Introduction
Exosomes are microvesicles (40–100 nm in diameter)
of endocytic origin that are released from various cells into the
extracellular space. In 1983, Pan and Johnstone were the first to
show the presence of exosomes as secreted vesicles from sheep
reticulocytes (1). It has been
thought that exosomes are formed by the inward budding of
multivesicular bodies (MVBs) and are released from the cell when
intracellular MVBs fuse with the plasma membrane of the cell
(2). Exosomes are present in cell
culture supernatants as well as in body fluids such as
serum/plasma, urine, amniotic fluid, and ascites fluid (2–6).
Exosomes are composed of a lipid bilayer, and they
contain some proteins such as tetraspanins on the surface of their
membrane (2). Tetraspanins comprise
a large superfamily of cell surface-associated membrane proteins
characterized by four transmembrane domains (7). There are 33 tetraspanin family genes
in the human genome. These membrane domains control the
proliferation and migration of cells through various cell adhesion
and growth factor receptors (7,8). For
example, it has been reported that TSPAN29 (CD9) is associated with
ICAM-1, and TSPAN28 (CD81) is associated with CD19 (8–10).
Among tetraspanin family proteins, CD63, CD9, and CD81 have been
known to be located frequently on the surface of exosomes (2,11–13).
Therefore, the tetraspanin proteins have been used as collection
markers of exosomes. However, the appropriate collection markers of
exosomes in each cell are not known because the components within
exosomes vary according to the type of secreted cell.
The worldwide incidence of colorectal cancer (CRC)
is high, particularly in developed nations. Recently, the
development of useful biomarkers for the early diagnosis of CRC has
been investigated. Microarray analyses have demonstrated that the
differential expressions of mRNAs are observed between the CRC and
normal colon tissues (14–18). In addition, it has been suggested
that a large number of non-coding RNAs including microRNAs and
natural antisense RNAs may be involved in the development of CRC
(19,20). Valadi et al reported that the
intracellular mRNAs and microRNAs are enclosed in exosomes and are
secreted into the extracellular space (21). It has been suggested that RNAs
within exosomes can be used as new diagnostic biomarkers because
RNA components within exosomes differ according to the type and the
physiological state of cells. Taylor et al identified
microRNAs within exosomes to be as useful diagnostic biomarkers of
ovarian cancer (22). Similarly, it
has been suggested that the CRC cell lines secrete exosomes
containing various RNAs and proteins, but the information on the
type of components within exosomes is poor to date.
In the present study, we investigated the
tetraspanin family proteins CD63, CD9, and CD81 as useful
collection markers of exosomes derived from the three CRC cell
lines HCT-15, SW480, and WiDr. In addition, we performed the
detection of mRNAs, microRNAs, and natural antisense RNAs within
exosomes secreted from these CRC cell lines. We also examined
whether exosomes derived from these three CRC cells were
transferred into the hepatoma cell line HepG2 and lung cancer cell
line A549.
Materials and methods
Cell lines and cell culture
The human colorectal cancer cell line WiDr
(JCRB0224), human hepatoma cell line HepG2 (JCRB1054), and human
lung cancer cell line A549 (JCRB0076) were purchased from the
Health Science Research Resources Bank (Osaka, Japan). The human
colorectal cancer cell lines HCT-15 (CCL-225) and SW480 (CCL-228)
were purchased from the American Type Culture Collection (ATCC,
Manassas, VA). WiDr, HepG2, and A549 cells were cultured in
Dulbecco’s minimum essential medium (D-MEM), which was supplemented
with 10% fetal bovine serum (FBS), 100 U/ml penicillin, and 100
μg/ml streptomycin. HCT-15 and SW480 cells were cultured in
RPMI-1640 medium, supplemented with 10% FBS, 100 U/ml penicillin,
and 100 μg/ml streptomycin. Cell cultures were performed at 37°C in
an atmosphere of 5% CO2.
Isolation of exosomes from culture
supernatants
The three CRC cell lines HCT-15, SW480, and WiDr
were plated onto collagen-coated 10-cm dishes at a concentration of
1×106 cells/dish using each of the media described
above. After 48 h, the culture media were discarded, and the cells
were washed three times in phosphate-buffered saline (PBS). Next,
new media supplemented with 10% exosome-free FBS (by
ultracentrifugation overnight) and the antibiotics mentioned above
was added to the cells, and the cells were cultured. After 72 h,
cell culture media were collected and sequential centrifugations
were performed. Cell culture media were centrifuged at 300 × g for
3 min at 4°C to remove floating cells. These supernatants were then
centrifuged at 2,000 × g for 15 min at 4°C, and at 12,000 × g for
35 min at 4°C to remove cell debris. These supernatants were then
passed through a 0.22 μm filter. The filtrates were
ultracentrifuged at 120,000 × g for 3 h at 4°C to collect exosomes
using an Optima TLX Ultracentrifuge (Beckman Coulter, Brea, CA).
Exosomal pellets were washed in PBS and were further
ultracentrifuged at 120,000 × g for 3 h at 4°C. The final exosomal
pellets were stored at −80°C until use or were resuspended in 100
μl of PBS.
Measurement of exosomal protein
concentration
Exosomal pellets were solved in 1% sodium dodecyl
sulfate (SDS) solution and sonicated. The concentrations of
exosomal proteins were measured using a Pierce BCA Protein assay
kit (ThermoFisher Scientific, Wilmington, DE) and a Benchmark
Microplate Reader (Bio-Rad, Hercules, CA) according to the
manufacturer’s instructions.
SDS-polyacrylamide gel electrophoresis
(SDS-PAGE)
Exosomal proteins (10 μg) were mixed in equal volume
with 2× sample buffer (0.125 mol/l Tris-HCl, 4% SDS, 20% glycerol,
10% 2-mercaptoethanol, 0.002% bromophenol blue, pH 6.8). These
samples were boiled at 99°C for 5 min, and immediately cooled on
ice. Electrophoresis of the exosomal proteins was performed using
10% Mini Protean TGX Precast Gels (Bio-Rad), Precision Plus Protein
Dual Color Standards (Bio-Rad), and electrophoresis buffer (25 mM
Tris, 192 mM glycine, 0.1% SDS, pH 8.3) at 200 V and 0.03 A for 30
min.
