Introduction
A number of epidemiological studies have provided
indications that workers chronically exposed to medium-to-high
levels of extremely low frequency (ELF) magnetic fields (MFs),
including those of a power frequency 50–60 Hz have an increased
risk of developing various types of cancer (1–3) or
degenerative diseases (4,5), although other studies have not
detected such increases in risk (6,7). The
International Agency for Research on Cancer (IARC) has classified
ELF MF exposure as a ‘possible carcinogen to humans’ (8) on the basis of the epidemiological and
experimental evidence, particularly on adult and childhood
leukemia. Nevertheless, uncertainty persists on this matter, and
other international bodies, including the International Commission
on Non-Ionizing Radiation Protection (ICNIRP), consider that the
current evidence on the carcinogenic potential of weak ELF fields
(B≤0.5 mT at 50 Hz) is insufficient, primarily due to the present
lack of a mechanistic basis that can explain the phenomena
underlying the biological interactions of those fields (9,10).
Experimental studies in vivo have yielded inconclusive or
contradictory results. The lack of properly standardized exposure
parameters may have contributed to such negative or inconsistent
results (11–13). It should, moreover, be considered
that animal models may be unsuitable for studies examining various
aspects of the carcinogenic potential of weak ELF fields (14). As for the cellular response, a large
number of studies have reported in vitro effects of an
ELF-MF on different cellular processes in different cell species
(reviewed in ref. 15). However, to
date there is no general agreement on the specific primary
biological targets of power frequency MFs, on the biophysical and
chemical basis of the MF interactions, nor to which extent the
reported in vitro bioeffects may be indicative of a human
susceptibility to these fields. Concerning potential targets
related to cancer risk, ELF MFs have been shown to alter gene and
protein expression (16–18) as well as intracellular calcium
concentration and apoptosis rates (19–23)
and to induce oxidative stress (24–26)
and DNA damage (27–31). The interest in the potential adverse
effects of power frequency MFs has also been focused on the
possibility that these fields may influence tumor promotion by
increasing the rate of cell proliferation and/or modifying the
activity of molecules implicated in their regulation. However, the
number of studies examining these processes under exposure to low
magnetic flux densities, B≤100 μT, which can be found in certain
occupational environments, is rather scarce. On the other hand, it
has been described that the cellular alterations due to ELF-MF
exposure, combined with certain risk factors and/or in combination
with the action of other physiological or environmental agents, may
influence tumorigenic processes (32,33).
In this respect, we previously reported that a 50-Hz
MF at 100 μT significantly increased the cell number and BrdU
incorporation into DNA in the human cell lines NB69 (neuroblastoma)
and HepG2 (hepatocarcinoma). These effects were modulated by
all-trans retinol, the metabolic precursor of retinoic acid
(34). The present study assessed
the responsiveness of the NB69 cell line exposed to a 50-Hz MF at
10 and 100 μT in combination with the retinoid retinoic acid (RA),
in regards to cell proliferation. RA plays a crucial role in the
growth and differentiation of normal, premalignant and malignant
tissues, and has received significant attention because of its
potential interest in cancer therapy (35–37).
RA is currently being used as a tool in standard treatment
protocols for high-risk neuroblastomas, and is one of the
well-established inducers of neuronal differentiation and/or
apoptosis in neuroblastoma cells (38–40).
For example, in the neuroblastoma cell line LAN-5, exposure to a
50-Hz MF at 1 mT was reported to exert an antagonistic effect on
cell differentiation induced by 5 μM RA (24). The flux density of 10 μT tested in
the present study, is found in certain occupational environments
(41), corresponds to 10 and 2%,
respectively, of the reference levels proposed by the ICNIRP for
the protection of the general public (100 μT) and workers (500 μT)
against the harmful effects of short-term exposure to a 50-Hz MF
(9,10). In the present study, as an initial
step to determine whether an MF at 10 μT induces proliferative
effects similar to those we reported at 100 μT, the potential
changes in cell viability and proliferation were evaluated by cell
counting (trypan blue exclusion), by spectrophotometric analysis of
total protein and DNA content and by the immunocytochemical
expression of the proliferative marker, proliferating cell nuclear
antigen (PCNA). Subsequently, the responsiveness of the NB69 cell
line to MF at 10 and 100 μT when in the presence of RA was assessed
in regards to cell viability and proliferation.
Materials and methods
Magnetic field exposure
Fifty-hertz, sine wave magnetic fields at 10 or 100
μT were generated by a set of coil pairs in a Helmholtz
configuration and energized using a Newtronics Model 200MSTPC wave
generator (Madrid, Spain). The exposure set-up used in these
experiments was based on that developed by Blackman et
al(42), and has been described
elsewhere (34). Briefly, each of
the two identical sets consisted of a pair of 1000-turn, 20-cm
diameter coils of enamelled copper wire, aligned coaxially 10 cm
apart and oriented to produce vertically polarized magnetic fields.
Five or twenty 60-mm Petri dishes (Nunc, Spain) were placed in the
uniform MF space within the coils for exposure or sham-exposure.
Currents in the coils were adjusted and monitored using a
multimeter (model 974A; Hewlett Packard, Loveland, CO, USA) after
establishing the DC and AC flux densities with fluxgate
magnetometers (Bartington model Mag-3; GMW Associates, San Carlos,
CA, USA and EFA-3 model BN 2245/90.20; Wandel & Goltermann S.A,
Eningen, Germany). The coil sets were mounted in the center of
magnetically shielded, Co-Netic alloy boxes (Amuneal Corp.,
Philadelphia, PA, USA) housed in incubators (Forma models 3121 and
3194) in an atmosphere of 5% CO2 at 37°C. The magnetic
shielding allowed for reduced environmental fields at the sample
locations, with DC MF=0.02–0.08 μT (rms) and 50 Hz AC MF=0.06–0.09
μT (rms). Two identical sets of coils, shielding rooms and
incubators were used. In each experimental run, only one set of
coils was energized at random. The samples in the unenergized set
were considered sham-exposed controls. The exposure settings and
the dosimetry were assessed and validated immediately before and
after each experimental run.
Cell culture
The NB69 human neuroblastoma cell line was provided
by Dr M.A. Mena (Hospital Ramón y Cajal, Madrid, Spain). The cells
were plated at a density of 4.5×104 cells/ml in Petri
dishes and grown in Dulbecco’s minimum Essential medium (DMEM;
Gibco, Carlsbad, CA, USA) supplemented with 15% heat inactivated
fetal calf serum (FCS, Gibco), 4 mM L-glutamine and 100 U/ml
penicillin plus 100 U/ml streptomycin. In each experimental run,
cells seeded in groups of 10 or 40 dishes were grown for three days
in humidified incubators with an atmosphere of 5% CO2 at
37°C. When appropriate, the medium was supplemented at plating with
2 μM RA (all-trans-retinol; Worthington Biochemical Corp.,
Lakewood, NJ, USA), or with the corresponding vehicle, absolute
ethanol (controls). At day 3 post-plating the cultures received
fresh medium supplemented with a second dose of 2 μM RA or with the
matched vehicle concentration.
Assessment of cellular response to
RA
Cells were seeded and supplemented with RA
concentrations within the physiological range in mammals: 0.5, 1.0,
2.0 or 5.0 μM. RA untreated controls, supplemented with the highest
vehicle concentration used in these experiments, were also included
in each run. Seventy-two hours after plating, the medium was
renewed and supplemented with the corresponding concentrations of
RA or vehicle. Each concentration was quadruplicate tested (a total
of 24 Petri dishes/experimental replicate). A total of 3
experimental replicates were carried out. At the end of five days
of incubation in the presence of RA or vehicle, the cells were
collected, counted and analyzed for cell viability and
proliferation.
