Introduction
The most important role of 1,25-dihydroxyvitamin
D3(1,25D) is to regulate calcium and phosphate levels by
enhancing their intestinal absorption, renal reabsorption and by
regulating bone mineralization. However, it is well documented that
the actions of 1,25D extend beyond a role in mineral metabolism, as
the compound is also important for cell proliferation (1), cell differentiation (2) and immunomodulation (3). Since differentiation of prostate
cancer, breast cancer and myeloid leukemia cells may have
beneficial therapeutic effects in pathological conditions,
therapeutic applications for 1,25D have been postulated.
Unfortunately, a major limitation for therapeutic use of 1,25D are
its potent calcemic and phosphatemic activities. The doses of
1,25D, which are necessary to induce differentiation and inhibit
cell proliferation in vivo, cause hypercalcemia and
hyperphosphatemia that may be potentially life-threatening.
Therefore there is a need of semi-selective 1,25D analogs that
retain high differentiating and anti-proliferative activities with
minimal or tolerable calcemic and phosphatemic activities (4).
1,25D is a steroid compound, which is either
produced in skin from 7-dehydrocholesterol or delivered with food.
There are two signal transduction pathways activated by 1,25D in
target cells. The most important and the best documented is so
called ‘genomic pathway’ that consists in activation of vitamin D
receptor (VDR). VDR belongs to the superfamily of nuclear receptors
for steroid and thyroid hormones. In order to be active VDR
heterodimerizes with the retinoid X receptor (RXR). VDR upon
ligation translocates to the cell nucleus and undergoes
conformational changes, that allow binding to specific sequences
called vitamin D response elements (VDRE) localized in promoter
regions of target genes. Binding of 1,25D to VDR enhances
heterodimerization with RXR and allows binding of the coactivators,
known as vitamin D receptor-interacting protein complex (DRIP)
(5). The complex includes histone
acetylase which relaxes chromatin structure to make DNA accessible
for RNA polymerase and to initiate transcription of target genes
(6). The less precisely described
is ‘non-genomic pathway’ which consists of intracellular signalling
molecules, such as mitogen-activated protein kinases (MAPKs),
phosphatidylinositol 3-kinase (PI3K) and others activated by a
putative membrane VDR (mVDR) (7).
In our recent paper we described pro-differentiating
effects of a series of vitamin D2 analogs with extended
and branched side-chains. Analogs with side-chains extended by one
(PRI-1906) or two carbon units (PRI-1907) displayed elevated
cell-differentiating activity in comparison to 1,25D (8). Moreover, previous studies in mice have
shown that PRI-1906 is less calcemic, while PRI-1907 was more
calcemic than 1,25D (9,10). Analogs PRI-1906 and PRI-1907 were
more active than 1,25D in inducing cell differentiation of HL60,
NB-4, MV4-11, U-937 and MOLM-13 acute myeloid leukemia (AML) cells.
Moreover, towards some cell lines PRI-1907 was more active than
PRI-1906, and both were more active than 1,25D. However, these
three compounds had similar potency to inhibit proliferation of
prostate cancer cells PC-3 (8). The
ability of these analogs to induce expression of CYP24A1
gene in HL60 cells was tested, and the increase in CYP24A1
mRNA appeared to be lower in cells exposed to PRI-1907 than in
cells exposed to either 1,25D or PRI-1906. The above mentioned
results have shown that these two analogs are selective not only in
calcemic and pro-differentiating activities, but also are
differentialy active in different cell lines.
As it was discussed recently (11), the potential mechanisms through
which selectivity of analogs could be achieved include interactions
with serum transporting proteins, efficiency of cellular uptake,
interactions with intracellular binding proteins, intracellular
metabolism to inactive endproducts or to active intermediary
metabolites, ligand-induced conformational changes in the VDR that
affect binding to the DNA or to the complex of protein coregulators
and eventually, activation of the non-genomic pathways through a
putative mVDR. In case of our analogs the interactions with serum
proteins could not have been of any importance, because experiments
were done in vitro. All other mechanisms mentioned above
could have an impact on selective biological actions of PRI-1906
and PRI-1907. Observed differences in regulation of CYP24A1 by our
analogs in HL60 cells encouraged us to study if in other cell lines
similar differences occur. We also wanted to verify if differences
in CYP24A1 mRNA levels in HL60 cells were followed by
different CYP24A1 protein levels in the cell mitochondria.
Eventually, in attempts to find the mechanism of selectivity of
PRI-1906 and PRI-1907, we examined changes in expression and
subcellular localization of VDR protein and downstream
transcription factors in response to our analogs.
Materials and methods
Cell lines
HL60 cells were obtained from the European
Collection of Cell Cultures. NB-4 cells were a kind gift from
Professor G.P. Studzinski (University of Medicine and Dentistry of
New Jersey). U-937, MV4-11 and MOLM-13 were purchased from German
Ressource Center for Biological Material (Deutsche Sammlung von
Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany). The
cells were propagated as suspension cultures in RPMI-1640 medium
supplemented with 10% fetal calf serum (FCS, Sigma, St. Louis, MO),
100 U/ml penicillin and 100 μg/ml streptomycin (Sigma). The cells
were kept at standard cell culture conditions, i.e., humidified
atmosphere of 95% air and 5% CO2 at 37°C. The cell
number and viability were determined by hemocytometer counts and
trypan blue (0.4%) exclusion. For all experiments the cells were
suspended in fresh medium containing 1,25D, analog or the
equivalent volume of ethanol as a vehicle control.
Chemicals and antibodies
1,25D and all analogs were synthesized in the
Pharmaceutical Research Institute (Warsaw, Poland). The compounds
were aliquoted and stored in glass ampoules under argon at −20°C.
Amount of the analog in an ampoule was determined by UV
spectrometry at 264 nm, compound was dissolved in an absolute
ethanol to 100 μM, and subsequently diluted in the culture medium
to the required concentration.