Exosomal protein staining
Exosomal proteins in the loading gel were stained
with Bio-Safe Coomassie brilliant blue (CBB) G250 Stain (Bio-Rad)
according to the manufacturer’s instructions. Stained gels were
photographed using a GS-800 Calibrated Densitometer (Bio-Rad) and a
Quantity One software (Bio-Rad).
Western blotting
The exosomal proteins of electrophoresed gels were
transferred to Hybond-P membranes (GE Healthcare, Buckinghamshire,
UK) using transfer buffer (25 mM Tris, 192 mM glycine, 0.01% SDS,
20% methanol, pH 8.3) at 100 V and 0.2 A for 60 min. Membranes were
blocked in TBST buffer (25 mM Tris-HCl, 150 mM NaCl, 0.05%
Tween-20, pH 7.2) containing 5% non-fat milk for 60 min at room
temperature. After blocking, membranes were incubated with each
primary antibody in TBST buffer containing 5% non-fat milk
overnight at 4°C. As primary antibodies, 1:200 dilution each of
anti-CD63 (sc-15363; Santa Cruz Biotechnology, Santa Cruz, CA),
anti-CD9 (ab92726; Abcam, Cambridge, UK), and CD81 (ab79559; Abcam)
were used. Membranes were washed five times for 5 min with TBST
buffer and incubated for 60 min at room temperature with
anti-rabbit horseradish peroxidase (HRP)-linked antibody (GE
Healthcare) or anti-mouse HRP-linked antibody (GE Healthcare) at
1:5000 dilution prepared in TBST buffer containing 5% non-fat milk.
Membranes were washed five times for 5 min with TBST buffer. Bound
antibodies were visualized by chemiluminescence using an ECL Plus
Western Blotting detection system (GE Healthcare). Luminescent
images were analyzed using a ChemiDoc XRS (Bio-Rad) and the
Quantity One software (Bio-Rad).
Isolation and detection of exosomal
RNAs
Exosomal RNAs were extracted using exosomal pellets
and an ISOGEN II (Nippon Gene, Tokyo, Japan) according to the
manufacturer’s instructions. Concentrations of exosomal RNAs
extracted were examined using a Quant-iT RiboGreen RNA Reagent and
kit (Life Technologies, Carlsbad, CA) and a Fluoroskan Ascent
(ThermoFisher Scientific) according to the manufacturer’s
instructions.
To confirm the size of the exosomal RNAs derived
from the three CRC cell lines, exosomal RNAs were electrophoresed
using an Agilent 2100 Bioanalyzer (Agilent Technologies, Foster
City, CA) and an Agilent RNA 6000 Pico kit (Agilent Technologies)
according to the manufacturer’s instructions. One nanogram of each
of the exosomal RNAs were used for analyses.
Reverse transcription-polymerase chain
reaction (RT-PCR)
To obtain cDNAs derived from the mRNAs within
exosomes, exosomal RNAs (1 ng) were used for the reverse
transcription reaction. Reverse transcription reactions were
performed using High Capacity cDNA Reverse Transcriptase kits (Life
Technologies) according to the manufacturer’s instructions. These
synthesized cDNAs were subjected to PCR in a 45 μl reaction mixture
containing 1× buffer, 1.5 mM MgCl2, 0.1 mM of each dNTP,
0.025 U/μl of BioTaq HS DNA polymerase (Nippon Genetics, Tokyo,
Japan), 0.5 μM of the primer pairs (Table I) and the cDNA template. PCRs were
performed using a Veriti 96 Well Thermal Cycler (Life Technologies)
under the following conditions: 7 min at 95°C, followed by 40
cycles each of 95°C for 30 sec, 60°C for 30 sec, and 72°C for 30
sec and a final elongation step at 72°C for 5 min.
 | Table IPrimer sequences for detection of
mRNAs. |
Table I
Primer sequences for detection of
mRNAs.
| Primer name | Sequences | Size (mer) | PCR products
(bp) |
|---|
| ACTB | F:
5′-CCAACCGCGAGAAGATGA-3′ | 18 | 97 |
| R:
5′-CCAGAGGCGTACAGGGATAG-3′ | 20 | |
| GAPDH | F:
5′-AGCCACATCGCTCAGACAC-3′ | 19 | 66 |
| R:
5′-GCCCAATACGACCAAATCC-3′ | 19 | |
| RPL13A | F:
5′-GCATGAGCTTGCTGTTGTACAC-3′ | 22 | 90 |
| R:
5′-CATGGGCGATGCCTGTAAC-3′ | 19 | |
| HMBS | F:
5′-GAGAAGTCCAAGCAACAGC-3′ | 19 | 61 |
| R:
5′-CCTTCAGAACTGGTTTATTAGTAGG-3′ | 25 | |
| B2M | F:
5′-CATGGTTGTGGTTAATCTG-3′ | 19 | 72 |
| R:
5′-GAGATAACACATCAAGTTTTATG-3′ | 23 | |
| TBP | F:
5′-CAGTATTGCAGGACAGAATATATG-3′ | 24 | 83 |
| R:
5′-TTGTACAGAGTACTCTGAAGAAAG-3′ | 24 | |
In order to synthesize cDNAs derived from microRNAs
within exosomes, reverse transcription reactions were performed
using exosomal RNAs (1 ng), a TaqMan MicroRNA Reverse Transcription
kit (Life Technologies), and TaqMan MicroRNA Assays (Life
Technologies) according to the manufacturer’s instructions
(Table II). PCRs were performed
using a TaqMan Universal PCR Master mix II (Life Technologies) and
a StepOne Plus Real-Time PCR system (Life Technologies) under the
following conditions: 10 min at 95°C, followed by 40 cycles each of
95°C for 15 sec and 60°C for 60 sec.
| Table IIThe list of TaqMan microRNA
assays. |
Table II
The list of TaqMan microRNA
assays.
| Assay name | Assay ID |
|---|
| hsa-miR-21 | 000397 |
| hsa-miR-34a | 000426 |
| hsa-miR-143 | 002249 |
| hsa-miR-192 | 000491 |
| hsa-miR-215 | 000518 |
| hsa-miR-221 | 000524 |
| U6 snRNA | 001973 |
To synthesize cDNAs derived from natural antisense
RNAs, reverse transcription reactions were performed using exosomal
RNAs (1 ng), the forward primers shown in Table III, and an AMV Reverse
Transcriptase (Promega, Madison, WI) (19,23,24).