Exposure to a 50-Hz MF
Two series of experiments were conducted. In the
first series, a total of 10 experimental replicates at a magnetic
flux density B=10 μT, and 5 replicates at 100 μT, were carried out.
In each experimental replicate 10 dishes with cells were grown for
3 days inside a commercial incubator. On day 3, the medium was
renewed and each group of 5 dishes was transferred to one of two
identical incubators, (atmosphere of 5% CO2 at 37°C and
100% humidity) with the shielded chambers and Helmholtz coils
inside, as previously described. These incubators were used for
MF-exposure or sham-exposure in alternating runs. The MF-treated
groups were exposed to the fields intermittently, 3 h on/3 h off,
for 42 h.
In the second experimental series, a total of 9
replicates, 4 at B=10 μT and 5 at 100 μT, were carried out. In each
replicate a total of 40 dishes (20 dishes for the 42-h studies and
20 for the 90-h studies) was distributed in groups of 5 Petri
dishes and exposed or sham-exposed to RA and/or MF, according to
the following combinations: MF−/RA-, MF−/RA+, MF+/RA- and MF+/RA+.
The MF and RA treatments, as well as the respective
sham-treatments, were applied according to the procedures
previously described. At the end of the 42-h or 90-h exposure
and/or incubation, the cell growth and viability of the NB69 cells
were determined using a hemocytometer. Spectrophotometric analysis
of total protein and DNA contents were conducted following the
methods described below.
Cell counting and spectrophotometric
analysis of protein and DNA contents
In all experiments the MF was applied at day 3
post-plating. The cell counting was performed at the end of 42 h or
90 h of treatment and/or incubation, which corresponds to days 5 or
7 post-plating, respectively. At the end of these periods the cells
were scrape-detached from the culture dishes,
pipette-disaggregated, collected and divided in aliquots. The total
cell number and viability were determined by trypan blue exclusion
and each sample was double-counted in a manner blinded to the
treatment method. The remaining aliquots were used for
spectrophotometric quantification of protein and DNA contents.
Protein content was determined using Bradford’s technique (43), using bovine serum albumin as a
standard. For DNA quantification, the Burton’s method (44) was applied, using 2-deoxy-D-ribose
(Sigma, Steinheim, Germany) as standard.
Immunocytochemical analysis of PCNA
Proliferating cell nuclear antigen (PCNA), the
auxiliary component of DNA polymerase δ, is a 36-kDa nuclear
protein synthesized in the late G1 and S phases of the cell cycle,
and constitutes a useful proliferation marker (45,46).
Cell samples were seeded on coverslips placed at the bottom of
60-mm Petri dishes (n=2 coverslips per dish; 8 dishes per
experimental replicate), incubated for three days and exposed
intermittently for an additional 42 h to a 50-Hz MF at 10 or 100
μT. In the experiments at 10 μT, additional samples were seeded
(n=8 dishes per experimental replicate) for cell counting by trypan
blue exclusion. PCNA expression was analyzed through indirect
immunofluorescence. For this purpose, at the end of the exposure
and/or incubation interval, cells on coverslips were incubated with
the monoclonal antibody anti-PCNA (FL-261; Santa Cruz
Biotechnology, Quimigen S.L., Madrid, Spain) and with an Alexa
Fluor Green secondary antibody (Molecular Probes, Invitrogen, Prat
de Llobregat, Barcelona, Spain). Hoechst 33342 was added to the
mounting medium as a counterstain for nuclei. Background controls
without the primary antibody were also analyzed. The samples were
evaluated by a photomicroscope (Nikon Eclipse TE300; Melville, NY,
USA) and computer-assisted image analysis (AnalySIS: Soft Imaging
Systems GmbH, Munich, Germany). In each of a total of 8
experimental replicates, 4 dishes were studied per experimental
condition, MF or sham exposure. Twenty random microscopic fields
per coverslip were evaluated, and ~5,000 cells were studied per
replicate and experimental group. The percentage of PCNA-positive
cells was calculated against the total number of cells.
Statistical analysis
All experimental and analytical procedures were
conducted in a blinded manner to the treatment method. The data
were normalized over the respective control samples, and the values
were presented as the means ± SEM of at least three independent
experimental replicates. The data corresponding to treated groups
and their respective controls were compared by the two-sample
Student’s t-test. The multifactorial one-way analysis of variance,
ANOVA, was used to assess differences between multiple sets of
data. The limit of statistical significance was set at
P<0.05.
Results
First experimental series: cell growth
and viability after MF exposure
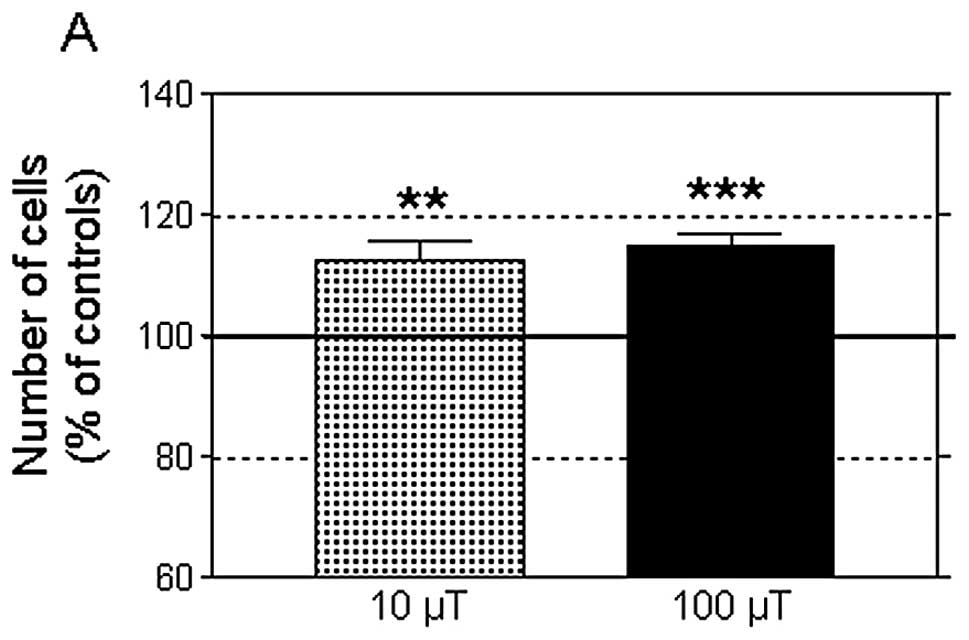
A 42-h intermittent exposure to an MF at 10 or 100
μT significantly increased the average number of NB69 cells when
compared to the corresponding controls (12.5 and 14.8%,
respectively, Fig. 1A). The MF
treatments did not influence cell viability, which was ~85%, both
in the MF-treated and in the sham-exposed samples. Nor were
significant changes observed in the percentages of necrotic cells
after exposure to 10 or 100 μT MF (2 and 4.5% over the controls,
respectively) as analyzed by trypan blue dye exclusion staining.
The total protein content, quantified through spectrophotometric
analysis, revealed no differences between samples exposed to MF at
10 or 100 μT and their controls (data not shown). However, the DNA
content was significantly increased by 8.1 and 17.4% over the
controls at 10 and 100 μT MF, respectively (Fig. 1B). As a whole, these results
confirmed previously reported findings that intermittent exposure
to 50-Hz MF stimulates cell growth in human neuroblastoma cultures
(34,47) and showed that an MF at 10 and 100 μT
induces equivalent proliferative responses.
