Introduction
Epstein-Barr virus (EBV) is a ubiquitous human
γ-herpesvirus and was the first human virus linked to cancer
(1). EBV can infect and immortalize
B-lymphocytes in vitro, giving rise to lymphoblastoid cell
lines (LCLs). Consistent with this feature, persistent, latent EBV
infection is present in several lymphoid malignancies including
Burkitt’s lymphoma (BL) and Hodgkin’s lymphoma (HL) (2).
BL represents a subtype of high-grade mature B-cell
non-Hodgkin’s lymphoma and is characterized by rapid cell
proliferation and a generally aggressive clinical course. BL is
typically seen in areas where malaria is endemic and can represent
up to 50% of childhood cancers in some regions. In this endemic
form of BL disease, EBV is present in all cases. BL also occurs
sporadically at lower incidence throughout the world with
associated lower levels of EBV positivity (2). The frequency of BL has increased in
low-incidence countries since the 1980s, following the advent of
human immunodeficiency virus/acquired immunodeficiency syndrome,
and among these cases 30–40% of tumors are EBV-positive. Patients
with human immunodeficiency virus-associated lymphoma pose
additional therapeutic challenges, particularly the risk of
overwhelming opportunistic infections (3).
HL is a B cell lymphoma generally comprising only a
small proportion of the characteristic malignant
Hodgkin/Reed-Sternberg (HRS) cells, but with abundant non-malignant
immunocytes recruited by various cytokines secreted by HRS cells.
EBV is present in the HRS cells in ~40% of classical HL cases
(2). Although multidrug approaches
have been very successful in the treatment of HL, unfortunately,
latent toxicity of these agents appear after several years in the
form of secondary malignancies and cardiovascular disease (4). In addition, some patients will suffer
from refractory disease or experience a relapse. Therefore, the
current goal in HL treatment is to find new therapies that
specifically target the deregulated signaling cascades that cause
HRS cell proliferation and resistance to apoptosis. In many
cancers, nuclear factor-κB (NF-κB) is constitutively activated,
protecting the tumor cells against apoptosis. Indeed, lesions from
patients with EBV-associated B-cell lymphomas show activation of
NF-κB, and thus inhibition of NF-κB is a much sought-after
therapeutic target in a variety of cancers (5,6).
Carotenoids are a family of natural pigments that
include fucoxanthin (FX), an abundant constituent of edible brown
algae. FX has several reported biological functions including
anti-oxidant, anti-inflammatory, anti-cancer, anti-obesity,
anti-diabetic, anti-angiogenic and anti-malarial activities
(7). FX is also an inhibitor of
pivotal proinflammatory mediators including nitric oxide and
cytokines, and affects signaling molecules involved in the
inflammatory processes such as NF-κB and mitogen-activated protein
kinases (8). In mammals, dietary FX
is deacetylated into fucoxanthinol (FXOH) in the intestinal tract
by lipases and esterases from the pancreas or intestinal cells, and
is then incorporated as FXOH into the blood circulation (7).
This study tested the hypothesis that FX and FXOH,
through the inhibition of NF-κB, could be potentially useful
therapeutic agents in the treatment of BL and HL.
Materials and methods
Extraction and isolation
FX was extracted from the brown seaweed
Cladosiphon okamuranus Tokida using acetone as the solvent,
and purified by column chromatography, liquid-liquid partition, and
then recrystallization up to >95% purity. It was further
purified by RP-HPLC up to >98% purity, in preparation for in
vitro assays. FXOH was prepared by enzymatic hydrolysis of the
purified FX using porcine pancreatic lipase. Briefly, 195 mg of FX,
2 g of sodium taurocholate and 2 g of porcine pancreatic lipase
(Type II; Sigma-Aldrich, St. Louis, MO) were dissolved in 30 ml of
0.1 M sodium phosphate buffer (pH 7.0). The reaction buffer was
incubated at 37°C for 3 h. FXOH was purified by ODS column
chromatography, liquid-liquid partition and recrystallization. We
prepared 142 mg of purified FXOH for this study (>95% purity,
72% yield). The FXOH was also further purified up to >98% purity
by RP-HPLC, for in vitro assays. The identity and purity of
the products were confirmed by comparison with reference FX (Wako
Pure Chemical Industries, Osaka, Japan) and data in the
literature.
Cells and cultures
Raji and Daudi are EBV-positive BL cell lines,
whereas BJAB and Ramos are EBV-negative BL cell lines. B95-8/Ramos
and B95-8/BJAB are Ramos and BJAB cells, respectively, infected
with the B95-8 strain of EBV. LCL-Ka and LCL-Ku are
EBV-immortalized human B-cell lines generated from peripheral blood
mononuclear cells (PBMC) of healthy volunteers. PBMC were isolated
by Ficoll-Paque density gradient centrifugation (GE Healthcare
Biosciences, Uppsala, Sweden) and washed with phosphate-buffered
saline. L428, KM-H2, HDLM-2 and L540 are HL cell lines. All cell
lines were cultured in Roswell Park Memorial Institute-1640 medium
supplemented with 10 or 20% heat-inactivated fetal bovine serum, 50
U/ml penicillin and 50 μg/ml streptomycin.
Cell viability and assay of
apoptosis
The effects of FX and FXOH on cell viability were
assessed using water-soluble tetrazolium (WST)-8 (Wako Pure
Chemical Industries). Briefly, 1×105 cells/ml were
incubated in a 96-well microculture plate in the absence or
presence of various concentrations of FX or FXOH. After 24-h
culture, WST-8 (5 μl) was added for the last 4 h of incubation and
the absorbance at 450 nm was measured using an automated microplate
reader. Mitochondrial dehydrogenase cleavage of WST-8 to formazan
dye provided a measure of cell proliferation. Apoptotic events in
cells were detected by staining with phycoerythrin-conjugated
Apo2.7 monoclonal antibody (Beckman-Coulter, Marseille, France)
(9) and analyzed by flow cytometry
(Epics XL, Beckman-Coulter, Fullerton, CA).
