Introduction
In human inflammatory bowel disease (IBD), the
subtypes of ulcerative colitis (UC) and Crohn’s disease are
chronic, relapsing, and remitting conditions that characterized by
diarrhea, bloody stools, abdominal pain and weight loss. Currently,
UC is believed to be caused by multiple factors, including genetic
and environmental factors. Histologically, UC is characterized by
crypt abscesses, crypt distortion and loss, ulceration, and by the
infiltration of large numbers of neutrophils, monocytes and
lymphocytes. IBD affects at least 1 in 1,000 people in Western
countries (1). Most importantly, UC
is associated with an increased risk of colorectal cancer (2). Despite many investigations into IBD,
its etiology and pathogenic mechanisms remain poorly
understood.
Experimental animal models of colitis have been
developed in order to investigate the underlying physiologic
mechanisms and to improve medical therapies for IBD. In the most
commonly-used models, colitis is induced by administering sulfated
polysaccharides such as dextran sulfate sodium (DSS) or carrageenan
(3). In general, the history of the
development of DSS-induced colitis has paralleled the history of
antipepsin agents. Since the mucosal protection theory for gastric
mucus was suggested, many antipepsin agents have been developed.
More recently, amylopectin sulfate was developed in the United
States, and DSS was developed in Sweden. At the same time, the
major side effects of antipepsin agents were discovered, and they
were found to induce intestinal and cecal ulceration in animal
experiments (4). Hence, DSS has
been used for the production of experimental colitis. DSS is a
heparin-like polysaccharide consisting of anhydro-D-glucose
(α-1,6-glucosidic linkage) moieties and sulfate. DSS contains
approximately 17% sulfur, with up to three sulfate groups per
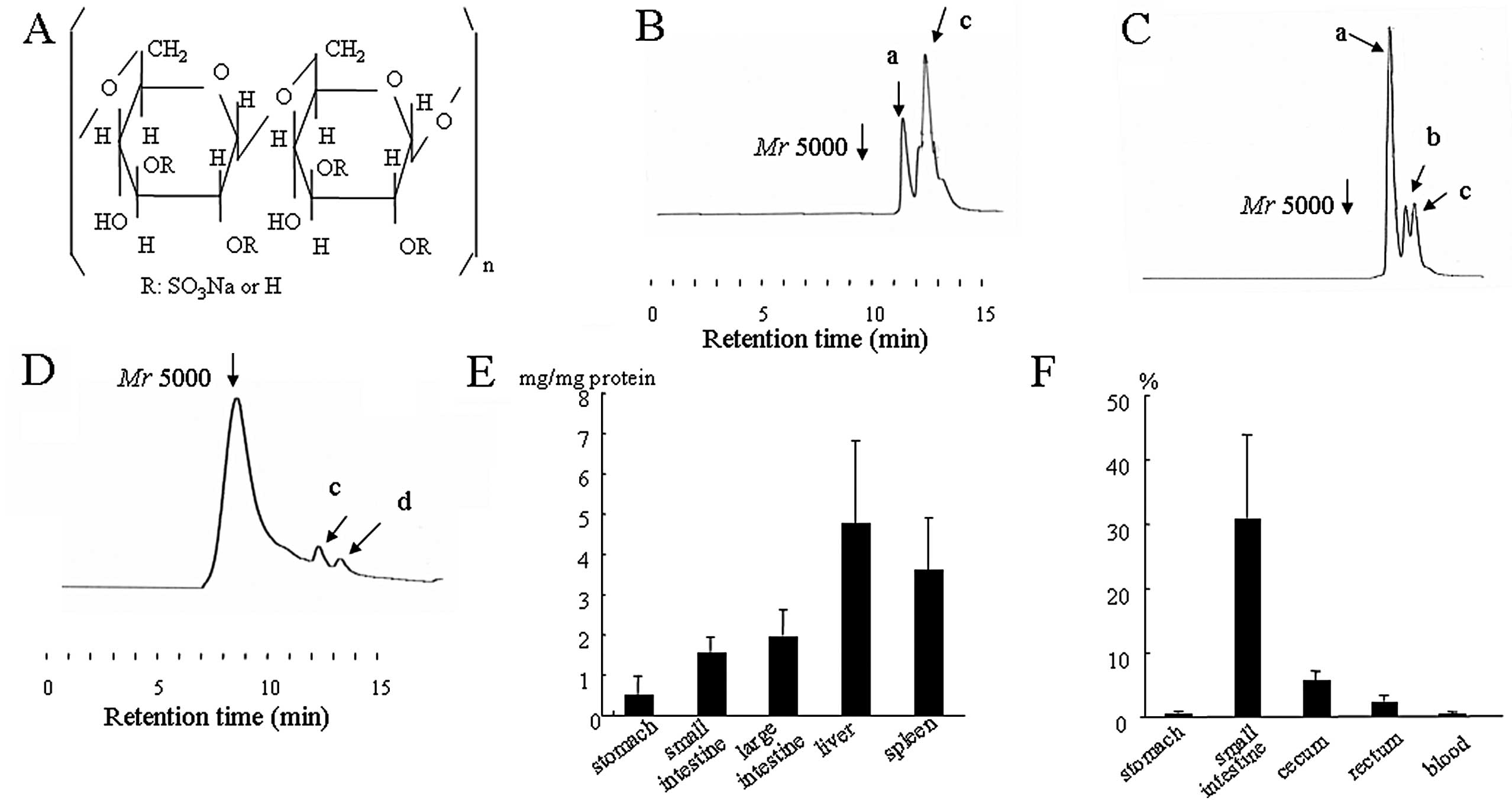
glucose molecule (5) (Fig. 1A). DSS induced-colitis exhibits some
of the clinical and histological features of UC. For example,
DSS-induced colitis starts from the rectum and subsequently spreads
to the anal side. Histologically, DSS-induced colitis is
characterized by crypt distortion and loss, ulceration, and the
infiltration of large numbers of inflammatory cells, which is
similar to UC. In addition, the dysplasia and adenocarcinomas
observed in UC patients are also recognized in this mode (6,7). It
has been reported that the development of colitis is dependent on
the molecular mass and sulfation of the DSS, in addition to the
dosage and duration of the administration (8,9).
However, it is unknown how DSS actually induces colitis. Therefore,
an investigation into the pathogenic factors in this colitis model
may help elucidate the mechanisms responsible for human UC. Despite
many studies, the pathogenic factors in DSS-induced colitis remain
unknown, although DSS has been often used as a UC model. Even with
respect to the metabolism of DSS in the gut lumen, there have been
no detailed studies. Therefore, the main purpose of this study was
to investigate the metabolism of DSS in the gut lumen. In the
present study, we used 2-aminopyridine labeling and HPLC for
detecting DSS in biological materials according to our previous
reports (10,11). Additionally, we also propose a
plausible mechanism for DSS-induced colitis based on our findings
for the unique metabolism of DSS.
Materials and methods
Chemicals
Mr 5000 and 500k DSS and 2-aminopyridine were
obtained from Wako Pure Chemical Industries, Ltd. (Osaka, Japan).
Mr 8000, 10000 DSS and D-glucose 3-sulfate were obtained from Sigma
Chemical Co. (St. Louis, MO). Mr 2500 DSS was obtained from Tokyo
Chemical Industry Co., Ltd. (Tokyo, Japan).
Labeling of DSS
The pyridylamination of the reducing termini of
sugar chains has been useful for the structural analysis and
metabolic studies on N- or O-glycosidically-linked
sugar chains. The labeling of each molecular mass of DSS using
2-aminopyridine was carried out according to our previous report
(10,11). We obtained pure pyridylamino-DSS
(PA-DSS) of each molecular mass, and the PA-D-glucose 3-sulfate as
a PA-monomer.
