Introduction
Hepatocellular carcinoma (HCC) is one of the most
common fatal malignancies observed worldwide, with an annual
incidence of ~600,000 deaths (1).
Chronic infection with hepatitis B virus (HBV) or hepatitis C virus
(HCV) is the most clearly established risk factor for HCC (2). In Japan, ~80% of HCC cases are due to
chronic HCV infection, while the vast majority of the remaining
cases are associated with HBV infection (3,4).
Despite recent advances in patient management, the 5-year survival
rate of HCC patients is as low as 26–50%, while disease-free
survival is only 13–29% after curative surgery (5). This poor prognosis can be largely
explained by early intrahepatic recurrence (IHR) due to metastatic
spread of cancer cells via portal vein invasion (PVI) following
surgery (6–8). Given this poor prognosis, there has
been intense interest in identifying a strategy that may either
prevent PVI, or effectively treat HCC with PVI. For this purpose,
it is crucial to identify the key genes that play a central role in
the development of PVI of HCC. With this in mind, we have pooled
DNA microarray data of >10,000 genes in 60 primary sites of HCC
(7,9). By collecting and analyzing these data,
we were able to identify RD RNA binding protein (RDBP) as a unique
candidate gene responsible for PVI.
The RDBP gene localizes to the class III
region of the major histocompatibility complex (MHC) on chromosome
6, and encodes a 44-kDa nuclear protein (10,11).
The encoded protein is a member of the negative elongation factor
(NELF) transcription elongation regulatory complex that represses
RNA polymerase II transcript elongation by acting with DRB
(5,6-dichloro-1-β-D-ribofuranosylbenzimidazole)
sensitivity-inducing factor (DSIF), resulting in the
transcriptional pause of RNA polymerase II (12). Deletion of the RNA-recognition motif
(RRM) has demonstrated that this region plays the most critical
role in the transcriptional pausing among the RDBP motifs (13,14).
However, to the best of our knowledge, studies examining the
biological role of RDBP in human cancer cells have yet to be
undertaken.
In the present study, we demonstrated for the first
time that increased RDBP levels were associated with PVI and early
IHR of HCV-related HCC, and that RDBP may serve as a therapeutic
target for HCC with a highly malignant phenotype.
Patients and methods
Patients and tissue samples
HCV-related HCC and corresponding non-cancer liver
tissues were obtained from patients who had undergone surgical
resection in the Department of Digestive Surgery and Surgical
Oncology, Yamaguchi University Medical Hospital between 1997 and
2007. For reverse transcription polymerase chain reaction (RT-PCR)
analysis, tumor tissue samples and their corresponding non-tumor
liver tissues were collected from 57 patients with HCC who
underwent curative hepatectomy. For immunohistochemical analysis,
88 non-HCC liver and HCC tissues were collected, fixed in 10%
formaldehyde solution and embedded in paraffin. Sixty-four (73%) of
the 88-patient samples collected for this portion of the study had
undergone curative hepatectomy. In this study, we defined IHR
within 2 years of surgery as early IHR, as has been described
previously (15). All samples were
obtained with the patients' informed consent. The study protocol
was undertaken according to the REMARK criteria (http://www.cancerdiagnosis.nci.nih.gov/assessment/progress/remark.htm.),
and was approved by the Institutional Review Board for the Use of
Human Subjects at Yamaguchi University School of Medicine.
Cell culture
The human HCC cell line HLE was used for all
functional analyses in the present study. HLE cells were maintained
in DMEM containing 10% fetal bovine serum (FBS) and antibiotics at
37°C in a 5% humidified CO2 atmosphere.
RNA extraction and RT-PCR
In 57 sample sets that were independent of the
sample sets assigned to the DNA microarray experiment, we evaluated
the reproducibility of the relationship between RDBP mRNA
expression levels with PVI of HCC using semi-quantitative real-time
RT-PCR. For this analysis, total RNA was extracted from 57 paired
samples of frozen HCC tissue and adjacent hepatic tissue using the
TRIzol method (Gibco, Carlsbad, CA, USA) according to the
manufacturer's instructions. Semi-quantitative real-time RT-PCR was
undertaken as described previously (16) with minor modifications. We measured
mRNA levels semi-quantitatively using the D/D threshold cycle
method. In addition, arginine/serine-rich splicing factor 4 (SFRS4)
(17) and glyceraldehyde phosphate
dehydrogenase (GAPDH) were used as the reference genes. cDNA
solution corresponding to 10 ng of the initial RNA was used for PCR
amplification steps that were designed using the Roche Universal
Probe Library (https://www.roche-applied-science.com/sis/rtpcr/upl/index.jsp).
PCR primers used were sense (5′-gcagaagaaattcaacaagctca-3′) and
antisense (5′-tgtgctgctgctactttgct-3′) for the RDBP gene
(NM_002904), sense (5′-gatggcagttacggttctgg-3′) and antisense
(5′-gccatatttatctcggccact-3′) for the SFRS4 gene (NM_005626) and
sense (5′-agccacatcgctcagacac-3′) and antisense
(5′-gcccaatacgaccaaatcc-3′) for the GAPDH gene (NM_002046). The
size of the PCR products for RDBP, SFRS4 and GAPDH were 76, 66 and
66 bp, respectively. The Universal Probe Library probes nos. 21, 86
and 60 (Roche Diagnostics GmbH, Mannheim, Germany) were used for
measurement of RDBP, SFRS4 and GAPDH levels, respectively.
