Introduction
Human cervical cancer progression is strongly
associated with the infection of high-risk human papillomaviruses
(HR-HPVs) (e.g. HPV16 and 18), which are detected in nearly all
human cervical cancer (1). HPV is a
small, non-enveloped DNA virus expressing three key oncoproteins:
E5, E6 and E7, which possess the ability of transforming certain
human cells in vitro and are considered to be associated
with human cervical carcinogenesis in vivo(2). E6 and E7 are well known for their
ability to inhibit the function of tumor suppressor p53 and pRb,
respectively (3). However, the role
of E5 in cellular transformation is less understood. HPV16 E5
protein is a hydrophobic, 83-amino acid polypeptide that associates
with the Golgi apparatus, endoplasmic reticulum and perinuclear
membrane (4). E5 is capable of
altering growth and differentiation of epithelial cells via a
number of pathways, including conferring resistance to apoptosis
and affecting several cellular pathways involved in cell adhesion
and cell motility signaling (2,5,6).
Co-expression of E5 with either E6 or E7 promotes transformation to
a greater extent than with either oncoprotein alone (7). Although, multiple studies (8,9)
strongly suggest that E5 plays an important role in carcinogenesis
at the early stage of HPV-related cancer, the involvement of E5
oncoproteins in multi-step carcinogenesis remains poorly understood
and is an intriguing area of active study.
In the present study, we prepared four different
cell models to examine the function of the HR-HPV E5 protein. The
SiHa and HeLa cell lines were stably transfected with the whole
length of the HPV16 E5 or HPV18 E5 gene, respectively. Meanwhile,
we transfected C33A (an HPV-negative human cervical cancer cell
line) and HaCaT cells (immortal human keratinocytes used as
non-related control) with the whole length HPV16 E5, then screened
for stably expressed clones. We demonstrated that E5 protein and
mRNA levels increased in the transfected cells compared with the
controls. We proved that E5 promotes cell proliferation, migration
and invasion in the positive groups. In addition, using an in
vivo mouse tumor model, we provided evidence that increased E5
expression promotes both HPV or non-HPV-derived tumor growth.
Collectively, our findings indicate that HPV16 E5 plays a critical
role in human cervical cancer progression.
Materials and methods
Construction of recombinant DNA
expression vectors
HPV16/18 E5 genes were amplified using PCR from
plasmid pBR322-HPV16/18 that included the complete genome of
HPV16/18 (kindly provided by Professor H. zur Hausen, Heidelberg
University, Germany). PCR was performed using high-fidelity DNA
polymerase (Invitrogen Life Technologies, USA) in a 50-μl reaction
mixture with primers as shown in Table
I(10). The reaction conditions
were 30 cycles of denaturation at 94°C for 30 sec, annealing at
56°C for 30 sec and extension at 72°C for 45 sec. E5 PCR products
were ligated into pEGFP-C1 (Clontech, USA) downstream of their
respective CMV promoters. Plasmids were transformed into E.
coli DH5α competent cells and selected by kanamycin or
ampicillin resistance. Positive colonies were screened by PCR then
sequenced to confirm the identity of the DNA inserts.
 | Table IHPV16/18 E5 PCR primers. |
Table I
HPV16/18 E5 PCR primers.
HPV16 E5
| HPV18 E5
|
|---|
| F: CCCAAGCTTACTGCATCCACAACATTACTGGC | F: GCGAATTCCATGTTATCACTTATTTTTTTATTTTGC |
| R: GGGATCCATTATGTAATTAAAAAGCGTGCA | R: CCGGATCCCAACCTATACAATTACTGTAAAGACAA |
Preparation of stable cells expressing
HPV16/18 E5
Unless specified, all chemicals used in this study
are from regular commercial sources. Cells were maintained at 37°C
in a 5% CO2-95% O2 atmosphere. SiHa (human
cervical cancer cell line which contains an integrated HPV16 genome
from ATCC HTB-35™), HeLa (human cervical cancer cell line which
contains HPV18 sequences from ATCC CCL-2™), C33A (human cervical
cancer cell line which is negative for HPV DNA and RNA from ATCC
HTB-31™, to eliminate potential effects of other HPV parts as the
controls) and HaCaT cell lines (immortal human keratinocytes from
the Wuhan University Typical Object Preserve Center, China, as the
controls) were grown in DMEM containing 10% FBS and supplemented
with 100 U/ml of penicillin, 100 μg/ml of streptomycin and 0.25
μg/ml of amphotericin B. Cells were transfected with E5-containing
plasmids (HPV16 E5 was transfected to SiHa, C33A and HaCaT cells,
and HPV18 E5 was transfected to HeLa cells) or empty vectors (as
mock groups) using Lipofectin™ (Invitrogen Life Technologies) and
grown in DMEM/FBS containing 600-1,000 mg/l G418 (a dose cytotoxic
for non-transfected cells within 1 week) for 3 weeks. Individual
colonies of G418-selected cells were isolated and expanded in
DMEM/FBS. Cell lines were examined for E5 mRNA expression using an
adaptation reverse transcriptase-polymerase chain reactions
(RT-PCR) described by Biswas et al(11). The cells were tested for the
presence of the GFP-E5 fusion protein (no commercial E5 antibody)
by western blot analyses as described below.
Cell lysis and western blot analyses
Cell lysates were prepared in lysis buffer. Lysates
normalized for protein content (Bradford protein assay; Bio-Rad)
were prepared in Laemmli buffer, heated, subjected to sodium
dodecyl sulfate-polyacrylamide gel (SDS-PAGE) on 10% acrylamide
gels and transferred onto nitrocellulose membranes. For western
blot analyses, membranes were blocked in TBS (Tris-buffered saline)
and then incubated overnight at 4°C with indicated primary
antibodies [anti-p-paxillin antibody (Tyr118; Cell Signaling
Technology, Inc., USA), anti-E-cadherin antibody (BD Biosciences,
USA], and anti-β-actin antibody (Sigma-Aldrich, USA) to control
equal loading appropriately diluted in the blocking solution.