HPLC conditions
Since PA-DSS is strongly negatively charged in
aqueous solution, a strong interaction between PA-DSS and the
stationary phase is present. Therefore, 0.2 M phosphate buffer at
pH 3.0 was used as the mobile phase according to our previous
report (10,11). The mobile phases were delivered
isocratically at a flow rate of 1.0 ml/min. We used an HPLC LC6A
apparatus (Shimadzu, Kyoto, Japan). The retention time of DSS was
determined by gel filtration chromatography on a Cosmosil 5Diol-120
Packed column (7.5×300 mm; Nacalai Tesque Inc., Kyoto, Japan) at
60°C. For the detection, the fluorescence detector RF-535
(Shimadzu) was used at excitation and emission wavelengths of 320
nm and 400 nm, respectively.
Animals
Specific pathogen-free male BALB/cA Jcl mice, 6-week
old, were purchased from Nippon Clea Inc. (Tokyo, Japan). They were
housed in a room with controlled temperature (20–22°C), humidity
(50–60%) and a preset light-dark cycle (12 h:12 h). This study was
carried out in strict accordance with the recommendations in the
Guide for the Care and Use of Laboratory Animals of the National
Institutes of Health. The protocol was approved by the Animal Care
and Use Committee of the Shiga University of Medical Science
(Permit no: 2006-7-6). All surgery was performed under sodium
pentobarbital anesthesia, and all efforts were made to minimize
suffering.
Mouse PA-DSS-induced colitis
At the beginning of the experiment, the mice were
fed the standard diet (MF, Oriental Yeast Co,. Ltd., Tokyo, Japan)
containing 5% (w/w of diet) Mr 5000 or 2500 PA-DSS for 8 days
(n=5). During the experimental period, body weight was measured
every other day. On the final day, blood samples were collected by
cardiac puncture. The contents of the gut lumen were removed,
diluted adequately by PBS, centrifuged to remove any insoluble
material (14,000 rpm, for 20 min), and then the supernatants were
used for the HPLC analysis. Each organ was resected, irrigated with
chilled PBS, placed in 0.5 ml of a hexadecyltrimethylammonium
bromide solution (0.5%, w/w), homogenized, sonicated and subjected
to three rapid cycles of freezing and thawing. The samples were
then centrifuged and the supernatants were used for HPLC analysis.
The blood samples were centrifuged, and the supernatants were used
for HPLC analysis. On the other hand, a specimen at 2 cm distance
from the anal margin (middle colon) was removed, frozen and then
cut into 5 μm sections. The sections were observed under a
fluorescence microscope, or stained with hematoxylin and eosin
(H&E) to observe under light microscopy. The mucosal damage was
determined according to a previously described method (12). Briefly, the following three
parameters were used: surface epithelium loss, crypt destruction
and inflammatory cell infiltration into the mucosa. A score of 0–4
was assigned to each of three parameters according to the extent
and the severity of the changes. The sum of the scores from the
three parameters represented the mucosal damage score in each
animal.
Caco-2 cell culture
Caco-2 (a human colon cancer cell line) cells were
purchased from the American Type Culture Collection (Rockville,
USA). The cells were cultured in Dulbecco’s modified Eagle’s
minimum essential medium (DMEM, pH 7.4) supplemented with 25 mM
glucose, 10% inactivated fetal bovine serum (FBS), 1%
penicillin-streptomycin and 1% non-essential amino acid solution at
37°C in a humidified 5% CO2 atmosphere.
Findings of Caco-2 cells using
microscopy
We reported that the Mr 5000 DSS was depolymerized
to Mr 1800, and ~70% of the sulfate groups were depleted from each
DSS molecule following autoclave treatment (at 115°C for 15 min,
1.7 atm) (13). Therefore, filter
sterilization is recommended. The Caco-2 cells were incubated on
culture slides. After the cells reached confluence, they were
incubated with 3% Mr 5000 PA-DSS in a growth medium for 24 h. Next,
the cells were washed with PBS and incubated with growth medium
alone for 24 h. Then, the cells were fixed with ethanol, permeated
with acetone, stained with a 0.05% toluidine blue solution for 20
min and observed under light microscopy. The cells were also
washed, fixed in 1% glutaraldehyde and observed under a
fluorescence microscope (Labophot-2, Nikon, Tokyo, Japan).
Metabolism of PA-DSS in Caco-2 cells
When confluent and allowed to mature on permeable
inserts, the Caco-2 monolayer formed tight junctions and attained
many of the morphological and functional characteristics of the
intestinal epithelium with normal barrier function (14). We investigated the metabolism of Mr
5000 PA-DSS during passage through this monolayer. Briefly, the
Caco-2 cells were plated on Millicell-HA 0.4 μm permeable filters
(12-mm) (Millipore, Bedford, MA). The transepithelial electrical
resistance (TEER) of these cells was measured with an electrical
resistance system, Millicell-ERS (Millipore), according to our
previous methods (15). Cells with
stable TEER readings >500 Ωcm2 were used (4–5 weeks
post plating). A 250 μl aliquot of 1% Mr 5000 PA-DSS solution was
inserted inside the porous filters, and 500 μl of medium outside of
the porous filters was collected at the indicated times and
analyzed by HPLC.
Cell cycle arrest and apoptosis in
DSS-induced colitis and Caco-2 cells
We observed each sample using immunohistochemical
staining. Anti-mouse or anti-human Ki-67 antibodies (Dako
Cytomation, Denmark) and the TdT-FragEL™ DNA Fragmentation
Detection kit (Calbiochem, USA) were used to analyze the cell cycle
and apoptosis, respectively according to the manuals, provided by
the manufacturers.
Statistical analysis
The results are presented as means ± SEM. The
variance was analyzed by the F test. Subsequently, the Student’s
t-test for unpaired values was performed to compare the means of
the normally distributed data. The Mann-Whitney U test was also
performed to compare the means of non-parametric or abnormally
distributed data. Differences were regarded as statistically
significant if the P-values were <0.05.
Results
Metabolism of Mr 5000 PA-DSS
First, we investigated the metabolism of DSS in the
gut lumen using 2-aminopyridine labeling and HPLC systems. No
PA-DSS was recognized in the esophageal mucosa. Surprisingly, Mr
5000 PA-DSS was depolymerized in the gastric contents (Fig. 1B). The two main peaks were
speculated to be ~Mr 2000 (peak a) and 1200 (peak c), according to
the relationship between the retention time and the molecular
masses of Mr 5000, 10000 PA-DSS and PA-D-glucose-3-sulfate,
respectively. The molecular mass distribution in the gastric mucosa
was the same as in the gastric contents. Both in the contents and
mucosa of the small intestine, the three main peaks were speculated
to be approximately Mr 2000 (peak a), 1500 (peak b) and 1200 (peak
c), respectively (Fig. 1C).
However, the main peaks in the contents of the cecum were
speculated to be Mr 5000 (Fig. 1D).
Small peaks of Mr 1200 (peak c) and 750 (peak d) were also
recognized. In the cecal mucosa, the main peaks were approximately
Mr 2000, 1500, 1200 and 750. The distribution in the liver, spleen
and blood was the same as in the small intestine. Remarkably, the
maximum molecular mass of PA-DSS in each organ was below Mr 2000.
Fig. 1E and F show the
quantification of PA-DSS in each organ and its contents,
respectively. The levels of PA-DSS were the highest in the liver
and spleen, and the concentration of PA-DSS in the contents of the
small intestine reached ~30% (w/v). In the cecum, the concentration
was ~5% (w/w). Interestingly, this concentration was the same as in
the diet.
