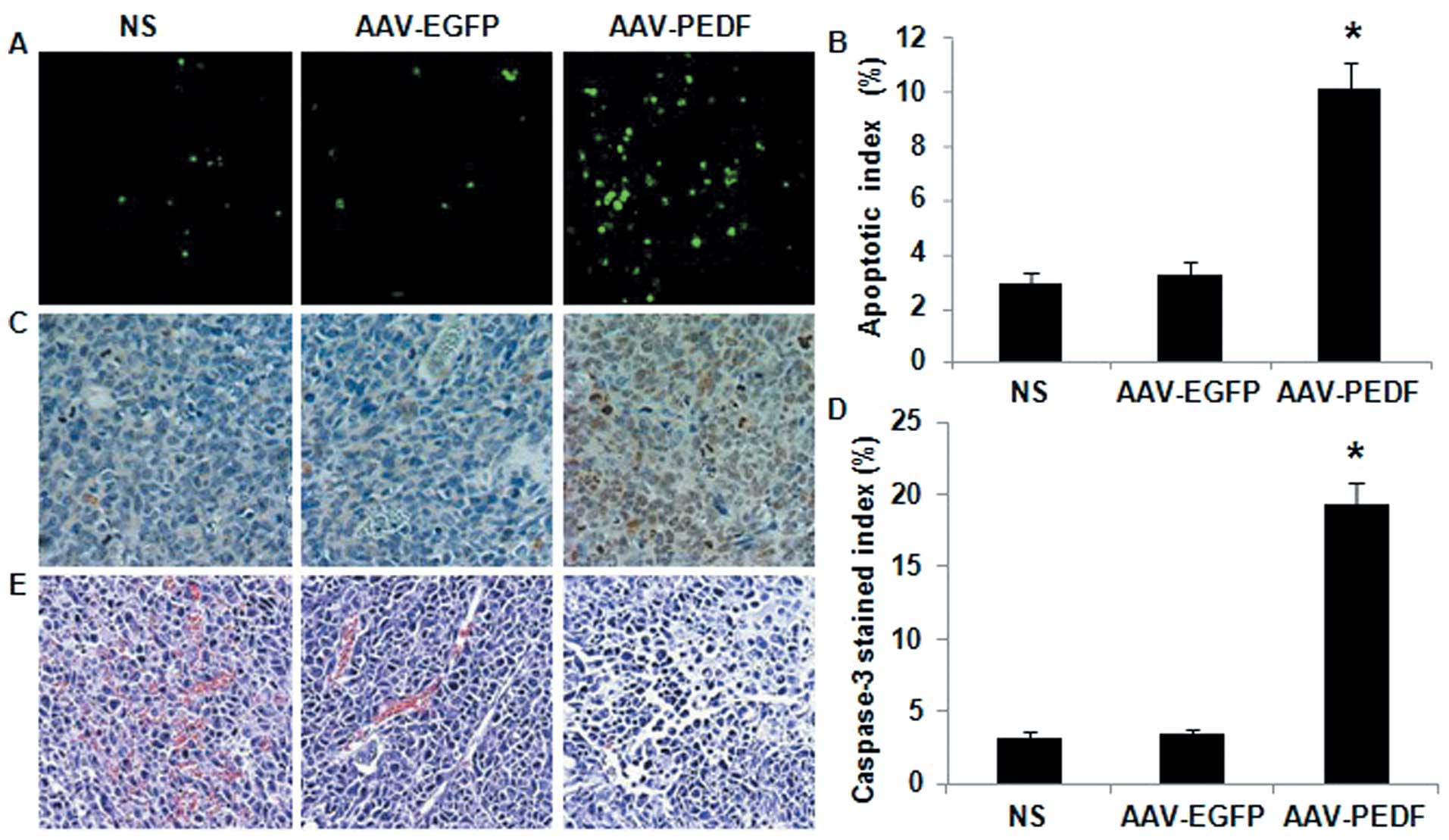 | Figure 4TUNEL, caspase-3 immunohistochemical
assay, and histological staining for tumour tissue. (A) Apoptotic
cells within LLC tumours were detected by the TUNEL assay (original
magnification, ×400). (B) The apoptotic index (%) of LLC tumours
treated with NS, AAV-EGFP, and AAV-PEDF (*P<0.05).