Finally, a signal was detected using ECL Western blotting detection
system (GE Healthcare, UK) and exposure to XR film (12).
Proliferation assays
Cell proliferation assays were performed using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
method. Cells (1×105) were plated in 96-well tissue
culture plates and grown to 70% confluence. Seventy-two hours later
the cells were stained with 5 mg/ml MTT reagent (Sigma-Aldrich).
The absorbance of each well was measured at a wavelength of 570 nm
and data were expressed as a raw OD at 570 nm or as a ratio of OD
at a specific time point over the initial OD on the first day.
Wells without cells but containing medium were used as a blank
value that was subtracted from all values. Each assay was performed
in triplicate.
Wound healing assays
Cell migration was assessed in classical wound
healing assays (12). Confluent
monolayer cells in a 6-well plate were wounded using a plastic
pipette tip (P200) and rinsed with PBS before being replaced in the
culture medium. The bottoms of the wells were marked to indicate
where the initial images of the wounded area were captured. After
24-h incubation at 37°C, images (x10) of the same areas were
recorded using an Axiovert 200M microscope (Zeiss) equipped with a
CoolSNAP™ ES camera (Photometric®; Roper Scientific) and
closure of the wound was evaluated using Metamorph®
(V6.3; Molecular Devices Corp.).
Cell invasion assays using
Transwells
In vitro invasion was determined in a
Matrigel-based Transwell assay essentially as previously described
by Pelletier et al(13). The
upper chambers of 24-well Transwell plates (Corning Costar,
Cambridge, MA, USA) with a pore size of 8 μm were coated with
Matrigel (0.7 mg/ml; Sigma-Aldrich) and the lower compartments were
filled with serum-free conditioned DMEM of NIH3T3. HPV16 or HPV18
E5 (HPV16/18 E5)-positive cells (1×105) re-suspended in
a 10% serum-containing medium were added to the top chamber of the
Transwell (8 μm, BD Biosciences, Franklin Lakes, NJ, USA)
pre-coated with Matrigel™ diluted in ice-cold PBS (175 μg/ml) at a
total of 35 μg/well and allowed to migrate for 24 h. Then, the
total number of invasive cells were estimated using Calcein AM (BD
Biosciences) as recommended by the manufacturer.
Immunofluorescence
Cells were stably transfected with pEGFP-C1 or
GFP-E5 and fixed as usual, followed by nuclear staining with
phospholine iodide (PI). To observe the cytoskeleton, cells were
blocked with 2.5% BSA plus 1% goat serum and doubled stained for
F-actin using tetramethylrhodamine B isothiocyanate-phalloidin
(Sigma-Aldrich). Cells were examined for fluorescence using laser
scanning microscopy (Olympus FluoView FV1000).
Tumor growth in vivo
HPV16/18 E5-positive cells were grown in DMEM/FBS to
80% confluency. Cells (5×105) resuspended in 200 μl of
physiological saline were injected subcutaneously in the right
flank of 6-week-old nude mice (Chinese Medical Science College,
Experiment Animal Research Center, China) (n=5 for each group). All
studies involving mice were approved by the Huazhong University of
Science and Technology Animal Care and Use Committee. Tumor
development was followed for 10 weeks. Mice were monitored twice a
week for tumor growth. Tumor size was measured every week with
calipers to assess tumor volume ([length × width2]/2).
The incidence of the tumors and the survival of mice were
recorded.
Statistical analysis
Statistical analyses and graphical presentations
were conducted by SPSS 13.0 and SigmaPlot 10.0. Results are
presented as the means ± standard error of the mean (SEM).
Statistical analysis of significance was based on analysis of
variance or χ2 analysis; P-values <0.05 were
considered to indicate a statistically significant difference.
Results
Analysis of the expression level of E5
before and after transfected in different cell models
To explore the role of HPV16/18 E5, we prepared four
cell lines stably expressing HPV16/18 E5 as described in Materials
and methods. As a result, HPV16/18 E5 was expressed as a GFP fusion
protein.
To determine the expression of HPV16/18 E5 before
and after transfection, we compared both instances by RT-PCR. As
shown in Fig. 1A, we observed a
high level of HPV16/18 E5 in all the transfected cells. HPV18 E5
also appeared at low levels in the mock (transfected with empty
plasmids, as the blank control) and untransfected HeLa cells. To
further substantiate these results the protein extracts were
blotted with the GFP antibody. We discovered bands with similar
molecular weights (GFP+E5) as shown in Fig. 1B. Further analyses of the
subcellular localization of the GFP+E5 fusion protein in HPV18 E5
stably transfected HeLa cells revealed that most of the GFP+E5
fluorescence accumulated at the perinuclear region of the cells
(Fig. 1C).
