Introduction
Epigallocatechin-3-gallate (EGCG), which is also
known as catechin, is the most abundant polyphenolic compound
extracted from green tea (1).
Numerous reports have demonstrated the beneficial health effects of
EGCG in cancer, anti-oxidation, inflammation and hypertension
(2–6). Moreover, EGCG exhibits an
anti-photoaging effect in skin cells. Ultraviolet (UV) radiation is
the main inducer of photoaging, which is characterized by collagen
loss, reactive oxygen species (ROS) generation, cell senescence and
apoptosis. In vitro studies have demonstrated that EGCG
protects against oxidative cellular damage in skin cells caused by
UV radiation (7). Studies in mice
demonstrated that EGCG prevents photocarcinogenesis through DNA
repair (8). Furthermore, EGCG
pretreatment inhibited UVB-mediated thinning of the epidermis,
thereby restoring epidermal thickness and rendering the basal layer
more compact in living skin equivalents (9). Together, these results suggest that
EGCG is a potential agent for preventing photoaging in skin.
Although the anti-photoaging effect of EGCG on skin
cells has been broadly researched, molecular studies investigating
the EGCG-mediated UVB protective effect have been limited. In
keratinocytes, EGCG inhibited UVB-induced activation of activator
protein-1 (AP-1) and p38 mitogen-activated protein kinase (MAPK)
(10). In addition, UVB-induced
NF-κB activation and IL-6 expression were attenuated by EGCG
treatment (11). One molecular
study investigating dermal fibroblasts reported that EGCG may
prevent UVB-induced collagenolytic MMP production by interfering
with the MAPK pathways (12).
Despite this knowledge, the molecular mechanisms underlying the
anti-photoaging effect of EGCG remain largely unknown.
microRNAs (miRNAs) are small (~19 nt), non-coding
RNA molecules expressed in eukaryotes that regulate gene expression
by inhibiting the translation of their target mRNA (13). These small RNA molecules play
central roles in several biological and disease processes,
including cell survival, apoptosis, metabolism, cancer and diabetes
(14). miRNAs have also been
associated with several important functions in skin cells. For
example, miR-203 is expressed only in terminally differentiated
cells and downregulates p63, an essential regulator of stem-cell
maintenance, to enable pluripotent cells to differentiate into the
stratified layers that compose the skin (15). Moreover, DGCR8-mediated miRNA
biogenesis is essential for skin development, indicating that
miRNAs are an important regulator of mammalian skin development
(16). Recent miRNA expression
profiling analyses of UVB-irradiated normal human keratinocytes
have revealed several specific miRNA expression patterns (17). However, characterization of miRNA
expression associated with the UVB-protective ability in human
dermal fibroblasts has yet to be performed. In this current study,
we conducted miRNA expression profiling of EGCG-treated normal
human dermal fibroblasts (NHDFs). Our data demonstrate that the
anti-photoaging effect of EGCG induced several specific miRNA
expression patterns involved in cell proliferation.
Materials and methods
Cell culture
NHDFs were purchased from Lonza (Basel, Switzerland)
and cultured in Dulbecco's modified Eagle's medium (DMEM, Gibco,
Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (FBS; Sigma-Aldrich, St. Louis, MO, USA), penicillin (100
U/ml) and streptomycin (100 μg/ml). The cells were incubated at
37°C in a humidified atmosphere containing 5% CO2.
Cytotoxicity and UVB protection
assay
NHDFs (3×103) were seeded into 96-well
plates and incubated overnight before treatment with several
concentrations of EGCG for 24 h. The cytotoxicity of EGCG was
measured by the WST-1 assay (EZ-CytoX Enhanced cell viability assay
kit; Daeil Lab Service, Seoul, Korea) according to the
manufacturer's instructions.
To assess UVB protection, NHDFs were first
pre-treated with DMSO (Sigma-Aldrich) or EGCG (Sigma-Aldrich) for 3
h. Then the cells were washed and exposed to 100 mJ/cm2
UVB without protective filters. After irradiation, the cells were
cultured in tissue culture media with DMSO or EGCG for an
additional 24 h before cell viability was measured using the WST-1
assay.
RNA purification
Total RNA was extracted using TRIzol reagent
(Invitrogen) according to the manufacturer's instructions. The
integrity of each RNA sample was verified with an Agilent 2100
Bioanalyzer® (Agilent Technologies, Santa Clara, CA,
USA). A260/A230 and A260/A280 ratios between 1.8 and 2.1 were
confirmed in all RNA samples using the MaestroNano®
spectrophotometer (Maestrogen, Las Vegas, NV, USA).