Second experimental series: cell growth
response to combined treatment with MF and retinoic acid
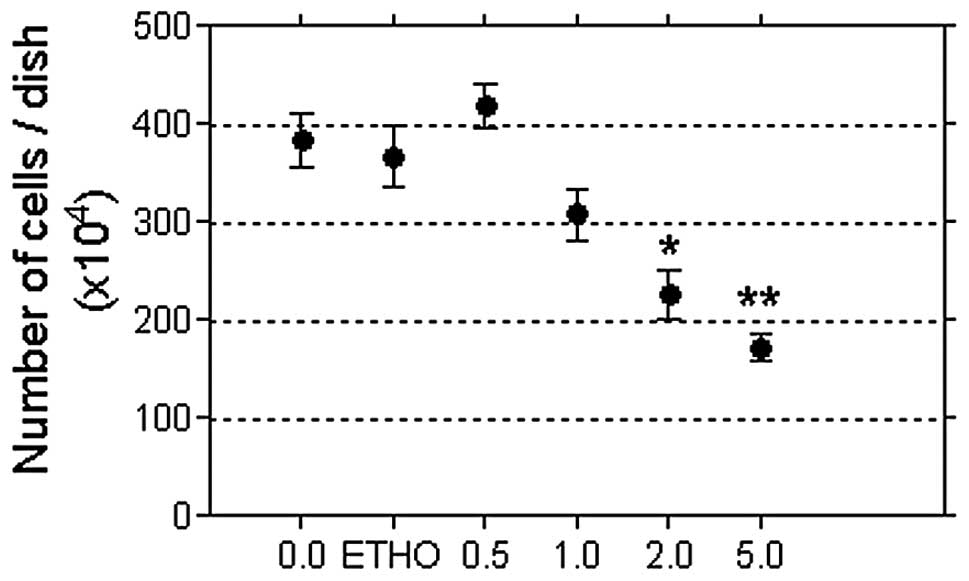
Fig. 2 shows the
results of a pilot study testing the NB69 cell growth response to 5
days of treatment with RA at concentrations of 0.5 to 5.0 μM, which
fall within the physiological range in mammals. RA induced a
linear, dose-dependent decrease in cell number (Pearson’s
correlation coefficient, r=−0.8915; P<0.05). Based on this
result, the intermediate dose of 2 μM RA was selected to
investigate the influence of MF on the cell growth response to
RA.
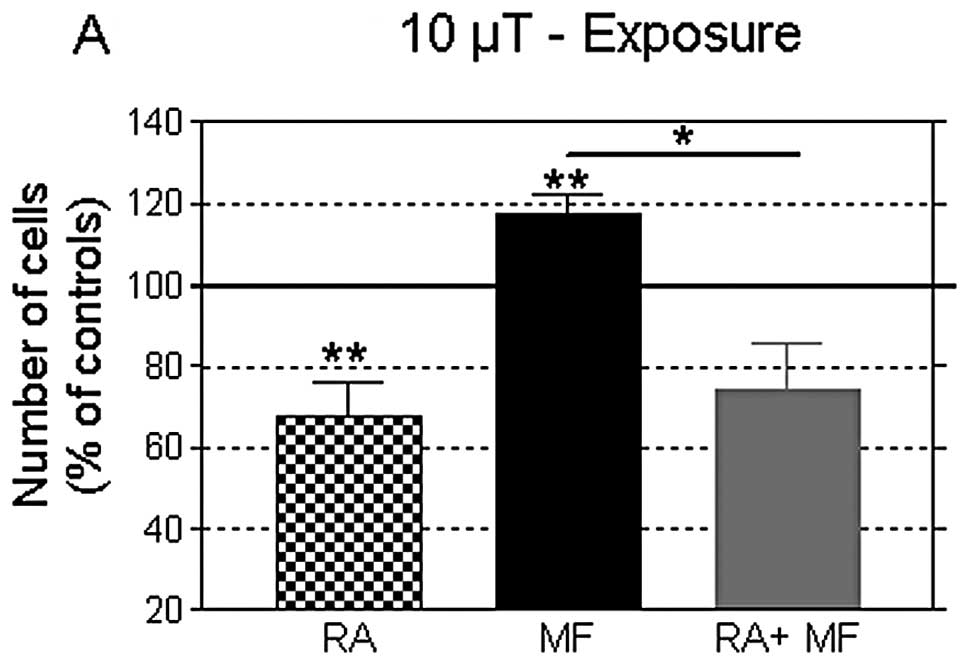
As shown in Fig. 3,
at the end of day 5 post-plating, a 42-h exposure to MF at 10 or
100 μT induced significant increases in cell growth (17.7 and 11.0%
over controls; P<0.01 and P<0.001, respectively), confirming
the results obtained in the first experimental series. In turn,
this growth-promoting effect elicited by the MFs was fully blocked
by the presence of 2 μM RA in the culture medium. Furthermore, the
cell number at the end of the combined treatment was significantly
lower than that in the controls, and equivalent to that in samples
treated with RA only. The cell viability was not significantly
altered in any of the conditions tested (data not shown). These
results indicated that RA caused NB69 cells to be irresponsive or
insensitive to the MF-induced cytoproliferative effects.
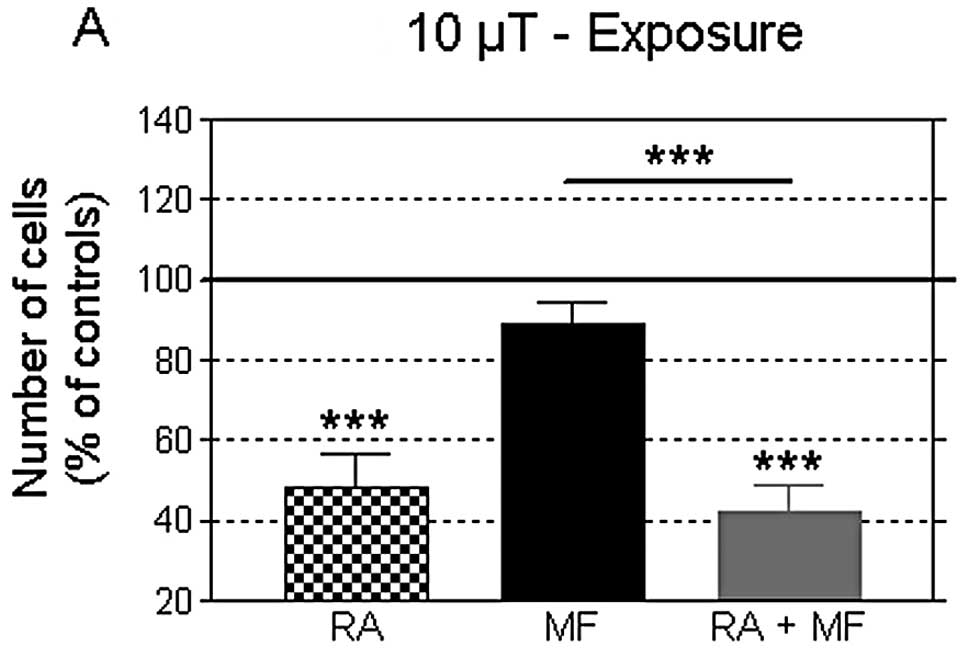
The data in Fig. 4
show that at day 7 post-plating, after 90 h of exposure, no changes
were induced in the cell growth by any of the two flux densities
tested. Thus, the proliferative response at 42 h of MF exposure was
not observed when the treatment was prolonged for an additional
48-h period. In turn, on day 7 post-plating, the RA-induced
decrease in cell number was more pronounced than that observed on
day 5, both in the presence or in the absence of the MF. The cell
viability was not significantly altered in any of the conditions
tested (data not shown).