Cell cycle analysis
Cell cycle analysis was performed with the
Cycletest™ Plus DNA DNA reagent kit (Becton-Dickinson
Immunocytometry Systems, San Jose, CA). Briefly, 1×106
cells were washed with a buffer solution containing sodium citrate,
sucrose and dimethyl sulfoxide, suspended in a solution containing
RNase A, and then stained with 125 μg/ml propidium iodide for 10
min. Cell suspensions were analyzed on a Coulter EPICS XL using
EXPO32 software. The population of cells in each cell cycle phase
was determined with MultiCycle software.
In vitro measurement of caspase
activity
Caspase activity was measured with the colorimetric
caspase assay kits (MBL, Nagoya, Japan). Cell extracts were
prepared using cell lysis buffer and assessed for caspases-3 and -9
activities using colorimetric probes. Colorimetric caspase assay
kits are based on detection of the chromophore
p-nitroanilide after cleavage from caspase-specific-labeled
substrates. Colorimetric readings were performed in an automated
microplate reader at an optical density of 405 nm.
Western blotting
Cells were lysed in a buffer containing 62.5 mM
Tris-HCl (pH 6.8), 2% sodium dodecyl sulphate (SDS), 10% glycerol,
6% 2-mercaptoethanol and 0.01% bromophenol blue. Equal amounts of
protein (20 μg) were subjected to electrophoresis on
SDS-polyacrylamide gels followed by transfer to a polyvinylidene
difluoride membrane and probing with the specific antibodies. The
bands were visualized by enhanced chemiluminescence (GE Healthcare,
Buckinghamshire, UK).
Antibodies to cyclin D2, cIAP-2, IκBα and NF-κB
subunits p50, p65, c-Rel, p52 and RelB were purchased from Santa
Cruz Biotechnology (Santa Cruz, CA). Antibodies to Bax, Bcl-2 and
actin were purchased from NeoMarkers (Fremont, CA). Antibodies to
XIAP and cyclin D1 were obtained from Medical & Biological
Laboratories (MBL, Nagoya, Japan). Antibodies to survivin,
caspase-3, -9, Bcl-xL and phospho-IκBα (Ser32 and Ser36)
were purchased from Cell Signaling Technology (Beverly, MA). The
antibody to poly(ADP-ribose) polymerase (PARP) was purchased from
BD Transduction Laboratories (San Jose, CA).
Preparation of nuclear extracts and
electrophoretic mobility shift assay (EMSA)
Cells were cultured and examined for inhibition of
NF-κB after exposure to FXOH for 24 h. Nuclear proteins were
extracted as described by Antalis and Godbolt (10) with modifications, and NF-κB binding