Microarray analysis of miRNA
profiles
miRNA profiling analysis was performed using
SurePrint G3 Human V16 miRNA 8×60K microarrays (Agilent
Technologies) containing probes for 1205 human and 144 viral
miRNAs. Each RNA sample was dephosphorylated, labeled with Cyanine
3-pCp using T4 RNA ligase, dried completely and then treated with
GE blocking agent (Agilent Technologies). The RNA was applied to
the microarrays and then placed in the Agilent microarray
hybridization chamber (Agilent Technologies) for 20 h. The
microarrays were imaged using an Agilent scanner, and quantitative
data for the miRNA profiles were extracted from the images using
the Feature Extraction program (Agilent Technologies). These data
were analyzed with GeneSpring GX software version 7.3 (Agilent
Technologies). miRNAs with flags present in at least one sample
were filtered and subjected to fold-change analysis. The
fold-change analysis was conducted by a factor of 1.5-fold between
the samples.
Computational analysis of miRNA
expression
miRNAs that exhibited significant differential
expression were selected, and their putative cellular target genes
were determined using microCosm Target version 5 (www.ebi.ac.uk/enright-srv/microcosm/htdocs/targets/v5/).
Target genes were categorized into four groups, namely aging,
apoptosis, cell proliferation, and skin development, using the Gene
Ontology analysis tool AmiGO (amigo.geneontology.org/cgi-bin/amigo/browse.cgi). The
genes were then further categorized into groups such as
anti-apoptosis, activation of MAPKK activity, Ras protein signal
transduction, small GTPase-mediated signal transduction, positive
or negative regulation of cell growth, cell proliferation, cell
cycle and regulation of transcription.
Results and Discussion
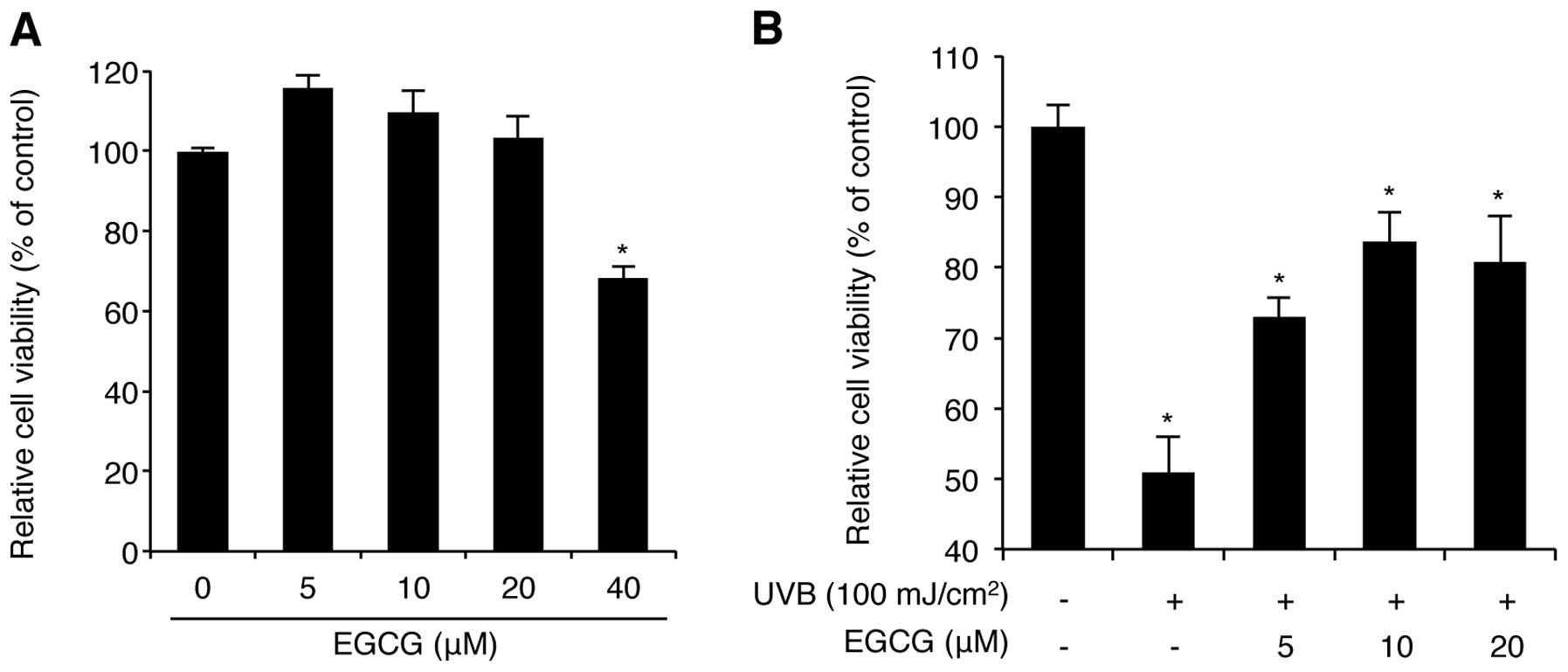
Before assessing the protective activity of EGCG
against UVB radiation, we investigated the cytotoxicity of this
compound on NHDFs at various doses. WST-1-based cell proliferation