Mr 5000 PA-DSS-induced colitis
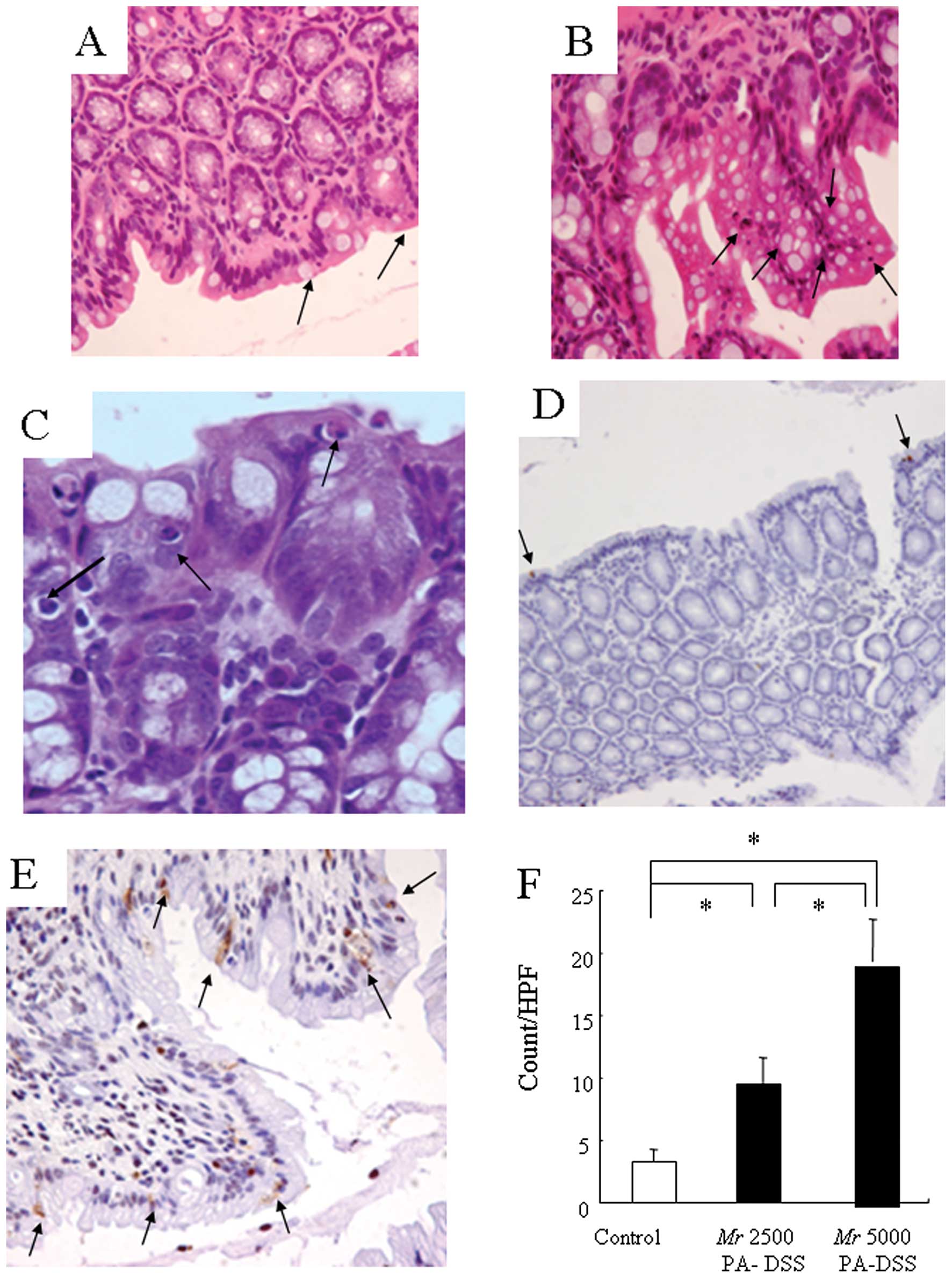
Next, we observed DSS-induced colitis both
macroscopically and microscopically. After the Mr 5000 PA-DSS
administration, diarrhea occurred on Days 3–4. Macroscopic
examination of the colon revealed hyperemia, erosions and
occasional tiny blood clots in the distal colon on Day 5. Using
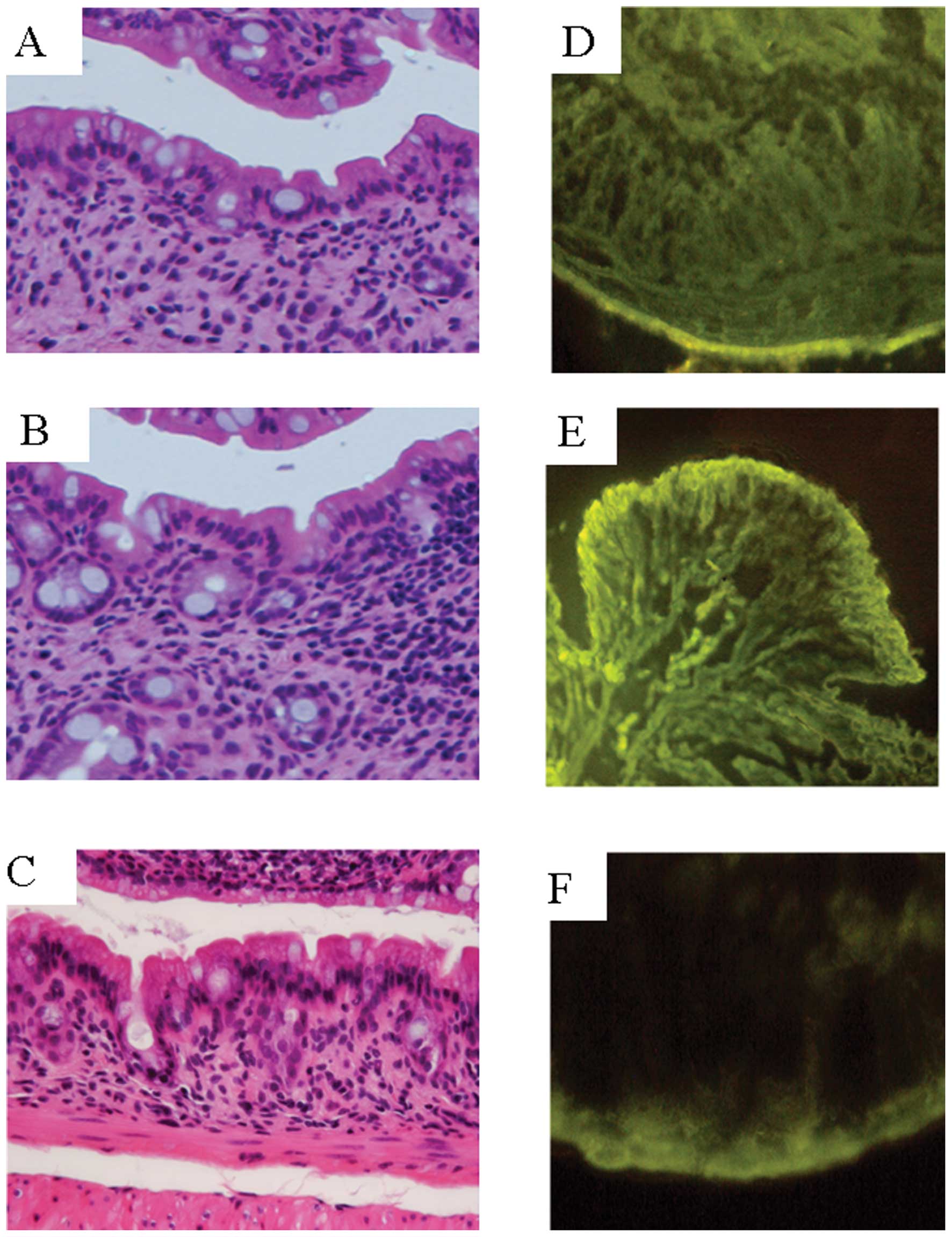
H&E staining, there was obvious evidence of inflammatory cell
infiltration into the mucosa and submucosa. Crypt shortening,
entire crypt loss, and earthenware mortar-like deformity of the
crypts, surface epithelial loss and mucosal edema were also evident
(Fig. 2A-C). Fig. 2D and E show the fluorescence signals
on Day 8. The signal increased significantly, and was localized
throughout the entire intestinal wall as compared to the control
mice, which had very weak signals localized mainly to the serosa
(Fig. 2F). These results suggest
that PA-DSS permeated the intestinal epithelium in a form under Mr
2000. In addition, PA-DSS subsequently invaded into the mucosa,
submucosa, and muscle layers.