activity to the NF-κB element was examined by EMSA. Briefly, 5 μg
of nuclear extracts were preincubated in a binding buffer
containing 1 μg poly-deoxy-inosinic-deoxycytidylic acid (GE
Healthcare Biosciences), followed by the addition of
32P-labeled oligonucleotide probes containing the NF-κB
element. The mixtures were incubated for 15 min at room
temperature. The DNA protein complexes were separated on 4%
polyacrylamide gels and visualized by autoradiography. The probes
and competitors used were prepared by annealing the sense and
antisense synthetic oligonucleotides as follows: a typical NF-κB
element from the IL-2 receptor α chain (IL-2Rα) gene
(5′-gatcCGGCAGGGGAATCTCCCTCTC-3′) and an AP-1
element of the IL-8 gene (5′-gatcGTGATGACTCAGGTT-3′). The above
underlined sequences represent the NF-κB and AP-1 binding sites,
respectively.
Results
FX and FXOH reduce cell viability of
EBV-immortalized human B-cell lines, BL and HL cell lines
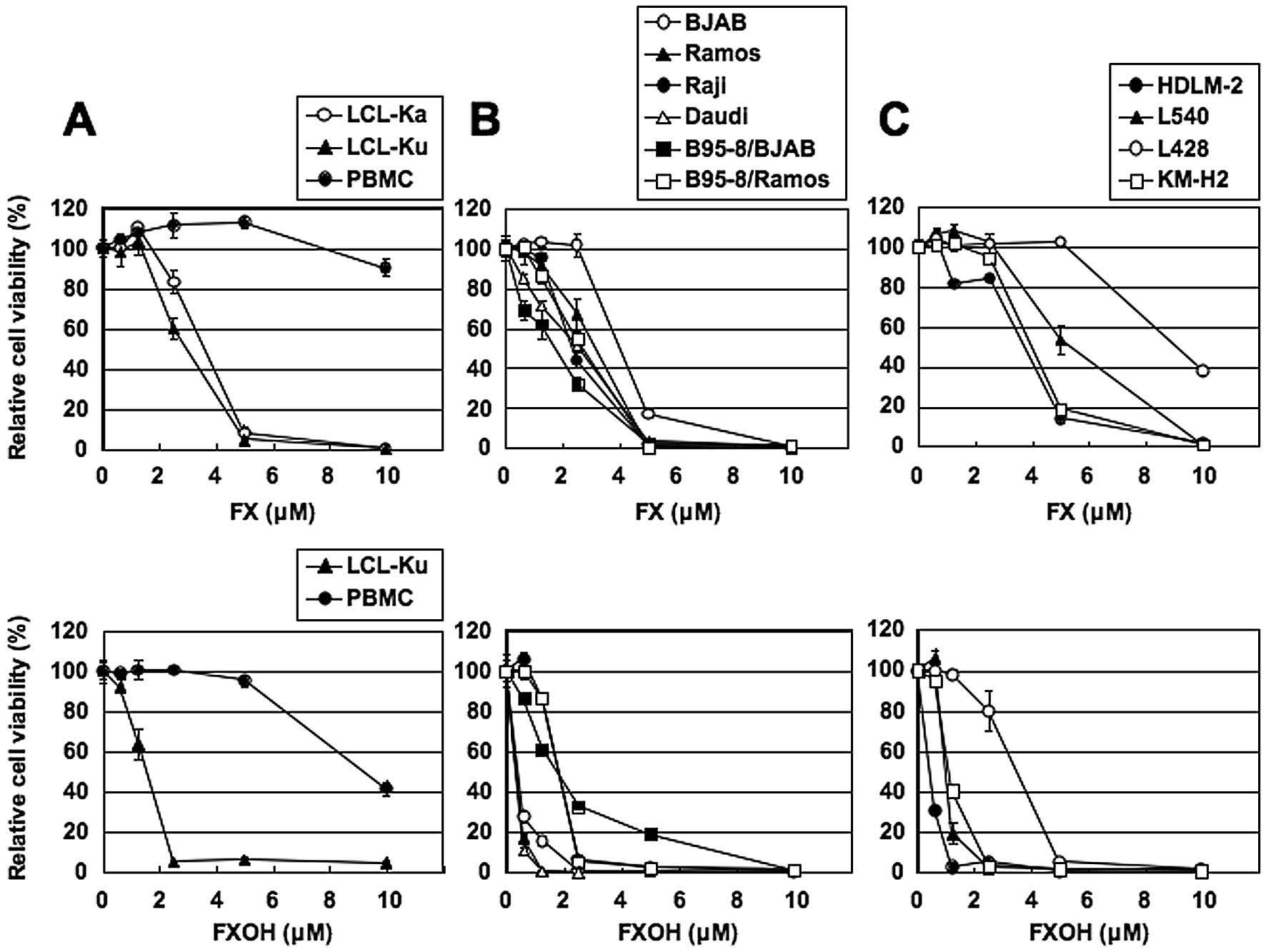
We first examined the effects of FX and FXOH on the
cell viability of EBV-immortalized human B-cell lines. Cells
cultured in the presence of various concentrations of FX or FXOH
for 24 h showed dose-dependent decrease in cell viability in two
EBV-immortalized human B-cell lines, as assessed by WST-8 assay
(Fig. 1A). PBMC from healthy
volunteers were less susceptible to FX and FXOH than the human
B-cell lines. Treatment of EBV-positive and -negative BL cell lines
(Fig. 1B) and HL cell lines
(Fig. 1C) with FX or FXOH also
resulted in lower cell viability. The FXOH-induced suppression of
cell viability was more pronounced than that induced by FX.
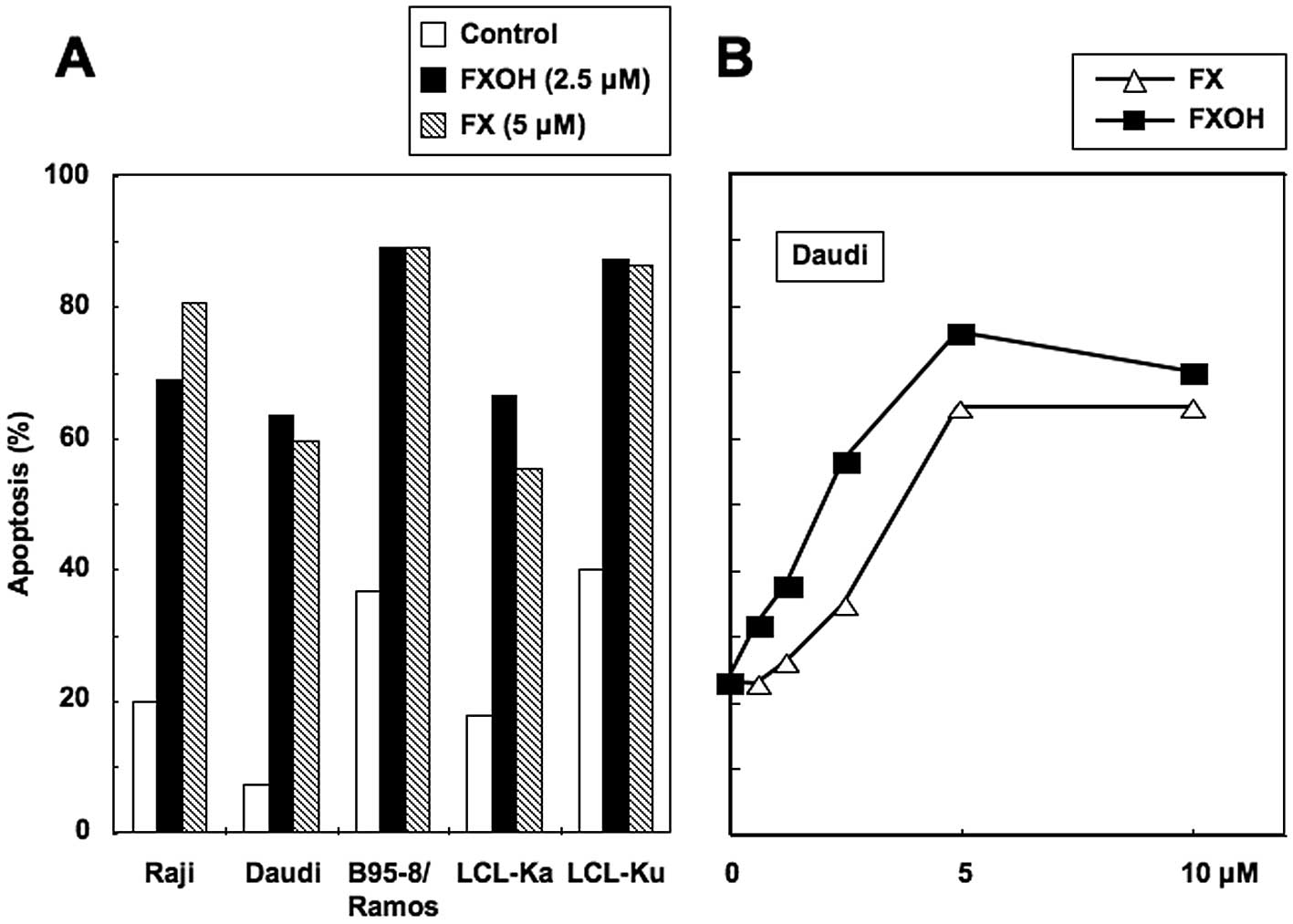
FX and FXOH induce apoptosis
We next examined whether induction of apoptosis
accounted for the reduced cell viability observed in these cell
lines. Cells were treated with 5 μM of FX or 2.5 μM of FXOH and
then probed with the Apo2.7 monoclonal antibody. FX (5 μM) and FXOH
(2.5 μM) increased the proportion of apoptotic cells in
EBV-immortalized human B-cell lines and BL cell lines by similar
levels (Fig. 2A). Exposure of Daudi
cells to FX and FXOH also induced apoptosis in a dose-dependent
manner (Fig. 2B). The FXOH-induced
apoptosis was more pronounced than that induced by FX. These
results indicate that the inhibitory effects of FX and FXOH on the
viability of cell lines reflect the agents’ pro-apoptotic
properties.