Antibodies CD14-PE and isotypic control-PE were from
ImmunoTools (Friesoythe, Germany). Chemiluminescence Blotting
Substrate was from Roche Diagnostics (Mannheim, Germany). Mouse
monoclonal anti-Hsp90, rabbit polyclonal anti-C/EBPβ and anti-VDR
antibodies were from Santa Cruz Biotechnology, Inc. (Santa Cruz,
CA). Goat anti-rabbit IgG and anti-mouse IgG conjugated to
peroxidase were from Jackson ImmunoResearch (West Grove, PA).
Rabbit anti-actin antibodies were from Sigma.
Determination of cell
differentiation
The expression of cell surface markers of monocytic
differentiation was determined by flow cytometry. The cells were
incubated with 1,25D or analogs and then stained with 1 μl of
CD14-PE for 1 h on ice. Next, they were washed three times with
ice-cold PBS and suspended in 0.5 ml PBS prior to analysis on FACS
Calibur flow cytometer (Becton-Dickinson, San Jose, CA). The
acquisition parameters were set for an isotype control. Data
analysis was performed with use of WinMDI 2.8 software (freeware by
Joseph Trotter). Differentiation assays were repeated from 3 to 5
times.
Preparation of cell lysates
In order to prepare cytosolic and nuclear lysates,
the cells (5×106/sample) were washed 3 times with PBS
and lysed for 20 min on ice in 80 μl of lysis buffer (20 mM Tris,
150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton X-100; pH 7.5)
containing protease inhibitor cocktail (Roche Diagnostics). The
lysates were separated by centrifugation for 5 min, at 18000 × g,
at 4°C. Supernatants were designated the cytoplasmic (C) fraction,
and the nuclei remaining in pellets after one washing were
sonicated for 10 sec in the same volume of lysis buffer as before
(80 μl/5×106 cells). Following sonication nuclei were
centrifuged again for 5 min, at 18000 × g, at 4°C and the final
supernatants were designated the nuclear (N) fraction. Samples were
denatured by adding 20 μl of 5× sample buffer and boiling for 5
min.
In order to prepare cytosolic, membrane, nucleosolic
and chromatin fractions 6×106 cells/sample (equivalent
of 20 μl packed cell volume) were washed with PBS and lysed using
Pierce Subcellular Protein Fractionation kit according to the
user’s manual. Obtained lysates were denatured by adding 5× sample
buffer (1/4 volume of the lysate) and boiled for 5 min.
To prepare mitochondrial extracts 2×107
cells/sample were washed with PBS and then the mitochondria were
isolated using Pierce Mitochondria Isolation kit for cultured cells
according to the user’s manual. Obtained mitochondrial pellets were
lysed, denatured in 80 μl of 1× sample buffer and boiled for 5
min.
Western blot analysis
For western blotting 30 μl of cell lysates were
separated on 12.5% SDS-PAGE gels and transferred to PVDF membranes.
The membranes were then dried, and incubated sequentially with
primary and a horseradish peroxidase-conjugated secondary antibody
(1 h, room temperature). The protein bands were visualized with
chemiluminescence. Then the membranes were stripped, dried again
and probed with subsequent antibodies. Western blots were repeated
from 2 to 5 times.
cDNA synthesis and real-time PCR
Total RNA was isolated using TriPure (Roche
Diagnostics) as per manufacturer’s recommendations. RNA quantity
was determined using Nanodrop (Thermo Fisher Scientific, Inc.
Worcester, MA) and the quality of RNA was verified by gel
electrophoresis. RNA was transcribed into cDNA using High Capacity
cDNA Reverse Transcription kit (Applied Biosystems, Foster City,