analysis demonstrated that 5-20 μM of EGCG for 24 h did not affect
cell viability, whereas a 40 μM dose decreased cell viability to
below 65% (Fig. 1A). In fact, a
previous report showed that treatment of NHDFs with >50 μM EGCG
reduced cell viability by 50% as determined by the MTT assay
(18). Thus, high doses (>40 μM)
of EGCG decrease cell viability, while low doses (<40 μM) have
no cytotoxic effects on NHDFs. Therefore, our analysis of
EGCG-mediated UVB protection was performed using 5, 10, and 20 μM
of EGCG. We confirmed that treatment with 10 μM of EGCG rescued
NHDFs from UVB radiation (100 mJ/cm2)-mediated cell
death up to 83.7% compared with the control NHDFs (Fig. 1B). These results are consistent with
previous studies (12), indicating
that EGCG has a strong photo-protective effect against UVB
radiation on NHDFs.
Studies of the molecular details underlying this
photo-protective effect have been focused on changes in MAPK
activation, including JNK, p38 MAPK and ERK1/2 phosphorylation.
However, these changes are not specific to EGCG-treated NHDFs. UVB
radiation induces MAPK phosphorylation in a variety of cell types,
including NHDFs, keratinocytes, melanocytes and cancer cells
(19). Moreover, UVB radiation does
not increase MAPK protein synthesis in cells, but rather the level
of MAPK activity (19). Notably,
several natural and chemical agents reported to exhibit a UVB
photo-protective effect share similar mechanisms, including MAPK
phosphorylation (20–23). These data indicate that, although
the MAPK pathways are important regulators in photo-protective
mechanisms, their activity is not specific to the protection
against UVB radiation. Besides these signaling pathways,
EGCG-mediated anticancer properties have been associated with
changes in the expression of specific miRNAs, namely miR-16 and
miR-210, indicating that EGCG may function by regulating miRNA
expression (24,25). Therefore, we proceeded to identify
the specific regulators of the EGCG-mediated photo-protective