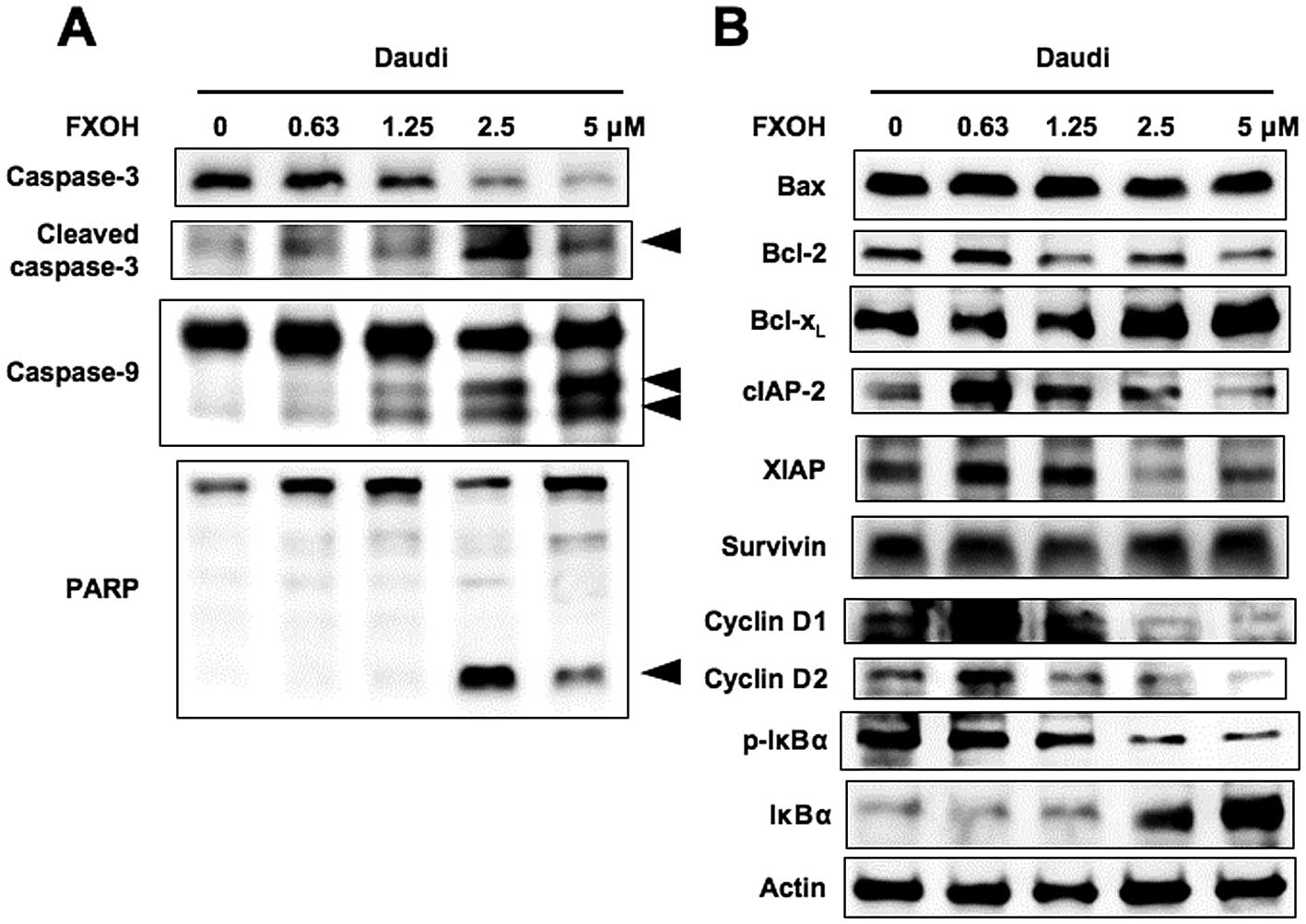
FX- and FXOH-induced apoptosis is
caspase-dependent
We next investigated whether the observed apoptosis
was due to caspase activation. Cell extracts were obtained after
various treatments and processed for immunoblot analysis. As shown
in Fig. 3A, immunoblot analysis
demonstrated the cleaved products of PARP, caspases-3 and -9
induced by FXOH in a dose-dependent manner. The immunoblotting
allowed us to examine the processing of caspases, but did not
indicate whether the cleavage products were enzymatically active.