CA). Real-time RT-PCR reaction was performed using SYBR-Green PCR
Mix (A&A Biotechnology, Gdansk, Poland) and Applied Biosystems
StepOne system. The sequences of CYP24A1 and GAPDH primers and
reaction conditions were as described previously (8). Real-time PCR assays were repeated 3
times.
Results
Comparison of pro-diffrentiating
activities of 1,25D and analogs
Previously, we showed that various AML cell lines
when exposed to either 1,25D or vitamin D2 analogs
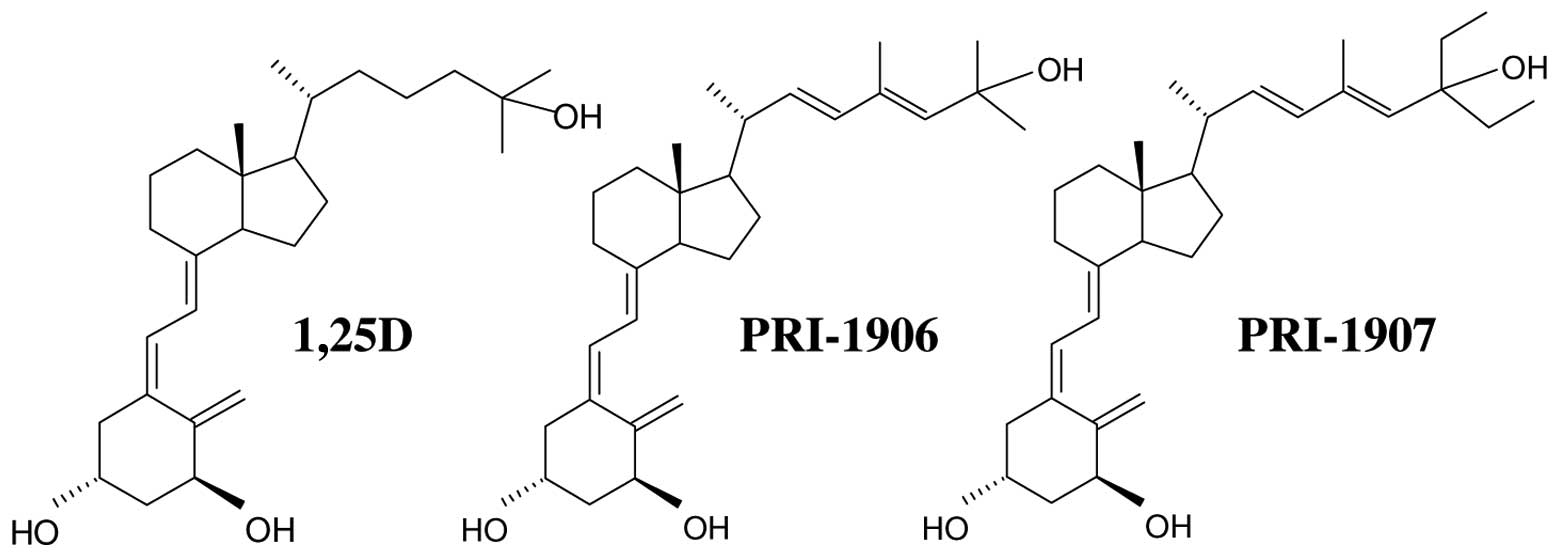
PRI-1906 and PRI-1907 (structures given in Fig. 1) acquire cell differentiation
markers characteristic of monocytes. The most strongly upregulated
is a key macrophage marker CD14, which functionally is a
co-receptor for bacterial lipopolysaccharide (12). In order to compare potencies of
these compounds in inducing cell differentiation we calculated from
differentiation graphs the concentrations of the analogs that cause
50% (C50%) of treated cells to be CD14 positive. The
values of C50%, given in Table I, show that in all cell lines
studied PRI-1906 and PRI-1907 were more effective than 1,25D, what
means that C50% for PRI-1906 and PRI-1907 were from 4 to
14 times lower than C50% for 1,25D. Moreover, in three
cell lines (HL60, NB-4 and MV4-11) C50% values for
PRI-1907 were about 4 times lower than for PRI-1906. At
concentrations around 10 nM, differentiating effects of the three
compounds were similar, but when the analogs studied were applied
at concentrations of 1 nM and lower, the differences in their
potencies were evident.
 | Table IPro-diffrentiating activities of 1,25D
and analogs towards human AML cell lines. |
Table I
Pro-diffrentiating activities of 1,25D
and analogs towards human AML cell lines.
| Compound | HL60 | NB4 | MV4-11 | U-937 | MOLM-13 |
|---|
| 1,25D | 1.19 | 1.26 | 1.39 | 5.96 | 1.81 |
| PRI-1906 | 0.39 | 0.43 | 0.40 | 1.09 | 0.39 |
| PRI-1907 | 0.12 | 0.15 | 0.10 | 1.28 | 0.33 |
Expression of CYP24A1 mRNA in response to
1,25D and analogs
CYP24A1 gene encodes an enzyme,
24-hydroxylase of 1,25D, which is the key enzyme in degradation of
1,25D to calctirioic acid and it was documented that CYP24A1
is the most strongly regulated out of all 1,25D-target genes
(13). We have reported that in
HL60 cells, the expression of CYP24A1 was indeed very
strong, but slowly upregulated in response to 1,25D (8). Surprisingly in HL60 the expression of
CYP24A1 cells was upregulated less by the analog PRI-1907
than by either 1,25D or PRI-1906. To determine whether similar
pattern of regulation can be observed in other AML cells, for these
studies two other AML cell lines were used. MV4-11 cell line, in
which the analogs studied were similarly efficient in inducing CD14
expression as in HL60 cells, indicating that PRI-1907 was more
active than PRI-1906 and both were more active than 1,25D and
MOLM-13 cells in which PRI-1906 and PRI-1907 were similarly active,
and both were more active than 1,25D. Since our previous
experiments have shown that expression of mRNA for CYP24A1
increases very slowly in response to 1,25D, for current experiments
we selected 96 h of exposure to the compounds at 10 nM
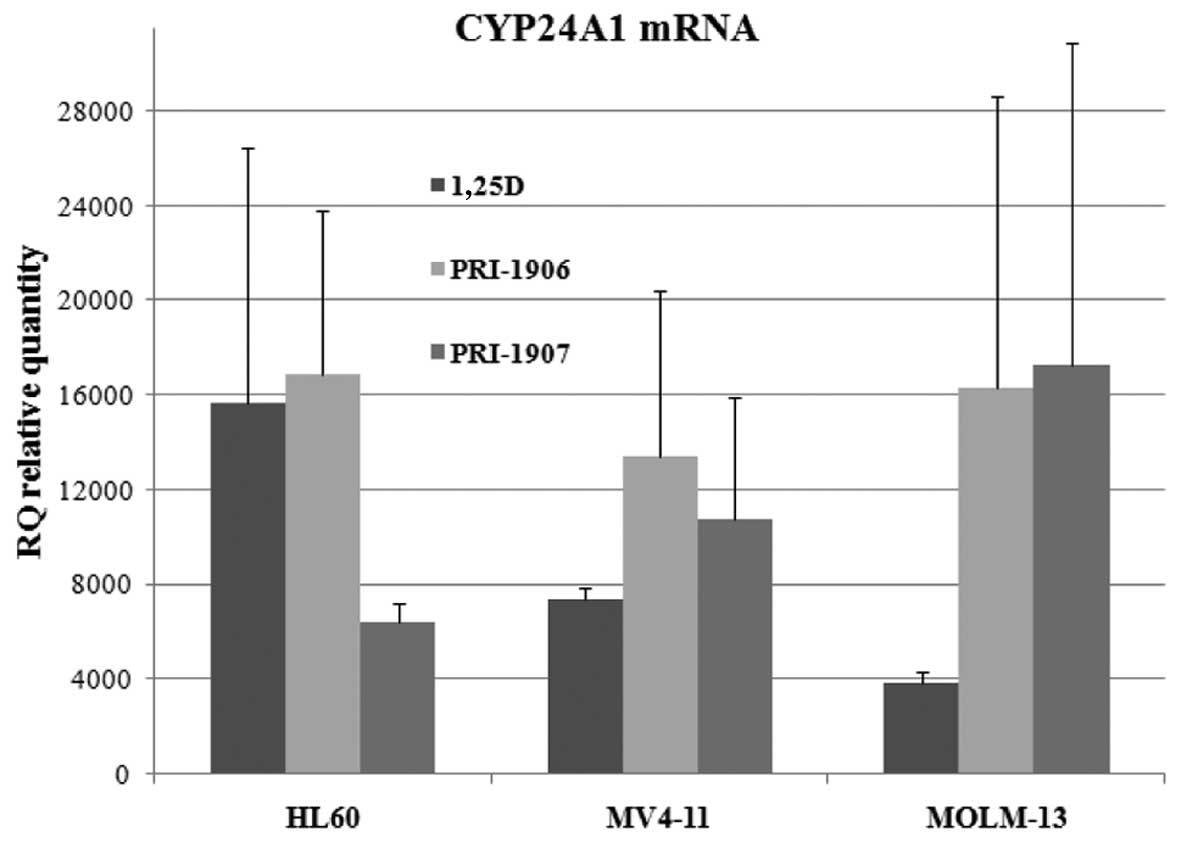
concentrations. Results which are presented in Fig. 2 show that unlike in HL60 cells, in
MV4-11 and in MOLM-13 cells, analogs PRI-1906 and PRI-1907
upregulated expression of CYP24A1 stronger than 1,25D, however due
to high standard deviations, differences were not significant.