PCRs were performed using cDNAs derived from natural antisense
RNAs, a Power SYBR Green Master mix (Life Technologies), the primer
pairs described in Table III, and
the StepOne Plus Real-Time PCR system (Life Technologies) under the
following conditions: 10 min at 95°C, followed by 40 cycles each of
95°C for 15 sec and 60°C for 60 sec.
| Table IIIPrimer sequences for detection of
natural antisense RNAs. |
Table III
Primer sequences for detection of
natural antisense RNAs.
| Primer name | Sequences | Size (mer) | PCR products
(bp) |
|---|
| LRRC24 | F:
5′-TACGTTCGCACAGCTAGAGG-3′ | 20 | 60 |
| R:
5′-TTGATGACGAACATCTCGTGGC-3′ | 22 | |
| MDM2 | F:
5′-AGACAACCAATTCAAATGATTGTGC-3′ | 25 | 60 |
| R:
5′-CTCTTATAGACAGGTCAACTAGG-3′ | 23 | |
| CDKN1A | F:
5′-TTGATTAGCAGCGGAACAAGG-3′ | 21 | 60 |
| R:
5′-TCCATAGCCTCTACTGCCA-3′ | 19 | |
The PCR products obtained above were electrophoresed
using 4% agarose gels. Detection of amplified fragments was
achieved by ethidium bromide staining using the ChemiDoc XRS
(Bio-Rad) and the Quantity One software (Bio-Rad).
PKH67 labeling of exosomes and their
uptake into HepG2 and A549 cells
Exosomes derived from the three CRC cell lines were
isolated as described above. They were then washed by being
resuspended in PBS and ultracentrifuged at 120,000 × g for 3 h at
4°C. Exosomes were labeled using PKH67 Fluorescent Cell Linker kits
(Sigma-Aldrich, St. Louis, MO) according to the manufacturer’s
instructions, with minor modifications in the washing process. The
washed exosomal pellets from the 100 ml culture media were
resuspended in 700 μl of Diluent C (exosomal solution). PKH67 dye
(1 μl) was diluted in 250 μl of Diluent C (PKH67 solution). Then,
250 μl exosomal solution and 250 μl PKH67 solution were mixed in a
4.7 ml centrifugation tube. Samples were mixed gently for 4 min,
and 4.2 ml of 1% BSA was added to bind the excess PKH67 dye.
PKH67-labeled exosomes were ultracentrifuged at 120,000 × g for 3 h
at 4°C using the Optima TLX Ultracentrifuge (Beckman Coulter).
Exosomal pellets were washed three times in PBS by
ultracentrifugation. Finally, PKH67-labeled exosomes were
resuspended in D-MEM or RPMI-1640 medium. As the negative controls,
no PKH67 control and no exosome control were prepared. Exosomes
were collected by ultracentrifugation without PKH67 dye, and then
D-MEM or RPMI-1640 medium were added to the centrifuged tubes (no
PKH67 control). After PKH67 dye was washed by ultracentrifugation
without exosomes, the supernatant was discarded and media described
above were added to the centrifuged tubes (no exosome control).
To examine the uptake of exosomes into other cells,
HepG2 or A549 cells were plated in 8-well chamber slides
(1×104 cells/well) using each medium. After 24 h, the
slides were washed three times in PBS, and each medium containing
PKH67-labeled exosomes or negative control samples was added into
each well. Cells were cultured for 48 h at 37°C in an atmosphere of
5% CO2. After incubation, the slides were washed three
times in PBS, and 4% paraformaldehyde solution then added to the
slides. These were fixed for 10 min at room temperature. The slides
were washed three times in PBS again. Nucleus staining was
performed using a ProLong Gold Antifade Reagent with DAPI (Life
Technologies) and the slide was covered with cover glass. Finally,
the cells were visualized under a confocal laser scanning
microscope LSM710 (Carl Zeiss, Oberkochen, Germany) under the same
conditions.