Immunohistochemical staining
Immunohistochemical staining for RDBP was performed
on formalin-fixed, paraffin-embedded tissue sections using the
Envision+ system (Dako, Glostrup, Denmark) following the
manufacturer's instructions. For antigen retrieval, slides were
boiled in 0.01 M sodium citrate buffer (pH 6.0) for 20 min in a
microwave oven. After blocking with 3% hydrogen peroxide
(H2O2), the slides were incubated with rabbit
polyclonal antibody 10705-AP (Proteintech Group, Chicago, IL, USA)
diluted 1:80 at 4°C overnight. The slides were then washed in
buffer, incubated with biotinylated secondary antisera, and
streptavidin-biotin complex/horseradish peroxidase applied.
Finally, we calculated the percentage of positive cells in each
lesion using a Katikati counter (http://www.vector.co.jp/soft/dl/win95/art/se347447.html).
We scored the staining intensity as follows; 0, no staining; 1+,
mild staining; 2+, moderate staining; 3+, intense staining. The
area of staining was evaluated as follows; 0, no staining of cells
in any microscopic fields; 1+, <30% of nucleus stained positive;
2+, between 30 and 60% stained positive; 3+, >60% stained
positive. RDBP expression was evalueded by combined assessing of
staining intensity and extension. The criteria used in this study
has been widely accepted previously (18). HCC samples were categorized into
weak and strong expression groups according to RDBP expression
score in the nucleus. HCC with RDBP expression score equal to or
<4 and those with RDBP positivity of >4 were categorized into
weak and strong expression groups, respectively.
Western blot analysis
Total protein was extracted from cell lines using
the protein extraction solution M-PER® Mammalian Protein
Extraction Reagent (Thermo Fisher Scientific, Rockford, IL, USA).
Aliquots of total protein (50 μg for clinical samples and 30 μg for
cell lines) were electrophoresed on 10–20% gradient precast gels
(System Instruments Co., Tokyo, Japan) and electroblotted onto pure
nitrocellulose membranes [iBlot Transfer Stack, Mini
(Nitrocellulose), Invitrogen, Carlsbad, CA USA].
RDBP protein was then detected using rabbit
polyclonal antibody 10705-AP (Proteintech Group) diluted 1:500.
RDBP protein levels were normalized to the level of GAPDH (SC
25778, Santa Cruz Biotechnology, Santa Cruz, USA). Blots were
developed with horseradish peroxidase-linked anti-rabbit
immunoglobulin SC 2004 (Santa Cruz Biotechnology) diluted 1:7500.
Supersignal West Pico Trial Kit Reagents (Thermo Fisher Scientific)
were used to detect antigen-antibody reactions.
siRNA transfection
Small interfering RNA (siRNA) for RDBP was custom
synthesized by Qiagen, (Qiagen, Hilden, Germany). Target sequences
were as follows: si RDBP-1 5′-CGGGATCGG GATCGAGATCGA-3′ and si
RDBP-2 5′-CAAGGTGGTGTCA AACGCTCA-3′. Non-specific control siRNA was
obtained from B-Bridge International (S10C-0600, Cupertino, CA).
SiRNA to RDBP1,2 or non-specific siRNA (both 100 pmol/ml) was
transfected using Lipofectamine 2000 reagent (Invitrogen) in
serum-free Opti-MEM I (Invitrogen) according to the manufacturer's
instructions and as described previously (16).
Proliferation assay
To evaluate cell survival and proliferation,
CellTiter 96RAQueous One Solution Cell Proliferation Assay (MTS
assay, Promega, Madison, WI, USA) was performed. Cells
(2×104) were seeded into the wells of 96-well plates
after the transfection of siRNA and incubated at 37°C in a
humidified atmosphere with 5% CO2. At the appropriate
time, 20 μl of CellTiter 96RAQueous one solution was added to each
well and the plates incubated for a further 4 h at 37°C. The
optical density was then measured at 490 nm using a 96-well plate
reader. Triplicate wells were analyzed in each assay.
Invasion assay
HLE cells transfected either with RDBP siRNA or
non-specific control siRNA were cultured in DMEM containing 5% FBS.
The cells were then harvested by trypsinization, washed in DMEM
without serum and suspended in DMEM at 4×104 cells/ml.
Prior to the preparation of the cell suspension, a dried layer of
Matrigel matrix (Becton-Dickinson Biosciences, San Jose, CA, USA)
was rehydrated with DMEM for 2 h at room temperature. DMEM (0.75
ml) containing 2% FBS was then added to the lower chambers of a
24-well Matrigel invasion chamber, and 0.5 ml (2×104
cells) of cell suspension added to each insert of the upper
chamber. The plates and inserts were incubated for 24 h at 37°C.
After incubation, the cells invading through the Matrigel were
fixed and stained with hematoxylin. As a control, uncoated
polycarbonate membrane (Becton-Dickinson) was used instead of the
Matrigel chamber. The number of cells in each membrane was also
counted under a microscope (magnification ×50) using a Katikati
counter. Triplicate wells were analyzed for each assay.
Statistical analysis
Student's t-test and Mann-Whitney U test were used
to evaluate differences between two or more continuous variables.