(C) Micrographs show tumour tissue sections stained with
anti-caspase-3 antibody (original magnification, ×400) after NS,
AAV-EGFP or AAV-PEDF treatment. (D) The caspase-3 stained index (%)
of tumours treated with NS, AAV-EGFP, and AAV-PEDF
(*P<0.05). (E) LLC tumours from NS, AAV-EGFP, and
AAV-PEDF treated mice were stained with H&E (original
magnification, ×400); noticeable necrosis was observed in tumours
from AAV-PEDF treated mice. |
AAV-PEDF inhibition of angiogenesis in
vitro and in vivo
Endothelial cells form tube and capillary-like
structures on a Matrigel membrane through a process involving
attachment, alignment, and migration. Treatment with conditioned
medium (CM) from LLC cells infected in vitro with AAV-PEDF
significantly decreased this process (Fig. 3C). Quantitative analysis showed that
AAV-PEDF restricted tube formation by 71 and 76%, respectively, in
comparison to the AAV-EGFP or NS controls (Fig. 3D; P<0.01).
Alginate implants in AAV-PEDF-treated animals
exhibited less vascularisation than AAV-EGFP- or NS-treated animals
(Fig. 3E). We observed that the
accumulation of FITC-dextran at the implant site in the AAV-PEDF
group was significantly lower compared to the other groups
(Fig. 3F, P<0.05).
The data confirmed that AAV-PEDF-mediated PEDF gene
transfer and expression could suppress tumour angiogenesis in the
studied tumour model.
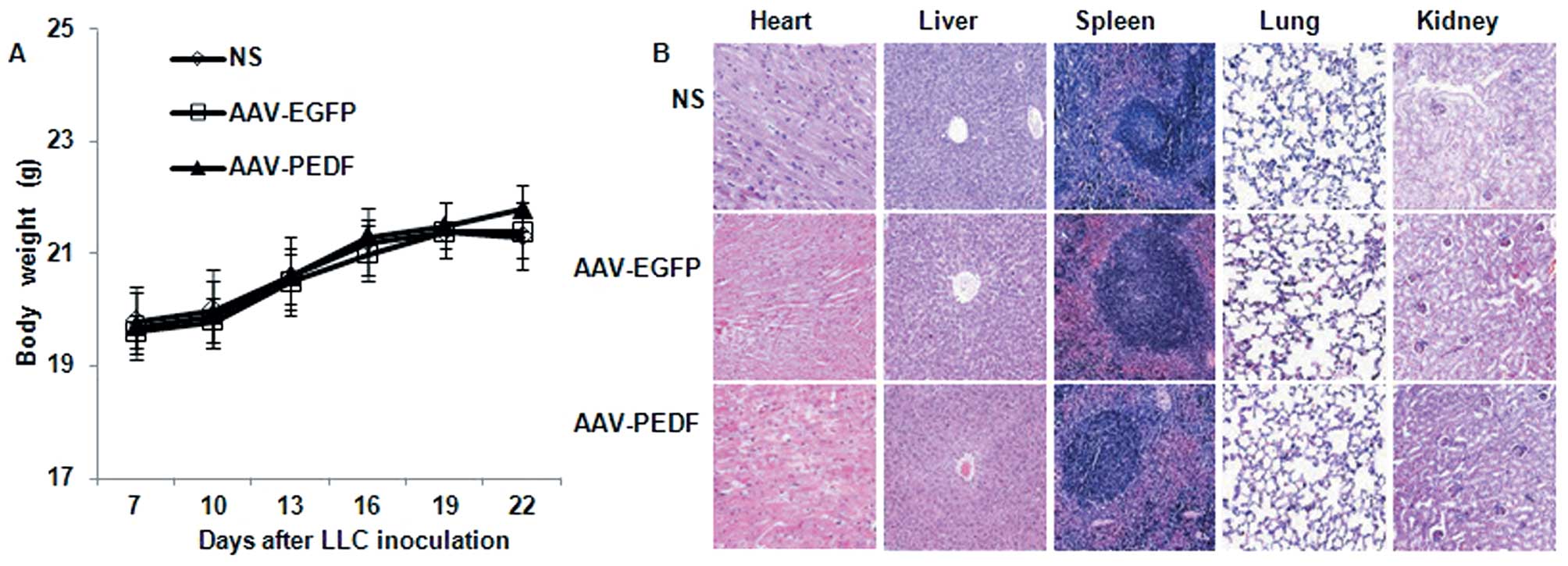
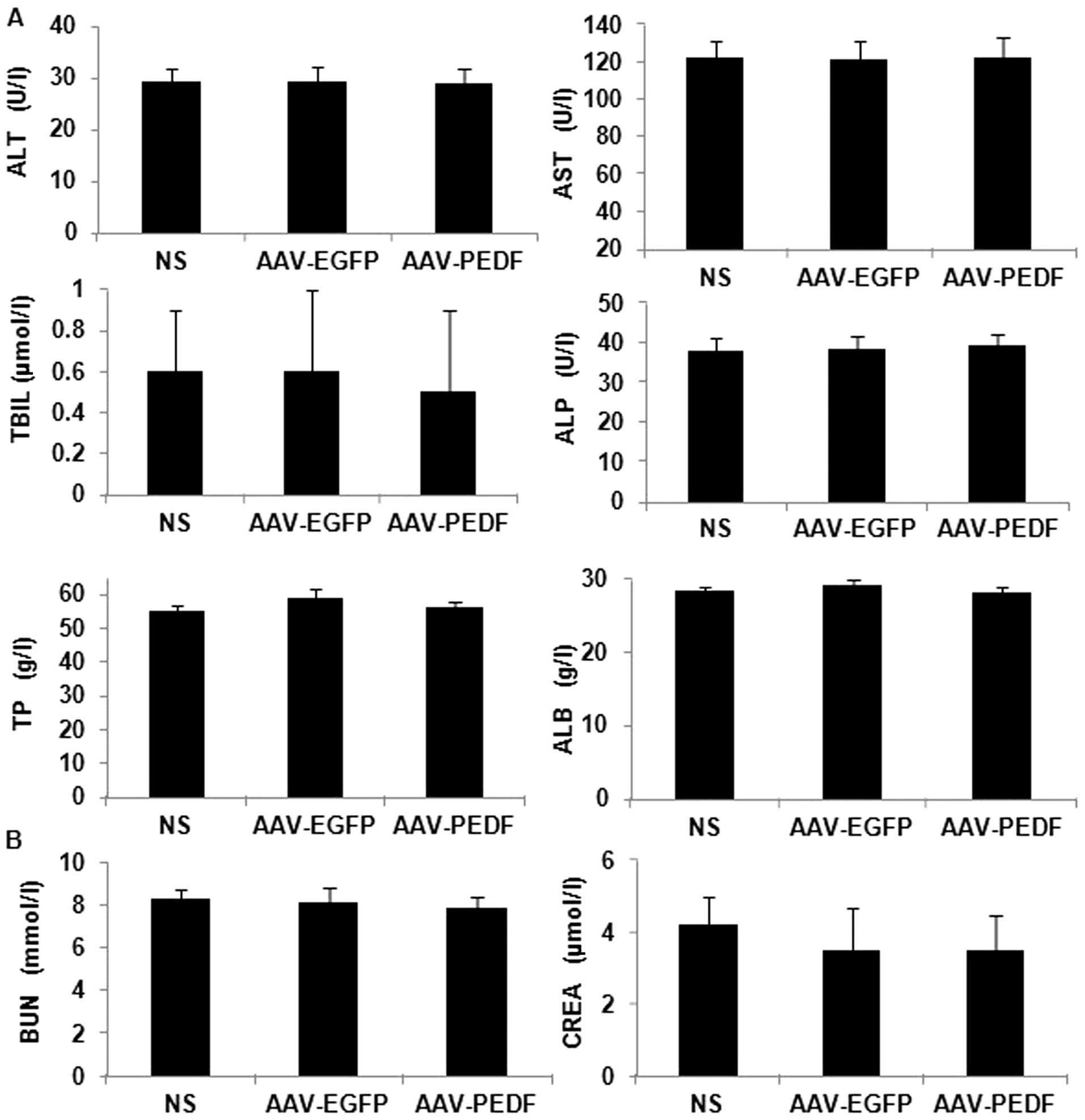
AAV toxicity in mice