effect on NHDFs using miRNA expression profiling analysis that also
demonstrated tissue- and process-specific expression of miRNAs in
various reports (26,27).
The Agilent SurePrint G3 Human v16 miRNA microarray,
which contains probes representing 1205 human and 144 viral miRNAs,
was used to investigate changes in miRNA expression induced by EGCG
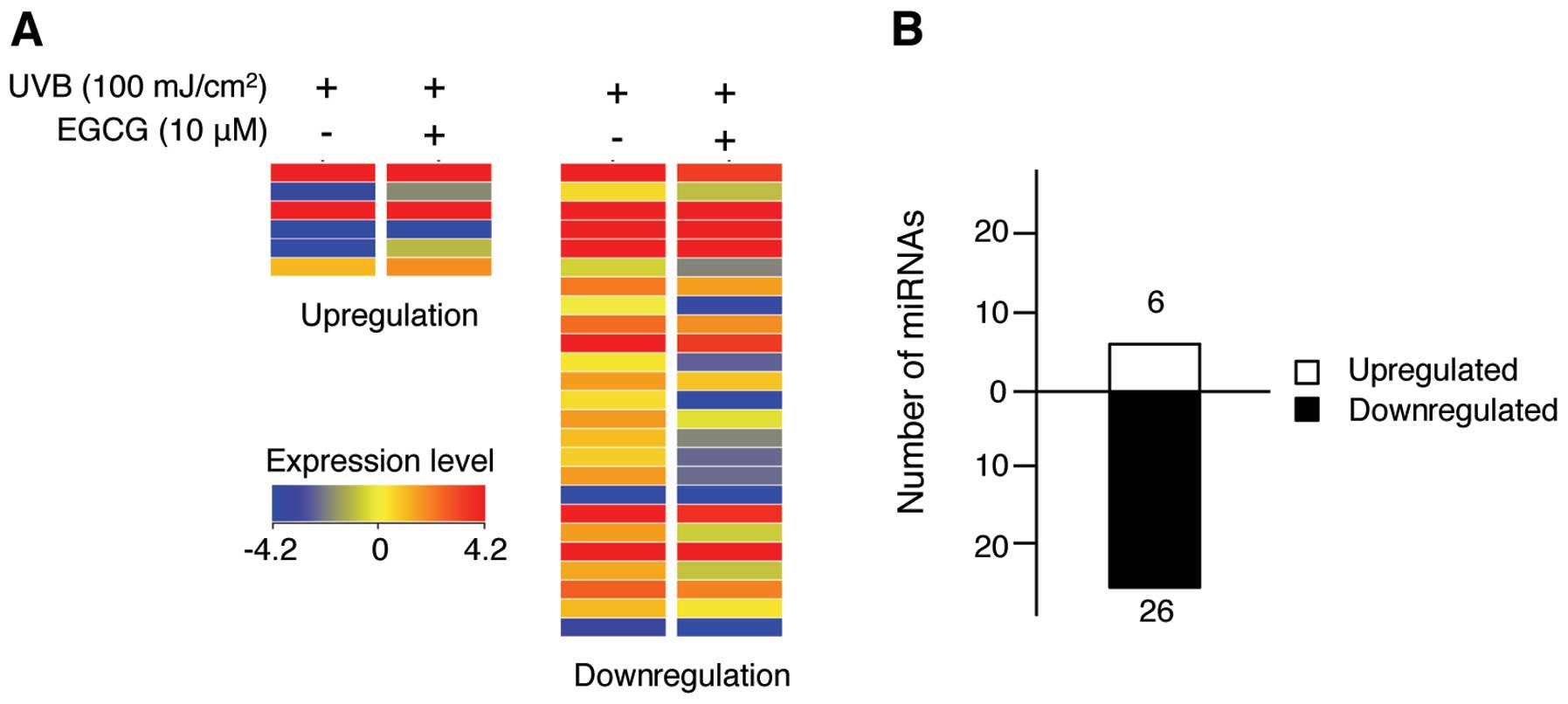
treatment of UVB-irradiated NHDFs. Our study revealed that 6 and 26
miRNAs were upregulated and downregulated greater than 1.2-fold,
respectively (Fig. 2). These miRNAs
are listed in Table I. With a
2.7-fold increase, miR-636 exhibited an increased upregulation.
miR-3907, which displayed a 2.30-fold decrease, was downregulated
to the greatest extent in this experiment. These data suggest that
these miRNAs may be novel targets of EGCG. Of note, the microarray
results showed that the number of downregulated miRNAs was higher
than the number of upregulated miRNAs, indicating that the
EGCG-mediated photo-protective effect is associated more with miRNA
downregulation. miR-133a and miR-212, which were downregulated 1.8-
and 1.61-fold, respectively, have been reported to induce apoptosis
in bladder cancer and non-small cell lung cancer cells (28,29).
In addition, miR-513a-5p, which was downregulated 1.74-fold in our
study, reportedly mediates TNF-α- and LPS-induced apoptosis in
human umbilical vein endothelial cells (30). Although such miRNAs have not been
associated with UVB-mediated apoptosis, these results suggest that
EGCG inhibits UVB-mediated NHDF cell death by downregulating
apoptosis-related miRNAs. Overall, these results indicate that EGCG
may regulate specific miRNA expression levels to mediate UVB
photoprotection.
 | Table ImiRNAs exhibiting a greater than
1.2-fold change in expression upon EGCG treatment in UVB-exposed
NHDFs. |
Table I
miRNAs exhibiting a greater than
1.2-fold change in expression upon EGCG treatment in UVB-exposed
NHDFs.