Fisher's exact test or χ2 test was used to evaluate the
differences between discontinuous variables. We carried out
multivariate analysis to assess independent factors for early IHR
using the multiple logistic regression models. All statistical
analyses were performed using SPSS 11.0J (SPSS, Inc., Chicago, IL,
USA) software. P<0.05 was considered significant.
Results
Identification of RDBP as a candidate
gene procedure
Using pooled DNA array data, we identified 40 genes
demonstrating expression levels that were higher by >4-fold in
HCC tissues when compared with non-cancer liver tissues. We next
filtered 18 genes for which expression levels were significantly
higher in HCC with PVI than in HCC without PVI. We subsequently
ranked the 18 genes according to the magnitude of their mRNA levels
and then examined immunohistochemically their protein levels in
several sample sets of HCC and corresponding non-HCC liver tissues.
Our preliminary examination demonstrated that RDBP expression
levels were abundant in HCC tissues and were significantly higher
in HCC with PVI than in HCC without PVI.
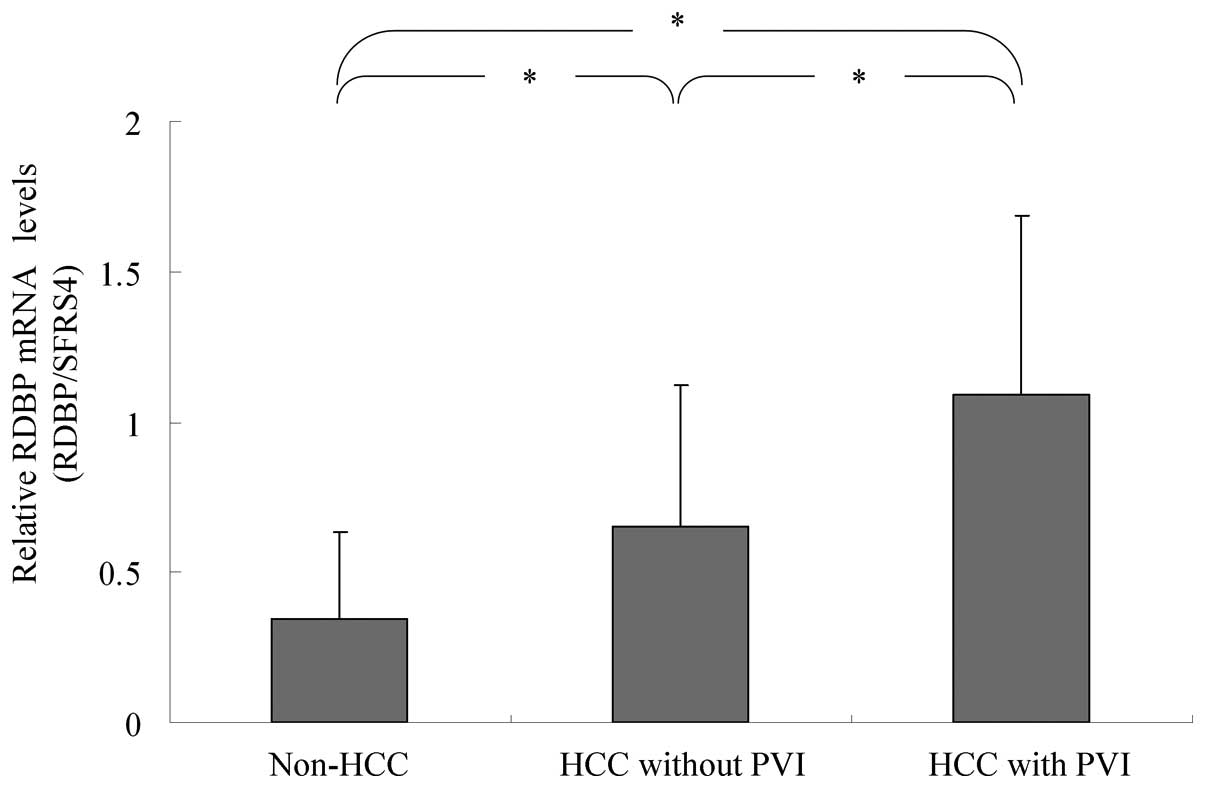
RDBP mRNA expression
RDBP mRNA expression was significantly higher in HCC
without PVI (n=39) than in non-HCC liver samples (n=57, 0.65±0.47
vs. 0.34±0.29 (mean ± SD), P<0.01). RDBP mRNA levels were also
significantly higher in HCC with PVI (n=18) than in HCC without PVI
(n=39, 1.09±0.60 vs. 0.65±0.47, P<0.01, Fig. 1).