We evaluated AAV toxicity in mice by monitoring
changes in mouse weight, basic organ structure, and histological
morphology, and analyzing mouse liver and kidney function. No
significant difference was observed in body weight among AAV-EGFP-,
AAV-PEDF- or NS-treated mice (Fig.
5A, P>0.1). Liver and kidney function data are shown in
Fig. 6 (P>0.1). In addition, no
apparent pathological changes were observed in the heart, liver,
spleen, lung, and kidney tissue from the different groups as
indicated by H&E staining (Fig.
5B).
Discussion
Angiogenesis, the complex biological process by
which new blood vessels develop from pre-existing ones, is known to
play an essential role in supporting progressive tumour growth
(1–3). Therefore, targeting neoangiogenesis or
signals that promote neovessel growth is a promising anticancer
therapeutic strategy (4–6). However, neovasculature growth is
controlled by maintaining a balance between pro- and
anti-angiogenic factors (7,8). For this reason, overexpression of
anti-angiogenesis factors could be excellent therapeutic tools in
combating tumour angiogenesis.
Combating lung cancer remains a major clinical
challenge. Existing therapeutic protocols are very disappointing.
Previous studies have demonstrated the efficacy of
anti-angiogenesis therapy (4–6). In
this study, we demonstrated that overexpression of PEDF mediated by
the AAV vector exerts a remarkable suppression of tumour growth and
prolongs animal survival in a C57BL/6 mouse model. The tumours