Mouse Mr 2500 PA-DSS-induced colitis
We next investigated whether this difference in the
molecular mass distribution of PA-DSS in the lumen explain the fact
that Mr 5000 PA-DSS mainly induces inflammation in the colon, but
not in the stomach or small intestine. Specifically, we examined
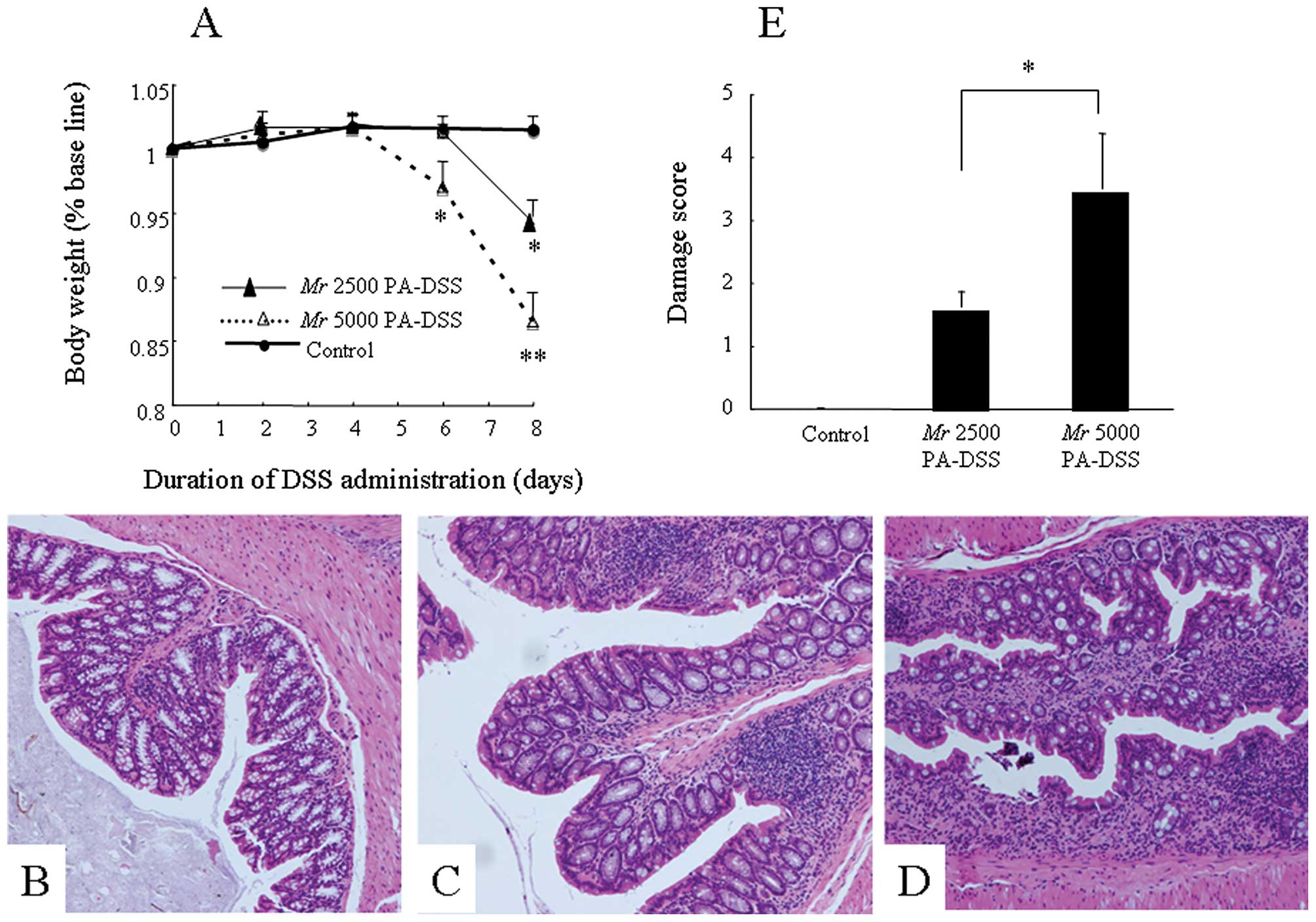
whether Mr 2500 PA-DSS could induce colitis or not. After the Mr
2500 PA-DSS administration, the appearance of diarrhea was delayed
until Days 6–7. The body weight loss was also less than that in
mice fed Mr 5000 PA-DSS (Fig. 3A).
This difference between Mr 2500 and 5000 DSS was statistically
significant on Day 6. Mr 5000 PA-DSS-induced colitis was
histologically more severe than Mr 2500 PA-DSS-induced colitis
(Fig. 3B, control; C, Mr 2500
PA-DSS; and D, Mr 5000 PA-DSS). This difference between both groups
was statistically significant (Fig.
3E). In this section, Mr 2500 PA-DSS was also depolymerized in
the stomach. Interestingly, the majority of the PA-DSS in the cecal
contents was also Mr 2500 DSS, at a concentration of ~5% (w/w),
which was the same as that in the diet.
Cell cycle arrest in PA-DSS-induced
colitis
Subsequently, we examined how DSS contributes to the
induction of colitis. We previously found that DSS strongly and
rapidly inhibited the intracellular energy metabolism in Caco-2
cells (16). Therefore, we focused
on the dynamics of the epithelial cells, especially the cell cycle
status in this section.
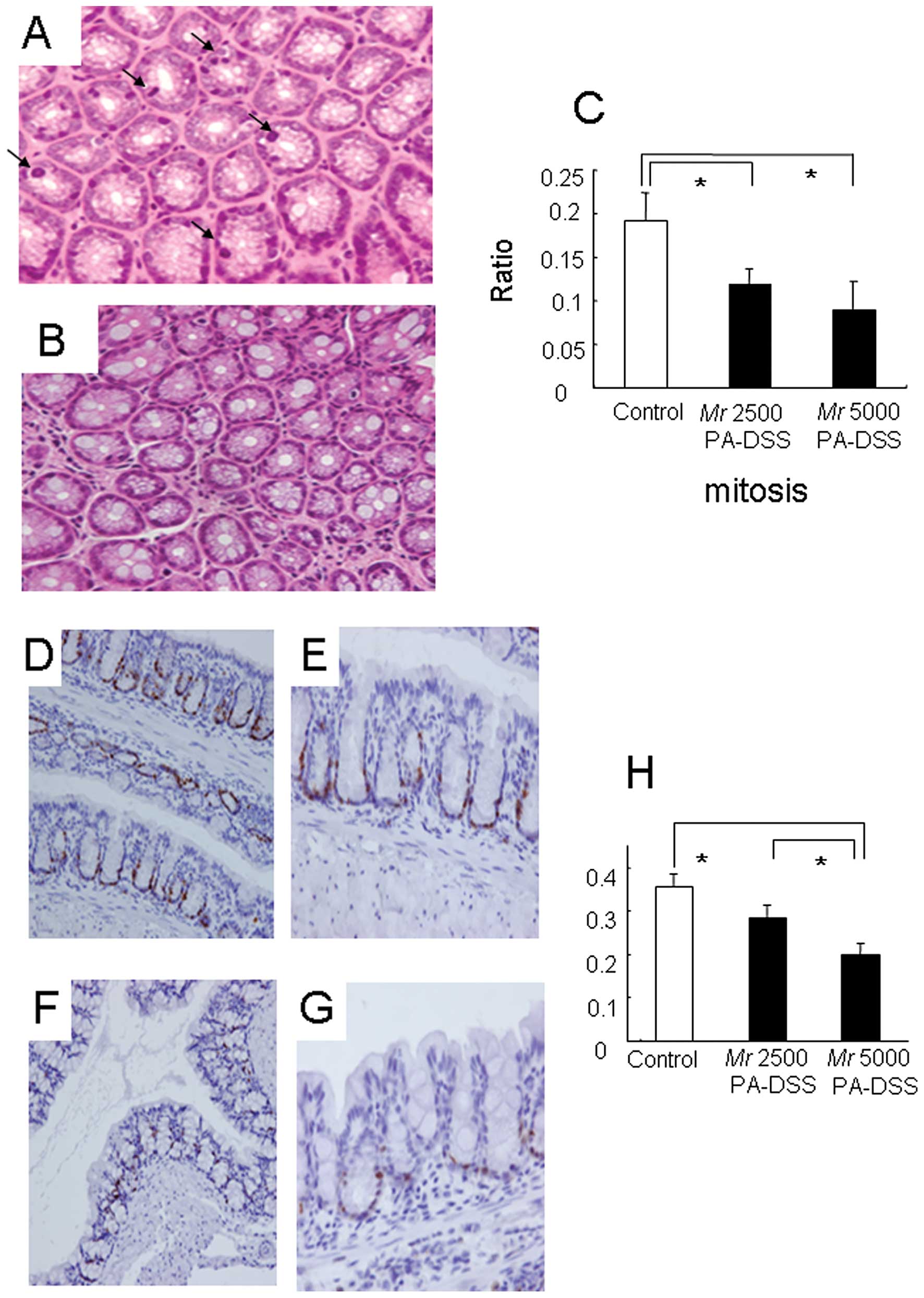
In the control mice, there was sporadic mitosis
present in the crypts using H&E staining (Fig. 4A). However, this mitosis was much
less frequent in mice fed Mr 5000 PA-DSS (Fig. 4B). To quantify this mitotic
activity, the ratio of the cell count with mitosis to the total
cell count in the crypt was calculated. Mr 5000 PA-DSS
significantly reduced this mitosis ratio as compared to the control
mice (Fig. 4C). With respect to Mr
2500 PA-DSS, the reductions in the mitotic activity were somewhere
between the control mice and the mice fed Mr 5000 PA-DSS.