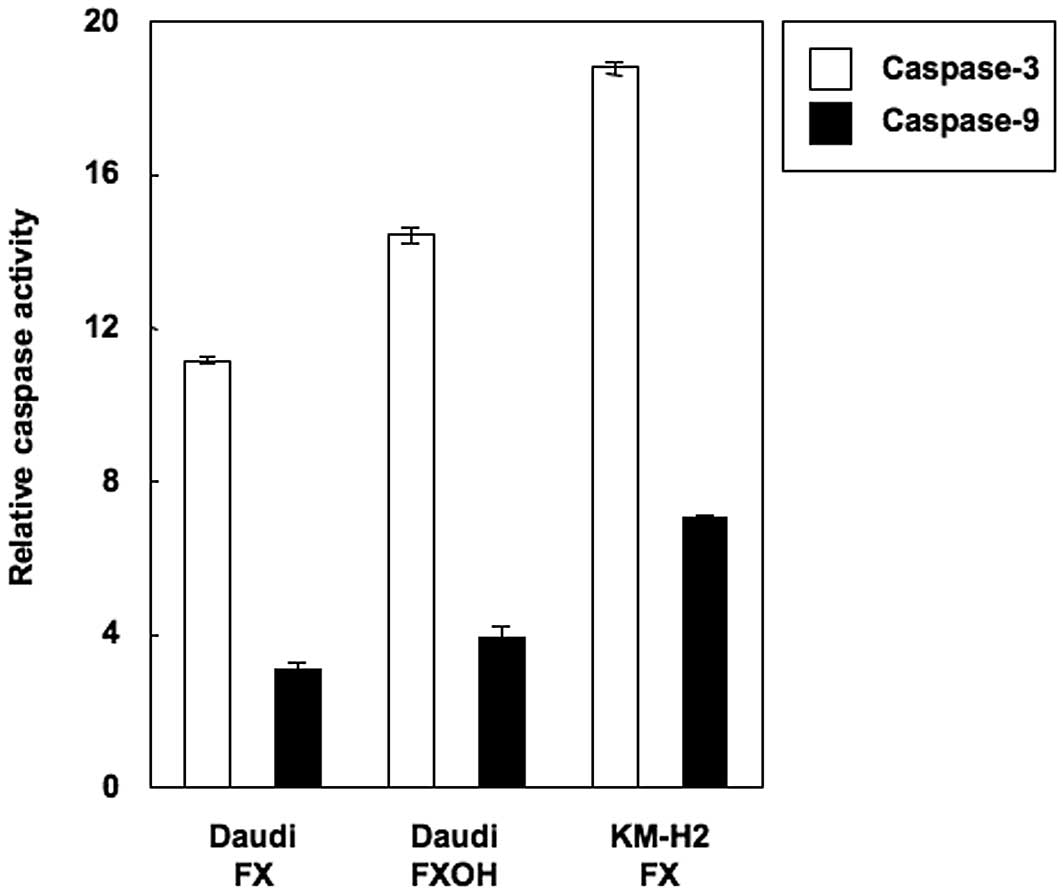
Therefore, caspases-3 and -9 activities were determined by cleavage
of caspase-specific-labeled substrates in colorimetric assays. Both
FX and FXOH activated caspase-3 and -9 in Daudi and KM-H2 cells
(Fig. 4). These results indicated
that caspase activation plays a role in the FX- and FXOH-induced
apoptosis observed in these cell lines.
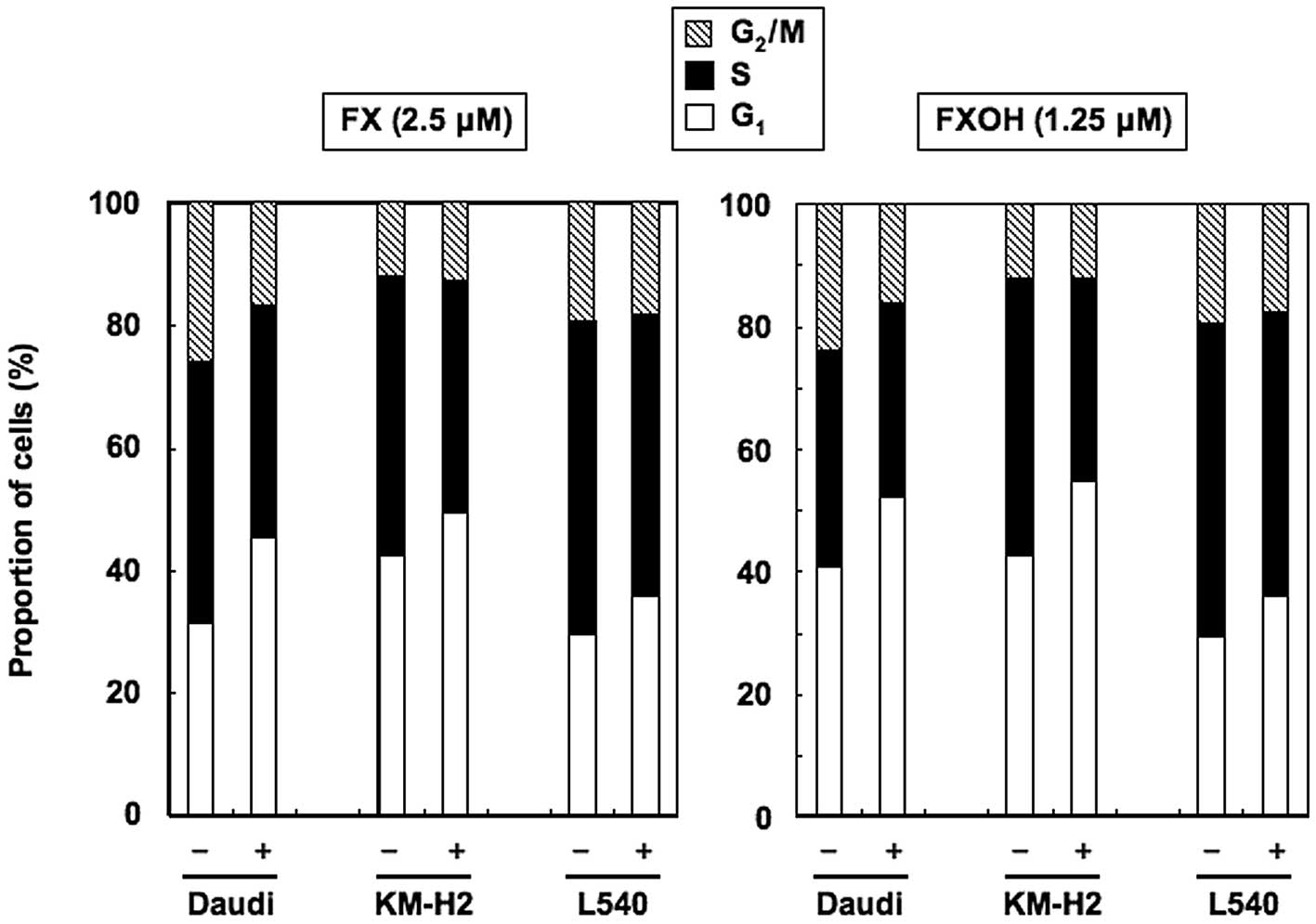
FX and FXOH cause G1 cell
cycle arrest
We next examined the effects of FX and FXOH on cell
cycle-regulatory mechanisms using Daudi, KM-H2 and L540 cells.