treated with a single intratumoural injection of AAV-PEDF began to
grow more slowly than the other two groups on Day 6 after treatment
with maximum tumour growth inhibition observed on Day 15 after
treatment (56% and 58% inhibition, respectively, compared with
AAV-EGFP- or NS-treated mice). Tumour growth suppression was
related to decreased microvessel density and increased apoptosis in
AAV-PEDF-treated tumours. A 73% decreased MVD in the
AAV-PEDF-treated tumours (Fig. 3A and
B) closely paralleled the 58% reduction in tumour size,
implying a direct relationship between lung carcinoma vascularity
and growth. The antiangiogenic activity of PEDF was further
demonstrated by tube formation and the alginate-encapsulated tumour
cell assay (Fig. 3C-F). The average
percentage of apoptosis in the AAV-PEDF group (10%) was
significantly increased, compared with the AAV-EGFP (3%) and NS
(3%) groups (Fig. 4B; P<0.05).
These observations suggest that hindering the vascular supply to a
tumour significantly curbs its ability to grow and that restriction
of tumour angiogenesis caused an increase in tumour cell apoptosis
in AAV-PEDF treated mice.
The mechanisms by which PEDF reduces
neovascularisation are still largely unknown. Nevertheless, 2 major
pathways have been implicated, namely, endothelial cell apoptosis
via activation of the Fas/Fas-L death pathway (28) and disruption of the crucial balance
between pro- and anti-angiogenic factors, in particular
downregulation of VEGF expression (29–31).