Influence of CYP24A1 inhibitors to
1,25D-induced cell differentiation
Data obtained using prostate cancer models show that
ketoconazole and other CYP24A1 inhibitors are able to enhance
anti-proliferative action of 1,25D in vitro and in
vivo(14-16). Since CYP24A1 is highly inducible by
1,25D in human AML cells, presumably resulting in an increased
1,25D degradation, an inhibition of the enzyme should cause its
delayed catabolism and enhanced differentiation effect. Therefore
in our next experiments HL60 cells were exposed simultaneously to
1,25D and various inhibitors of cytochrome P450 family of enzymes,
which among others inhibit activity of CYP24A1. The inhibitors used
were ketoconazole at concentrations ranging from 0.1 to 6 nM,
proadifen at concentrations ranging from 0.1 to 10 nM and
1-aminobenzotriazole at concentrations ranging from 0.1 to 100 nM.
The concentrations were verified as not toxic to the cells, before
starting differentiation assays. The cells were exposed to either 1
or to 10 nM 1,25D ± the inhibitors given above. The cells were
exposed for 96 h and then the expression of CD11b and CD14 was
tested in flow cytometry. Surprisingly, neither inhibitor was able
to induce any changes to 1,25D-induced expression of cell surface
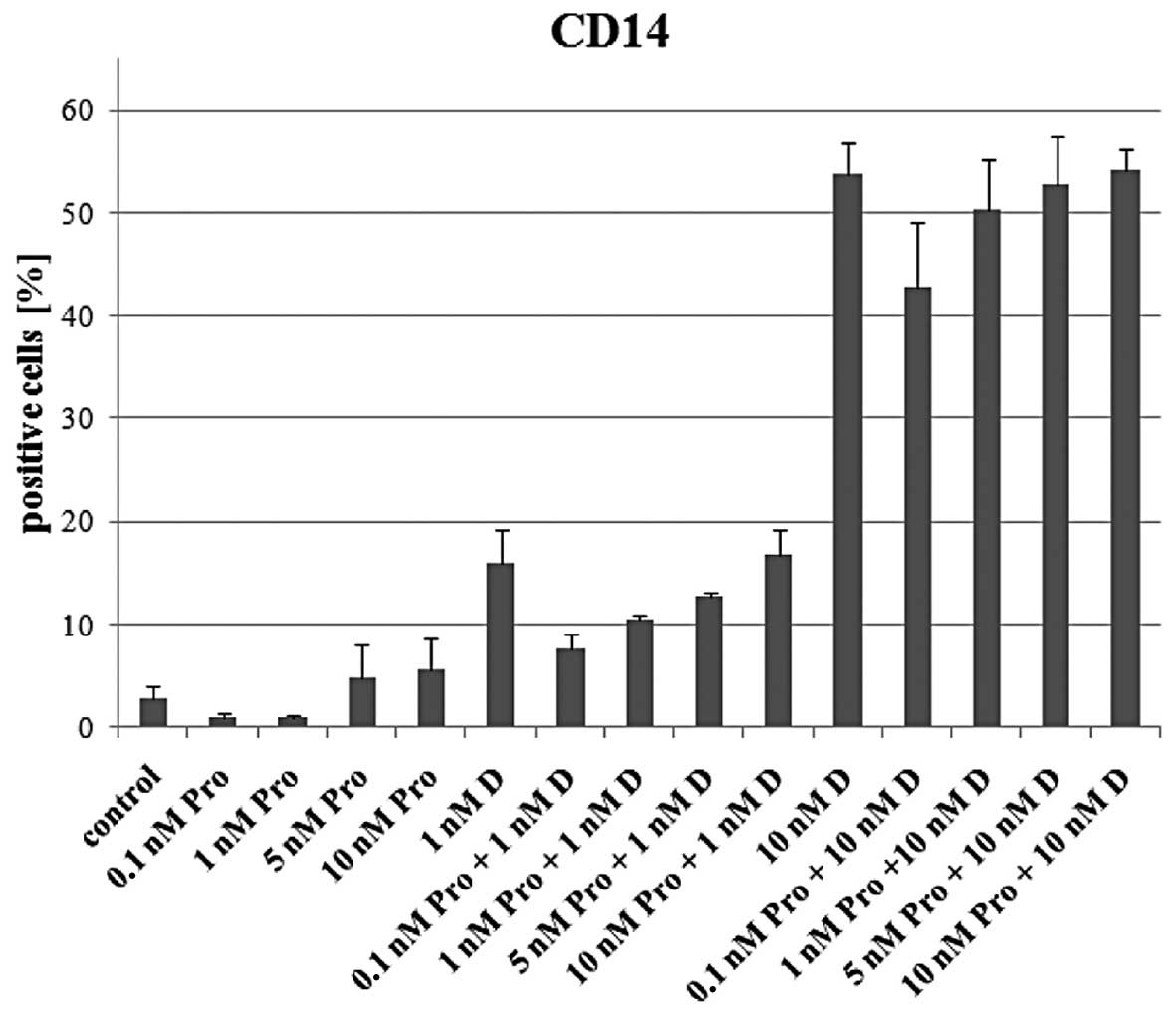
markers. As an example the graph showing expression of CD14 in
cells exposed to 1,25D and proadifen is presented in Fig. 3. The values of CD14 expression in
cells exposed to 1,25D and in cells exposed to proadifen and 1,25D
are comparable.