On the other hand, we observed the cell cycle status
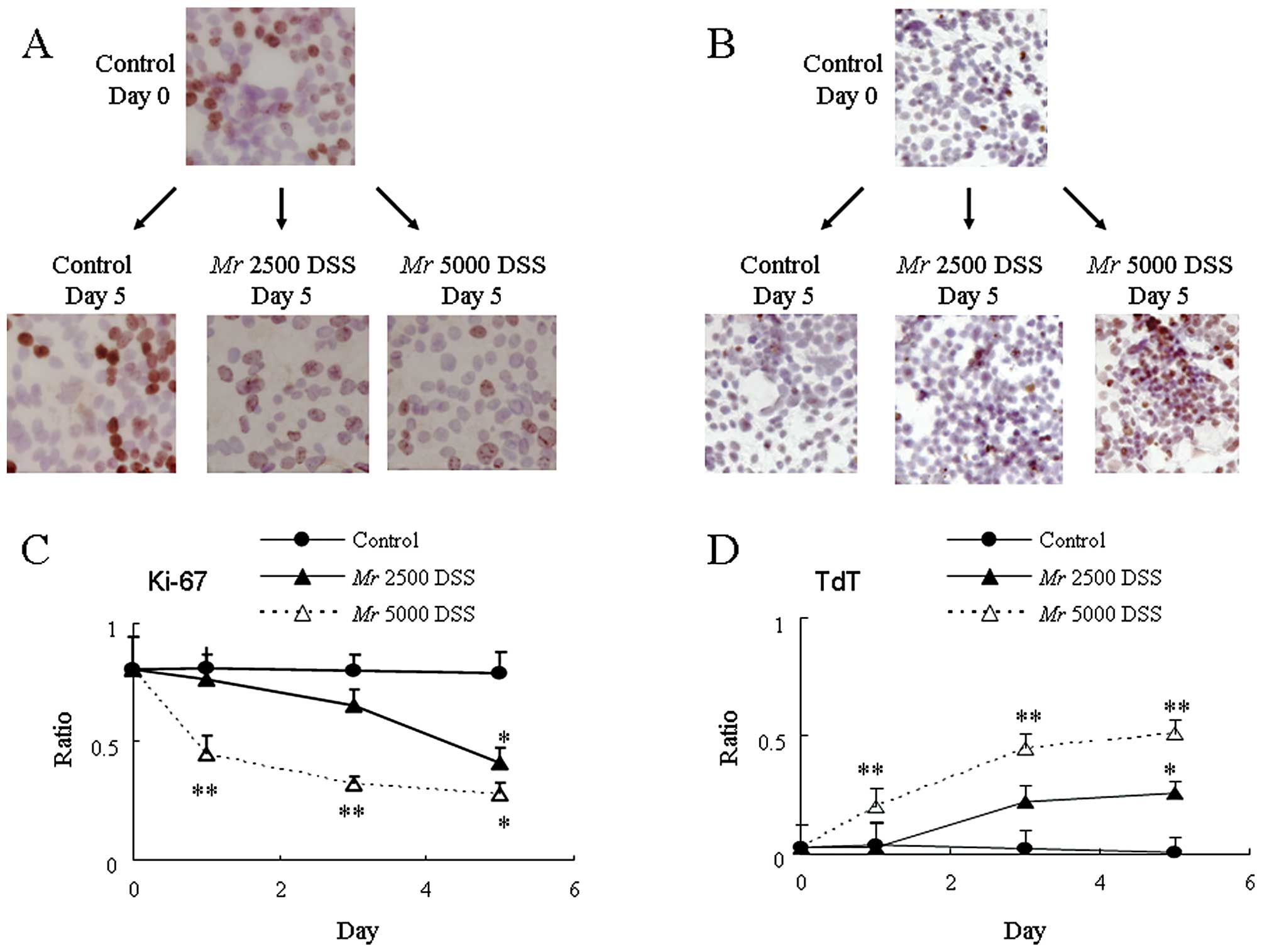
of the colonic epithelium. Fig.
4D-G show the immunostaining using anti-mouse Ki-67 antibody,
which labels all active phases of the cell cycle (G1, S, G2 and M
phases). In the control mice, there were anti-Ki-67 immunopositive
cells in the lower part of the crypt (Fig. 4D and E). However, these anti-Ki-67
immunopositive cells were far fewer in the lower part of the crypt
in mice fed Mr 5000 PA-DSS (Fig. 4F and
G). To quantify these anti-Ki-67 immunopositive cells, the
ratio of the anti-Ki-67 immunopositive cell count to the total cell
count in the crypt was calculated. Mr 5000 PA-DSS significantly
reduced the anti-Ki-67 immunopositive cell ratio as compared to the
control mice (Fig. 4H). With
respect to Mr 2500 PA-DSS, the reductions in the anti-Ki-67
immunopositive cell ratio was somewhere between the control mice
and the mice fed Mr 5000 PA-DSS.
Apoptosis in PA-DSS-induced colitis
It has become clear that DSS arrested the cell cycle
of the colonic epithelium. Therefore, we investigated whether DSS
induces apoptosis of the colonic epithelium or not as the dynamics
of the epithelial cells.
There were a few apoptotic bodies present,
especially at the top of the villi, in the control mice (Fig. 5A). These apoptotic bodies were much
more frequent in mice fed Mr 5000 PA-DSS (Fig. 5B and C). Fig. 5D and E show the staining using the
TUNEL assay. In the control mice, there were a few TdT positive
cells in the epithelium (Fig. 5D).
However, these TdT-positive cells were far more frequent in mice
fed Mr 5000 PA-DSS (Fig. 5E). We
randomly calculated the TdT positive cell count in high power field
(×400). Mr 5000 PA-DSS significantly increased the number of TdT
positive cells as compared to the control mice (Fig. 5F). With respect to Mr 2500 PA-DSS,
the increase in the TdT positive cells was somewhere between the
control mice and mice fed Mr 5000 PA-DSS. These results suggest
that DSS essentially induced cell cycle arrest, especially at the
G0 phase, and apoptosis of the colonic epithelial cells
in vivo.