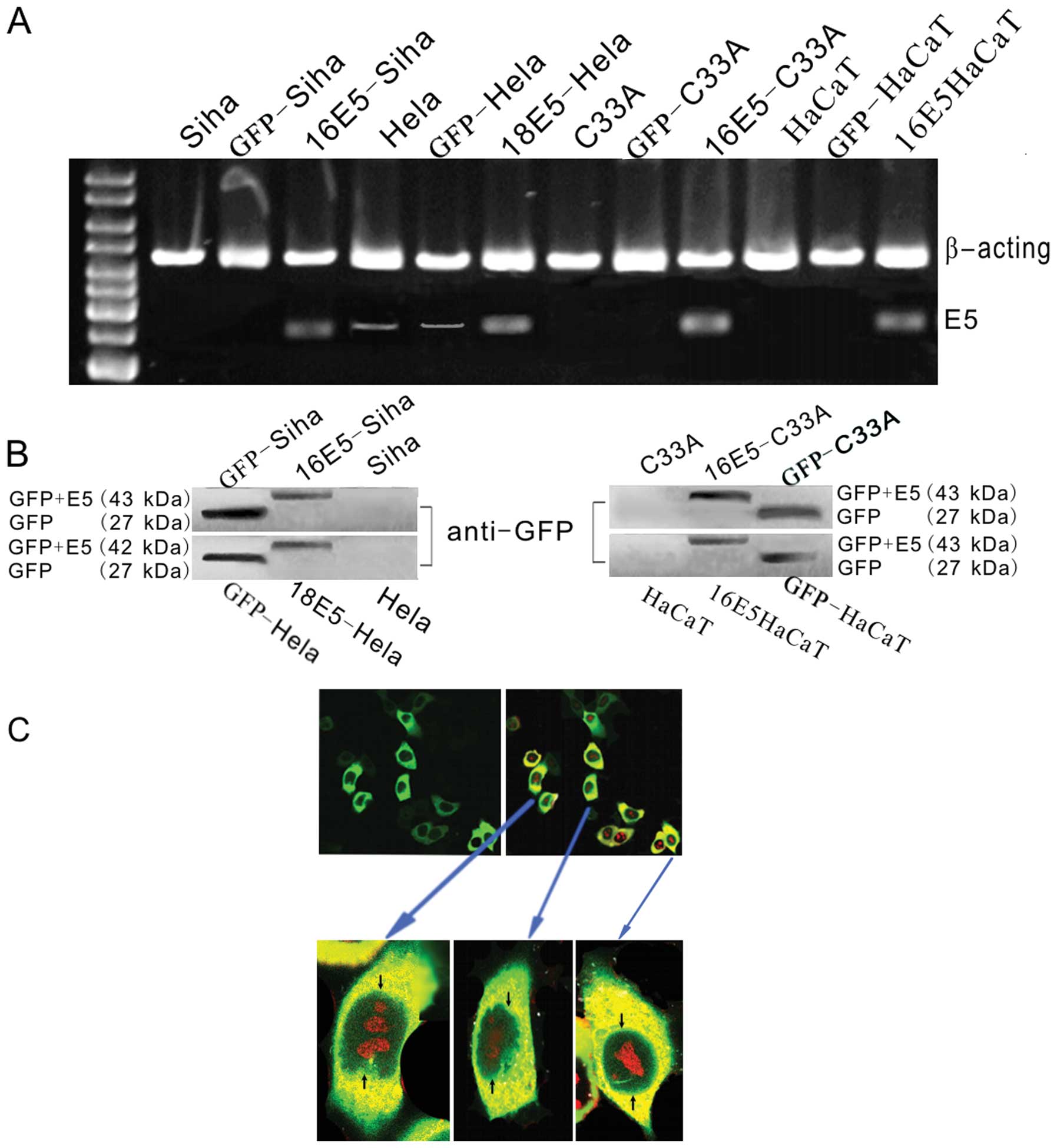 | Figure 1Analysis of the expression of HPV16/18
E5 in the recombinant plasmids. (A) RT-PCR for HPV16/18 E5
expression in different cell lines. In the E5-stably transfected
cells (16 E5-SiHa, 18 E5-HeLa, 16 E5-C33A and 16 E5-HaCaT), E5 was
detected. In the empty plasmid pEGFP-C1-transfected groups
(GFP-SiHa, GFP-HeLa, GFP-C33A and GFP-HaCaT), E5 was not detected.
In the untreated SiHa, HeLa, C33A, HaCaT cell lines, an
insignificant E5 expression in HeLa cells before transfection was
detected while in the other groups no clear E5 band was observed.
(B) Western blot analysis of the HPV16/18 E5 by GFP fusion
recombinants. Using the anti-GFP antibodies, a positive band was
noted with a molecular weight corresponding to GFP + HPV16/18 E5
(27+16/15 kDa) in the E5-stably transfected cells. In the empty
plasmid pEGFP-C1-transfected groups, only a GFP band (27 kDa) was
detected. In the untreated SiHa, HeLa, C33A and HaCaT cell lines, a
clear band was not discovered. (C) Immunofluorescence for the
subcellular localization of E5, photographed by a confocal
fluorescence microscope. Most of the fluorescence accumulated at
the perinuclear region of the 18 E5-HeLa cells stably transfected
with pEGFP-C1-HPV18 E5 (cell nuclear staining was performed using
phospholine iodide). |
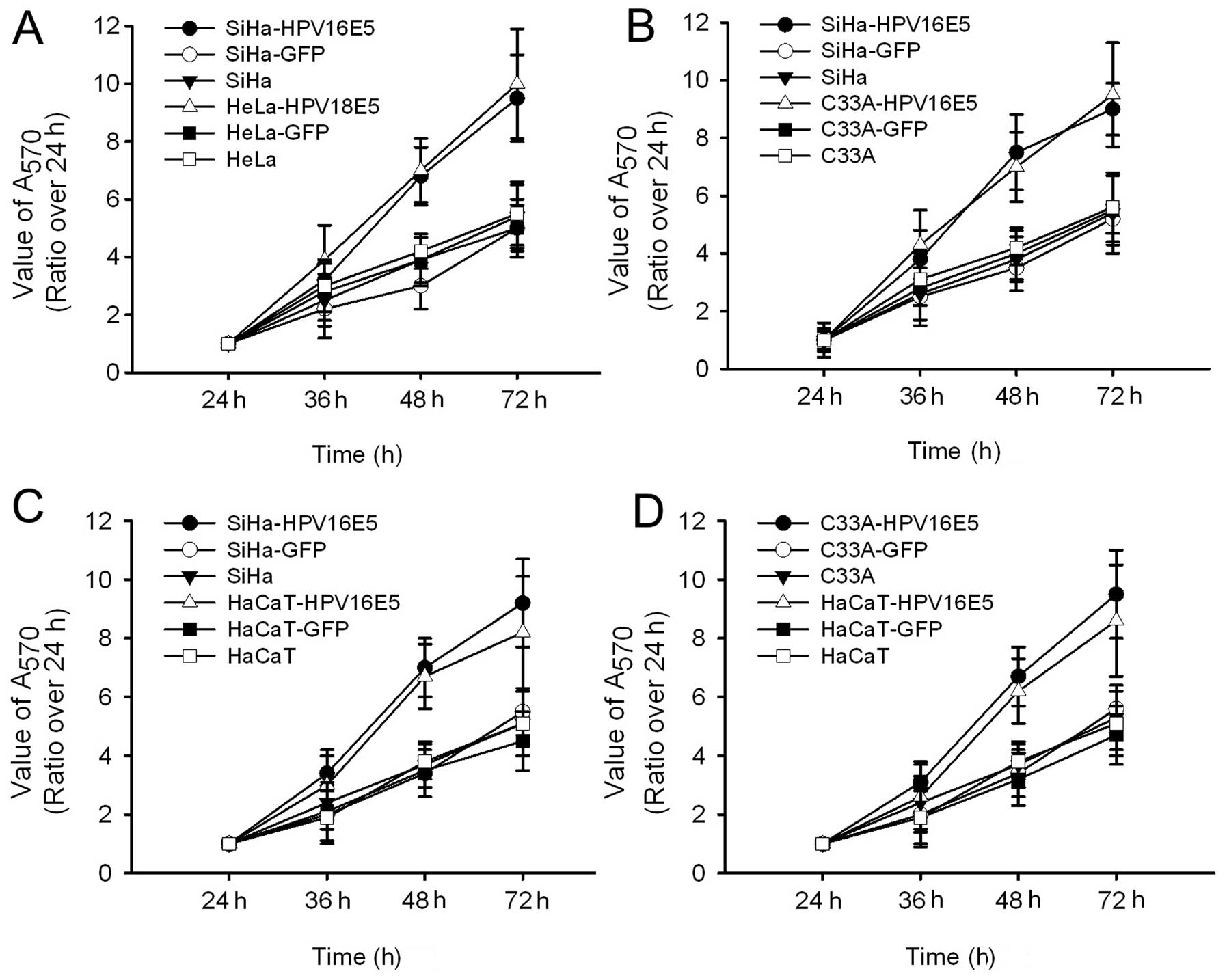
HPV16/18 E5 promotes cell
proliferation
To demonstrate the role of HPV16/18 E5 in cell
functions, we first examined whether E5 regulates cell
proliferation in different cell lines. We found that overexpression
of HPV16/18 E5 significantly enhanced cell proliferation and showed
higher proliferative abilities compared to its paired groups
(Fig. 2). As expected, we found no
significant difference in HPV16 E5 and HPV18 E5 (Fig. 2A).
To assess whether other HPV parts affect cell
proliferation, we used C33A-HPV16 E5 as HPV negative controls and
HaCaT-HPV16 E5 as the non-related controls to eliminate any
influence from other HPV parts. As demonstrated in Fig. 2B-D we demonstrated that the cell