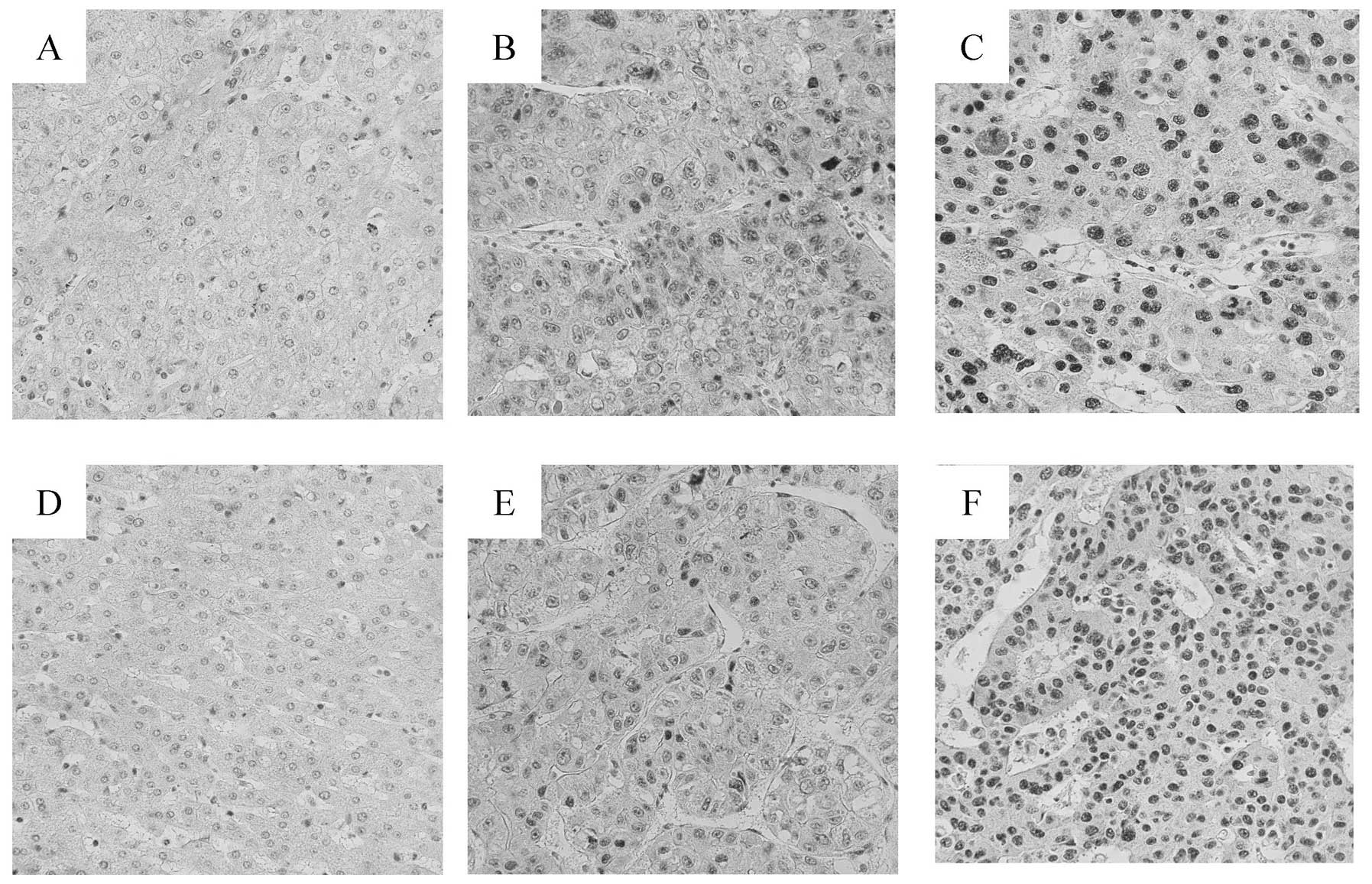
RDBP protein expression
To evaluate whether RDBP protein levels were
associated with the metastatic potential of HCC, we performed
immunohistochemical staining for RDBP on 88 sample sets of HCC and
corresponding non-HCC liver tissues, and 10 normal liver tissues in
which normal liver function was confirmed by a blood test. RDBP
protein was predominantly expressed in the nucleus of HCC cells
(Fig. 2). RDBP protein was
expressed in the majority of HCC tissues (84/88, 95%). In contrast,
staining for RDBP protein was observed in only 20% of the non-HCC
liver tissues (Fig. 2A and D).
According to the protein expression score, we determined that
staining for RDBP was weak in 33 (37.5%) and strong in 51 (58.0%)
of the HCC tissues. Staining for RDBP was weak in 16 (18.2%) and
strong in 2 (2.3%) of the non-HCC liver tissues. Thus, RDBP protein
levels were found to be significantly higher in HCC tissues than in
non-HCC liver tissues (P<0.0001 by Fisher's exact test). No
staining for RDBP protein was observed in the 10 control liver
tissues (Fig. 2 and Table I).
 | Table IRDBP protein expression in human liver
samples. |
Table I
RDBP protein expression in human liver
samples.
| HCC patient | | |
|---|
|
| | |
|---|
| RDBP protein
expression | HCC (n=88) | Non-tumor (n=88) | Normal (n=10) | P-value |
|---|
| Negative | 4 (4.5%) | 70 (79.5%) | 10 | <0.01 |
| Weak | 33 (37.5%) | 16 (18.2%) | 0 | |
| Strong | 51 (58.0%) | 2 (2.3%) | 0 | |
Correlation of RDBP expression with
clinicopathological features
RDBP protein level was positively associated with
PVI and de-differentiation grade (P<0.05 for both, Table II). In contrast, RDBP protein level
was not associated with other factors such as age, gender, tumor
size and pathological TNM classification of malignant tumors (TNM)
stages.