Metabolism of PA-DSS in Caco-2 cells
The molecular mass in the feces was predominantly Mr
5000, whereas that in the mucosa of the colon was below Mr 2000.
Does this result mean that some of the DSS was depolymerized when
it passed through the epithelial cells? In addition, can DSS also
induce cell cycle arrest and apoptosis in culture cells? We
investigated the metabolism of PA-DSS using Caco-2 cells in order
to clarify these possibilities. In general, a mixture of DSS and
toluidine blue exhibits a color change from blue to violet called
metachromasia. This color change is derived from the shift of the
UV absorbance from 620 to 560 nm and a minimum molecular mass of
2,500 Da was required for a metachromatic reaction according to our
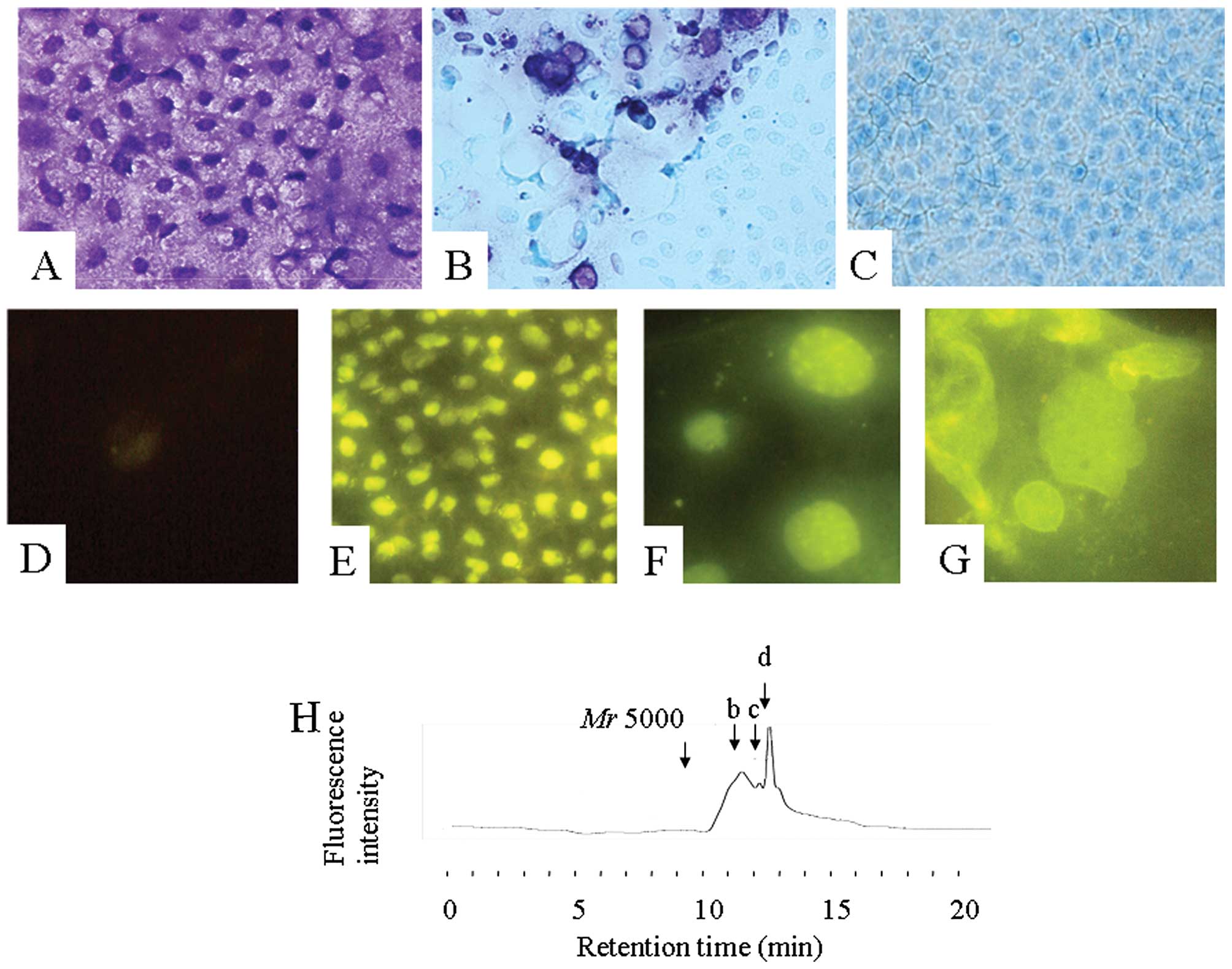
previous study (13). Fig. 6A shows the toluidine blue staining
of Caco-2 cells after 24 h of co-incubation with Mr 5000 DSS in
medium. The metachromatic reaction was present mainly in the
nucleus. At 3 h after replacing with medium alone, the
metachromatic reaction disappeared gradually (Fig. 6B) and was no longer detectable at 24
h (Fig. 6C). These findings
strongly suggest that Mr 5000 DSS entered the cells and was
depolymerized to under Mr 2500. Under a fluorescence microscope,
the fluorescence intensity of the control CaCo-2 cells was very
weak (Fig. 6D). Mr 5000 PA-DSS also
entered the cells, and was bound mainly to the nucleus (Fig. 6E). On the surface of the nucleus,
several more densely-bound areas were recognized (Fig. 6F). Some cells exhibited apoptotic
features including nuclear shrinkage or fragmentation (Fig. 6G). The fluorescence intensity was
also gradually reduced after replacing with medium alone.
The Mr 5000 PA-DSS was also depolymerized while it
passed through the Caco-2 monolayer. Fig. 6H shows a chromatogram of PA-DSS in
the medium outside of a porous filter at 5 h after 1% Mr 5000
PA-DSS in the medium was inserted inside the filter. At this time
point, three peaks were recognized: approximately Mr 1500 (peak b),
1200 (peak c) and 750 (peak d), respectively. The 1% Mr 5000 PA-DSS
solution did not reduce the TEER, indicating that Mr 5000 PA-DSS
passed through a transcellular and, not through paracellular
pathway. These results confirmed that the Mr 5000 PA-DSS was
depolymerized in Caco-2 cells.
Cell cycle arrest and apoptosis in Caco-2
cells
It has become clear that Caco-2 cells can also
depolymerize PA-DSS. We thus investigated if DSS can also induce
the cell cycle arrest and apoptosis in Caco-2 cells.
Fig. 7A shows the
immunostaining using an anti-mouse Ki-67 antibody on Days 0 and 5
after co-incubation with Mr 5000 or 2500 DSS in the medium. We
calculated the ratio of the anti-Ki-67 immunopositive cell count to
the total cell count in randomly-observed fields (Fig. 7C). These anti-Ki-67 immunopositive
cells gradually decreased after co-incubation with 3% Mr 5000 DSS
in the medium. On the other hand, Fig.
7B shows the immunostaining using the TUNEL assay on Days 0 and
5 after co-incubation with Mr 5000 or 2500 DSS in medium. We
calculated the ratio of the TdT positive cell count to the total
cell count in randomly-observed fields (Fig. 7D). These TdT positive cells
gradually increased after co-incubation with 3% Mr 5000 DSS in the
medium. With respect to 3% Mr 2500 DSS in the medium, the
reductions in the anti-Ki-67 immunopositive cell ratio and increase
in the TdT positive cell ratio were somewhere between the control
and co-incubation with 3% Mr 5000 DSS in the medium values. These
results suggest that DSS also essentially induced cell cycle
arrest, especially at the G0 phase, and apoptosis of
Caco-2 cells in vitro.
Discussion
When investigating the pathogenesis of DSS-induced
colitis, the metabolism of DSS is a problem that cannot be avoided.