Concerning protein and DNA levels, the
administration of 2 μM RA alone significantly reduced their levels,
both at day 5 and 7 of treatment (Tables I and II), coincidentally with the observed
reduction in cell number. Again, this response to RA was not
affected by the simultaneous MF exposure for 42 or 90 h. Under most
experimental conditions tested in the second series of experiments,
treatment with MF alone did not change the total protein and DNA
content. Exceptions to this was the DNA content at 42 h of exposure
to MF at 10 μT (Table II) and the
amount of protein at 90 h of treatment with 100 μT (Table I), which were modestly but
significantly increased. This could indicate that an increase in
the cell number may also occur at long-term exposures, after 90 h
of MF stimulation starting at day 3 post-plating. However, the cell
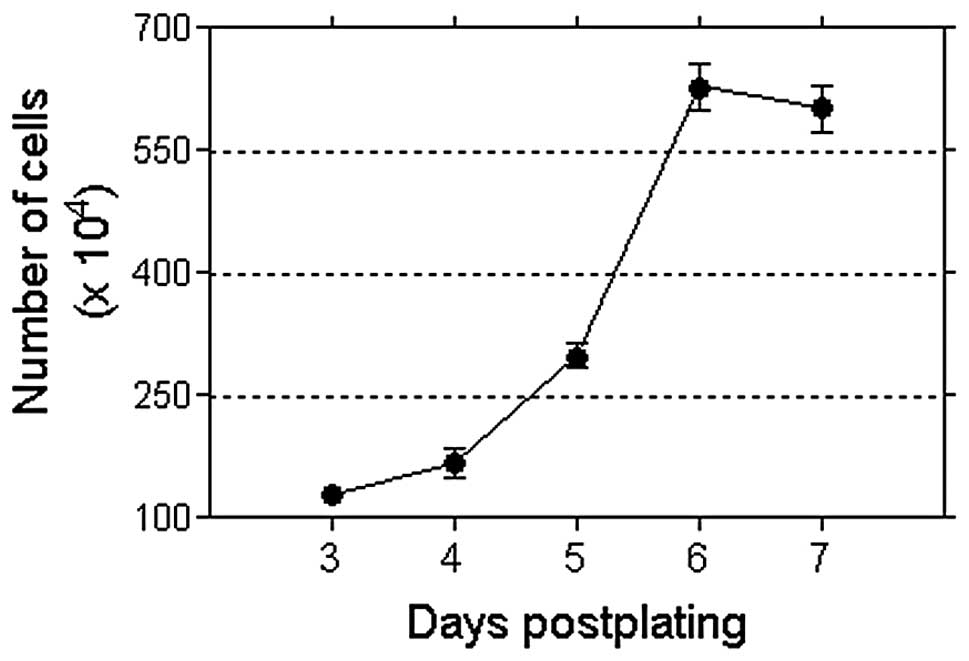
confluence in the cultures, almost reaching saturation at that time
(Fig. 5) could have prevented the
growth response to the MF. Experiments at a lower cell density
would be needed to test this hypothesis. It should be noted that in
this experimental series, the cell growth response induced by a
42-h exposure to MF at 100 μT was not found to be associated with
incremental changes in DNA content (Table II), which is in contrast with the
results obtained in the first series using the same flux density
and exposure interval. This apparent discrepancy may be due to the
relatively limited sensitivity of the spectrophotometric technique,
which may fail to reveal differences in the DNA content when the
differences in the cell number are in the rank of 10% or below.
 | Table IChanges in total protein content. |
Table I
Changes in total protein content.
| Day 5
post-plating | Day 7
post-plating |
|---|
|
|
|
|---|
| RA | MF (42 h) | RA+MF | RA | MF (90 h) | RA+MF |
|---|
| 10 μT | 85.34±3.21b | 110.10±6.56 | 83.56±3.56b | 73.14±4.46c | 95.75±3.09 | 72.53±1.55c |
| 100 μT | 76.81±5.33b | 105.30±3.39 | 82.47±2.85c | 63.66±4.50c | 105.60±2.39a | 68.44±7.52b |
| Table IIChanges in DNA content. |
Table II
Changes in DNA content.
| Day 5
post-plating | Day 7
post-plating |
|---|
|
|
|
|---|
| RA | MF (42 h) | RA+MF | RA | MF (90 h) | RA+MF |
|---|
| 10 μT | 71.35±2.64c | 124.8±9.83a | 74.90±9.21a | 60.45±7.61b | 98.67±3.68 | 56.20±3.62c |
| 100 μT | 69.67±3.33c | 98.33±3.74 | 71.06±2.28c | 51.33±3.20c | 96.43±3.98 | 48.35±2.37c |
Immunocytochemical analysis of PCNA
Assessment of the proliferative response to a 42-h
exposure to MF at 10 or 100 μT was carried out through analysis of
the levels of the proliferation marker PCNA at the end of the
treatment. PCNA expression is known to be correlated with the
proliferative activity in neuroblastomas (48). In addition, PCNA is necessary for
nucleotide-excision repair of DNA (49). The analysis of PCNA expression
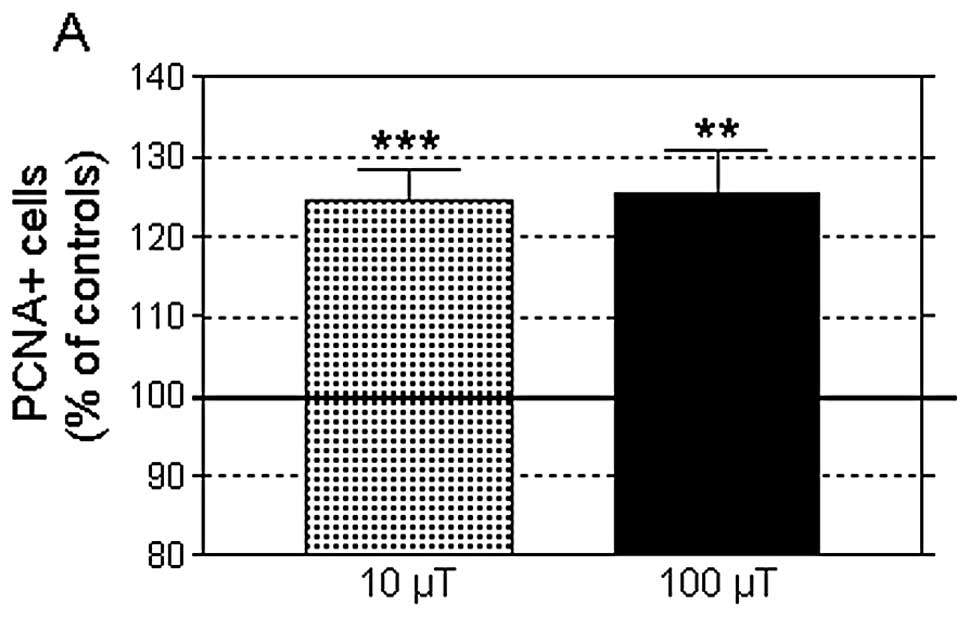
revealed that both MF densities, 10 or 100 μT, significantly
increased the proportion of PCNA-positive cells (24.5±4.07 and
25.6±5.27% over controls, respectively; Fig. 6), which confirmed the observed
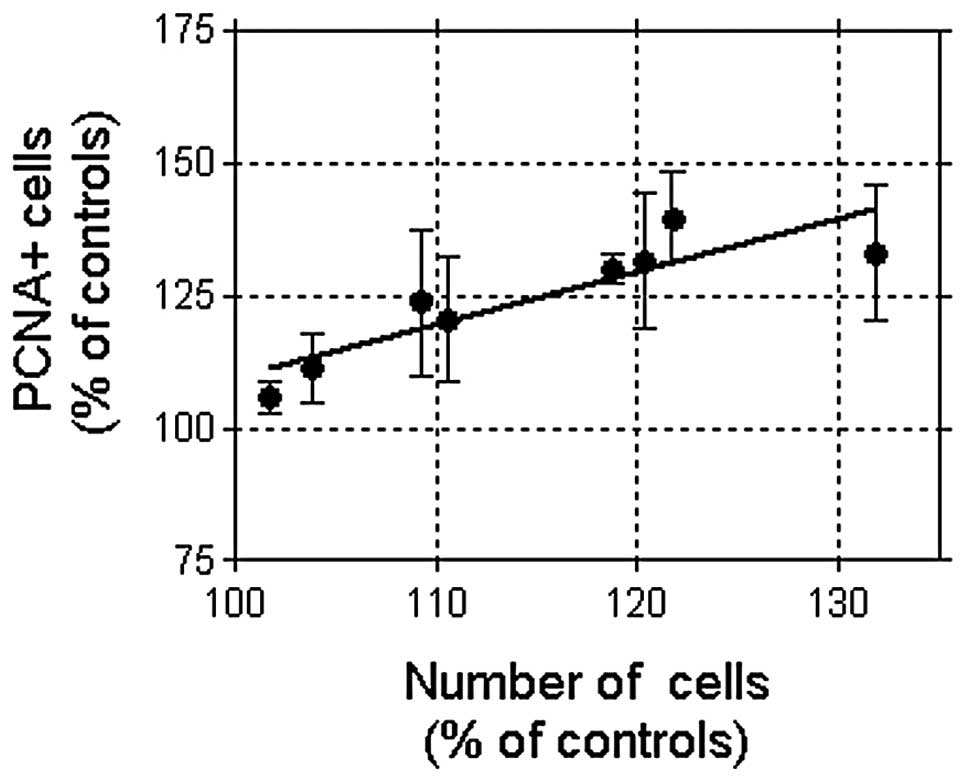
effects on cell growth. At MF at 10 μT the response was associated
with an increased cell number (14.80±3.59% over the controls;
P<0.01); both parameters, PCNA-positive cells and total cell
number, being linearly correlated (8 experimental replicates,
Pearson’s r=0.8832; P<0.01, Fig.