Protein levels of CYP24A1 in HL60 cells
exposed to 1,25D and analogs
In the next experiments we wanted to verify if
differences seen in CYP24A1 mRNA levels in HL60 cells are
mirrored by CYP24A1 protein levels. Since CYP24A1 is a
mitochondrial inner-membrane cytochrome P450 enzyme, in order to
precisely identify its levels, mitochondria were isolated from HL60
cells exposed either to 1,25D or to the analogs. At first, the
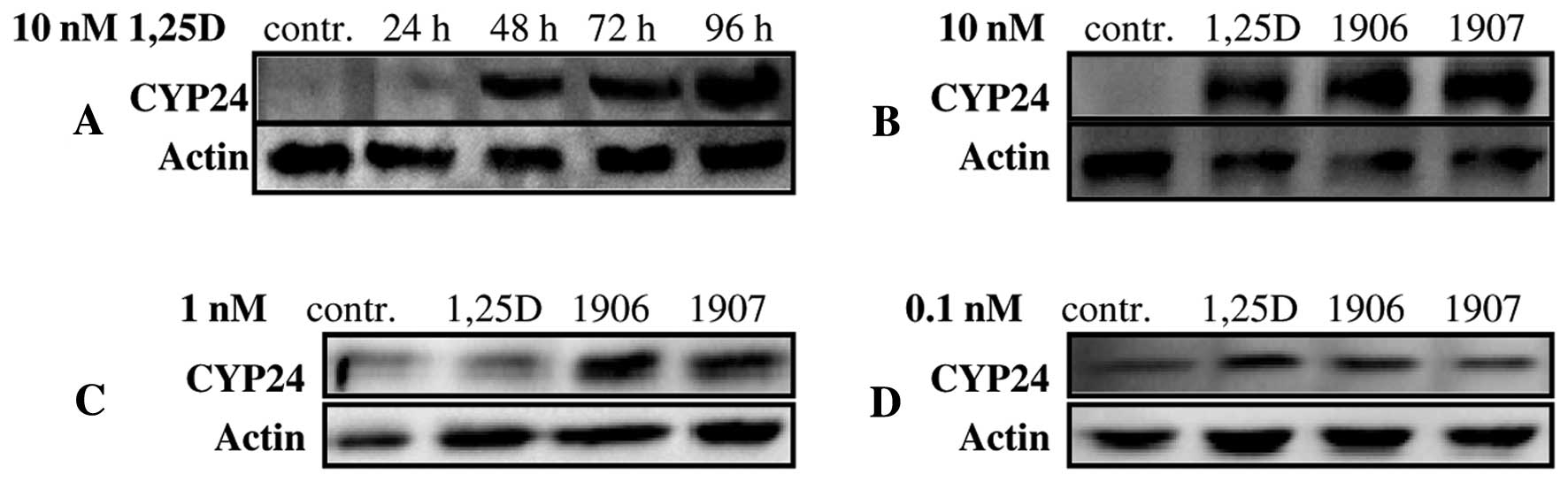
kinetics of CYP24A1 upregulation in cells exposed to 10 nM 1,25D
was studied. Data from Fig. 4A show
that CYP24A1 is expressed as a protein in HL60 cells at a very low,
hardly detectable level, but after exposure to 1,25D it grows
slowly, but significantly. Then the 96 h exposure-time was chosen
to compare abilities of 1,25D and of the analogs to increase levels
of CYP24A1 in mitochondria. As presented in Fig. 4B 1,25D, PRI-1906 and PRI-1907 at 10
nM concentrations induced comparable levels of CYP24A1 in HL60
cells. Then the analogs were applied at 1 and 0.1 nM
concentrations, and CYP24A1 was detected in western blots. When the
cells were exposed to 1 nM compounds, CYP24A1 was detectable, but
levels induced by PRI-1906 and PRI-1907 were higher than levels
induced by 1,25D (Fig. 4C). It
appeared that levels of CYP24A1 in cells exposed to 0.1 nM
compounds were similar to the control, and the western blot
membranes had to be overexposed to see the bands (Fig. 4D).