Cultivation with 2.5 μM of FX or 1.25 μM of FXOH for 24 h increased
the population of cells in the G1 phase, with reduced
numbers of cells in the S phase, relative to untreated cells
(Fig. 5). Thus, FX and FXOH
suppressed the proliferation of BL and HL cell lines by arresting
the cells in the G1 phase of the cell cycle.
Effects of FXOH on the expression of
apoptosis and cell cycle regulatory proteins
To clarify the molecular mechanisms of FX- and
FXOH-induced inhibition of cell growth and apoptosis, we used
western blot analysis to investigate the mechanism of FXOH-induced
changes in the expression of several intracellular regulators of
cell cycle and apoptosis. As shown in Fig. 3B, FXOH did not alter the expression
levels of anti-apoptotic proteins Bcl-xL and survivin,
or pro-apoptotic protein Bax. In contrast, FXOH downregulated the
expression levels of anti-apoptotic proteins Bcl-2, cIAP-2 and XIAP
in a dose-dependent manner. Furthermore, FXOH downregulated the
expression levels of cell cycle regulatory proteins cyclins D1 and
D2 dose-dependently. The results demonstrated that FXOH-mediated
growth inhibition and apoptosis was associated with reduced
expression of Bcl-2, cIAP-2, XIAP, cyclin D1 and cyclin D2 in the
Daudi cells.
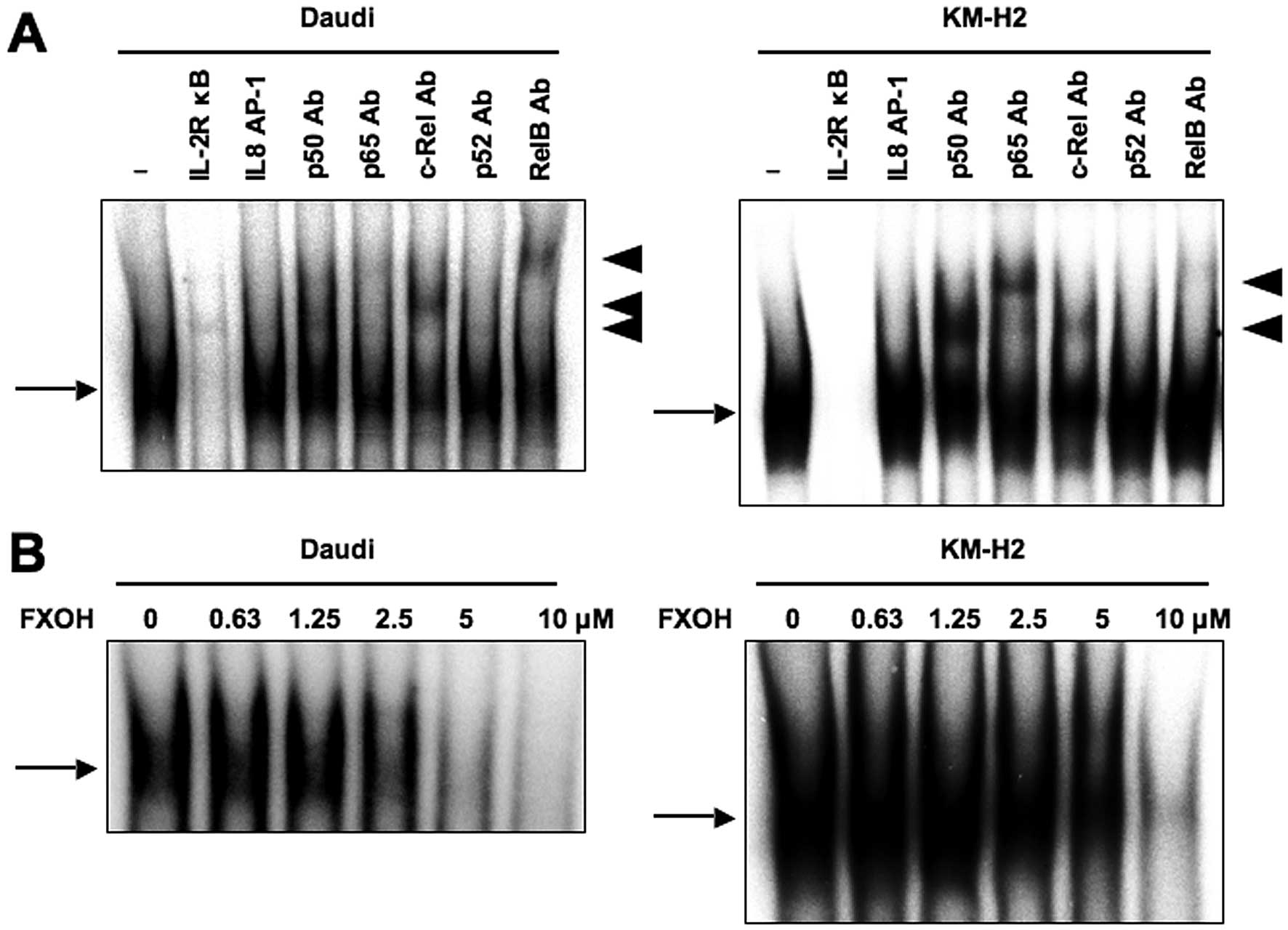
Inhibitory effects of FXOH on NF-κB
activity
Mammalian NF-κB characterizes a family of five
transcription factors: RelA/p65, c-Rel, RelB, NF-κB1 (p50) and
NF-κB2 (11). After activation, the
NF-κB heterodimer is rapidly translocated to the nucleus, where it
activates the transcription of target genes (12). Because Bcl-2, cIAP-2, XIAP, cyclin
D1 and cyclin D2 are NF-κB target genes (12–18),
we examined whether FXOH directly inhibits the NF-κB pathway. To
study the DNA-binding activity of NF-κB, we performed EMSA with
radiolabeled double-stranded NF-κB oligonucleotides and nuclear
extracts from Daudi and KM-H2 cells, and confirmed the constitutive
activation of NF-κB in these cells. Supershift analysis showed that
the NF-κB bands were composed of p50, p65, c-Rel and RelB subunits
(Fig. 6A). We next examined the
effects of FXOH on Daudi and KM-H2 cells in this context by EMSA
and found reduced DNA binding to NF-κB in a dose-dependent manner
(Fig. 6B), suggesting that FXOH
could inhibit the DNA-binding activity of NF-κB.