Results
Detection of tetraspanins in exosomes
derived from the three CRC cell lines
It is known that exosomes contain various proteins,
especially those of the tetraspanin family. CD63, CD9, and CD81
proteins were examined in exosomes derived from the three CRC cell
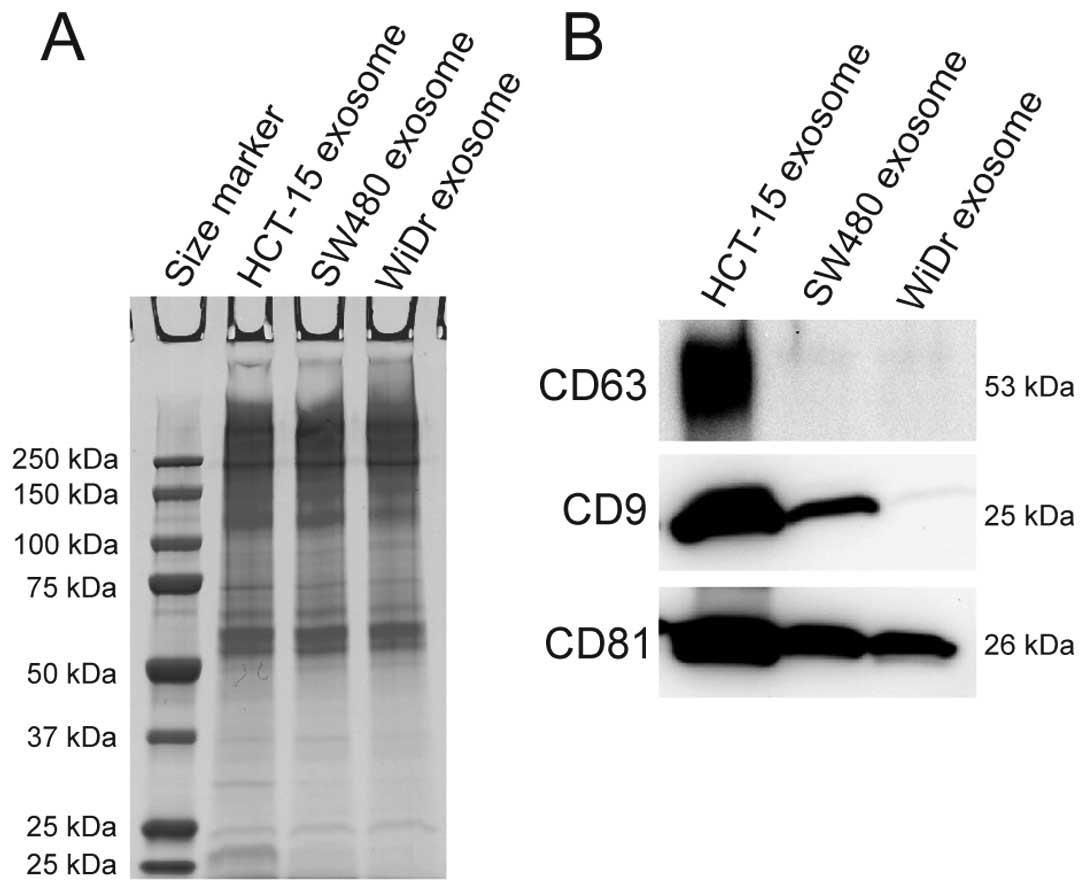
lines HCT-15, SW480, and WiDr by western blotting. As shown in
Fig. 1A, CBB staining showed that
exosomes derived from the three CRC cell lines contained various
proteins, and these proteins were similar in sizes with the
exosomes of the three CRC cell lines. This finding suggests that
the exosomes between the three CRC cell lines may have similar
proteins. As shown in Fig. 1B, CD63
was detected in exosomes derived from HCT-15 cells; CD9 was
detected in exosomes derived from HCT-15 and SW480 cells; and CD81
was detected in exosomes derived from HCT-15, SW480, and WiDr cells
(Fig. 1B). These results indicate
that exosomes derived from all the CRC cell lines examined contain
CD81. Therefore, CD81 can be a collection marker of exosomes
derived from the three CRC cell lines.
Detection of exosomal RNAs in the three
CRC cell lines
To confirm whether exosomes derived from the three
CRC cell lines contained RNAs, the exosomal RNAs were extracted
using the ISOGEN II as described in Materials and methods. The size
of exosomal RNAs was examined using the Agilent 2100 Bioanalyzer as
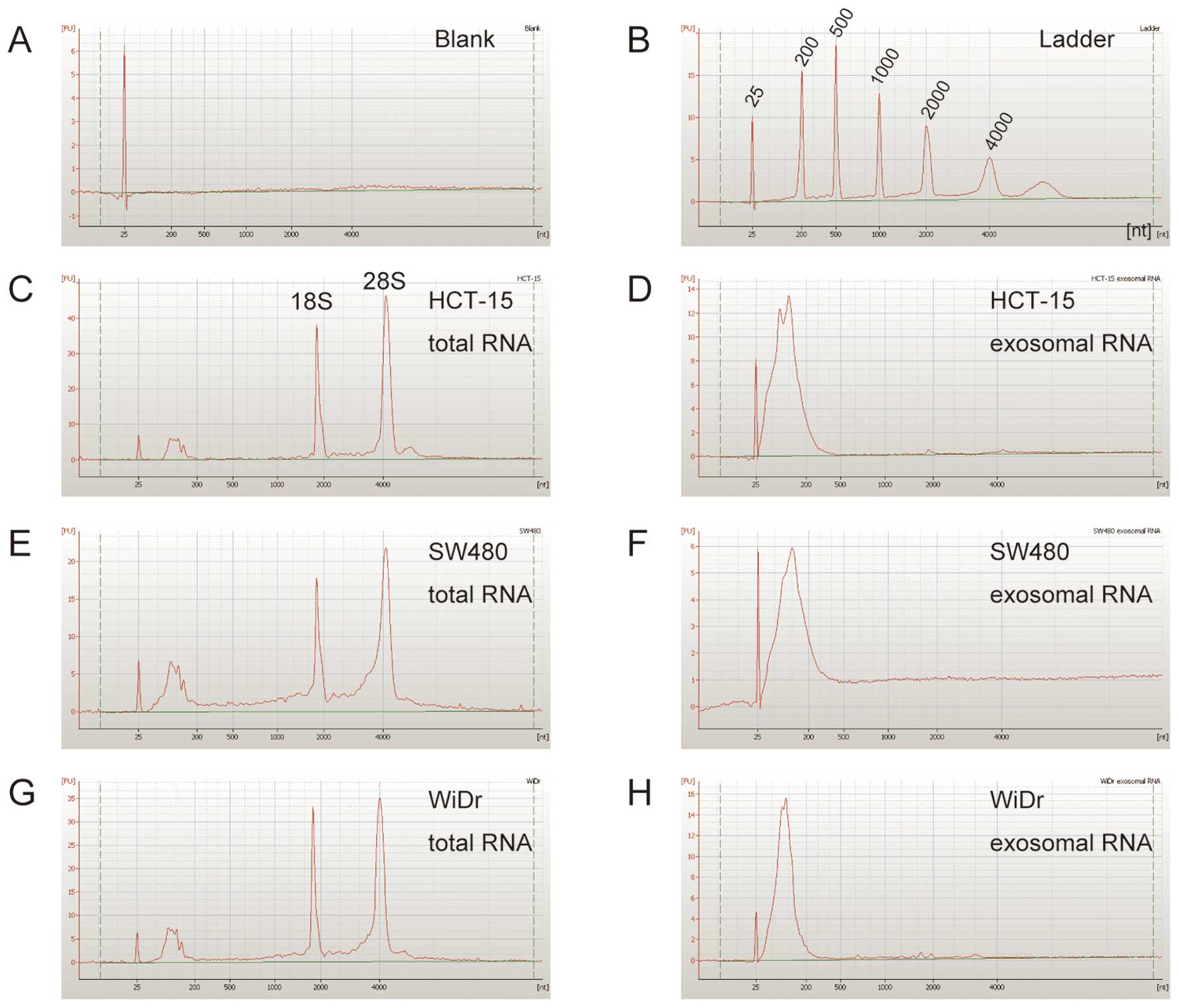
described in the Materials and methods. As shown in Fig. 2, exosomes derived from HCT-15,
SW480, and WiDr cells contained a large number of small RNAs, and
their small RNAs were rarely detected in the size range: 25–200
nucleotides (nt).