Upregulation and subcellular localization
of VDR and C/EBPβ in HL60 cells exposed to 1,25D and analogs
In light of the data presented above, the
differences in expression of CYP24A1 do not seem responsible for
different activities of 1,25D analogs. However, as mentioned
before, differences in the analog activities were the most evident
when compounds were applied at low concentrations (≤1 nM). Thus
HL60 cells were exposed to 1,25D, PRI-1906 and PRI-1907 at 0.1 and
1 nM concentrations, and the levels of VDR and
CCAAT-enhancer-binding protein β (C/EBPβ) were tested in cell
lysates. C/EBPβ was studied because this transcription factor is a
master regulator of monocyte differentiation, and its upregulation
and importance for differentiation of cells exposed to 1,25D were
documented (17,18). C/EBPβ is translated from its mRNA in
three isoforms of different lengths. The two larger isoforms, full
length C/EBPβ-1 and slightly shorter version C/EBPβ-2 contain
several transactivation domains, whereas the much shorter variant,
C/EBPβ-3 lacks them (19). At first
the expression of VDR and C/EBPβ was tested in HL60 cells
fractionated into cytosol and nuclei. Previously we documented that
this fractionation method efficiently separates cytosolic proteins
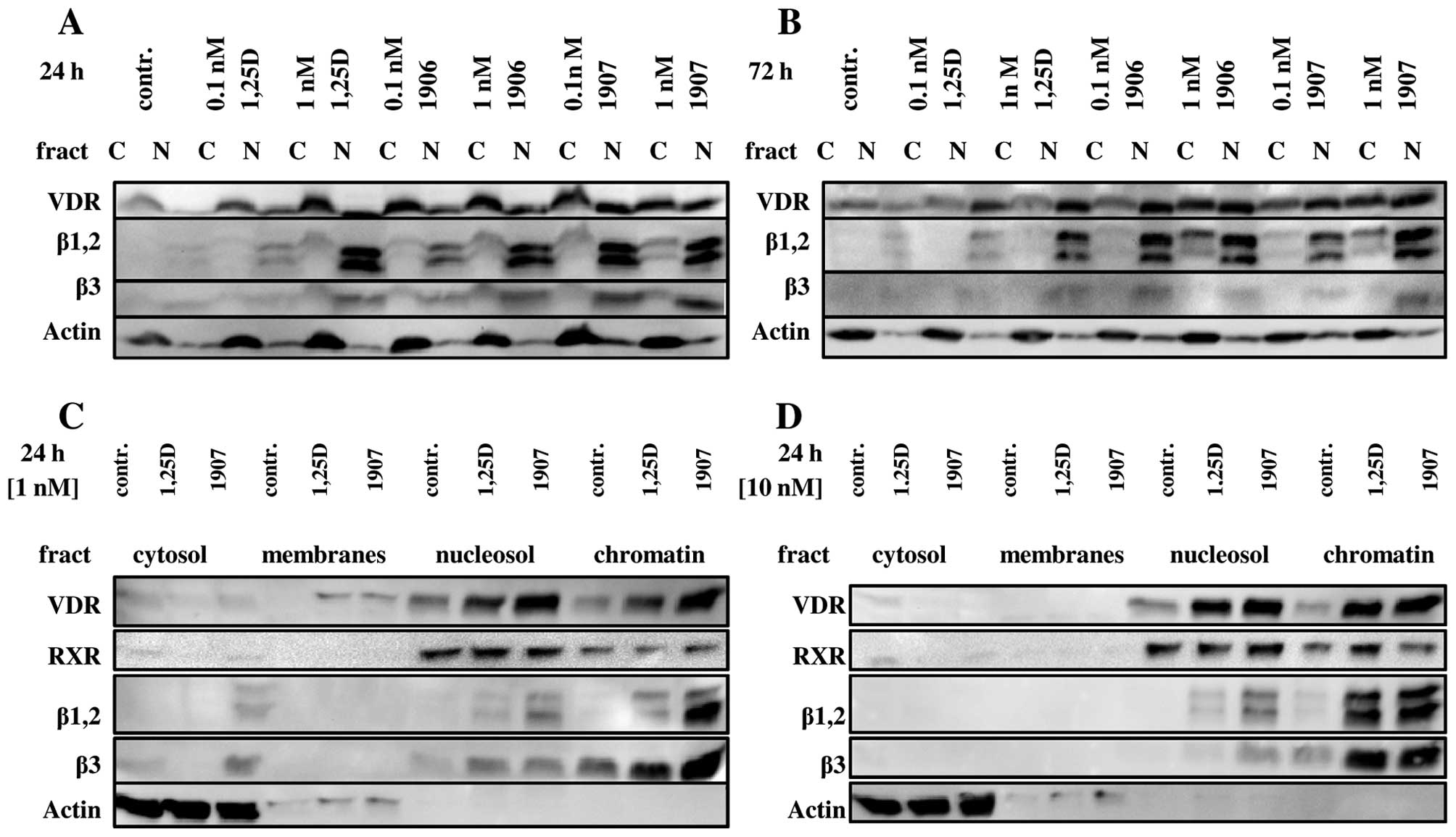
from nuclear proteins (8,20). As presented in Fig. 5A and B, 0.1 nM 1,25D slightly
increased nuclear content of VDR, and even less the levels of
C/EBPβ isoforms. In contrast, 0.1 nM PRI-1907 strongly increased
nuclear content of VDR and C/EBPβ, particularly after 24 h
exposure. Also PRI-1906 applied at 0.1 nM concentration increased
both VDR and C/EBPβ isoforms in nuclei of treated cells,
predominantly after 72 h exposure. In the next series of
experiments HL60 cells exposed either to 1,25D or to PRI-1907 were
divided into more discrete cellular fractions using commercially
available Pierce Subcellular Protein Fractionation kit, namely into
cytosol, membranes, nucleosol and chromatin. When this
fractionation protocol was applied, further differences were
observed. First of all, almost equal distribution of VDR between
nucleosol and chromatin was detected (Fig. 5C and D). When 1,25D and PRI-1907
were applied at 1 nM concentrations differences in VDR protein
levels were clearly visible, but they were less evident when the
compounds were used at 10 nM concentrations. Isoforms of C/EBPβ
presented different pattern of distribution, as they were
predominantly bound to chromatin. All isoforms of C/EBPβ were
upregulated by the compounds studied, however 0.1 nM PRI-1907 was
more efficient than 0.1 nM 1,25D in upregulation of chromatin-bound
C/EBPβ isoforms.
Discussion
The major goal of many chemical laboratories is to
develop vitamin D analogs that retain clinically useful activities
of 1,25D with minimal calcemic and phosphatemic activities
(4). There are some promising
analogs that retain high pro-differentiating and anti-proliferative
activities in vitro and express low calcemic activities
in vivo. Even though many interesting analogs have been
made, a clear pattern of structure-function relationship did not
emerge (21). How complicated such
structure-function relationships could be is well illustrated by a
series of vitamin D2 analogs with extended and branched
side-chains (22). These analogs
have an additional double bond in the side-chain, which is further
extended by carbon units in an aliphatic and not alicyclic manner.