7). However, the number of replicates at 100 μT (n=3) was
insufficient to provide a statistical correlation.
Discussion
The present data showed that 42 h of intermittent
exposure to a 50-Hz sinus wave MF induced similar cytoproliferative
responses in the NB69 cell line at flux densities of 10 and 100 μT.
On the other hand, 2 μM retinoic acid significantly reduced the
cell number, as well as the protein and DNA contents at the
different times assayed. This antiproliferative effect of RA was
not significantly affected by simultaneous exposure to MF at 10 or
100 μT, indicating that RA inhibits the growth-promoting effects
induced by an MF when administered alone.
Consistent evidence exists that power frequency MF
can influence proliferation in different cell types when
administered at densities of 1 mT or above. For instance, Delle
Monache et al(50) reported
that exposure to a sine wave 50-Hz MF at 1 mT increases
proliferation in endothelial cells from human umbilical vein in a
time-dependent manner, with statistically significant effects
achieved at exposure periods of 6 h or longer. Vianale et
al(18) also demonstrated that
a 48-h exposure to a 50-Hz MF at 1 mT modulates keratinocyte growth
through inhibition of the nuclear factor κ-light-chain-enhancer of
activated B cell (NF-κB) signaling pathway. Wolf et
al(51) reported that 24–72 h
of exposure to 50-Hz MF at 0.5 to 1 mT, induced a dose-dependent
increase in cell proliferation and in DNA damage (DNA strand
breaks) in normal and cancer cells, including HL-60 leukemia cells,
Rat-1 fibroblasts and WI-38 diploid fibroblasts. Proliferation of
neuroblastoma cell lines was also found to be sensitive to ELF MF.
For instance, 7 days of continuous exposure to a 50-Hz MF at 1 mT
significantly increased, the proliferation rate by 10% in the
neuroblastoma cell line LAN-5 (24). Exposure to a 50-Hz ELF-MF at 1 mT
also induced cytoproliferation and dedifferentiation in SH-SY5Y
neuroblastoma cells, triggering overexpression of proteins related
to high malignant potential, drug resistance, cytoskeleton
re-arrangement and enhanced defense against oxidative stress
(52).
Besides of the above, a large number of studies have
reported a heterogeneous variety of in vitro responses to MF
(53–56) that, when taken together, have often
been considered contradictory. However, at least part of such
apparent contradictions can be attributable to the fact that the
cellular response to ELF MF is dependent on a number of biological
and physical parameters or factors, including the exposure time or
the specific cell type used (57).
For instance, the intermittency of exposure has recently been
revealed as a critical factor in the proliferative response of NB69
cells to power frequency MF (47)
which, in turn, supports the results and hypothesis of other
authors (28,58).
Thus, the biological effects of weak ELF EMF, in
particular those concerning promotion of cell proliferation, remain
a matter of debate (reviewed in ref. 59). Among studies that have focused on
the proliferative effects of power frequency MF, only a few have
assayed magnetic flux density values below the safety levels
recommended by ICNIRP for occupational protection against
field-induced health effects (500 μT at f=50 Hz). In the present
study, a proliferative effect was observed at the end of 42 h of
intermittent exposure to weak magnetic flux densities of 10 and 100
μT, below the ICNIRP’s reference levels and much lower than those
assayed in most of the studies described above. However, such a
proliferative effect was not observable when the intermittent
exposure was extended for an additional time duration of 48 h
(total exposure time 90 h). At that time no differences were
detected with respect to sham-exposed samples, except for a small,
although statistically significant increase in total protein
levels. In this regard, it must be taken into account that a
spontaneous reduction in the cell number occurred in control
samples at day 7 after seeding. This decline in cell proliferation
may be attributable to the cell density-dependent regulation of
growth, as well as to depletion of nutrients in the medium
(60). Thus, in the event that the
cell sensitivity to MF extended for periods longer than the 42 h of
exposure, as indicated by the above mentioned increase in protein
content, it would not be detectable in terms of changes in cell
growth because of the described spontaneous decline in cell
proliferation.
In order to identify the cellular events involved in
the proliferative effects of an ELF MF, a number of studies have
investigated the in vitro response to the fields when
administered in combination with different chemical agents
(61,62). Tonini et al(63) reported inhibition of proliferation
in the neuroblastoma × glioma hybrid cell line NG108-15 when
treated with the differentiating agent Bt2cAMP. This chemical
inhibition of cell proliferation was counterbalanced by exposure to
a 50-Hz MF at 120 or 240 μT. Retinoic acid is known to be one of
the most potent inducers of differentiation in human neuroblastoma
(64–66); however, the molecular mechanisms and
signaling pathways that are responsible for RA-mediated
neuroblastoma cell differentiation remain unclear. Retinoids are
signaling molecules that are involved in cell proliferation,
differentiation and apoptosis via both, non-receptor- and nuclear
receptor-mediated pathways, thereby altering gene expression.
Treatment of human neuroblastoma cell lines with retinoic acid
causes a significant decrease in MYCN RNA expression and arrest of
cell proliferation (65,67). The non-genomic effects of RA on
neuroblastoma SH-SY5Y cells are mediated by the classical nuclear
receptor, the retinoic acid receptor (RAR), which promotes
activation of the PI3K and MAPK signaling pathways that intervene
in RA-induced differentiation (68). The present results reveal that,
administered at the physiological concentration of 2 μM, RA induces
a significant decrease in cell number, associated with decreased
protein and DNA contents, indicating that RA exerts
antiproliferative effects in NB69 cells. These results support
reports that RA induces specific phenotype expression and growth
inhibition in a number of cell types, including NB69 (69–72).
Our data also provide further support and additional rationale to
the medical application of RA, alone or in combination with other
chemicals, aimed to induce cell differentiation, apoptosis and
growth arrest in tumors (37,67).