| Table IIClinicopathologic findings of HCC in
relation to RDBP expression. |
Table II
Clinicopathologic findings of HCC in
relation to RDBP expression.
| RDBP
expression | |
|---|
|
| |
|---|
| Clinicopathologic
factor | High (N=51) | Low (N=37) | P-value |
|---|
| Age | 68.0±8.4 | 67.8±7.2 | 0.90 |
| Gender | | | 0.44 |
| Male | 38 | 29 | |
| Female | 13 | 8 | |
| Tumor size | | | 0.08 |
| <3 cm | 24 | 11 | |
| ≥3 cm | 27 | 26 | |
| Degree of
differentiation | | | <0.01 |
| Well | 5 | 12 | |
| Moderate | 35 | 24 | |
| Poor | 11 | 1 | |
| Portal vein
invasion (PVI) | | | <0.01 |
| Negative | 31 | 33 | |
| Positive | 20 | 4 | |
| TNM stage | | | 0.28 |
| I+II | 21 | 19 | |
| III+IV | 30 | 18 | |
| Early intrahepatic
recurrence | High (N=40) | Low (N=24) | 0.02 |
| Negative | 15 | 16 | |
| Positive | 25 | 8 | |
RDBP overexpression as an import
predictive marker for early intrahepatic recurrence
RDBP protein levels were significantly higher in HCC
with early IHR (n=33) than in HCC without early IHR (n=31, Table II). To identify independent risk
factors for early IHR, 5 variables including primary tumor number,
tumor size, tumor differentiation, portal invasion, and RDBP
expression were entered into a multivariate regression analysis.
The logistic regression analysis selected only one variable, RDBP
expression group (P=0.026) as risk factors for early IHR (Table III).
| Table IIIIndependent risk factors for early
intrahepatic recurrence (IHR). |
Table III
Independent risk factors for early
intrahepatic recurrence (IHR).
| Factors | Regression
coeffcient | Standard error | Risk ratio (95%
CI) | P-value |
|---|
| RDBP group | 4.928 | 0.542 | 3.333
(1.151–9.650) | 0.026 |
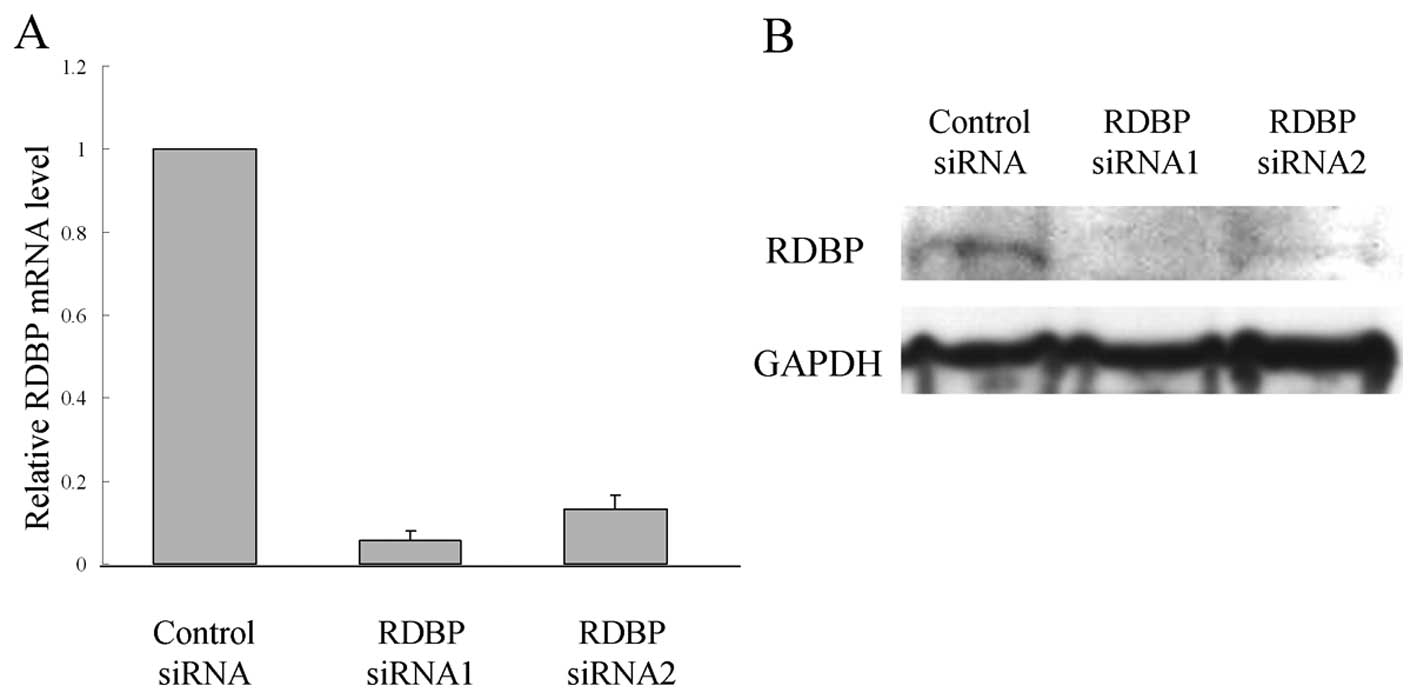
RDBP function in cell proliferation
To assess whether RDBP is essential for growth or
survival of HCC cells, we used siRNA technology to reduce RDBP
levels. When we transfected siRNAs against RDBP (RDBP si-RNA1 and
si-RNA2) into HLE cells, we found that HLE-siRNA1 and HLE-siRNA2
cells showed markedly lower levels at RDBP mRNA and protein levels
than the si-control cells (Fig. 3A and
B). Down-regulation of RDBP expression by siRNA also led to a
significant growth inhibition and a significant reduction in the
cellular activity of HLE cells when compared with the si-control
cells (Fig. 3C and D).