The differentiating activity in AML cells increased from the analog
with side-chain extended by a pair of methyl groups (PRI-1906),
reaching maximum for the analog extended by a pair of ethyl groups
at C-25 (PRI-1907). Further extensions of side-chain by either
n-propyl (PRI-1908) or n-butyl (PRI-1909) groups caused sudden drop
in pro-differentiating activities (8). Surprisingly, the minimum calcemic
activity in vivo was observed for the analog PRI-1906, and
was significantly lower than for 1,25D, as well as for PRI-1907
(10). Our earlier results
suggested that the mechanism of superagonistic differentiating
activity of PRI-1907 in HL60 cells could lie in its weak ability to
upregulate expression of CYP24A1, a major catabolizing enzyme for
vitamin D. This hypothesis was addressed in the current study.
However, the results presented here demonstrate that such
explanation is very unlikely. First of all, in another cell line in
which differentiating activity of PRI-1907 was stronger than the
activities of 1,25D and PRI-1906, namely in MV4-11 cell line,
levels of CYP24A1 mRNA in response to PRI-1907 were higher
than in response to 1,25D. Moreover, inhibition of catalytic
activity of CYP24A1 protein using its inhibitors, have not
increased 1,25D-induced monocytic differentiation, and finally the
protein levels of CYP24A1 in HL60 cells exposed to 1,25D, PRI-1906
and PRI-1907 have not reflected mRNA levels detected in real-time
PCR. We assume that it is important that mRNA and protein levels of
CYP24A1 in AML cells increase very slowly after exposure of the
cells to vitamin D compounds. The differentiation markers can be
observed faster than CYP24A1 (23,24).
Therefore we suppose that signals for differentiation are already
‘on’ when catabolism of vitamin D compounds starts. Consequently,
we assume that altered catabolism of PRI-1907 is not a cause of its
superagonistic differentiating actions in certain AML cell lines.
There are some other possible mechanisms of superagonistic
activities of analogs, for example higher efficiency of cellular
uptake or altered interactions with intracellular binding proteins
(11). Since the biggest
differences between differentiating activities of 1,25D and
PRI-1906 and PRI-1907 can be noticed when the compounds are applied
at concentrations of 1 nM and less, we studied intracellular levels
of VDR and C/EBPβ isoforms in HL60 cells exposed to 0.1 and 1 nM
compounds. As was presented in our previous publications VDR is a
rapid and sensitive indicator of intracellular presence of 1,25D
and its analogs, as the protein is immediately stabilized and
accumulated in cell nuclei after ligation (25,26).
Therefore, the observation that nuclear levels of VDR are similar
after exposure of the cells to either 0.1 nM PRI-1906 or 0.1 nM
PRI-1907 as after 1 nM 1,25D may suggest that the two analogs are
either more efficiently transported through cellular membrane, or
have higher affinities to VDR. Nuclear accumulation of C/EBPβ
transcription factor is more delayed response of the cells to
1,25D, as it can be observed not earlier than at 24 h, and reaches
its maximum at 72 h from the exposure time (18). C/EBPβ is a regulator of monocytic
differentiation, it directly regulates transcription of many
monocyte-specific proteins, such as CD14, lactoferrin or lyzozyme
(19). Moreover, recently an
involvement of two longer isoforms C/EBPβ-1 and C/EBPβ-2 in
differentiation-related inhibition of proliferation was reported
(27). As presented herein, all
three isoforms of C/EBPβ accumulate in the nuclei of cells exposed
to 0.1 nM PRI-1906 or 0.1 nM PRI-1907, while for similar effect 1
nM 1,25D is needed. Almost entire nuclear content of C/EBPβ remains
bound to chromatin, consequently suggesting transcriptional
activity of these proteins. On the other hand the observation that
at least half of the nuclear content of VDR remains in the soluble
nucleosol, most probably in dimers with RXR, is surprising and
suggests that the role of VDR in the nuclei of AML cells is not
limited to transcription. In summary, we conclude that
superagonistic differentiating activities of analogs PRI-1906 and
PRI-1907 do not seem to be caused by their altered catabolism, but
most probably by altered interactions with VDR and resulting
downstream proteins.
Acknowledgements
The research was supported by Wroclaw Research
Center EIT+ under the Project ‘Biotechnologies and
advanced medical Technologies-BioMed’ (POIG 01.01.02-02-003/08-00)
financed by the European Regional Development Fund (Operational
Program Innovative Economy, 1.1.2).
References
|
1
|
McCarthy D, San Miguel J, Freake H, Green
P, Zola H, Catovsky D and Goldman J: 1,25-Dihydroxyvitamin
D3 inhibits proliferation of human promyelocytic
leukaemia (HL60) cells and induces monocyte-macrophage
differentiation in HL60 and normal bone marrow cells. Leuk Res.
7:51–55. 1983.
|
|
2
|
Abe E, Miyaura C, Sakagami H, Takeda M,
Konno K, Yamazaki T, Yoshiki S and Suda T: Differentiation of mouse
myeloid leukemia cells induced by 1-alpha,25-dihydroxyvitamin
D3. Proc Natl Acad Sci USA. 78:4990–4994. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van Etten E and Mathieu C:
Immunoregulation by 1,25-dihydroxyvitamin D3: basic
concepts. J Steroid Biochem Mol Biol. 97:93–101. 2005.PubMed/NCBI
|
|
4
|
Brown A and Slatopolsky E: Vitamin D
analogs: therapeutic applications and mechanisms for selectivity.
Mol Aspects Med. 29:433–452. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rachez C, Lemon B, Suldan Z, Bromleigh V,
Gamble M, Naar A, Erdjument-Bromage H, Tempst P and Freedman L:
Ligand-dependent transcription activation by nuclear receptors
requires the DRIP complex. Nature. 398:824–828. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aranda A and Pascual A: Nuclear hormone
receptors and gene expression. Physiol Rev. 81:1269–1304.