With regard to the cellular response to combined
treatments with MF and RA, recent experimental data (73) showed that sinusoidal MF exposure (50
Hz, 1 mT), apart from enhancing the antiproliferative response
induced by RA (5 μM), has a synergistic effect with RA-induced
neural differentiation in the human neuroblastoma cell line
BE(2)C. Moreover, Pirozzoli et
al(24) found that RA induced
an antiproliferative effect in the human neuroblastoma line LAN-5,
which was significantly inhibited (by ~22%) by a 72-h exposure to a
50-Hz MF at 1 mT. In contrast to this, herein we report that a 42-h
exposure to a 50-Hz MF at 10 or 100 μT did not inhibit or revert
the antiproliferative effect nor the decrease in protein and DNA
contents induced in NB69 cells by 2 μM RA. Rather, our data
revealed that the presence of RA prevented or antagonized the
proliferative effect induced by the MF. The differences between
these 3 studies concerning the cellular response to RA in
combination with an MF, could be due to the differences in the MF
parameters and exposure protocols or in the RA concentrations
tested. In addition, differences in the cellular genetics,
including the varied presence of multiple copies of the MYCN
oncogene and/or their transcriptional activation, which have been
shown to be implicated in ELF MF responses (74) may be among the causes for the
dissimilarities between these studies.
Moreover, we previously showed that the
proliferative response of NB69 cells to a 50-Hz MF at 100 μT is
mediated by the MAPK-ERK signaling pathway (47) that can elicit heterogeneous
responses through cell type-specific regulatory mechanisms
(75). Since the effects of RA on
cell differentiation, proliferation and apoptosis are also mediated
by MAPK-ERK (76), it is possible
that RA can prevent the proliferative response to an MF by acting
on the common ERK growth signaling pathway. The ERK pathway, also
called the MEK-ERK cascade, is one of the main cytoplasmic
signaling transduction systems that regulate processes of
proliferation and survival in eukaryotic cells. Upregulation of
ERK1/2 has been proposed to be implicated in tumor progression and
metastasis in different cancer cell types (77,78).
In the nervous system ERK1/2 has been connected to neuronal
responses to stimuli, both functional (modulating neuronal
survival, differentiation and plasticity) and pathological (such as
Alzheimer’s or Parkinson’s disease) (79–81).
Although the specific mechanism underlying the MF proliferative
effect mediated by the MEK-ERK1/2 cascade is yet to be identified,
it has been shown that ERK1/2 can be transiently activated by a
variety of signals, including ELF MF (82) or UHF radio waves (83), as well as ionizing radiation
(84).
Concerning DNA damage, several studies have reported
increased DNA aberrations under specific MF exposure conditions
(85,86). By contrast, others have not found
such effects (87,88). Since it has been comfirmed that low
frequency magnetic or electromagnetic fields do not transmit energy
high enough to break chemical bonds, there is general agreement
that these fields are unable of directly damaging DNA (89). Nevertheless, several hypotheses have
been proposed on how electromagnetic fields might indirectly affect
the structure of DNA (90–92). By reproducing experimental
conditions described by Ivancsits et al(28) and using Comet assay analysis, Focke
and coworkers (58) found a
significant increase in DNA fragmentation in primary cultures of
human fibroblasts exposed to a 50-Hz MF at 1 mT. The slight,
although significant effect on DNA integrity was dependent on the
intermittence of the exposure as well as on the cell line used; the
latter indicating that the effect was mediated by biological
factors of a genetic or physiological nature. These results also
suggest that the potential genotoxic impact of the field may be due
to slightly increased apoptosis and to disturbances in DNA
transactions, both associated with the S-phase of the cell cycle.
Although we do not know whether our exposure parameters induced DNA
damage through a similar phenomenon, our data revealed that a 42-h
exposure to an MF at 10 or 100 μT increased PCNA expression, which
may be related to disturbances in the S-phase. Moreover, since a
response was observed to be associated with an increased cell
number, the present data indicate that the cytoproliferative effect
of an MF may be mediated by stimulation of cell cycle progression
in S-phase. This indication receives support from recently
published data on increased BrdU incorporation in the DNA of NB69
cells intermittently exposed to a 50-Hz MF at 100 μT (34,47).
In conclusion, the herein reported results confirm
previous indications that 42 h of intermittent exposure to a weak
50-Hz MF enhances proliferation in human neuroblastoma NB69 cells.
The effects induced by two flux densities, 10 and 100 μT, were
equivalent, non-significantly different from each other. The
effects were manifested as a significant increase in the cell
number. This increase was associated and linearly correlated to
increased expression of PCNA, which was potentially related to the
observed MF-induced disturbances in the S-phase of the cell cycle.
If in fact these MF effects reflect an action exerted primarily on
cell cycle-control molecules, this would reinforce prior data that
the sensitive fraction of the cell population is the one undergoing
cell division during the exposure intervals (34). The response to MF was fully
inhibited or counteracted by 2 μM retinoic acid, which when
administered alone or in combination with the MF, significantly
reduced the cell number as well as the protein and DNA contents
with respect to the corresponding untreated controls. This blocking
of the MF effects may be exerted through RA-induced changes in the
MAPK-ERK signaling pathway, which is known to intervene in the
cellular response to both agents, RA and ELF MF (47,76).
Collectively, the present data are of potential relevance to
identify the mechanisms by which human cells are sensitive to ELF
MF at flux densities below the reference levels recommended by
ICNIRP (9,10) for protection against the deleterious
effects of occupational or residential MF exposure.
Acknowledgements
This study was supported by grants from the European
Union, under the program ‘Quality of Life and Management of Living
Resources’, Key Action 4 ‘Environment and Health’, REFLEX Project:
QLK4-CT-1999-01574 and Ministry of Science and Technology, Spain,
under the program ‘Special Actions of the National Plan for
Scientific Research, Development and Technological Innovation’:
AE00-0376.
References
|
1
|
Kheifets LI, Afifi AA, Buffler PA and
Zhang ZW: Occupational electric and magnetic field exposure and
brain cancer: a meta-analysis. J Occup Environ Med. 37:1327–1341.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kliukiene J, Tynes T and Andersen A:
Residential and occupational exposures to 50-Hz magnetic fields and
breast cancer in women: a population-based study. Am J Epidemiol.
159:852–861. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Davanipour Z and Sobel E: Long-term
exposure to magnetic fields and the risks of Alzheimer’s disease
and breast cancer: further biological research. Pathophysiology.
16:149–156. 2009.
|
|
4
|
Hakansson N, Gustavsson P, Johansen C and
Floderus B: Neurodegenerative diseases in welders and other workers
exposed to high levels of magnetic fields. Epidemiology.
14:420–426. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huss A, Spoerri A, Egger M and Röösli M:
Residence near power lines and mortality from neurodegenerative
diseases: longitudinal study of the Swiss population. Am J
Epidemiol. 169:167–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Feychting M and Forssen U: Electromagnetic
fields and female breast cancer. Cancer Causes Control. 17:553–558.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kheifets L, Bowman JD, Checkoway H,
Feychting M, Harrington JM, Kavet R, Marsh G, Mezei G, Renew DC and
van Wijngaarden E: Future needs of occupational epidemiology of
extremely low frequency electric and magnetic fields: review and
recommendations. Occup Environ Med. 66:72–80. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
International Agency for Research of
Cancer (IARC). IARC monograph on the evaluation of carcinogenic
risks to humans. 80:Non-ionizing radiation, Part 1: Static and
extremely low-frequency (ELF) electric and magnetic fields. IARC
Press; Lyon, France: 2002, Retrieved from: http://monographs.iarc.fr/ENG/Monographs/vol80/mono80.pdf.