RDBP function in cell invasion
Cell invasion was investigated using the Matrigel
cell invasion chamber. Down-regulation of RDBP expression by siRNA
led to a decrease in the number of invasive cells when compared
with the si-control cells (Fig.
3E), thus independently suggesting that RDBP plays an important
role in the invasive ability of HCC cells.
Discussion
It is well established that vessel invasion (VI)
including PVI, tumor number and tumor size are representative risk
factors of a poor prognosis for HCC patients undergoing hepatic
surgery (19). Among these factors,
PVI is considered to be a hallmark of the intrahepatic spread of
HCC cells and of poor outcome (20). It is therefore crucial for
hepatologists to indentify key genes or proteins that play a
central role in the PVI process of HCC. To this end, the present
study focused on PVI-related genes and successfully identified a
novel PVI-related gene, RDBP, in HCC from thousands of genes on the
DNA array without any bias. We found that mRNA levels of RDBP were
significantly higher in HCV-related HCC with PVI than in those
without PVI, and that the encoded protein was increased in parallel
to the progression of poorly differentiated HCC. Strikingly, the
protein level of RDBP was an independent risk factor for early IHR
within 2 years of surgery.
Several DNA microarray studies have identified
VI-related genes in a manner similar to our study. Chen et
al(21) identified 91 genes for
which expression levels were significantly correlated with the
presence or absence of VI. Okabe et al(22) identified 151 VI-related genes
including 110 ESTs, RHOC and two small GTPase-related genes
known as ARHGAP8 and ARHGEF6. A study by Tsunedomi
et al(23) focused on
moderately differentiated HCV-related HCC to minimize the bias of
gene selection, as VI is not detected in well-differentiated HCC,
but is frequently observed in moderately or less
well-differentiated HCC, and identified 35 VI-related genes. Tanaka
et al(24) identified 28
VI-related genes including AURKB using microarray clustering based
on macroscopic findings for VI. Thus, although much effort has been
devoted to identification of VI-related genes, there are, so far,
few genes or gene products that can be applied to the daily
clinical use of HCC treatment. One possible deficiency may be
explained by the fact that gene levels are not always related to
those of the encoded protein. In this regard, RDBP identified in
the present study is fascinating from the viewpoint of a target
molecule specific for HCC as its protein is abundant in HCC, but
not in non-HCC liver tissues and in various non-malignant epithelia
(data not shown).
It is generally accepted that HCC recurrence is a
complicated process. There are at least three modes of
postoperative recurrence (25)
including early and late IHR that appear in the remnant liver, and
extra-hepatic recurrence that appears in distant tissues and
organs. In this study, we found that RDBP protein level was
positively associated with early IHR, but not late IHR.
Additionally, our multivariate analysis revealed that RDBP was an
independent risk factor for early IHR. These results appear to be
reasonable, considering that late IHR is caused by de novo
hepatocarcinogenesis such as multicentric occurrence (25), which can be affected by the
background liver status in chronic liver disease, but not tumor
factors. In contrast, most early IHRs are due to intrahepatic
metastasis of cancer cells and are detected in 30–50% of patients
within two years of surgery, limiting the potential for surgical
cure of HCC (7,26,27).
Taken together, we strongly suggest that overexpression of RDBP may
account for the highly metastatic potential of HCC.
In the present study, our experimental finding
showed that knockdown of RDBP inhibited the invasive potential of
HCC cells, supporting our clinical finding that RDBP was
overexpressed in HCC with PVI. However, the precise role of RDBP in
the metastatic process of cancer cells remains unclear. An elegant
study by Narita et al(28)
showed that although knockdown of RDBP/NELF-E inhibited the
proliferation of HeLa cells, there were no significant difference
in the cell cycle distribution between the control and knockdown
cells. Consistent with this, we confirmed both the growth
inhibitory effects and the lack of change in cell cycle pattern in
HCC cells by RDBP knockdown (data not shown). Narita et
al(28) revealed that knockdown
of RDBP/NELF-E promoted the transcription of replication-dependent
histone genes. It remains unclear as to how histone protein family
is linked to invasion potentials of cancer cells. Therefore,
confirmation of this finding is beyond the scope of the present
study. Intriguingly, recent studies showed that COBRA1/NELF-B and
TH1/NELF-C/D were decreased in advanced breast cancer and that
knockdown of the two genes enhanced motility of the breast cancer
cells (29,30). Thus, NELF family proteins may have
opposite effects in cancer progression in a tissue-dependent
manner. Further examination is required to gain insight into this
concept.
The precise mechanisms underlying the up-regulation
of RDBP at a transcriptional level in HCC with PVI remain unknown.