proliferation ability had no significant difference in the
HPV-positive or -negative groups. Together these results indicate
that HPV16/18 E5 contributes to the control of cell
proliferation.
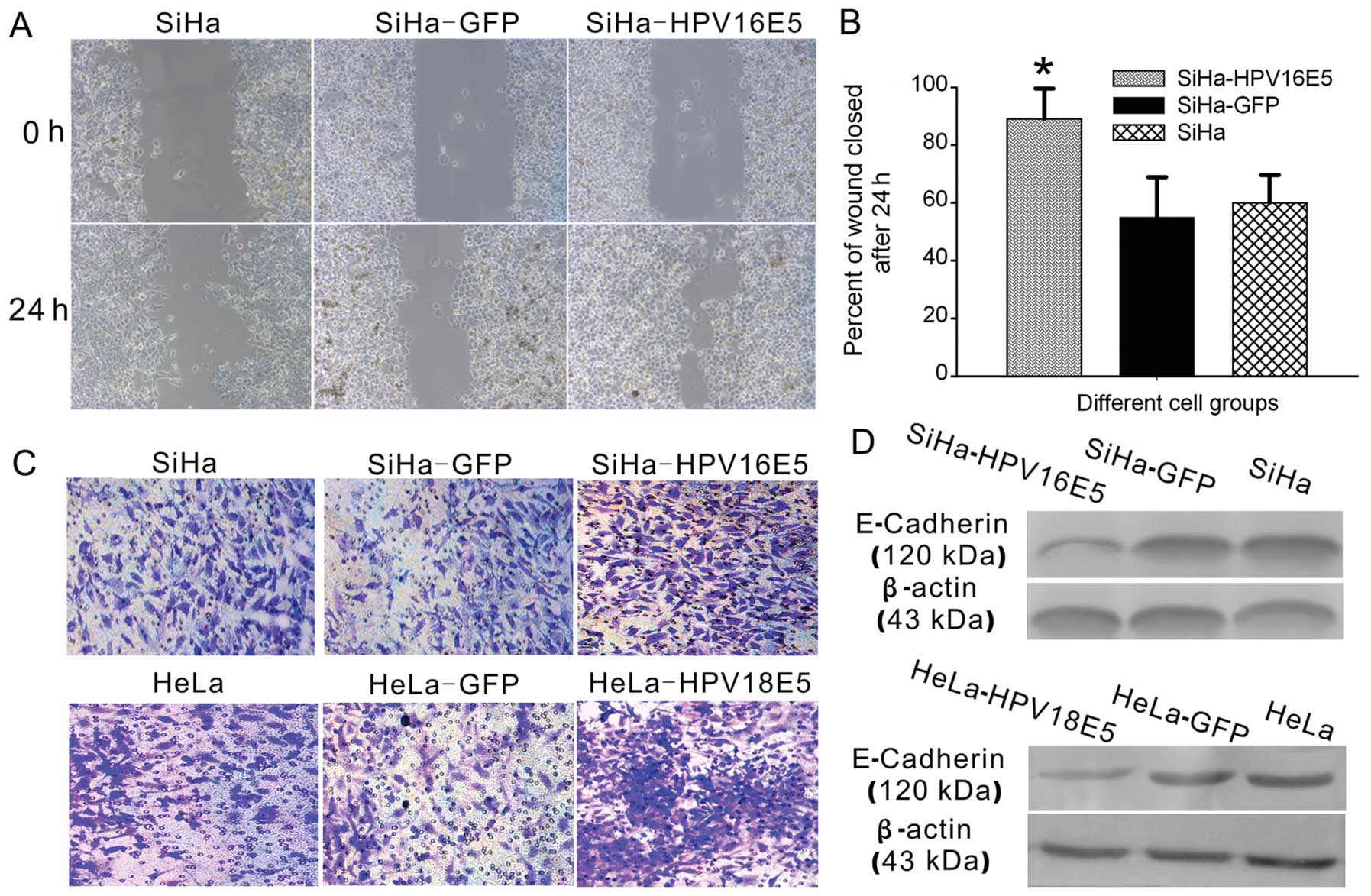
HPV16/18 E5 modulates migration and
invasion of human cervical cancer cells
Cancer progression involves the transformation of
cells to acquire motility and invasive abilities (12). Therefore, we next explored whether
HPV16/18 E5 is critical to human cervical cancer cell migration and
invasion. We compared the migration of SiHa-HPV16 E5 with SiHa-GFP
cells and the untreated SiHa cells in wound healing assays. As
shown in Fig. 3A and B, increasing
levels of HPV16 E5 in human cervical cancer cells promoted
migration and this was significant in SiHa-HPV16 E5 groups compared
with the controls. We then compared the migration of HeLa-HPV18 E5
cells with HeLa-GFP cells or the untreated HeLa cells and obtained
similar results.
To determine whether the expression of HPV16/18 E5
promotes invasion in a tumor-like context, we evaluated invasion of
human cervical cancer cells through Matrigel™ matrix in Transwells
assays with a collagen type I matrix (Fig. 3C). As expected, human cervical
cancer cells expressing HPV16/18 E5, including SiHa-HPV16 E5 and
HeLa-HPV18 E5 cells, grew significantly outward after 48 h in
culture. In contrast, in empty plasmid-transfected cells (SiHa-GFP
and HeLa-GFP) or untreated SiHa and HeLa cells the number of
invasive cells were less.
To explore the mechanism involved we studied whether
HPV16/18 E5 affected the expression of E-cadherin. E-cadherin
expression in cancer cells is associated with cancer progression,
invasion, metastasis and cytoskeleton rearrangement (14). E-cadherin expression was
significantly downregulated in SiHa-HPV16 E5/HeLa-HPV18 E5 cells
compared to the control cells (Fig.
3D). Together these results suggest that HPV16/18 E5 promotes
cell migration and invasion in human cervical cancer cells through
the E-cadherin pathway.
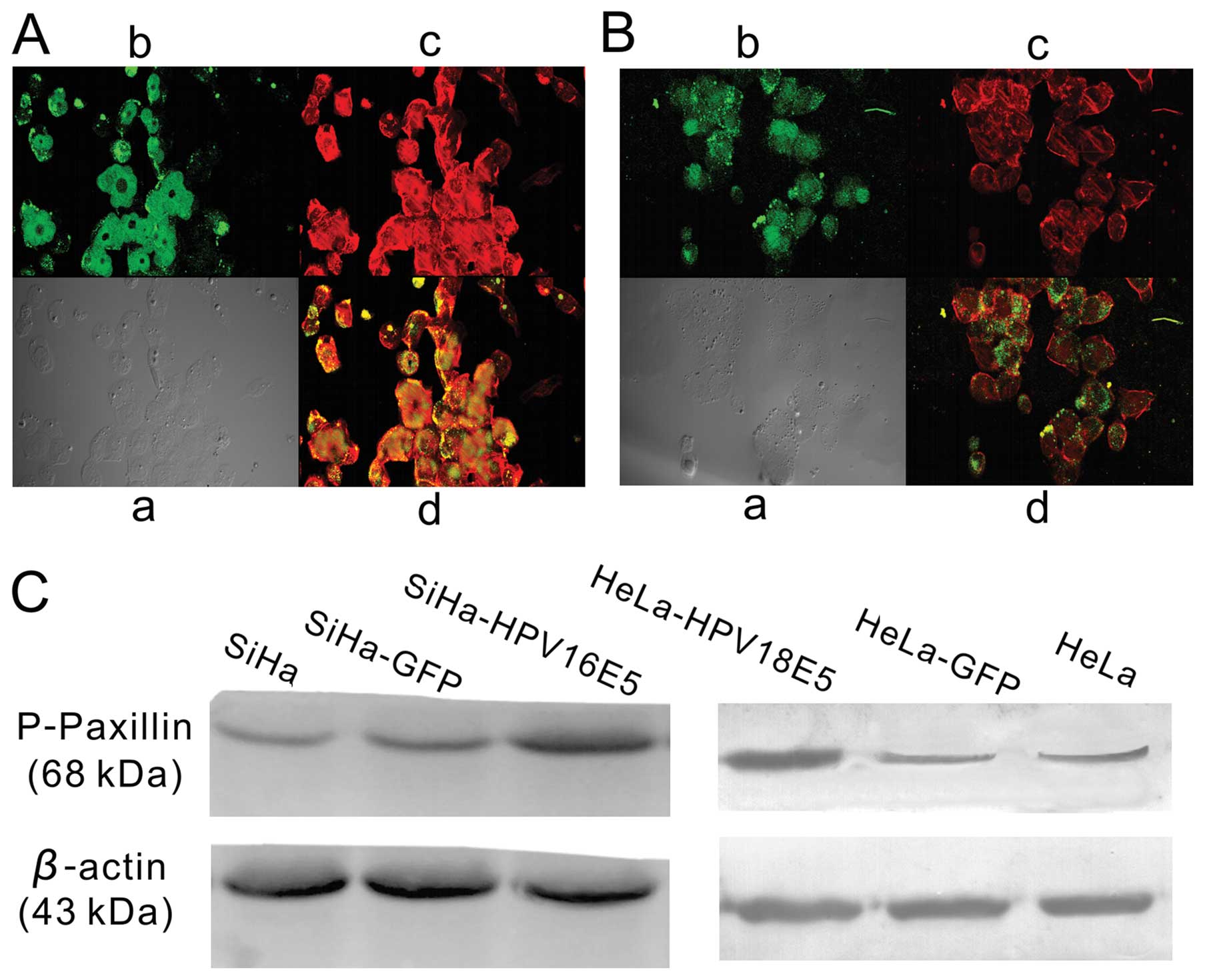