NF-κB is inactive in the cytosol where it is bound
to IκB, and only becomes active after IκB has been phosphorylated
and subsequently degraded (11).
Immunoblotting showed that in the absence of FXOH, the levels of
phosphorylated IκBα steadily increased in Daudi cells (Fig. 3B). FXOH thus reduced the
phosphorylation and degradation of IκBα in a dose-dependent manner.
These results indicated that FXOH inhibits NF-κB activation by
preventing the degradation of phosphorylated IκBα.
Discussion
The main issue addressed in this study was whether
FX and FXOH have inhibitory activity against human B-cell
malignancies including BL and HL, and the possible mechanisms
underlying such activities. Because of its central role in cell
proliferation and survival, the NF-κB transcription factor has
become an important molecular marker of the malignant
transformation of cells, especially in hematopoietic malignancies
(5,19), and particularly in the pathogenesis
of BL and HL (19–21). The present study therefore
characterized this transcription factor as a good target for the
treatment of BL and HL (19,20).
FX is one of the most biologically active and abundantly found
carotenoids (7). Our findings
strongly suggested that FX and its deacetylated product, FXOH,
inhibit constitutive NF-κB activation via the IκBα phosphorylation
mechanism, resulting in less cell proliferation and increased cell
death by apoptosis. Such results encourage the search for novel
carotenoids for the treatment of cancers characterized by aberrant
regulation of NF-κB.
FX was hydrolyzed to FXOH during uptake by Caco-2
cells, a tissue culture model for studying the absorption of
dietary compounds by human intestinal epithelium (22). Dietary FX is also thought to be
hydrolyzed to FXOH in the mammalian gastrointestinal tract by
digestive enzymes such as lipase and cholesterol esterase before
being absorbed into intestinal cells (22). It was also reported that FXOH is a
major metabolite of dietary FX in humans, implying that the major
compound in the circulation after FX intake is FXOH (23). Therefore, the bioavailability of
FXOH is higher than that of FX in the body. In addition, our study
found that FXOH was more potent in inducing apoptosis than FX in
the BL and HL cell lines. Taken together, it seems likely that most
dietary FX is converted to FXOH, which could then exert a
suppressive effect on cells at concentrations lower than the
effective concentrations of FX used in the present study.
A dose-dependent suppression of NF-κB constitutive
activity by FXOH was also observed in the BL and HL cell lines
studied here. In Daudi and KM-H2 cells, FXOH inhibited NF-κB-DNA
binding by preventing IκBα phosphorylation, which is a crucial
prerequisite to NF-κB-DNA binding. The capacity of FXOH to inhibit
nuclear NF-κB-DNA binding was associated with the inhibition of
NF-κB-regulated gene expression. In this regard, FXOH reduced the
expression of genes involved in cell proliferation (cyclins D1 and
D2) and anti-apoptosis (Bcl-2, cIAP-2 and XIAP). The decrease in
cell viability could be associated with a decreased cell
proliferation and/or an increase in apoptotic cell death, and both
D-type cyclins play a key role in cell proliferation through the
activation of cyclin-dependent kinases (24). Furthermore, both cyclins are
required for the progression of cells from the G1 phase
to the S phase of the cell cycle (24). This scenario could therefore explain
our findings that FXOH induces G1/S cell cycle arrest
and thus inhibits cell proliferation.
We found that both FX and FXOH induced apoptosis of
cells, accompanied by activation of caspases-3 and -9. A possible
mechanism underlying the induction of apoptosis by FXOH could be
its capacity to inhibit NF-κB-regulated anti-apoptotic proteins.
Bcl-2 prevents the process of mitochondrial release of
pro-apoptotic factors, such as cytochrome c (25), and treatment of Daudi cells with
FXOH in this study caused a reduction in Bcl-2 expression as well
as other NF-κB-dependent anti-apoptotic proteins, cIAP-2 and XIAP
(14,15). These IAP proteins are known to
inhibit both extrinsic (i.e., death receptor) and intrinsic (i.e.,
mitochondrial) pathways of apoptosis; cIAP-2 and XIAP directly bind
and inhibit effector caspases, acting downstream of the initiator
caspases. Our results demonstrated that FXOH induces apoptosis
through downregulation of NF-κB-dependent gene products, Bcl-2,
cIAP-2 and XIAP.
Notably, FX and FXOH up to 10 and 5 μM,
respectively, did not inhibit cell viability of PBMC from healthy
volunteers that presented no NF-κB activity. These results together
indicated that inhibition of NF-κB activity and of NF-κB-dependent
expression of cell survival proteins plays a major role in the
pro-apoptotic activity of FX and FXOH in B cell malignancies.
In conclusion, this study demonstrated that FX and
FXOH can inhibit NF-κB constitutive activation in BL and HL cell
lines. The ability of both carotenoids to reduce cell viability
reflects their capacity to decrease cell proliferation by causing
G1 cell cycle arrest and to induce apoptotic cell death.