One possible explanation is that this gene may be epigenetically
regulated via CpG islands at the promoter region. To test this
possibility, we undertook a preliminary examination to investigate
the epigenetic status of the promoter region of the RDBP
gene in four paired sets of HCC and corresponding non-HCC liver
tissue using methylation-specific PCR (MSP). Our MSP analysis
revealed that the RDBP gene remained unmethylated in all of
the samples tested (data not shown), raising the possibility that
RDBP was up-regulated by mechanisms other than promoter
methylation.
More recently, enhancement of HCC patient prognosis
using specific targeted agents such as Sorafenib has been reported
(31,32). Although Sorafenib inhibits
proliferation and angiogenesis in HCC, it is unclear whether it
directly inhibits tumor invasion and metastasis. In this regard,
and given the finding that RDBP is closely associated with the
proliferation and invasive potentials of HCC cells, we suggest that
RDBP may prove useful as a potential therapeutic target for HCC,
especially in advanced HCC with PVI.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
2
|
Thorgeirsson SS and Grisham JW: Molecular
pathogenesis of human hepatocellular carcinoma. Nat Genet.
31:339–346. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ohishi W, Kitamoto M, Aikata H, Kamada K,
Kawakami Y, Ishihara H, Kamiyasu M, Nakanishi T, Tazuma S and
Chayama K: Impact of aging on the development of hepatocellular
carcinoma in patients with hepatitis C virus infection in Japan.
Scand J Gastroenterol. 38:894–900. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ikai I, Arii S, Ichida T, Okita K, Omata
M, Kojiro M, Takayasu K, Nakanuma Y, Makuuchi M, Matsuyama Y and
Yamaoka Y: Report of the 16th follow-up survey of primary liver
cancer. Hepatol Res. 32:163–172. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thomas MB and Zhu AX: Hepatocellular
carcinoma: the need for progress. J Clin Oncol. 23:2892–2899. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shirabe K, Kanematsu T, Matsumata T,
Adachi E, Akazawa K and Sugimachi K: Factors linked to early
recurrence of small hepatocellular carcinoma after hepatectomy:
univariate and multivariate analyses. Hepatology. 14:802–805. 1991.
View Article : Google Scholar
|
|
7
|
Iizuka N, Oka M, Yamada-Okabe H, Nishida
M, Maeda Y, Mori N, Takao T, Tamesa T, Tangoku A, Tabuchi H, Hamada
K, Nakayama H, Ishitsuka H, Miyamoto T, Hirabayashi A, Uchimura S
and Hamamoto Y: Oligonucleotide microarray for prediction of early
intrahepatic recurrence of hepatocellular carcinoma after curative
resection. Lancet. 361:923–929. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shirabe K, Wakiyama S, Gion T, Motomura K,
Koyanagi T, Sakamoto S and Nagaie T: Clinicopathological risk
factors linked to recurrence pattern after curative hepatic
resection for hepatocellular carcinoma - results of 152 resected
cases. Hepatogastroenterology. 54:2084–2087. 2007.
|
|
9
|
Iizuka N, Oka M, Yamada-Okabe H, Mori N,
Tamesa T, Okada T, Takemoto N, Sakamoto K, Hamada K, Ishitsuka H,
Miyamoto T, Uchimura S and Hamamoto Y: Self-organizing-map-based
molecular signature representing the development of hepatocellular
carcinoma. FEBS Lett. 579:1089–1100. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Surowy CS, Hoganson G, Gosink J, Strunk K
and Spritz RA: The human RD protein is closely related to nuclear
RNA-binding proteins and has been highly conserved. Gene.
90:299–302. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheng J, Macon KJ and Volanakis JE: cDNA
cloning and characterization of the protein encoded by RD, a gene
located in the class III region of the human major
histocompatibility complex. Biochem J. 294:589–593. 1993.
|
|
12
|
Yamaguchi Y, Takagi T, Wada T, Yano K,
Furuya A, Sugimoto S, Hasegawa J and Handa H: NELF, a multisubunit
complex containing RD, cooperates with DSIF to repress RNA
polymerase II elongation. Cell. 97:41–51. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yamaguchi Y, Inukai N, Narita T, Wada T
and Handa H: Evidence that negative elongation factor represses
transcription elongation through binding to a DRB
sensitivity-inducing factor/RNA polymerase II complex and RNA. Mol
Cell Biol. 22:2918–2927. 2002. View Article : Google Scholar
|
|
14
|
Narita T, Yamaguchi Y, Yano K, Sugimoto S,
Chanarat S, Wada T, Kim DK, Hasegawa J, Omori M, Inukai N, Endoh M,
Yamada T and Handa H: Human transcription elongation factor NELF:
identification of novel subunits and reconstitution of the
functionally active complex. Mol Cell Biol. 23:1863–1873. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sakon M, Umeshita K, Nagano H, Eguchi H,
Kishimoto S, Miyamoto A, Ohshima S, Dono K, Nakamori S, Gotoh M and
Monden M: Clinical significance of hepatic resection in
hepatocellular carcinoma: analysis by diseasefree survival curves.