HPV16/18 E5 affects the cytoskeleton of
human cervical cancer cells
The actin cytoskeleton is critical for maintaining
cell morphology and is required for cell motility. HPV16/18 E5
played an important role in regulating cell proliferation,
migration and invasion as demonstrated above. Furthermore, we
compared actin staining in SiHa cells expressing either GFP+E5
(Fig. 4A) or GFP (Fig. 4B). We discovered that in SiHa-HPV16
E5 cells stably transformed by E5, the phalloidin staining revealed
multiple heavy actin cables brightly stained (Fig. 4A) whereas vector-transfected
SiHa-GFP cells contained only a few thin actin fibers (Fig. 4B). We compared the actin
cytoskeleton of HeLa-HPV18 E5 with HeLa-GFP cells, and obtained
exactly the same results.
Tyrosine phosphorylation of paxillin (p-paxillin)
plays a key role in the regulation of actin cytoskeleton
organization (15), As expected, we
demonstrated the increased expression of p-paxillin in SiHa-HPV16
E5/HeLa-HPV18 E5 cells as compared to the control cells (Fig. 4C). Collectively, these results
revealed that the increased expression of HPV16/18 E5 in human
cervical cancer cells significantly enhance cell motility.
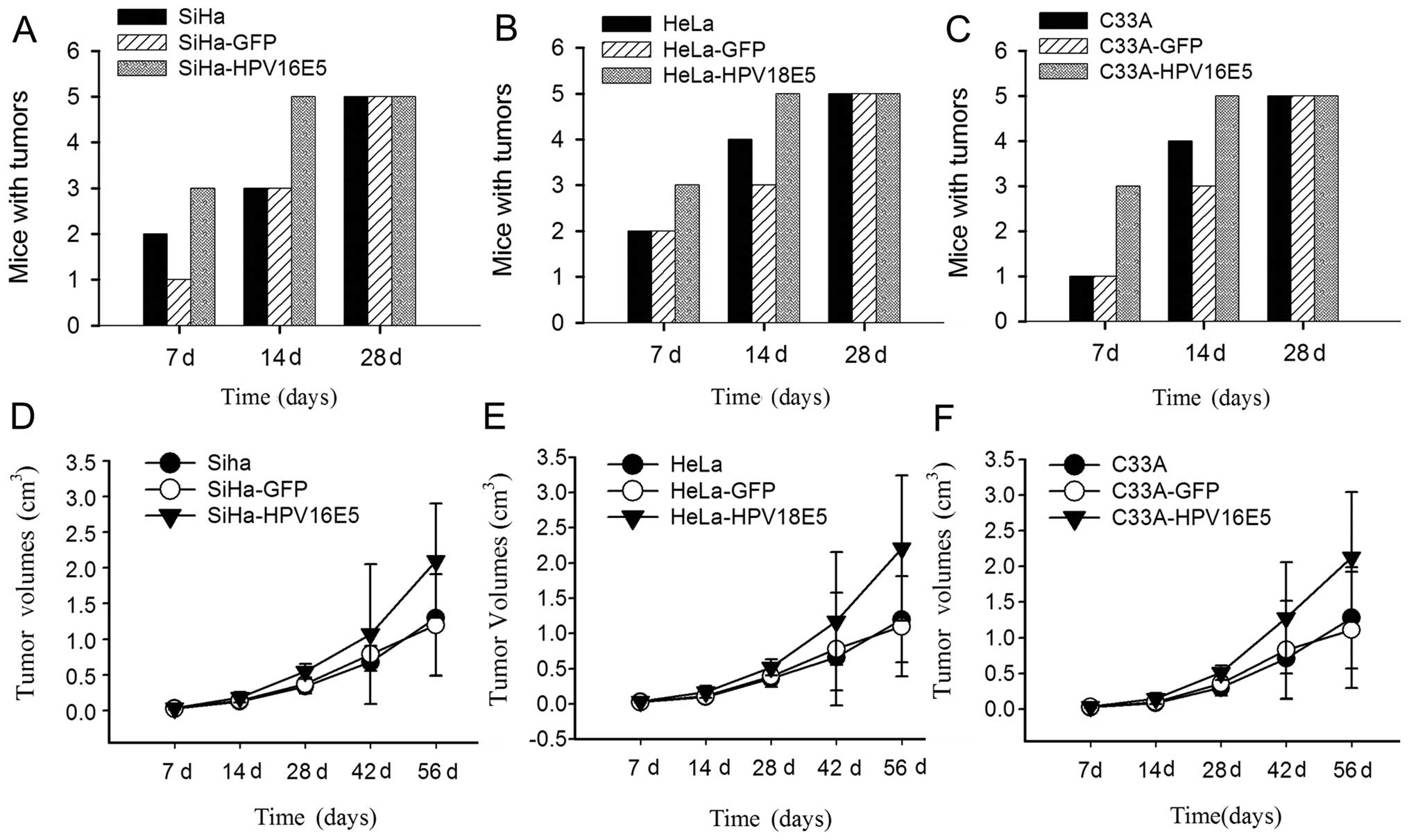
HPV16/18 E5 promotes tumor growth in
vivo
To establish the biological relevance of our
findings (12), we examined whether
the overexpression of HPV16/18 E5 in human cervical cancer cells
confers tumorigenic advantage in subcutaneous tumors using a mouse
model. To examine this, SiHa-HPV16 E5 and HeLa-HPV18 E5 cells,
parental SiHa or HeLa cells, SiHa-GFP or HeLa-GFP were injected
subcutaneously into nude mice. Tumors derived from SiHa-HPV16 E5
and HeLa-HPV18 E5 cells (Fig. 5A and
B) grew more rapidly and reached the maximal volume allowed
during 8 weeks post-injection compared with parental SiHa and HeLa
cells or SiHa-GFP and HeLa -GFP treated groups (Fig. 5D and E). Together, these
observations suggest that the increased expression of HPV16/18 E5
in human cervical cancer cells not only increases the tumor volume