In order to detect the mRNAs in exosomal RNAs,
ACTB, GAPDH, RPL13A, HMBS, B2M, and TBP mRNA of
housekeeping genes were examined by RT-PCR as described in
Materials and methods. As shown in Fig.
3A, all the mRNAs examined above were detected from the
exosomes derived from HCT-15 and SW480 cells; and the mRNAs, except
for TBP mRNA, were detected from the exosomes derived from
WiDr cells.
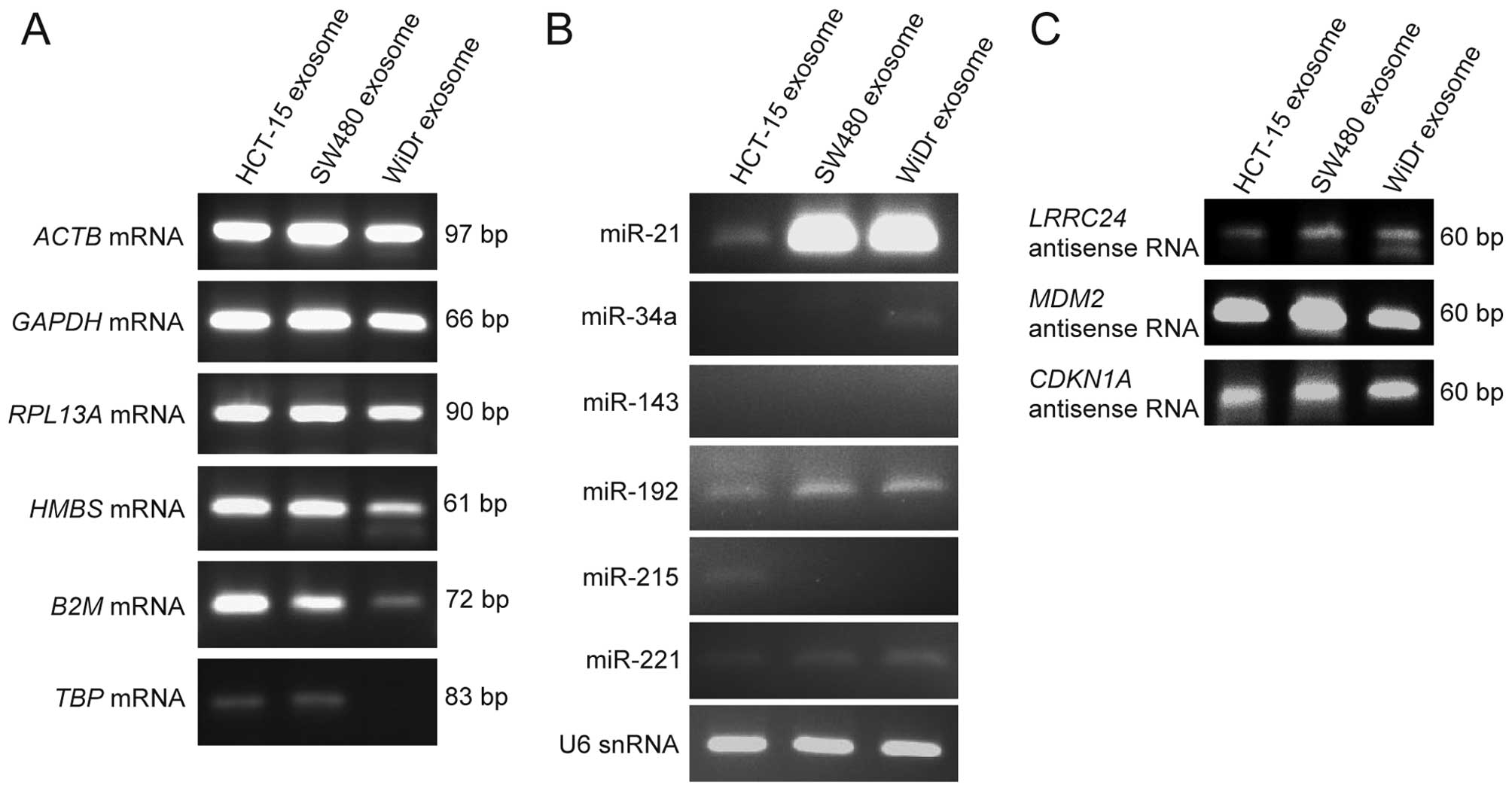 | Figure 3Identification of mRNAs, microRNAs,
and natural antisense RNAs within exosomes derived from the three
CRC cell lines by RT-PCR. The mRNAs, microRNAs, and natural
antisense RNAs within exosomes derived from the three CRC cell
lines were detected by RT-PCR as described in Materials and
methods. (A) Detection of ACTB, GAPDH, RPL13A, HMBS, B2M,
and TBP mRNAs. (B) Detection of miR-21, miR-34a, miR-143,
miR-192, miR-215 and miR-221 as microRNAs and of U6 snRNA as small
RNA. (C) Detection of the natural antisense RNAs of LRRC24,
MDM2 and CDKN1A genes. |
de Krijger et al stated that microRNAs such
as miR-21, miR-34a, miR-143, miR-192, miR-215, and miR-221 were
involved in CRC metastasis (25).