However, there has been no report that shows in detail the
metabolites of DSS found in the biological materials, including the
contents of the gut lumen. First, Mr 5000 PA-DSS was administer
orally, and was rapidly depolymerized in the stomach. We previously
reported that acidic but not alkaline conditions can depolymerize
PA-DSS (13). In particular, 1 N
HCl was best suited for this reaction without depleting the
sulfates in the molecule. Therefore, we speculated that the gastric
acid might depolymerize the Mr 5000 PA-DSS. After the Mr 5000
PA-DSS was depolymerized in the gastric lumen, PA-DSS entered the
lumen of the small intestine. The depolymerized PA-DSS
concentration in the lumen of the small intestine reached ~30-fold
greater than in the stomach contents. The amount of depolymerized
PA-DSS in the wall of the small intestine was 3-fold greater than
in the gastric mucosa. In addition, the molecular mass distribution
of depolymerized PA-DSS in the blood, spleen, and liver was the
same as the contents and mucosa of the small intestine. These
results suggest that depolymerized PA-DSS could be easily absorbed
in the small intestine. After entering the cecum, however, the
molecular mass distribution was remarkably altered; contrary to our
expectations, the Mr 5000 PA-SS became the main form. This Mr 5000
PA-SS possibly escaped absorption in the small intestine.
Regardless, it has become clear that Mr 5000 PA-DSS in the colon
essentially strongly induced cell cycle arrest, especially at the
G0 phase, and apoptosis of the colonic epithelial cells
than depolymerized PA-DSS in vivo and in vitro.
Therefore, this difference in the molecular mass distribution of
PA-DSS in the lumen may explain the fact that PA-DSS mainly induces
inflammation in the colon, but not in the stomach or the small
intestine.
In addition, the intestinal epithelial cells are
able to uptake and depolymerize PA-DSS. On the other hand, even Mr
500k DSS could enter Caco-2 cells, and the nucleus exhibited
metachromasia (data not shown). DSS could also enter other types of
cells, such as IEC-6 (a rat intestinal epithelial cell line) and
Hep G2 (a human hepatoblastoma cell line) cells, and became
depolymerized (data not shown). Since dextran (non-sulfated
polysaccharide) cannot enter these cells nor the epithelium, these
characteristics of DSS are quite remarkable. This DSS pathway,
however, has not been clarified yet. In addition, it is still
unclear how the DSS was depolymerized or what enzymes were
involved.
There have been a few previous reports showing in
part the metabolism of DSS in the biological materials. In those
reports, DSS was detected mainly using a metachromatic reaction
(17–19). However, our method using the
PA-labeling system seems more reliable. DSS depolymerized in the
gut lumen cannot induce a metachromatic reaction any more because a
minimum molecular mass of 2,500 Da is required for DSS to induce a
metachromatic reaction according to our previous study (13).
In the present study, the precise mechanisms
responsible for the induction of cell cycle arrest and apoptosis
remain unclear. Some previous reports, however, have suggested an
association between DSS and the cell cycle arrest. For example, the
cell cycle of B lymphocytes was arrested by DSS (20). In this process, DSS inhibited the
DNA topoisomerases, and controlled cell proliferation via DNA
polymerase (21). This effect was
the strongest at around Mr 4300 (22). In addition, we previously reported
that DSS strongly and rapidly inhibited reactive oxygen species
(ROS) generation in Caco-2 cells. In this study, ROS generation was
reduced to ~3% of the control value by 5 h after 1% DSS exposure
(16). The mechanisms responsible
for this reduction in ROS generation remain unclear. This
phenomenon, however, also suggests that DSS arrests intracellular
energy metabolism, because ROS are generated during energy
metabolism. Taken together, the early stage of DSS-induced colitis
might involve a cytostatic mechanism (cellular metabolic arrest and
cell cycle arrest), in other words, as if the cells were in
hibernation.
With respect to apoptosis, a few previous reports
showed the involvement of apoptosis in DSS-induced colitis
(23). Since intestinal epithelial
cells have a rapid cell cycle, it is logical to postulate that cell
cycle arrest and apoptosis in addition to cellular metabolic arrest
might lead to crypt shortening, entire crypt loss, and an
earthenware mortar-like deformity of the crypt, and surface
epithelial loss, which were partly reported by Cooper et al
(24). Consequently, the mucosa
might eventually develop breaks in the barrier of the epithelial
cells as an early stage event.
It is also very interesting that T and B cells are
not required for acute DSS-induced colitis, since DSS also produces
colitis in severe combined immunodeficient (SCID) mice (25). Another previous study also suggests
that DSS exhibits a direct toxicity against colonic epithelial
cells, IEC-18 cells (26).
Thereafter, the mucosal breaks would lead to mucosal inflammatory
changes (24), for example through
macrophage activation (27), the
invasion of intestinal bacteria, or a variety of cytokine cascades.
In that sense, many previous studies which reported that some
growth hormones or factors ameliorated DSS-induced colitis
(28–31) might be reasonable because these
kinds of agents have been reported to promote the cell cycle
(32,33). On the other hand, Kitajima et
al reported that the degree of colitis was stronger in the
order of Mr 40k DSS > Mr 5000 DSS in mice (17).
In addition, carrageenan is produced from seaweed
and is used as a gelling agent in food. Carrageenan is another type
of sulfated polysaccharide consisting of anhydro-D-galactose
(α-1,3-galactosidic link) and sulfate (over Mr 100k to 800k), and
also induces colitis (34).
Carrageenan also arrested the cell cycle (35). Interestingly, the acid-degraded form
of carrageenan (poligeenan, Mr 40k) more strongly caused colitis.
It is also said that a critical DSS load exceeding 30 mg/g body wt
was required to induce colitis (36). Taken together, the most important
finding might be that a certain amount of DSS with a certain range
of molecular masses is required to induce cell cycle arrest and
apoptosis after the DSS is depolymerized in the gut lumen, and
subsequently enters the cells. These results might attract the
interest of many researchers. When focusing on the etiology of IBS
or other types of colitis, including DSS-induced colitis, other
kinds of materials may induce the cytostatic effects, apoptosis and
eventually colitis.
Finally, several types of bacteria, including
Pseudomonas carrageenovora of marine origin, have been
identified which are able to hydrolyze carrageenan (37). We also reported that DSS was
depolymerized by Proteus species in the gut lumen (38). It is possible that some kinds of
bacteria in the gut lumen may depolymerize the DSS and modulate the
DSS-induced colitis.
In conclusion, we proposed one plausible mechanism
responsible for an early stage event in DSS-induced colitis. DSS is
depolymerized rapidly in the mouse stomach. This depolymerized DSS
cannot induce severe inflammation in the stomach or small
intestine. However, the majority of the DSS in the colonic lumen
returned to the Mr 5000 form. This Mr 5000 DSS induced even more
severe colitis through cell cycle arrest and apoptosis than the
depolymerized DSS.
References
|
1
|
Fiocchi C: Inflammatory bowel disease:
etiology and pathogenesis. Gastroenterology. 115:182–205. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bernstein CN, Blanchard JF, Kliewer E and
Wajda A: Cancer risk in patients with inflammatory bowel disease: a
population-based study. Cancer. 91:854–862. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Elson CO, Sartor RB, Tennyson GS and
Riddell RH: Experimental models of inflammatory bowel disease.
Gastroenterology. 109:1344–1367. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Watt J and Marcus R: Ulceration of the
colon in rabbits fed sulfated amylopectin. J Pharm Pharmacol.
24:68–69. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ricketts CR: Dextran sulphate-a synthetic
analogue of heparin. Biochem J. 51:129–133. 1952.PubMed/NCBI
|
|
6
|
Tanaka T, Kohno H, Suzuki R, Yamada Y,
Sugie S and Mori H: A novel inflammation-related mouse colon
carcinogenesis model induced by azoxymethane and dextran sodium
sulfate. Cancer Sci. 94:965–973. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suzuki R, Kohno H, Sugie S and Tanaka T:
Sequential observations on the occurrence of preneoplastic and
neoplastic lesions in mouse colon treated with azoxymethane and
dextran sodium sulfate. Cancer Sci. 95:721–727. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yamada M, Ohkusa T and Okayasu I:
Occurrence of dysplasia and adenocarcinoma after experimental
chronic ulcerative colitis in hamsters induced by dextran sulphate
sodium. Gut. 33:1521–1527. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Okayasu I, Hatakeyama S, Yamada M, Ohkusa
T, Inagaki Y and Nakaya R: A novel method in the induction of
reliable experimental acute and chronic ulcerative colitis in mice.