The observed effects combined with the well-established
pharmacological safety of both carotenoids (7) provide strong rationale for the
potential use of FX and FXOH as new therapeutic agents for patients
with BL and HL.
Acknowledgements
We acknowledge Dr Takeshi Sairenji for providing
B95-8/Ramos and B95-8/BJAB.
References
|
1
|
Epstein MA, Achong BG and Barr YM: Virus
particles in cultured lymphoblasts from Burkitt’s lymphoma. Lancet.
1:702–703. 1964.PubMed/NCBI
|
|
2
|
Taylor GS and Blackbourn DJ: Infectious
agents in human cancers: lessons in immunity and immunomodulation
from gammaherpesviruses EBV and KSHV. Cancer Lett. 305:263–278.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mounier N, Spina M and Gisselbrecht C:
Modern management of non-Hodgkin lymphoma in HIV-infected patients.
Br J Haematol. 136:685–698. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Re D, Thomas RK, Behringer K and Diehl V:
From Hodgkin disease to Hodgkin lymphoma: biologic insights and
therapeutic potential. Blood. 105:4553–4560. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chaturvedi MM, Sung B, Yadav VR, Kannappan
R and Aggarwal BB: NF-κB addiction and its role in cancer: ‘one
size does not fit all’. Oncogene. 30:1615–1630. 2011.
|
|
6
|
Liebowitz D: Epstein-Barr virus and a
cellular signaling pathway in lymphomas from immunosuppressed
patients. N Engl J Med. 338:1413–1421. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peng J, Yuan J-P, Wu C-F and Wang J-H:
Fucoxanthin, a marine carotenoid present in brown seaweeds and
diatoms: metabolism and bioactivities relevant to human health. Mar
Drugs. 9:1806–1828. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim K-N, Heo S-J, Yoon W-J, Kang S-M, Ahn
G, Yi T-H and Jeon Y-J: Fucoxanthin inhibits the inflammatory
response by suppressing the activation of NF-κB and MAPKs in
lipopolysaccharide-induced RAW 264.7 macrophages. Eur J Pharmacol.
649:369–375. 2010.PubMed/NCBI
|
|
9
|
Zhang C, Ao Z, Seth A and Schlossman SF: A
mitochondrial membrane protein defined by a novel monoclonal
antibody is preferentially detected in apoptotic cells. J Immunol.
157:3980–3987. 1996.PubMed/NCBI
|
|
10
|
Antalis TM and Godbolt D: Isolation of
intact nuclei from hematopoietic cell types. Nucleic Acids Res.
19:43011991. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vallabhapurapu S and Karin M: Regulation
and function of NF-κB transcription factors in the immune system.
Annu Rev Immunol. 27:693–733. 2009.
|
|
12
|
Pahl HL: Activators and target genes of
Rel/NF-κB transcription factors. Oncogene. 18:6853–6866. 1999.
|
|
13
|
Grossmann M, O’Reilly LA, Gugasyan R,
Strasser A, Adams JM and Gerondakis S: The anti-apoptotic
activities of Rel and RelA required during B-cell maturation
involve the regulation of Bcl-2 expression. EMBO J. 19:6351–6360.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wäldele K, Silbermann K, Schneider G,
Ruckes T, Cullen BR and Grassmann R: Requirement of the human
T-cell leukemia virus (HTLV-1) tax-stimulated HIAP-1 gene
for the survival of transformed lymphocytes. Blood. 107:4491–4499.
2006.PubMed/NCBI
|
|
15
|
Stehlik C, de Martin R, Kumabashiri I,
Schmid JA, Binder BR and Lipp J: Nuclear factor (NF)-κB-regulated
X-chromosome-linked iap gene expression protects endothelial
cells from tumor necrosis factor α-induced apoptosis. J Exp Med.
188:211–216. 1998.
|
|
16
|
Iwanaga R, Ohtani K, Hayashi T and
Nakamura M: Molecular mechanism of cell cycle progression induced
by the oncogene product Tax of human T-cell leukemia virus type I.
Oncogene. 20:2055–2067. 2001. View Article : Google Scholar
|
|
17
|
Huang Y, Ohtani K, Iwanaga R, Matsumura Y
and Nakamura M: Direct trans-activation of the human cyclin D2 gene
by the oncogene product Tax of human T-cell leukemia virus type I.
Oncogene. 20:1094–1102. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hinz M, Krappmann D, Eichten A, Heder A,
Scheidereit C and Strauss M: NF-κB function in growth control:
regulation of cyclin D1 expression and
G0/G1-to-S-phase transition. Mol Cell Biol.
19:2690–2698. 1999.
|
|
19
|
Jost PJ and Ruland J: Aberrant NF-κB
signaling in lymphoma: mechanisms, consequences, and therapeutic
implications. Blood. 109:2700–2707. 2007.
|
|
20
|
Kimura N, Miyakawa Y, Kohmura K, Umezawa
K, Ikeda Y and Kizaki M: Targeting NF-κB and induction of apoptosis
by novel NF-κB inhibitor dehydroxymethylepoxyquinomicin (DHMEQ) in
Burkitt lymphoma cells. Leuk Res. 31:1529–1535. 2007.
|
|
21
|
Küppers R: Molecular biology of Hodgkin
lymphoma. Hematology Am Soc Hematol Educ Program. 491–496.
2009.
|
|
22
|
Sugawara T, Baskaran V, Tsuzuki W and
Nagao A: Brown algae fucoxanthin is hydrolyzed to fucoxanthinol
during absorption by Caco-2 human intestinal cells and mice. J
Nutr. 132:946–951. 2002.PubMed/NCBI
|
|
23
|
Asai A, Yonekura L and Nagao A: Low
bioavailability of dietary epoxyxanthophylls in humans. Br J Nutr.
100:273–277. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sherr CJ and Roberts JM: Living with or
without cyclins and cyclin-dependent kinases. Genes Dev.
18:2699–2711. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Burstein E and Duckett CS: Dying for
NF-κB? Control of cell death by transcriptional regulation of the
apoptotic machinery. Curr Opin Cell Biol. 15:732–737. 2003.
|