Last accessed 1 August 2012
|
|
9
|
International Commission on Non-Ionizing
Radiation Protection (ICNIRP). Guidelines for limiting exposure to
time varying electric, magnetic and electromagnetic fields. Health
Phys. 74:494–522. 1998.PubMed/NCBI
|
|
10
|
International Commission on Non-Ionizing
Radiation Protection (ICNIRP). Guidelines for limiting exposure to
time varying electric and magnetic fields (1 Hz to 100 kHz). Health
Phys. 99:818–836. 2010.PubMed/NCBI
|
|
11
|
Fedrowitz M and Loscher W: Exposure of
Fischer 344 rats to a weak power frequency magnetic field
facilitates mammary tumorigenesis in the DMBA model of breast
cancer. Carcinogenesis. 29:186–193. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiménez-García MN, Arellanes-Robledo J,
Aparicio-Bautista DI, Rodríguez-Segura MA, Villa-Trevino S and
Godina-Nava JJ: Anti-proliferative effect of extremely low
frequency electromagnetic field on preneoplastic lesions formation
in the rat liver. BMC Cancer. 10:159–170. 2010.PubMed/NCBI
|
|
13
|
Wen J, Jiang S and Chen B: The effect of
100 Hz magnetic field combined with X-ray on hepatoma-implanted
mice. Bioelectromagnetics. 32:322–324. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Juutilainen J: Do electromagnetic fields
enhance the effects of environmental carcinogens? Radiat Prot
Dosimetry. 132:228–231. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Santini MT, Rainaldi G and Indovina PL:
Cellular effects of extremely low frequency (ELF) electromagnetic
fields. Int J Radiat Biol. 85:294–313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Zeng Q, Weng Y, Lu D, Jiang H and Xu
Z: Effects of ELF magnetic fields on protein expression profile of
human breast cancer cells MCF7. Sci China C Life Sci. 48:506–514.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lupke M, Frahm J, Lantow M, Maercker C,
Remondini D, Bersani F and Simko M: Gene expression analysis of
ELF-MF exposed human monocytes indicating the involvement of the
alternative activation pathway. Biochim Biophys Acta. 1763:402–412.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vianale G, Reale M, Amerio P, Stefanachi
M, Di Luzio S and Muraro R: Extremely low frequency electromagnetic
field enhances human keratinocyte cell growth and decreases
proinflammatory chemokine production. Br J Dermatol. 158:1189–1196.
2008. View Article : Google Scholar
|
|
19
|
Simko M, Kriehuber R, Weiss DG and Luben
RA: Effects of 50 Hz EMF exposure on micronucleus formation and
apoptosis in transformed and non-transformed human cell lines.
Bioelectromagnetics. 19:85–91. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nikolova T, Czyz J, Rolletschek A,
Blyszczuk P, Fuchs J, Jovtchev G, Schuderer J, Kuster N and Wobus
AM: Electromagnetic fields affect transcript levels of
apoptosis-related genes in embryonic stem cell-derived neural
progenitor cells. FASEB J. 19:1686–1688. 2005.PubMed/NCBI
|
|
21
|
Manikonda PK, Rajendra P, Devendranath D,
Gunasekaran B, Channakeshava, Aradhya RS, Sashidhar RB and
Subramanyam C: Influence of extremely low frequency magnetic fields
on Ca2+ signaling and NMDA receptor functions in rat
hippocampus. Neurosci Lett. 413:145–149. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gaetani R, Ledda M, Barile L, Chimenti I,
De Carlo F, Forte E, Ionta V, Giuliani L, D’Emilia E, Frati G,
Miraldi F, Pozzi D, Messina E, Grimaldi S, Giacomello A and Lisi A:
Differentiation of human adult cardiac stem cells exposed to
extremely low-frequency electromagnetic fields. Cardiovasc Res.
82:411–420. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Di Loreto S, Falone S, Caracciolo V,
Sebastiani P, D’Alessandro A, Mirabilio A, Zimmitti V and
Amicarelli F: Fifty hertz extremely low-frequency magnetic field
exposure elicits redox and trophic response in rat-cortical
neurons. J Cell Physiol. 219:334–343. 2009.PubMed/NCBI
|
|
24
|
Pirozzoli MC, Marino C, Lovisolo GA,
Laconi C, Mosiello L and Negroni A: Effects of 50 Hz
electromagnetic field exposure on apoptosis and differentiation in
a neuroblastoma cell line. Bioelectromagnetics. 24:510–516. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Falone S, Grossi MR, Cinque B, D’Angelo B,
Tettamanti E, Cimini A, Di Ilio C and Amicarelli F: Fifty hertz
extremely low-frequency electromagnetic field causes changes in
redox and differentiative status in neuroblastoma cells. Int J
Biochem Cell Biol. 39:2093–2106. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Eleuteri AM, Amici M, Bonfili L, Cecarini
V, Cuccioloni M, Grimaldi S, Giuliani L, Angeletti M and Fioretti
E: 50 Hz extremely low frequency electromagnetic fields enhance
protein carbonyl groups content in cancer cells: effects on
proteasomal systems. J Biomed Biotechnol. 2009:8342392009.
View Article : Google Scholar
|
|
27
|
Simko M, Kriehuber R and Lange S:
Micronucleus formation in human amnion cells after exposure to 50
Hz MF applied horizontally and vertically. Mutat Res. 418:101–111.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ivancsits S, Diem E, Pilger A, Rudiger HW
and Jahn O: Induction of DNA strand breaks by intermittent exposure
to extremely-low-frequency electromagnetic fields in human diploid
fibroblasts. Mutat Res. 519:1–13. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ivancsits S, Diem E, Jahn O and Rudiger
HW: Intermittent extremely low frequency electromagnetic fields
cause DNA damage in a dose-dependent way. Int Arch Occup Environ
Health. 76:431–436. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ivancsits S, Diem E, Jahn O and Rudiger
HW: Age-related effects on induction of DNA strand breaks by
intermittent exposure to electromagnetic fields. Mech Ageing Dev.
124:847–850. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fatigoni C, Dominici L, Moretti M,
Villarini M and Monarca S: Genotoxic effects of extremely low
frequency (ELF) magnetic fields (MF) evaluated by the
Tradescantia-micronucleus assay. Environ Toxicol. 20:585–591. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Simko M and Mattsson MO: Extremely low
frequency electromagnetic fields as effectors of cellular responses
in vitro: possible immune cell activation. J Cell Biochem.
93:83–92. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mannerling AC, Simkó M, Mild KH and
Mattsson MO: Effects of 50-Hz magnetic field exposure on superoxide
radical anion formation and HSP70 induction in human K562 cells.
Radiat Environ Biophys. 49:731–741. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Trillo MA, Martínez MA, Cid MA, Leal J and
Úbeda A: Influence of a 50 Hz magnetic field and of
all-trans-retinol on the proliferation of human cancer cell lines.
Int J Oncol. 40:1405–1413. 2012.PubMed/NCBI
|
|
35
|
Tulachan SS, Doi R, Kawaguchi Y, Tsuji S,
Nakajima S, Masui T, Koizumi M, Toyoda E, Mori T, Ito D, Kami K,
Fujimoto K and Imamura M: All-trans retinoic acid induces
differentiation of ducts and endocrine cells by
mesenchymal/epithelial interactions in embryonic pancreas.
Diabetes. 52:76–84. 2003. View Article : Google Scholar
|
|
36
|
Schenk T, Chen WC, Göllner S, Howell L,
Jin L, Hebestreit K, Klein HU, Popescu AC, Burnett A, Mills K,
Casero RA Jr, Marton L, Woster P, Minden MD, Dugas M, Wang JC, Dick
JE, Müller-Tidow C, Petrie K and Zelent A: Inhibition of the LSD1
(KDM1A) demethylase reactivates the all-trans-retinoic acid
differentiation pathway in acute myeloid leukemia. Nat Med.