but also accelerates the tumor growth rate both in HPV- and
non-HPV-derived tumors in vivo (Fig. 5C and 5F). Due to the appearance of a
spontaneous tumor necrosis core, mice were required to be
euthanized 8 weeks post-injection. Nevertheless, in line with our
previous in vitro studies, these results strongly support
the important role of HPV16/18 E5 in tumor growth in
vivo.
Discussion
Recent findings implicate HPV16 E5 as an important
mediator of oncogenic transformation (16). In this study, we provide evidence
that HPV16/18 E5 plays a role in promoting the proliferation,
migration and invasion of human cervical cancer cells in
vitro and accelerated the growth of human cervical
cancer-derived tumors in vivo. Our investigation revealed
that E5 overexpression induces cell proliferation in both
HPV-positive and -negative groups pointing towards E5 sufficiency
to promote cell growth (Fig. 2).
Venuti et al(17) suggested
that the expression of E5 is increased upon differentiation to
promote proliferation of differentiated cells and productive viral
replication. The localization of E5 was observed in internal cell
membranes (Fig. 1), as previously
reported (4). We suggest that the
localization of HPV E5 to the endoplasmic reticulum indicated that
its activity may be related to the trafficking of cytoplasmic
membrane proteins through this cellular compartment.
To explore the possible mechanism involved, we
performed a thorough investigation. In this study, we observed that
HPV16/18 E5 overexpression affects cell migration, invasion and
significantly downregulated E-cadherin protein expression (Fig. 3). E-cadherin, found at adherens
junctions, is the principal effector of cell-cell adhesion
(18). E-cadherin impairment
represents the hallmark of malignancy and is strongly associated
with poor prognosis of a number of tumors (19). Loss of E-cadherin expression in
cancer cells weakens cell-cell adhesion and is associated with
cancer progression, invasion, metastasis and cytoskeleton
rearrangement (14). Our results