Therefore, in order to detect their microRNAs within exosomes,
RT-PCR of their microRNAs and U6 snRNA were performed as described
in Materials and methods. As shown in Fig. 3B, miR-21, miR-192, miR-221, and U6
snRNA were detected from the exosomes of the three CRC cell lines,
especially miR-21 was strongly detected from the exosomes of SW480
and WiDr cells; miR-34a was weakly detected from the exosomes of
WiDr; miR-215 was weakly detected from the exosomes of HCT-15; and
miR-143 was not detected from the exosomes of the three CRC cell
lines.
Kohno et al reported that natural antisense
RNAs of LRRC24 gene was upregulated in the human CRC
tissues, compared with the normal colon tissues (19). In addition, we detected the natural
antisense RNAs of MDM2 and CDKN1A genes in human cell
lines in a previous study (23).
Therefore, in order to examine whether natural antisense RNAs are
present within exosomes, the natural antisense RNAs of LRRC24,
MDM2 and CDKN1A genes were examined by strand-specific
RT-PCR as described in Materials and methods. As shown in Fig. 3C, all the natural antisense RNAs
examined above were detected in the exosomes of the three CRC cell
lines. We discovered that their natural antisense RNAs were within
exosomes for the first time in the present study. These results
indicate that the mRNAs, microRNAs, and natural antisense RNAs
exist within the exosomes derived from the three CRC cell
lines.
HepG2 and A549 cells take up exosomes
derived from the three CRC cell lines
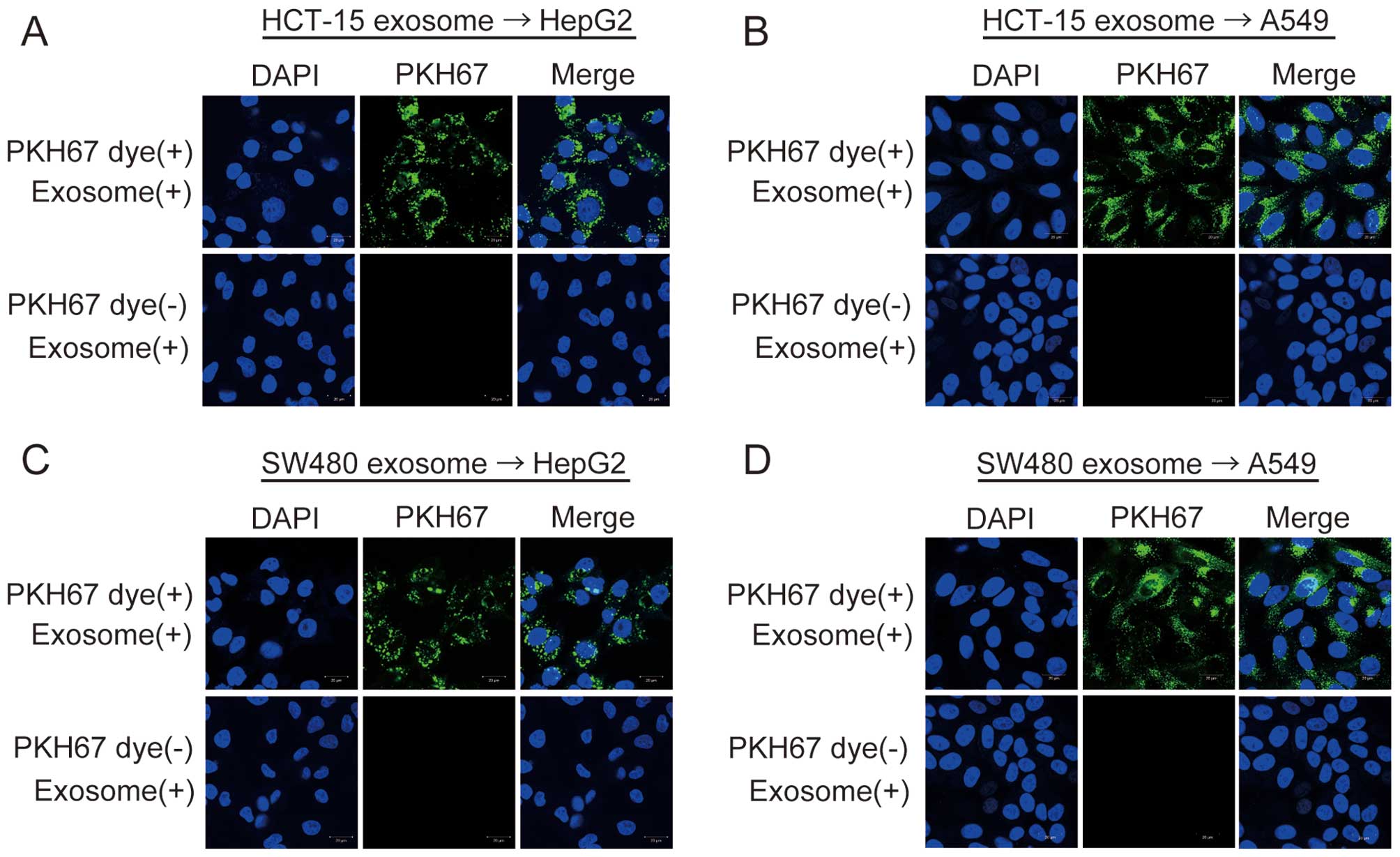
In order to examine whether exosomes derived from
HCT-15, SW480, and WiDr cells could be taken up in HepG2 or A549
cells, their exosomes were labeled with PKH67 dye (green
fluorescence) as described in Materials and methods. When
PKH67-labeled exosomes were added to the culture media of HepG2 or
A549 cells, green fluorescence was observed in HepG2 and A549 cells
using the confocal laser scanning microscopy (Fig. 4). Green fluorescence was not
observed in the two negative controls (Fig. 4). These results show that exosomes
derived from the three CRC cell lines are taken up into HepG2 and
A549 cells, and that exosomes containing various RNAs can shuttle
between cells.