Arch Surg. 135:1456–1459. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsunedomi R, Iizuka N, Tamesa T, Sakamoto
K, Hamaguchi T, Somura H, Yamada M and Oka M: Decreased ID2
promotes metastatic potentials of hepatocellular carcinoma by
altering secretion of vascular endothelial growth factor. Clin
Cancer Res. 14:1025–1031. 2008. View Article : Google Scholar
|
|
17
|
Waxman S and Wurmbach E: De-regulation of
common housekeeping genes in hepatocellular carcinoma. BMC
Genomics. 8:2432007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng AL, Huang WG, Chen ZC, Peng F, Zhang
PF, Li MY, Li F, Li JL, Li C, Yi H, Yi B and Xiao ZQ:
Identification of novel nasopharyngeal carcinoma biomarkers by
laser capture microdissection and proteomic analysis. Clin Cancer
Res. 14:435–445. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Poon RT, Ng IO, Fan ST, Lai EC, Lo CM, Liu
CL and Wong J: Clinicopathologic features of long-term survivors
and disease-free survivors after resection of hepatocellular
carcinoma: a study of a prospective cohort. J Clin Oncol.
19:3037–3044. 2001.PubMed/NCBI
|
|
20
|
Vauthey JN, Lauwers GY, Esnaola NF, Do KA,
Belghiti J, Mirza N, Curley SA, Ellis LM, Regimbeau JM, Rashid A,
Cleary KR and Nagorney DM: Simplified staging for hepatocellular
carcinoma. J Clin Oncol. 20:1527–1536. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen X, Cheung ST, So S, Fan ST, Barry C,
Higgins J, Lai KM, Ji J, Dudoit S, Ng IO, van De Rijn M, Botstein D
and Brown PO: Gene expression patterns in human liver cancers. Mol
Biol Cell. 13:1929–1939. 2002. View Article : Google Scholar PubMed/NCBI
|
|
22
|
Okabe H, Satoh S, Kato T, Kitahara O,
Yanagawa R, Yamaoka Y, Tsunoda T, Furukawa Y and Nakamura Y:
Genome-wide analysis of gene expression in human hepatocellular
carcinomas using cDNA microarray: identification of genes involved
in viral carcinogenesis and tumor progression. Cancer Res.
61:2129–2137. 2001.
|
|
23
|
Tsunedomi R, Iizuka N, Yamada-Okabe H,
Tamesa T, Okada T, Sakamoto K, Takashima M, Hamaguchi T, Miyamoto
T, Uchimura S, Hamamoto Y, Yamada M and Oka M: Identification of
ID2 associated with invasion of hepatitis C virus-related
hepatocellular carcinoma by gene expression profile. Int J Oncol.
29:1445–1451. 2006.PubMed/NCBI
|
|
24
|
Tanaka S, Mogushi K, Yasen M, Noguchi N,
Kudo A, Nakamura N, Ito K, Miki Y, Inazawa J, Tanaka H and Arii S:
Gene-expression phenotypes for vascular invasiveness of
hepatocellular carcinomas. Surgery. 147:405–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Iizuka N, Hamamoto Y, Tsunedomi R and Oka
M: Translational microarray systems for outcome prediction of
hepatocellular carcinoma. Cancer Sci. 99:659–665. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Llovet JM, Burroughs A and Bruix J:
Hepatocellular carcinoma. Lancet. 362:1907–1917. 2003. View Article : Google Scholar
|
|
27
|
Matoba K, Iizuka N, Gondo T, Ishihara T,
Yamada-Okabe H, Tamesa T, Takemoto N, Hashimoto K, Sakamoto K,
Miyamoto T, Uchimura S, Hamamoto Y and Oka M: Tumor HLA-DR
expression linked to early intrahepatic recurrence of
hepatocellular carcinoma. Int J Cancer. 115:231–240. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Narita T, Yung TM, Yamamoto J, Tsuboi Y,
Tanabe H, Tanaka K, Yamaguchi Y and Handa H: NELF interacts with
CBC and participates in 3′ end processing of replication-dependent
histone mRNAs. Mol Cell. 26:349–365. 2007.PubMed/NCBI
|
|
29
|
Sun J, Watkins G, Blair AL, Moskaluk C,
Ghosh S, Jiang WG and Li R: Deregulation of cofactor of BRCA1
expression in breast cancer cells. J Cell Biochem. 103:1798–1807.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zou W, Yang Y, Wu Y, Sun L, Chi Y, Wu W,
Yun X, Xie J and Gu J: Negative role of trihydrophobin 1 in breast
cancer growth and migration. Cancer Sci. 101:2156–2162. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Llovet JM, Ricci S, Mazzaferro V, Hilgard
P, Gane E, Blanc JF, de Oliveira AC, Santoro A, Raoul JL, Forner A,
Schwartz M, Porta C, Zeuzem S, Bolondi L, Greten TF, Galle PR,
Seitz JF, Borbath I, Häussinger D, Giannaris T, Shan M, Moscovici
M, Voliotis D and Bruix J: Sorafenib in advanced hepatocellular
carcinoma. N Engl J Med. 359:378–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheng AL, Kang YK, Chen Z, Tsao CJ, Qin S,
Kim JS, Luo R, Feng J, Ye S, Yang TS, Xu J, Sun Y, Liang H, Liu J,
Wang J, Tak WY, Pan H, Burock K, Zou J, Voliotis D and Guan Z:
Efficacy and safety of sorafenib in patients in the Asia-Pacific
region with advanced hepatocellular carcinoma: a phase III
randomised, double-blind, placebo-controlled trial. Lancet Oncol.
10:25–34. 2009. View Article : Google Scholar
|