| miR name | FC | Chr | miR name | FC | Chr |
|---|
| hsa-miR-1246 | 1.23 | - | hsa-miR-212 | −1.61 | Chr17 |
| hsa-miR-145* | 1.32 | Chr22 | hsa-miR-3141 | −1.23 | Chr1 |
| hsa-miR-4299 | 1.29 | Chr9 | hsa-miR-3610 | −2.17 | ChrX |
| hsa-miR-548c-3p | 1.36 | Chr16 | hsa-miR-362-3p | −1.70 | Chr1 |
| hsa-miR-636 | 2.70 | Chr19 | hsa-miR-3667-5p | −1.81 | Chr2 |
| hsa-miR-933 | 1.25 | Chr12 | hsa-miR-3679-3p | −1.77 | - |
| ebv-miR-BART12 | −1.45 | Chr8 | hsa-miR-3907 | −2.30 | - |
| ebv-miR-BART1-1 | −1.36 | Chr11 | hsa-miR-423-3p | −2.09 | - |
| hsa-miR-1202 | −1.22 | Chr19 | hsa-miR-4270 | −1.21 | - |
| hsa-miR-1207-5p | −1.22 | Chr7 | hsa-miR-455-5p | −1.77 | - |
| hsa-miR-1225-5p | −1.23 | Chr8 | hsa-miR-494 | −1.29 | Chr11 |
| hsa-miR-1227 | −1.24 | Chr18 | hsa-miR-513a-5p | −1.74 | Chr7 |
| hsa-miR-1271 | −1.23 | Chr20 | hsa-miR-660 | −1.21 | Chr11 |
| hsa-miR-133a | −1.80 | ChrX | hsa-miR-718 | −1.25 | Chr17 |
| hsa-miR-134 | −1.21 | Chr3 | kshv-miR-K12-10b | −1.92 | Chr1 |
| hsa-miR-181d | −1.32 | Chr15 | | | |
miRNAs function by engaging with their target mRNA
and inhibiting its translation (13). We used the MicroCosm Target tool to
identify putative miRNA target genes, analyze their gene ontology,
and categorize them into cellular processes including aging,
apoptosis, cell proliferation, and skin development (Tables II and III). Furthermore, target genes were
re-analyzed and categorized according to the cellular mechanisms to
which they are related, as represented in Table IV. Of note, the genes targeted by
the downregulated miRNAs are mainly involved in regulating
transcription, anti-apoptosis, cell division, cell cycle and small
GTPase-mediated signal transduction. However, these target genes
are not involved in MAPK-related mechanisms, indicating that the
miRNA-based EGCG-mediated photo-protective response is controlled
by specific miRNAs and target genes associated with transcription
and cell survival, but not MAPK activation, in NHDFs.
| Table IIPredicted target genes of miRNAs
upregulated in response to EGCG in UVB-exposed NHDFs. |
Table II
Predicted target genes of miRNAs
upregulated in response to EGCG in UVB-exposed NHDFs.
| Functions of target
genes |
|---|
|
|
|---|
| miRNA | Aging | Apoptosis | Cell
proliferation | Skin development |
|---|
| hsa-miR-1246 | | AQP1 | AQP1, AXIN2, GNRHR,
PTCH1 | - |
| hsa-miR-548c-3p | MME, SMC5, MNT, SMC6,
BBC3, ID2, NUAK1, NR3C1 | MNT, NR3C1, BBC3,
CDKN1B, BCL2L11, DLX1, FOXQ1, OGT, UBE2B, WNK3, TCFL2, BDNF, RFFL,
RPS6KA1, SNCA, TIA1, AKAP13, PPARGC1A, PREX1, CITED2, DLC1, ERBB4,
FLT3, FOXO1, HIPK2, LRP6, ATG5, NFKBIA, SMAD3, TNFSF12, TRIM24,
PAK2, PKP1, CBL, RPS6KA2, TGFBR2, PXN2, PRKAA1, PSMD3, TRAF4 | TNFSF12, TRIM24,
DDR1, PGR, UBR5, ODC1, TOB2, ID4, LRP6, HSF1, FOXO1, HIPK2, NFKBIA,
SMAD3, NUAK1, CITED2, ERBB4, MMP12, MMP14, PBRM1, MNT, NR3C1, ID2,
DLC1, STAT3, VASH2, TCF7L2, ODZ1, TOB1, RPS6KA2, FLT3, CDKN1B,
HMX2, IRF6, TGFBR2, BDNF | DDR1, IRF6, TCF7L1,
TCF7L2, FRAS1 |
| hsa-miR-636 | SOCS3 | RPS6KA3, SENP1, CBL,
PKN2, SOCS3, MITF, SFRP2, TRAF5, PCGF2, PRKCE, PROC, RTN3, ACTN1,
ARF6, GRIK2, ITSN1, YWHAZ, TCF7L2, RPS6KA2, TGFBR2 | TRAF5, BCAT1, FBXW7,
MITF, SFRP2, RPS6KA2, TCF7L2, RNF139, TOB1, EMX2, LIFR, SSR1,
TGFBR2 | TCF7L2 |
| hsa-miR-933 | - | BDNF | BDNF | - |
| Table IIIPredicted target genes of miRNAs
downregulated in response to EGCG in UVB-exposed NHDFs. |
Table III
Predicted target genes of miRNAs
downregulated in response to EGCG in UVB-exposed NHDFs.