2001.PubMed/NCBI
|
|
7
|
Mizwicki M and Norman A: Vitamin D
sterol/VDR conformational dynamics and non-genomic actions. Vitamin
D. Feldman D, Pike J and Adams J: 3rd edition. Academic Press;
London: pp. 271–297. 2011, View Article : Google Scholar
|
|
8
|
Baurska H, Klopot A, Kielbinski M, Chrobak
A, Wijas E, Kutner A and Marcinkowska E: Structure-function
analysis of vitamin D2 analogs as potential inducers of
leukemia differentiation and inhibitors of prostate cancer
proliferation. J Steroid Biochem Mol Biol. 126:46–54. 2011.
|
|
9
|
Ji Y, Kutner A, Verstuyf M, Verlinden L
and Studzinski G: Derivatives of vitamins D2 and
D3 activate three MAPK pathways and upregulate pRb
expression in differentiating HL60 cells. Cell Cycle. 1:410–415.
2002.
|
|
10
|
Wietrzyk J, Nevozhay D, Milczarek M, Filip
B and Kutner A: Toxicity and antitumor activity of the vitamin D
analogs PRI-1906 and PRI-1907 in combined treatment with
cyclophosphamide in a mouse mammary cancer model. Cancer Chemother
Pharmacol. 62:787–797. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brown AJ: Mechanisms for the selective
actions of vitamin D analogs. Vitamin D. Feldman D, Pike JW and
Adams JS: 3rd edition. Academic Press; London: pp. 1436–1459.
2011
|
|
12
|
Chow CW, Grinstein S and Rotstein OD:
Signaling events in monocytes and macrophages. New Horiz.
3:342–351. 1995.PubMed/NCBI
|
|
13
|
Kahlen J and Carlberg C: Identification of
a vitamin D receptor homodimer-type response element in the rat
calcitriol 24-hydroxylase gene promoter. Biochem Biophys Res
Commun. 202:1366–1372. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Muindi JR, Yu WD, Ma Y, Engler KL, Kong
RX, Trump DL and Johnson CS: CYP24A1 inhibition enhances the
antitumor activity of calcitriol. Endocrinology. 151:4301–4312.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yee SW, Campbell MJ and Simons C:
Inhibition of Vitamin D3 metabolism enhances VDR
signalling in androgen-independent prostate cancer cells. J Steroid
Biochem Mol Biol. 98:228–235. 2006.
|
|
16
|
Peehl DM, Seto E, Hsu JY and Feldman D:
Preclinical activity of ketoconazole in combination with calcitriol
or the vitamin D analogue EB 1089 in prostate cancer cells. J Urol.
168:1583–1588. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ji Y and Studzinski G: Retinoblastoma
protein and CCAAT/enhancer-binding protein β are required for
1,25-dihydroxyvitamin D3-induced differentiation of HL60
cells. Cancer Res. 64:370–377. 2004.
|
|
18
|
Marcinkowska E, Garay E, Gocek E, Chrobak
A, Wang X and Studzinski G: Regulation of C/EBPβ isoforms by MAPK
pathways in HL60 cells induced to differentiate by
1,25-dihydroxyvitamin D3. Exp Cell Res. 312:2054–2065.
2006.
|
|
19
|
Ramji D and Foka P: CCAAT/enhancer-binding
proteins: structure, function and regulation. Biochem J.
365:561–575. 2002.PubMed/NCBI
|
|
20
|
Wiedlocha A, Nilsen T, Wesche J, Sorensen
V, Malecki J, Marcinkowska E and Olsnes S:
Phosphorylation-regulated nucleocytoplasmic trafficking of
internalized fibroblast growth factor-1. Mol Biol Cell. 16:794–810.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
DeLuca HL and Plum LA: Alterations in
1,25-dihydroxyvitamin D3 structure that produce profound
changes in in vivo activity. Vitamin D. Feldman D, Pike JW
and Adams JS: 3rd edition. Academic Press; London: pp. 1429–1435.
2011
|
|
22
|
Chodynski M, Wietrzyk J, Marcinkowska E,
Opolski A, Szelejewski W and Kutner A: Synthesis and
antiproliferative activity of side-chain unsaturated and
homologated analogs of 1,25-dihydroxyvitamin D2.
(24E)-(1S)-24-Dehydro-24a-homo-1,25-dihydroxyergocalciferol and
congeners. Steroids. 67:789–798. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marcinkowska E, Wiedlocha A and
Radzikowski C: Evidence that phosphatidylinositol 3-kinase and
p70S6K protein are involved in differentiation of HL-60 cells
induced by calcitriol. Anticancer Res. 18:3507–3514.
1998.PubMed/NCBI
|
|
24
|
Marcinkowska E, Wang X and Studzinski G:
C/EBPβ: a candidate for a major player in vitamin D-induced
monocytic differentiation of human leukemia cells. Vitamin D: New
Research. Stolzt V: Nova Science Publishers Inc; New York, NY: pp.
25–40. 2006
|
|
25
|
Gocek E, Kielbinski M, Wylob P, Kutner A
and Marcinkowska E: Side-chain modified vitamin D analogs induce
rapid accumulation of VDR in the cell nuclei proportionately to
their differentiation-inducing potential. Steroids. 73:1359–1366.
2008. View Article : Google Scholar
|
|
26
|
Gocek E, Kielbinski M and Marcinkowska E:
Activation of intracellular signaling pathways is necessary for an
increase in VDR expression and its nuclear translocation. FEBS
Lett. 581:1751–1757. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gutsch R, Kandemir J, Pietsch D, Cappello
C, Meyer J, Simanowski K, Huber R and Brand K:
CCAAT/enhancer-binding protein β inhibits proliferation in
monocytic cells by affecting the retinoblastoma protein/E2F/cyclin
E pathway but is not directly required for macrophage morphology. J
Biol Chem. 286:22716–22729. 2011.
|