Gastroenterology. 98:694–702. 1990.PubMed/NCBI
|
|
10
|
Araki Y, Andoh A, Fujiyama Y, Hata K,
Makino J, Okuno T, Nakanura F and Bamba T: Application of
2-aminopyridine fluorescence labeling in the analysis of in vivo
and in vitro metabolism of dextran sulfate sodium by size-exclusion
high-performance liquid chromatography. J Chromatogr B Biomed Sci
Appl. 753:209–215. 2001. View Article : Google Scholar
|
|
11
|
Arak Y, Andoh A, Fujiyama Y, Hata K,
Makino J, Shimada M, Bamba H, Okuno T, Urushiyama N and Bamba T:
Analysis of α-amylase-derived pyridylamino-dextran sulfate
oligomers by the combination of size-exclusion and reversed-phase
high-performance liquid chromatography. J Chromatogr B Analyt
Technol Biomed Life Sci. 766:351–356. 2002.
|
|
12
|
Araki Y, Kanauchi O, Sugihara H, Fujiyama
Y and Hattori T: Germinated barley foodstuff suppresses dextran
sulfate experimental colitis in rats: the role of mast cells. Int J
Mol Med. 19:257–262. 2007.PubMed/NCBI
|
|
13
|
Araki Y, Mukaisyo K, Sugihara H and
Hattori T: Decomposition of dextran sulfate sodium under alkaline,
acidic, high temperature and high pressure conditions. Oncol Rep.
20:147–149. 2008.PubMed/NCBI
|
|
14
|
Hidalgo IJ, Raub TJ and Borchardt RT:
Characterization of human colon carcinoma cell line (Caco-2) as a
model system of intestinal epithelial permeability.
Gastroenterology. 96:736–749. 1989.PubMed/NCBI
|
|
15
|
Araki Y, Katoh T, Ogawa A, Bamba S, Andoh
A, Koyama S, Fujiyama Y and Bamba T: Bile acid modulates
transepithelial permeability via the generation of reactive oxygen
species in the Caco-2 cell line. Free Radic Biol Med. 39:769–780.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Araki Y, Sugihara H and Hattori T: In
vitro effects of dextran sulfate sodium on a Caco-2 cell line and
plausible mechanisms for dextran sulfate sodium-induced colitis.
Oncol Rep. 16:1357–1362. 2006.PubMed/NCBI
|
|
17
|
Kitajima S, Takuma S and Morimoto M:
Histological analysis of murine colitis induced by dextran sulfate
sodium of different molecular weights. Exp Anim. 49:9–15. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hoshi O, Iwanaga T and Fujino MA:
Selective uptake of intraluminal dextran sulfate sodium and senna
by macrophages in the cecal mucosa of the guinea pig. J
Gastroenterol. 31:189–198. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hiebert LM, Wice SM, Jaques LB, Williams
KE and Conly JM: Orally administered dextran sulfate is absorbed in
HIV-positive individuals. J Lab Clin Med. 133:161–170. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Burg DL, Harrison ML and Geahlen RL: Cell
cycle-specific activation of the PTK72 protein-tyrosine kinase in B
lymphocytes. J Biol Chem. 68:2304–2306. 1993.PubMed/NCBI
|
|
21
|
Umemura K, Yanase K, Suzuki M, Okutani K,
Yamori T and Andoh T: Inhibition of DNA topoisomerases I and II,
and growth inhibition of human cancer cell lines by a marine
microalgal polysaccharide. Biochem Pharmacol. 66:481–487. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Furukawa K and Bhavanandan VP: Influences
of anionic polysaccharides on DNA synthesis in isolated nuclei and
by DNA polymerase alpha: correlation of observed effects with
properties of the polysaccharides. Biochim Biophys Acta.
740:466–475. 1983. View Article : Google Scholar
|
|
23
|
Vetuschi A, Latella G, Sferra R, Caprilli
R and Gaudio E: Increased proliferation and apoptosis of colonic
epithelial cells in dextran sulfate sodium-induced colitis in rats.
Dig Dis Sci. 47:1447–1457. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cooper HS, Murthy SN, Shah RS and
Sedergran DJ: Clinicopathologic study of dextran sulfate sodium
experimental murine colitis. Lab Invest. 69:238–249. 1993.
|
|
25
|
Dieleman LA, Ridwan BU, Tennyson GS,
Beagley KW, Bucy RP and Elson CO: Dextran sulfate sodium-induced
colitis occurs in severe combined immunodeficient mice.
Gastroenterology. 107:1643–1652. 1994.
|
|
26
|
Ni J, Chen SF and Hollander D: Effects of
dextran sulfate sodium on intestinal epithelial cells and
intestinal lymphocytes. Gut. 39:234–241. 1996. View Article : Google Scholar
|
|
27
|
Yoshino T, Nakase H, Honzawa Y, Matsumura
K, Yamamoto S, Takeda Y, Ueno S, Uza N, Masuda S, Inui K and Chiba
T: Immunosuppressive effects of tacrolimus on macrophages
ameliorate experimental colitis. Inflamm Bowel Dis. 16:2022–2033.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao J, de Vera J, Narushima S, Beck EX,
Palencia S, Shinkawa P, Kim KA, Liu Y, Levy MD, Berg DJ, Abo A and
Funk WD: R-spondin1, a novel intestinotrophic mitogen, ameliorates
experimental colitis in mice. Gastroenterology. 132:1331–1343.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Egger B, Carey HV, Procaccino F, Chai NN,
Sandgren EP, Lakshmanan J, Buslon VS, French SW, Büchler MW and
Eysselein VE: Reduced susceptibility of mice overexpressing
transforming growth factor alpha to dextran sodium sulfate induced
colitis. Gut. 43:64–70. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Williams KL, Fuller CR, Dieleman LA,
DaCosta CM, Haldeman KM, Sartor RB and Lund PK: Enhanced survival
and mucosal repair after dextran sodium sulfate-induced colitis in
transgenic mice that overexpress growth hormone. Gastroenterology.
120:925–937. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tessner TG, Cohn SM, Schloemann S and
Stenson WF: Prostaglandins prevent decreased epithelial cell
proliferation associated with dextran sodium sulfate injury in
mice. Gastroenterology. 115:874–882. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lepique AP, Moraes MS, Rocha KM and
Eichler CB: c-Myc protein is stabilized by fibroblast growth factor
2 and destabilized by ACTH to control cell cycle in mouse Y1
adrenocortical cells. J Mol Endocrinol. 33:623–638. 2004.
View Article : Google Scholar
|
|
33
|
Slominski A, Zbytek B, Pisarchik A,
Slominski RM, Zmijewski MA and Wortsman J: CRH functions as a
growth factor/cytokine in the skin. J Cell Physiol. 206:780–791.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Watt J and Marcus R: Carrageenan induced
ulceration of the large intestine in the guinea pig. Gut.
12:164–171. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ling KY, Bhalla D and Hollander D:
Mechanisms of carrageenan injury of IEC18 small intestinal
epithelial cell monolayers. Gastroenterology. 95:1487–1495.
1988.PubMed/NCBI
|
|
36
|
Vowinkel T, Kalogeris TJ, Mori M,
Krieglstein CF and Granger DN: Impact of dextran sulfate sodium
load on the severity of inflammation in experimental colitis. Dig
Dis Sci. 49:556–564. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McLean MW and Williamson FB:
kappa-Carrageenase from Pseudomonas carrageenovora. Eur J Biochem.
93:553–558. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Araki Y, Mukaisho K, Sugihara H, Fujiyama
Y and Hattori T: Proteus mirabilis sp intestinal microflora
grow in a dextran sulfate sodium-rich environment. Int J Mol Med.
25:203–208. 2010.
|