| Functions of the
target genes |
|---|
|
|
|---|
| miRNA | Aging | Apoptosis | Cell
proliferation | Skin
development |
|---|
| hsa-miR-1202 | SLC1A2 | DSP, NAIF1, ETS1,
SOS1, UBD, RALB | ARNT, GABBR1,
BCAT1, HOOK3, ETS1 | DSP |
|
hsa-miR-1207-5p | LRP1 | IGF1, MKL1, FGFR1,
UBE2Z, CBL, NOL3, TNS4, EGLN2 | LRP1, FGFR1,
ACVRL1, IGF1, CYP27B1 | - |
|
hsa-miR-1225-5p | - | KIAA1324,
PEG10 | ELN, TAL1 | - |
| hsa-miR-1227 | TGFBR1, SOCS3 | RTNFAIP8, TGFBR1,
SOCS3 | TGFBR1, IRF2 | - |
| hsa-miR-1271 | MAP2K1, EDNRA,
CASP2, DDIT3, PTEN | TGFBR1, CASP2, ALK,
DDIT3, MED1, PROK2, DOCK1, OGT, KPNB1, MBD4, SORT1, TRIB, EDNRA,
FOXO1, TNFSF13B, ECE1, PRKCE, PTEN | PROK2, LAMC1,
RNF139, TNFSF13B, CD164, ALK, EDNRA, MED1, MYO16, FOXO1, LIPG,
MAP2K1, TNS3, NEUROD4, IRS1, KRAS, PTEN | - |
| hsa-miR-133a | ZNF354A, PML | RB1CC1, FOXC1,
FOXL2, PML, MCL1, SGK1, CYLD, EPHA7, SOX4, RFFL, SGPP1, FOXQ1 | PML, SGK1, SOX4,
FGF1, APPL2, BRD4, TXLNA, CNN2, FOXC1, ENPEP, LHX5, NPPC | - |
| hsa-miR-134 | SEPRINE1 | BDNF, STAT5B,
ANGPTL4, PDCD7, WWOX, SERPINE1 | BDNF, STAT5B,
FOXP2, SERPINE1 | - |
| hsa-miR-181d | ATM, TIMP3, ADRBK1,
PRKCD, SIRT1 | IL1A, GATA6, CBX4,
HSP90B1, ITSN1, RAD21, RNF34, UBE2B, TNF, PRKCD, BAG4, BCL2L11,
CARD11, USP47, ATM, DDIT4, BIRC6, INSL3, IRS2, NOTCH2, PDCD6IP,
UNC5A, TRIM2, SIRT1 | S1PR1, KRAS, SIRT1,
LIF, PRKCD, BIRC6, CARD11, ATM, IL1A, INSL3, IRS2, PROX1, RBBP7,
GATA6, NOTCH2, CDON, ING5, PLAU, PDAP1, PRDM4, LRRC32, TNF, MCC,
CBLB | - |
| hsa-miR-212 | CTGF | CTGF, EP300, FOXA1,
MAPK3, FOXO3, ISL1, SGK3, MAPT, RB1, RASA1 | CTGF, FOXO3, ISL1,
RB1, SGK3, EGR1, SPRY1, ZEB2, SOX11 | - |
| hsa-miR-362-3p | GRB2 | CUL2, KRIT1,
BCLAF1, GRIK2, BLCAP, PTPRJ, PRUNE2, SHB, PRKCA | BMPR2, CSF1R,
PTPRJ, CDK2, TXLNA, VSX2, GPC3, OSM, PRKCA | - |
| hsa-miR-455-5p | LRP2, SOCS3 | FZD5, SOCS3, TJP1,
ETS1, GPI, KPNA1, KDR | LRP2, FZD5, KDR,
ETS1 PDGFRA, IRF2, SOX11 | - |
| hsa-miR-494 | BBC3, CNR1, SLC1A2,
PTEN, SIRT1 | BBC3, CNR1, ROCK1,
KPNA1, CUL3, FGFR2, IGF1R, GULP1, MTDH, UACA, PTEN, SIRT1,
INHBB | PHOX2B, FGFR2,
IGF1R, CUL3, IL12B, ARHGAP5, EVI5, GPNMB, PBRM1, RAP1B, PTEN,
TACC1, NFIB, PITX, SIRT1 | - |
|
hsa-miR-513a-5p | HMGCR, CHEK2,
SERP1, CDK6, GRB2 | RAG1, UNC5D,
ZNF346, EYA1, TRIM2, USP47, XIAP, MAPK7, CHEK2, NOD2 | MAGI2, PDS5B,
SMAD2, TBX19, KRAS, CBLB, CDK6, EYA1, NOD2, XIAP, ATF3, DDX11,
S1PR1, LIFR, VSX2 | - |
| hsa-miR-660 | - | TFAP2B, CDH13,
HIPK1 | TFAP2B, CDH13,
HIPK1, NAP1L1, LIFR | TFAP2B |
| Table IVGene ontology analysis of the
putative miRNA target genes. |
Table IV
Gene ontology analysis of the
putative miRNA target genes.