Discussion
In the present study, we showed that CD81 was an
appropriate collection marker of exosomes derived from HCT-15,
SW480, and WiDr cell lines using western blotting. We also
demonstrated that exosomes derived from the three CRC cell lines
contained the mRNAs, microRNAs, and natural antisense RNAs. Herein,
we discovered natural antisense RNAs of LRRC24, MDM2, and
CDKN1A genes within exosomes for the first time. Finally, we
showed that exosomes derived from the three CRC cell lines were
transferred into HepG2 and A549 cells.
Recently, the identification of exosomal RNA and
protein components have been performed by a number of researchers.
Valadi et al discovered various mRNAs and microRNAs within
exosomes derived from mouse and human mast cell lines by microarray
analyses for the first time (21).
Montecalvo et al detected microRNAs within exosomes derived
from immature and mature dendritic cells by microarray analyses
(26). Those studies showed that
RNA components within exosomes differed according to cell type.
Herein, we examined the types of exosomal marker proteins and RNAs
within exosomes derived from the three CRC cell lines. By detection
of the tetraspanin family within exosomes derived from the three
CRC cell lines by western blotting, CD63, CD9, and CD81 were
detected within exosomes derived from HCT-15 cells; CD9 and CD81
were detected within exosomes derived from SW480 cells; and CD81
was detected within exosomes derived from WiDr cells (Fig. 1B). These findings indicate that CD81
is the most useful exosome collection marker of these CRC cell
lines. Interestingly, Petersen et al found that exosome
production increased significantly after CD63-knockdown in B
lymphoblastoid cells (27). A type
of tetraspanin family and the quantity of such a family may be
involved in exosome production.
We identified various RNAs within exosomes. As shown
in Fig. 3, miR-21 was strongly
detected within exosomes derived from the CRC cell lines,
especially SW480 and WiDr cells. MiR-21 is the most common and
highly upregulated microRNA in the CRC cell lines, and it inhibits
the translation of PDCD4 protein (28). Overexpression of miR-21 induces the
invasion, intravasation, and metastasis of the CRC cell lines by
downregulation of PDCD4 protein (28). Taken together, these findings
suggest that miR-21 within exosomes derived from the CRC cell lines
may regulate the expression of target genes into HepG2 and A549
cells, and may induce various functions (e.g., invasion,
intravasation, and metastasis) in those cells.
Natural antisense RNAs, which are transcribed from
the DNA strand opposite to the sense strand, have been demonstrated
to be involved in the control of gene expression in eukaryotes.
Kohno et al discovered that the natural antisense RNA of
LRRC24 gene was upregulated in the human CRC tissues,
compared with normal colon tissues (19). Furthermore, we detected the natural
antisense RNAs of MDM2 and CDKN1A genes in human B
lymphoblastic cells in a previous study, and showed that their
natural antisense RNAs were in the cytoplasm (23). In the present study, it was
clarified that their intracellular natural antisense RNAs were
enclosed within exosomes. Their natural antisense RNAs within
exosomes derived from the CRC cell lines may also regulate the
expressions of target genes such as MDM2 and CDKN1A
mRNAs into the cytoplasm of HepG2 and A549 cells.
It has been reported that exosomes contain various
RNAs and are transferred into other cells. Valadi et al
demonstrated that mouse proteins could be synthesized into human
HMC-1 cells when exosomes derived from the mouse mast cell line
MC/9 were added to the supernatants of the cultured human mast cell
line HMC-1 (21). Zhang et
al demonstrated that microvesicles derived from the human
monocytic cells THP-1 delivered miR-150 into the human endothelial
cells HMEC-1, and that the elevated level of exogenous miR-150
effectively reduced c-Myb expression and enhanced cell migration in
HMEC-1 cells (29). Those findings
suggest that the mRNAs and microRNAs within exosomes are delivered
into different cells and can function in those cells. We revealed
that exosomes containing the mRNAs, microRNAs, and natural
antisense RNAs were transferred into HepG2 and A549 cells. The
transferred mRNAs into those recipient cells may be translated to
proteins, or the transferred microRNAs and natural antisense RNAs
may regulate the expression of target mRNAs. Thereby, their
exosomal RNAs may induce the development and malignancy of tumor
cells.
It has been reported that ceramide plays an
important role in the external secretion of exosomes (30,31).
Ceramide is synthesized from sphingomyelin by the action of neutral
sphingomyelinase 2 (nSMase2), and is a major component of exosomes.
Kosaka et al reported that nSMase2 was involved in the
external secretion of microRNAs within exosomes (32). It is thought that exosomes are
secreted from a cell by the vesicular transport pathway involved in
MVBs (2,11), but the detailed mechanisms are not
completely understood. Further studies are required to: understand
the mechanisms of exosome production via MVBs of the CRC cell
lines; identify comprehensively the RNA species in the CRC cell
lines; and examine whether the transferred RNAs in the CRC cell
lines function in recipient cells such as HepG2 and A549 cells.
In conclusion, we revealed that exosomes derived
from the three CRC cell lines contained the mRNAs, microRNAs, and
natural antisense RNAs, and were delivered into HepG2 and A549
cells. These important and novel findings indicate that exosomes
containing various RNAs can shuttle between cells, and may regulate
the gene expression into recipient cells.
Acknowledgements
This study was supported in part by a grant from
KAKENHI (no. 23790613), Grant-in-Aid for Young Scientists (B). This
work also received support from the Suzuken Memorial Foundation
(no. 11-076).