suggest that the enhancement of migration and invasion in HPV16/18
E5-overexpressing human cervical cancer cells may be, at least
partly, due to the E-cadherin downregulation. Further investigation
is required to elucidate the molecular events responsible for the
association of E-cadherin and E5, and this is an ongoing focus of
our research
Transformed cells commonly exhibit altered
morphology and reduced cell adherence due to the disruption of
cytoskeletal structures (15) and
cell migration is strictly regulated by the re-organization of the
actin cytoskeleton (20). It may be
possible that E5 was directly or indirectly involved in these
processes. Our investigation (rhodamine-phalloidin was used to
label F-actin) (Fig. 4)
demonstrated that the cell shape of the clones overexpressing
HPV16/18 E5, revealed polygonal cell shapes, which was brightly
stained and longitudinal actin bundles were formed. In contrast to
the control cells, we observed shrunken cell shapes and stress
fibers were almost invisible. Previous studies revealed that tumor
cells with different motile activities differ in terms of
morphology and that these differences may be attributable to the
reorganization of the actin cytoskeleton (21). Tyrosine phosphorylation of paxillin
(p-paxillin) was found to be involved in the regulation of actin
cytoskeleton organization (12). In
this study, we discovered that the overexpression of HPV16/18 E5
upregulated p-paxillin and may in this manner, contribute to cancer
progression by altering signaling pathways regulating the actin
cytoskeleton network supporting cell migration. We hypothesize that
the downregulation of E-cadherin and the upregulation of p-paxillin
by HPV16/18 E5 may endow human cervical cancer cells with altered
spatial relationships that favor uncontrolled proliferation,
migration and invasion.
The role of HPV16/18 E5 in carcinogenesis seems to
be limited to the early stages of cervical carcinogenesis since the
E5 gene is frequently deleted when the HPV genome is integrated
during malignant progression (22).