Whereas pro-angiogenic factors stimulate endothelial cells that
express Fas, PEDF increases FasL expression on the endothelial cell
surface. A caspase-dependent apoptotic cascade is subsequently
initiated, resulting in endothelial cell death. Another mechanism
underlying the antiangiogenic property of PEDF is that it also has
an inhibitory effect on VEGF-induced angiogenesis. A study by Cai
et al reported that PEDF activates γ-secretase-dependent
cleavage of the VEGF-R1 C terminus, which consequently modulates
angiogenic signalling via VEGF-R2 (31). Exogenous PEDF downregulated VEGF
expression at both the mRNA and protein levels in the MG63
osteosarcoma cell line (26).
In this present study, our data indicated that PEDF
not only acts to halt angiogenesis, but also has the ability to
increase apoptosis in tumours. However, the mechanisms of
PEDF-mediated apoptosis in tumours are still poorly understood. The
apoptotic activity is likely due to a distinct functional epitope
on the PEDF protein that was discovered by Filleur et al,
who reported increases in prostate cancer cell death in
vitro with the 34-mer peptide, but not with the 44-mer
(20). Significant tumour cell
apoptosis with full-length PEDF treatment in vitro has been
observes in melanoma and osteosarcoma cells (16,26).
This effect may be mediated by the Fas/Fas-L cascade similar to
endothelial cell apoptosis, as it was reversed by neutralizing
antibodies against Fas-L (16,26).
AAV has several excellent advantages making it a
particularly promising vector for gene therapy. AAV has long-term
transgene expression in experimental gene therapy methods compared
with other viral vectors (37).
More importantly, AAV is a non-pathogenic virus and has low
immunogenicity because the genes encoding the wild-type protein are
absent, and it has a replication-defective nature (32–34).
In our study, we did not detect AAV-induced toxicity as measured by
calculating the change in animal body weight, observing basic organ
structure and histological morphology, and analyzing mouse liver
and kidney function (Figs. 5 and
6). Several recent reports have
demonstrated the therapeutic efficiency of PEDF viral vector
intratumoural injections (20,38,39).
Intratumoural gene therapy allows for selective expression within
tumour cells contributing to higher intratumoural concentrations
while maintaining normal systemic levels, thus limiting the number
of adverse reactions. AAV vectors in gene therapy could be
relatively safe and efficient.
In conclusion, our results indicate for the first
time that AAV-PEDF by intratumoural injection can provide an
effective protocol for LLC treatment in a C57BL/6 mouse model.
Mechanisms of PEDF-mediated reduction of LLC tumour growth include
both inhibition of tumour angiogenesis and stimulation of tumour
cell apoptosis.
Acknowledgements
This study was funded by the National Key Basic
Research Program (973 Program) of China (2010CB529900).
Abbreviations:
|
PEDF
|
pigment epithelium-derived factor
|
|
AAV
|
adeno-associated virus
|
|
MVD
|
microvessel density
|
|
LLC
|
Lewis lung carcinoma
|
|
VEGF
|
vascular endothelial growth factor
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
NS
|
normal saline
|
|
VEGF-R1
|
vascular endothelial growth
factor-receptor 1
|
|
VEGF-R2
|
vascular endothelial growth
factor-receptor 2
|
References
|
1
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar
|
|
2
|
Folkman J and Ingber D: Inhibition of
angiogenesis. Semin Cancer Biol. 3:89–96. 1992.
|
|
3
|
Bicknell R and Harris AL: Anticancer
strategies involving the vasculature: vascular targeting and the
inhibition of angiogenesis. Semin Cancer Biol. 3:399–407.
1992.PubMed/NCBI
|
|
4
|
Augustin HG: Translating angiogenesis
research into the clinic: the challenges ahead. Br J Radiol.
76:3–10. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shimizu K and Oku N: Cancer
anti-angiogenic therapy. Biol Pharm Bull. 27:599–605. 2004.