| Target genes of the
upregulated miRNAs |
|---|
|
|---|
| Gene ontology | % of totala |
|---|
| Negative regulation
of transcription | 20.0 |
| Positive regulation
of transcription | 22.6 |
| Nerve growth factor
receptor signaling pathway | 13.0 |
| Cell cycle | 19.1 |
| Blood
coagulation | 20.0 |
| Neural tube
closure | 5.2 |
|
| Target genes of the
downregulated miRNAs |
|
| Gene ontology | % of totala |
|
| Chromatic
modification | 9.0 |
| Positive regulation
of transcription | 14.2 |
| Negative regulation
of transcription | 9.5 |
| Cell division | 10.4 |
| Anti-apoptosis | 6.8 |
| Gene
expression | 11.2 |
| Response to DNA
damage stimulus | 4.4 |
| Cell cycle | 10.6 |
| Small
GTPase-mediated signal transduction | 8.4 |
| Ras protein signal
transduction | 3.3 |
| Activation of
pro-apoptotic gene products | 2.2 |
| Positive regulation
of osteoblast differentiation | 2.2 |
| Androgen receptor
signaling pathway | 2.5 |
| Regulation of
sequence-specific DNA binding transcription factor activity | 3.0 |
| BMP signaling
pathway | 2.5 |
In summary, to the best of our knowledge, we
demonstrated for the first time that EGCG protects against UVB
radiation by regulating specific miRNAs that putatively target
transcription- and cell survival-related genes in NHDFs. Although
additional studies must be performed to verify the predicted miRNA
target genes identified in this study, our results suggest that
characterization of EGCG-specific miRNA changes may provide a
useful approach to understanding cellular responses to EGCG in
UVB-induced NHDF damage.
Acknowledgements
This work was supported by the Ministry of
Education, Science and Technology (grant 20110028646 to S. An) of
the Republic of Korea.
References
|
1
|
McKay DL and Blumberg JB: The role of tea
in human health: an update. J Am Coll Nutr. 21:1–13. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bettuzzi S, Brausi M, Rizzi F, Castagnetti
G, Peracchia G and Corti A: Chemoprevention of human prostate
cancer by oral administration of green tea catechins in volunteers
with high-grade prostate intraepithelial neoplasia: a preliminary
report from a one-year proof-of-principle study. Cancer Res.
66:1234–1240. 2006.
|
|
3
|
Yang CS, Lambert JD, Hou Z, Ju J, Lu G and
Hao X: Molecular targets for the cancer preventive activity of tea
polyphenols. Mol Carcinog. 45:431–435. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stuart EC, Scandlyn MJ and Rosengren RJ:
Role of epigallocatechin gallate (EGCG) in the treatment of breast
and prostate cancer. Life Sci. 79:2329–2336. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dona M, Dell'Aica I, Calabrese F, et al:
Neutrophil restraint by green tea: inhibition of inflammation,
associated angiogenesis, and pulmonary fibrosis. J Immunol.
170:4335–4341. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cavet ME, Harrington KL, Vollmer TR, Ward
KW and Zhang JZ: Anti-inflammatory and anti-oxidative effects of
the green tea polyphenol epigallocatechin gallate in human corneal
epithelial cells. Mol Vis. 17:533–542. 2011.PubMed/NCBI
|
|
7
|
Katiyar SK, Afaq F, Perez A and Mukhtar H:
Green tea polyphenol (−)-epigallocatechin-3-gallate treatment of
human skin inhibits ultraviolet radiation-induced oxidative stress.
Carcinogenesis. 22:287–294. 2001.
|
|
8
|
Meeran SM, Mantena SK, Elmets CA and
Katiyar SK: (−)-Epigallocatechin-3-gallate prevents
photocarcinogenesis in mice through interleukin-12-dependent DNA
repair. Cancer Res. 66:5512–5520. 2006.
|
|
9
|
Kim SY, Kim DS, Kwon SB, et al: Protective
effects of EGCG on UVB-induced damage in living skin equivalents.