References
|
1
|
Pan BT and Johnstone RM: Fate of the
transferrin receptor during maturation of sheep reticulocytes in
vitro: selective externalization of the receptor. Cell. 33:967–978.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mathivanan S, Ji H and Simpson RJ:
Exosomes: extracellular organelles important in intercellular
communication. J Proteomics. 73:1907–1920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Michael A, Bajracharya SD, Yuen PS, et al:
Exosomes from human saliva as a source of microRNA biomarkers. Oral
Dis. 16:34–38. 2010.PubMed/NCBI
|
|
4
|
Moon PG, You S, Lee JE, Hwang D and Baek
MC: Urinary exosomes and proteomics. Mass Spectrom Rev.
30:1185–1202. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brase JC, Wuttig D, Kuner R and Sultmann
H: Serum microRNAs as non-invasive biomarkers for cancer. Mol
Cancer. 9:3062010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lasser C, Alikhani VS, Ekstrom K, et al:
Human saliva, plasma and breast milk exosomes contain RNA: uptake
by macrophages. J Transl Med. 9:92011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Romanska HM and Berditchevski F:
Tetraspanins in human epithelial malignancies. J Pathol. 223:4–14.
2011. View Article : Google Scholar
|
|
8
|
Yanez-Mo M, Barreiro O, Gordon-Alonso M,
Sala-Valdes M and Sanchez-Madrid F: Tetraspanin-enriched
microdomains: a functional unit in cell plasma membranes. Trends
Cell Biol. 19:434–446. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hemler ME: Tetraspanin functions and
associated microdomains. Nat Rev Mol Cell Biol. 6:801–811. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Levy S and Shoham T: Protein-protein
interactions in the tetraspanin web. Physiology. 20:218–224. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pols MS and Klumperman J: Trafficking and
function of the tetraspanin CD63. Exp Cell Res. 315:1584–1592.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Escola JM, Kleijmeer MJ, Stoorvogel W,
Griffith JM, Yoshie O and Geuze HJ: Selective enrichment of
tetraspan proteins on the internal vesicles of multivesicular
endosomes and on exosomes secreted by human B-lymphocytes. J Biol
Chem. 273:20121–20127. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mathivanan S and Simpson RJ: ExoCarta: A
compendium of exosomal proteins and RNA. Proteomics. 9:4997–5000.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Croner RS, Foertsch T, Brueckl WM, et al:
Common denominator genes that distinguish colorectal carcinoma from
normal mucosa. Int J Colorectal Dis. 20:353–362. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ohmachi T, Tanaka F, Mimori K, Inoue H,
Yanaga K and Mori M: Clinical significance of TROP2 expression in
colorectal cancer. Clin Cancer Res. 12:3057–3063. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bertucci F, Salas S, Eysteries S, et al:
Gene expression profiling of colon cancer by DNA microarrays and
correlation with histoclinical parameters. Oncogene. 23:1377–1391.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bianchini M, Levy E, Zucchini C, et al:
Comparative study of gene expression by cDNA microarray in human
colorectal cancer tissues and normal mucosa. Int J Oncol. 29:83–94.
2006.PubMed/NCBI
|
|
18
|
Birkenkamp-Demtroder K, Christensen LL,
Olesen SH, et al: Gene expression in colorectal cancer. Cancer Res.
62:4352–4363. 2002.PubMed/NCBI
|
|
19
|
Kohno K, Chiba M, Murata S, et al:
Identification of natural antisense transcripts involved in human
colorectal cancer development. Int J Oncol. 37:1425–1432.
2010.PubMed/NCBI
|
|
20
|
Negrini M, Nicoloso MS and Calin GA:
MicroRNAs and cancer - new paradigms in molecular oncology. Curr
Opin Cell Biol. 21:470–479. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Valadi H, Ekstrom K, Bossios A, Sjostrand
M, Lee JJ and Lotvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chiba M, Miura T, Kasai K, et al:
Identification of up-regulated and down-regulated cis-natural
antisense transcripts in the human B lymphoblastic cell line IM-9
after X-ray irradiation. Mol Med Rep. 5:1151–1157. 2012.PubMed/NCBI
|
|
24
|
Chiba M, Kiyosawa H, Hiraiwa N, Ohkohchi N
and Yasue H: Existence of Pink1 antisense RNAs in mouse and their
localization. Cytogenet Genome Res. 126:259–270. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
de Krijger I, Mekenkamp LJ, Punt CJ and
Nagtegaal ID: MicroRNAs in colorectal cancer metastasis. J Pathol.
224:438–447. 2011.
|
|
26
|
Montecalvo A, Larregina AT, Shufesky WJ,
et al: Mechanism of transfer of functional microRNAs between mouse
dendritic cells via exosomes. Blood. 119:756–766. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Petersen SH, Odintsova E, Haigh TA,
Rickinson AB, Taylor GS and Berditchevski F: The role of
tetraspanin CD63 in antigen presentation via MHC class II. Eur J
Immunol. 41:2556–2561. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Asangani IA, Rasheed SA, Nikolova DA, et
al: MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor
suppressor Pdcd4 and stimulates invasion, intravasation and
metastasis in colorectal cancer. Oncogene. 27:2128–2136. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Y, Liu D, Chen X, et al: Secreted
monocytic miR-150 enhances targeted endothelial cell migration. Mol
Cell. 39:133–144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Marsh M and van Meer G: Cell biology. No
ESCRTs for exosomes. Science. 319:1191–1192. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Trajkovic K, Hsu C, Chiantia S, et al:
Ceramide triggers budding of exosome vesicles into multivesicular
endosomes. Science. 319:1244–1247. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kosaka N, Iguchi H, Yoshioka Y, Takeshita
F, Matsuki Y and Ochiya T: Secretory mechanisms and intercellular
transfer of microRNAs in living cells. J Biol Chem.
285:17442–17452. 2010. View Article : Google Scholar : PubMed/NCBI
|