Therefore, targeting E5 which is frequently expressed in earlier
stages of malignant transformation may be a rational approach for
preventing premalignant lesions from progressing into invasive
human cervical cancer and may be advantageous particularly in the
early stage of HPV infections and pre-cancerous lesions.
Acknowledgements
This study was partially supported by the National
Science Foundation of China (30901586, 30973205, 81172464), The
‘973’ Program of China (2009CB521800), the Huibei Province Science
Fund (2011CDB191) and the Central University Basic Science Research
Fund (2012QN188).
Abbreviations:
|
HR-HPV
|
high-risk human papillomavirus
|
|
RT
|
reverse transcriptase
|
|
PCR
|
polymerase chain reaction
|
|
SDS-PAGE
|
sodium dodecyl sulfate-polyacrylamide
gel
|
|
CIN
|
cervical intraepithelial neoplasm
|
|
FACS
|
fluorescence activated cell sorter
|
|
p-paxillin
|
phosphorylated paxillin
|
References
|
1
|
Zur Hausen H: Papillomavirus infections -
a major cause of human cancers. Biochim Biophys Acta. 1288:F55–F78.
1996.PubMed/NCBI
|
|
2
|
Kivi N, Greco D, Auvinen P and Auvinen E:
Genes involved in cell adhesion, cell motility and mitogenic
signaling are altered due to HPV 16 E5 protein expression.
Oncogene. 27:2532–2541. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Narisawa-Saito M and Kiyono T: Basic
mechanisms of high-risk human papillomavirus-induced
carcinogenesis: roles of E6 and E7 proteins. Cancer Sci.
98:1505–1511. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krawczyk E, Suprynowicz FA, Sudarshan SR
and Schlegel R: Membrane orientation of the human papillomavirus
type 16 E5 oncoprotein. J Virol. 84:1696–1703. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pedroza-Saavedra A, Lam EW,
Esquivel-Guadarrama F and Gutierrez-Xicotencatl L: The human
papillomavirus type 16 E5 oncoprotein synergizes with EGF-receptor
signaling to enhance cell cycle progression and the down-regulation
of p27(Kip1). Virology. 400:44–52. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kabsch K, Mossadegh N, Kohl A, Komposch G,
Schenkel J, Alonso A and Tomakidi P: The HPV-16 E5 protein inhibits
TRAIL- and FasL-mediated apoptosis in human keratinocyte raft
cultures. Intervirology. 47:48–56. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stöppler MC, Straight SW, Tsao G, Schlegel
R and McCance DJ: The E5 gene of HPV-16 enhances keratinocyte
immortalization by full-length DNA. Virology. 223:251–254.
1996.PubMed/NCBI
|
|
8
|
Gao P and Zheng J: High-risk HPV
E5-induced cell fusion: a critical initiating event in the early
stage of HPV-associated cervical cancer. Virol J. 7:2382010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Boulenouar S, Weyn C, Van Noppen M, et al:
Effects of HPV-16 E5, E6 and E7 proteins on survival, adhesion,
migration and invasion of trophoblastic cells. Carcinogenesis.
31:473–480. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kozak M: At least six nucleotides
preceding the AUG initiator codon enhance translation in mammalian
cells. J Mol Biol. 196:947–950. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Biswas C, Kell B, Mant C, et al: Detection
of human papillomavirus type 16 early-gene transcription by reverse
transcription-PCR is associated with abnormal cervical cytology. J
Clin Microbiol. 35:1560–1564. 1997.PubMed/NCBI
|
|
12
|
Labelle-Côté M, Dusseault J, Ismaïl S, et
al: Nck2 promotes human melanoma cell proliferation, migration and
invasion in vitro and primary melanoma-derived tumor growth in
vivo. BMC Cancer. 11:4432011.PubMed/NCBI
|
|
13
|
Pelletier AJ, Kunicki T and Quaranta V:
Activation of the integrin alpha v beta 3 involves a discrete
cation-binding site that regulates conformation. J Biol Chem.
271:1364–1370. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Perl AK, Wilgenbus P, Dahl U, Semb H and
Christofori G: A causal role for E-cadherin in the transition from
adenoma to carcinoma. Nature. 392:190–193. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tong X and Howley PM: The bovine
papillomavirus E6 oncoprotein interacts with paxillin and disrupts
the actin cytoskeleton. Proc Natl Acad Sci USA. 94:4412–4417. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moody CA and Laimins LA: Human
papillomavirus oncoproteins: pathways to transformation. Nat Rev
Cancer. 10:550–560. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Venuti A, Paolini F, Nasir L, et al:
Papillomavirus E5: the smallest oncoprotein with many functions.
Mol Cancer. 10:1402011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takeichi M: Morphogenetic roles of classic
cadherins. Curr Opin Cell Biol. 7:619–627. 1995. View Article : Google Scholar
|
|
19
|
Wells A, Yates C and Shepard CR:
E-cadherin as an indicator of mesenchymal to epithelial reverting
transitions during the metastatic seeding of disseminated
carcinomas. Clin Exp Metastasis. 25:621–628. 2008. View Article : Google Scholar
|
|
20
|
Friedl P and Wolf K: Tumor-cell invasion
and migration: diversity and escape mechanisms. Nat Rev Cancer.
3:362–374. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nemethova M, Auinger S and Small J:
Building the actin cytoskeleton: filopodia contribute to the
construction of contractile bundles in the lamella. J Cell Biol.
180:1233–1244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chang JL, Tsao YP, Liu DW, et al: The
expression of HPV-16 E5 protein in squamous neoplastic changes in
the uterine cervix. J Biomed Sci. 8:206–213. 2001. View Article : Google Scholar : PubMed/NCBI
|