Arch Pharm Res. 28:784–790. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen W, Dong Z, Valcic S, Timmermann BN
and Bowden GT: Inhibition of ultraviolet B-induced c-fos gene
expression and p38 mitogen-activated protein kinase activation by
(−)-epigallocatechin gallate in a human keratinocyte cell line. Mol
Carcinog. 24:79–84. 1999.PubMed/NCBI
|
|
11
|
Xia J, Song X, Bi Z, Chu W and Wan Y:
UV-induced NF-κB activation and expression of IL-6 is attenuated by
(−)-epigallocatechin-3-gallate in cultured human keratinocytes
in vitro. Int J Mol Med. 16:943–950. 2005.
|
|
12
|
Bae JY, Choi JS, Choi YJ, et al:
(−)Epigallocatechin gallate hampers collagen destruction and
collagenase activation in ultraviolet-B-irradiated human dermal
fibroblasts: involvement of mitogen-activated protein kinase. Food
Chem Toxicol. 46:1298–1307. 2008.
|
|
13
|
Ambros V: microRNAs: tiny regulators with
great potential. Cell. 107:823–826. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yi R, Poy MN, Stoffel M and Fuchs E: A
skin microRNA promotes differentiation by repressing ‘stemness’.
Nature. 452:225–229. 2008.
|
|
16
|
Yi R, Pasolli HA, Landthaler M, et al:
DGCR8-dependent microRNA biogenesis is essential for skin
development. Proc Natl Acad Sci USA. 106:498–502. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou BR, Xu Y, Permatasari F, et al:
Characterization of the miRNA profile in UVB-irradiated normal
human keratinocytes. Exp Dermatol. 21:317–319. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakagawa H, Hasumi K, Woo JT, Nagai K and
Wachi M: Generation of hydrogen peroxide primarily contributes to
the induction of Fe(II)-dependent apoptosis in Jurkat cells by
(−)-epigallocatechin gallate. Carcinogenesis. 25:1567–1574.
2004.PubMed/NCBI
|
|
19
|
Muthusamy V and Piva TJ: The UV response
of the skin: a review of the MAPK, NFkappaB and TNFalpha signal
transduction pathways. Arch Dermatol Res. 302:5–17. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yao K, Zhang L, Zhang Y, Ye P and Zhu N:
The flavonoid, fisetin, inhibits UV radiation-induced oxidative
stress and the activation of NF-kappaB and MAPK signaling in human
lens epithelial cells. Mol Vis. 14:1865–1871. 2008.PubMed/NCBI
|
|
21
|
Gu M, Dhanalakshmi S, Mohan S, Singh RP
and Agarwal R: Silibinin inhibits ultraviolet B radiation-induced
mitogenic and survival signaling, and associated biological
responses in SKH-1 mouse skin. Carcinogenesis. 26:1404–1413. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim DS, Kim SY, Lee JE, et al:
Sphingosine-1-phosphate-induced ERK activation protects human
melanocytes from UVB-induced apoptosis. Arch Pharm Res. 26:739–746.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sharma SD, Meeran SM and Katiyar SK:
Dietary grape seed proanthocyanidins inhibit UVB-induced oxidative
stress and activation of mitogen-activated protein kinases and
nuclear factor-kappaB signaling in in vivo SKH-1 hairless mice. Mol
Cancer Ther. 6:995–1005. 2007. View Article : Google Scholar
|
|
24
|
Tsang WP and Kwok TT: Epigallocatechin
gallate up-regulation of miR-16 and induction of apoptosis in human
cancer cells. J Nutr Biochem. 21:140–146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang H, Bian S and Yang CS: Green tea
polyphenol EGCG suppresses lung cancer cell growth through
upregulating miR-210 expression caused by stabilizing HIF-1α.
Carcinogenesis. 32:1881–1889. 2011.PubMed/NCBI
|
|
26
|
Liu H and Kohane IS: Tissue and process
specific microRNA-mRNA co-expression in mammalian development and
malignancy. PLoS One. 4:e54362009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hu HY, Guo S, Xi J, et al: MicroRNA
expression and regulation in human, chimpanzee, and macaque brains.
PLoS Genet. 7:e10023272011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Uchida Y, Chiyomaru T, Enokida H, et al:
MiR-133a induces apoptosis through direct regulation of GSTP1 in
bladder cancer cell lines. Urol Oncol. Mar 9–2011.(Epub ahead of
print).
|
|
29
|
Incoronato M, Garofalo M, Urso L, et al:
miR-212 increases tumor necrosis factor-related apoptosis-inducing
ligand sensitivity in non-small cell lung cancer by targeting the
antiapoptotic protein PED. Cancer Res. 70:3638–3646. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shin S, Moon KC, Park KU and Ha E:
MicroRNA-513a-5p mediates TNF-α and LPS induced apoptosis via
downregulation of X-linked inhibitor of apoptotic protein in
endothelial cells. Biochimie. 94:1431–1436. 2012.PubMed/NCBI
|