View Article : Google Scholar
|
|
6
|
Purow B and Fine HA: Progress report on
the potential of angiogenesis inhibitors for neuro-oncology. Cancer
Invest. 22:577–587. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Conway EM, Collen D and Carmeliet P:
Molecular mechanisms of blood vessel growth. Cardiovasc Res.
49:507–521. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yancopoulos GD, Davis S, Gale NW, Rudge
JS, Wiegand SJ and Holash J: Vascular-specific growth factors and
blood vessel formation. Nature. 407:242–248. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tombran-Tink J, Chader GG and Johnson LV:
PEDF: A pigment epithelium derived factor with potent neuronal
differentiative activity. Exp Eye Res. 53:11–14. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Becerra SP, Sagasti A, Spinella P, et al:
Pigment epithelium-derived factor behaves like a non-inhibitory
serpin. Nerothrophic activity does not require the serpin reactive
loop. J Biol Chem. 270:25992–25999. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Filleur S, Nelius T, de Riese W and
Kennedy RC: Characterization of PEDF: a multi-functional serpin
family protein. J Cell Biochem. 106:769–775. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tombran-Tink J and Johnson LV: Neuronal
differentiation of retinoblastoma cells induced by medium
conditioned by human RPE cells. Invest Ophthalmol Vis Sci.
30:1700–1707. 1989.PubMed/NCBI
|
|
13
|
Bouck N: PEDF: anti-angiogenic guardian of
ocular function. Trends Mol Med. 8:330–334. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dawson DW, Volpert OV, Gillis P, et al:
Pigment epithelium-derived factor: a potent inhibitor of
angiogenesis. Science. 285:245–248. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Houenou LJ, D’Costa AP, Li L, et al:
Pigment epithelium-derived factor promotes the survival and
differentiation of developing spinal motor neurons. J Comp Neurol.
412:506–514. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Abe R, Shimizu T, Yamagishi S, et al:
Overexpression of pigment epithelium-derived factor decreases
angiogenesis and inhibits the growth of human malignant melanoma
cells in vivo. Am J Pathol. 164:1225–1232. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garcia M, Fernandez Garcia NI, Rivas V, et
al: Inhibition of xenografted human melanoma growth and prevention
of metastasis development by dual antiangiogenic/antitumour
activities of pigment epithelium-derived factor. Cancer Res.
64:5632–5642. 2004. View Article : Google Scholar
|
|
18
|
Yang LP, Cheng P, Peng XC, et al:
Anti-tumour effect of adenovirus-mediated gene transfer of pigment
epithelium-derived factor on mouse B16-F10 melanoma. J Exp Clin
Cancer Res. 28:752009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Doll JA, Stellmach VM, Bouck NP, et al:
Pigment epithelium-derived factor regulates the vasculature and
mass of the prostate and pancreas. Nat Med. 9:774–780. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Filleur S, Volz K, Nelius T, et al: Two
functional epitopes of pigment epithelial-derived factor block
angiogenesis and induce differentiation in prostate cancer. Cancer
Res. 65:5144–5152. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guan M, Jiang H, Xu C, Xu R, Chen Z and Lu
Y: Adenovirus-mediated PEDF expression inhibits prostate cancer
cell growth and results in augmented expression of PAI-2. Cancer
Biol Ther. 6:419–425. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kojiro M, Hiroki I, Daisuke N, Keisuke H,
Kazuhiko N and Katsumi E: Antiangiogenic property of pigment
epithelium-derived factor in hepatocellular carcinoma. Hepatology.
40:252–259. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Uehara H, Miyamoto M, Kato K, et al:
Expression of pigment epithelium-derived factor decreases liver
metastasis and correlates with favorable prognosis for patients
with ductal pancreatic adenocarcinoma. Cancer Res. 64:3533–3537.
2004. View Article : Google Scholar
|
|
24
|
Zhang L, Chen J, Ke Y, Mansel RE and Jiang
WG: Down-regulation of PEDF expression by ribozyme transgene in
endothelial and lung cancer cells and its impact on angiogenesis
in vitro. Oncol Rep. 14:1615–1619. 2005.PubMed/NCBI
|
|
25
|
Ek ET, Dass CR, Contreras KG and Choong
PF: PEDF-derived synthetic peptides exhibit antitumour activity in
an orthotopic model of human osteosarcoma. J Orthop Res.
25:1671–1680. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Takenaka K, Yamagishi S, Jinnouchi Y,
Nakamura K, Matsui T and Imaizumi T: Pigment epithelium-derived
factor (PEDF)-induced apoptosis and inhibition of vascular
endothelial growth factor (VEGF) expression in MG63 human
osteosarcoma cells. Life Sci. 77:3231–3241. 2005. View Article : Google Scholar
|
|
27
|
Zhang T, Guan M, Xu C, Chen Y and Lu Y:
Pigment epithelium-derived factor inhibits glioma cell growth in
vitro and in vivo. Life Sci. 81:1256–1263. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Volpert OV, Zaichuk T, Zhou W, et al:
Inducer-stimulated Fas targets activated endothelium for
destruction by anti-angiogenic thrombospondin-1 and pigment
epithelium-derived factor. Nat Med. 8:349–357. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamagishi S, Amano S, Inagaki Y, et al:
Pigment epithelium-derived factor inhibits leptin-induced
angiogenesis by suppressing vascular endothelial growth factor gene
expression through anti-oxidative properties. Microvasc Res.
65:186–190. 2003. View Article : Google Scholar
|
|
30
|
Yamagishi S, Inagaki Y, Amano S, et al:
Pigment epithelium-derived factor protects cultured retinal
pericytes from advanced glycation end product-induced injury
through its antioxidative properties. Biochem Biophys Res Commun.
296:877–882. 2002. View Article : Google Scholar
|
|
31
|
Cai J, Jiang WG, Grant MB and Boulton M:
Pigment epithelium-derived factor inhibits angiogenesis via
regulated intracellular proteolysis of vascular endothelial growth
fact receptor 1. J Biol Chem. 281:3604–3613. 2006. View Article : Google Scholar
|
|
32
|
Nathwani AC, Davidoff A, Hanawa H, et al:
Factors influencing in vivo transduction by recombinant
adeno-associated viral vectors expressing the human factor IX cDNA.
Blood. 97:1258–1265. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Z, Zeng G, Zhang L, Xu C and Guo Y:
Extracellular domain of kinase domain region mediated by
adeno-associated virus inhibits growth and angiogenesis of bladder
cancer in Balb-c mice. Chin Med J (Engl). 115:1209–1212.
2002.PubMed/NCBI
|
|
34
|
Carter PJ and Samulski RJ:
Adeno-associated viral vectors as gene delivery vehicles. Int J Mol
Med. 6:17–27. 2000.PubMed/NCBI
|
|
35
|
Wu X, Dong X, Wu Z, Cao H, Niu D, et al: A
novel method for purification of recombinant adenoassociated virus
vectors on a large scale. Chinese Sci Bull. 46:485–488. 2001.
View Article : Google Scholar
|
|
36
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumour angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, Su C, Cao H, et al: A novel
triple-regulated oncolytic adenovirus carrying p53 gene exerts
poten antitumour efficacy on common human solid cancers. Mol Cancer
Ther. 7:1598–1603. 2008. View Article : Google Scholar
|
|
38
|
Hase R, Miyamoto M, Uehara H, et al:
Pigment epithelium-derived factor gene therapy inhibits human
pancreatic cancer in mice. Clin Cancer Res. 11:8737–8744. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mahtabifard A, Merritt RE, Yamada RE,
Crystal RG and Korst RJ: In vivo gene transfer of pigment
epithelium-derived factor inhibits tumour growth in syngeneic
murine models of thoracic malignancies. J Thorac Cardiovasc Surg.
126:28–38. 2003. View Article : Google Scholar : PubMed/NCBI
|