18:605–611. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang QJ, Zhou LY, Mu YQ, Zhou QX, Luo JY,
Cheng L, Deng ZL, He TC, Haydon RC and He BC: All-trans retinoic
acid inhibits tumor growth of human osteosarcoma by activating Smad
signaling-induced osteogenic differentiation. Int J Oncol.
41:153–160. 2012.PubMed/NCBI
|
|
38
|
Handler A, Lobo MD, Alonso FJ, Paíno CL
and Mena MA: Functional implications of the
noradrenergic-cholinergic switch induced by retinoic acid in NB69
neuroblastoma cells. J Neurosci Res. 60:311–320. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hölzel M, Huang S, Koster J, Ora I,
Lakeman A, Caron H, Nijkamp W, Xie J, Callens T, Asgharzadeh S,
Seeger RC, Messiaen L, Versteeg R and Bernards R: NF1 is a tumor
suppressor in neuroblastoma that determines retinoic acid response
and disease outcome. Cell. 142:218–229. 2010.PubMed/NCBI
|
|
40
|
Shih YY, Lee H, Nakagawara A, Juan HF,
Jeng YM, Tsay YG, Lin DT, Hsieh FJ, Pan CY, Hsu WM and Liao YF:
Nuclear GRP75 binds retinoic acid receptors to promote neuronal
differentiation of neuroblastoma. PLoS One. 6:e262362011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Di Nallo AM, Strigari L, Giliberti C,
Bedini A, Palomba R and Benassi M: Monitoring of people and workers
exposure to the electric, magnetic and electromagnetic fields in an
Italian National Cancer Institute. J Exp Clin Cancer Res.
27:162008.PubMed/NCBI
|
|
42
|
Blackman CF, Benane SG and House DE:
Evidence for direct effect of magnetic fields on neurite outgrowth.
FASEB J. 7:801–806. 1993.PubMed/NCBI
|
|
43
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of proteins utilizing
the principle of protein dye-binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Burton K: Study of the conditions and
mechanism of the diphenylamine reaction for the colorimetric
estimation of deoxyribonucleic acid. Biochem J. 62:315–323.
1956.PubMed/NCBI
|
|
45
|
Woods AL, Hall PA, Shepherd NA, Hanby AM,
Waseem NH, Lane DP and Levison DA: The assessment of proliferating
cell nuclear antigen (PCNA) immunostaining in primary
gastrointestinal lymphomas and its relationship to histological
grade, S+G2+M phase fraction (flow cytometric analysis) and
prognosis. Histopathology. 19:21–27. 1991.
|
|
46
|
Tan Z, Wortman M, Dillehay KL, Seibel WL,
Evelyn CR, Smith SJ, Malkas LH, Zheng Y, Lu S and Dong Z: Small
molecule targeting of PCNA chromatin association inhibits tumor
cell growth. Mol Pharmacol. 81:811–819. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Martínez MA, Úbeda A, Cid MA and Trillo
MA: The proliferative response of NB69 human neuroblastoma cells to
a 50 Hz magnetic field is mediated by ERK1/2 signaling. Cell
Physiol Biochem. 29:675–686. 2012.PubMed/NCBI
|
|
48
|
Kawasaki H, Mukai K, Yajima S, Tanaka R,
Takayama J, Takasaki Y and Ohira M: Prognostic value of
proliferating cell nuclear antigen (PCNA) immunostaining in
neuroblastoma. Med Pediatr Oncol. 24:300–304. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Stoimenov I and Helleday T: PCNA on the
crossroad of cancer. Biochem Soc Trans. 37:605–613. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Delle Monache S, Alessandro R, Iorio R,
Gualtieri G and Colonna R: Extremely low frequency electromagnetic
fields (ELF-EMFs) induce in vitro angiogenesis process in human
endothelial cells. Bioelectromagnetics. 29:640–648. 2008.PubMed/NCBI
|
|
51
|
Wolf FI, Torsello A, Tedesco B, Fasanella
S, Boninsegna A, D’Ascenzo M, Grassi C, Azzena GB and Cittadini A:
50-Hz extremely low frequency electromagnetic fields enhance cell
proliferation and DNA damage: possible involvement of a redox
mechanism. Biochim Biophys Acta. 1743:120–129. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sulpizio M, Falone S, Amicarelli F,
Marchisio M, Di Giuseppe F, Eleuterio E, Di Ilio C and Angelucci S:
Molecular basis underlying the biological effects elicited by
extremely low-frequency magnetic field (ELF-MF) on neuroblastoma
cells. J Cell Biochem. 112:3797–3806. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yoshizawa H, Tsuchiya T, Mizoe H, Ozeki H,
Kanao S, Yomori H, Sakane C, Hasebe S, Motomura T, Yamakawa T,
Mizuno F, Hirose H and Otaka Y: No effect of extremely
low-frequency magnetic field observed on cell growth or initial
response of cell proliferation in human cancer cell lines.
Bioelectromagnetics. 23:355–368. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Grassi C, D’Ascenzo M, Torsello A,
Martinotti G, Wolf F, Cittadini A and Azzena GB: Effects of 50 Hz
electromagnetic fields on voltage-gated Ca2+ channels
and their role in modulation of neuroendocrine cell proliferation
and death. Cell Calcium. 35:307–315. 2004.PubMed/NCBI
|
|
55
|
Bułdak RJ, Polaniak R, Bułdak L,
Zwirska-Korczala K, Skonieczna M, Monsiol A, Kukla M, Duława-Bułdak
A and Birkner E: Short-term exposure to 50 Hz ELF-EMF alters the
cisplatin-induced oxidative response in AT478 murine squamous cell
carcinoma cells. Bioelectromagnetics. 33:641–651. 2012.PubMed/NCBI
|
|
56
|
Hong MN, Han NK, Lee HC, Ko YK, Chi SG,
Lee YS, Gimm YM, Myung SH and Lee JS: Extremely low frequency
magnetic fields do not elicit oxidative stress in MCF10A cells. J
Radiat Res. 53:79–86. 2012. View Article : Google Scholar
|
|
57
|
Ivancsits S, Pilger A, Diem E, Jahn O and
Rüdiger HW: Cell type-specific genotoxic effects of intermittent
extremely low-frequency electromagnetic fields. Mutat Res.
583:184–188. 2005.PubMed/NCBI
|
|
58
|
Focke F, Schuermann D, Kuster N and Schär
P: DNA fragmentation in human fibroblasts under extremely low
frequency electromagnetic field exposure. Mutat Res. 683:74–83.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Repacholi M: Concern that ‘EMF’ magnetic
fields from power lines cause cancer. Sci Total Environ.
426:454–458. 2012.
|
|
60
|
Ba F, Pang PK and Benishin CG: The
establishment of a reliable cytotoxic system with SK-N-SH
neuroblastoma cell culture. J Neurosci Methods. 123:11–22. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Úbeda A, Trillo MA, House DE and Blackman
CF: A 50 Hz magnetic field blocks melatonin-induced enhancement of
junctional transfer in normal C3H/10T1/2 cells. Carcinogenesis.
16:2945–2949. 1995.PubMed/NCBI
|
|
62
|
Blackman CF, Benane SG and House DE: The
influence of 1.2 microT, 60 Hz magnetic fields on melatonin- and
tamoxifen-induced inhibition of MCF-7 cell growth.
Bioelectromagnetics. 22:122–128. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Tonini R, Baroni MD, Masala E, Micheletti
M, Ferroni A and Mazzanti M: Calcium protects differentiating
neuroblastoma cells during 50 Hz electromagnetic radiation. Biophys
J. 81:2580–2589. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Reynolds CP, Matthay KK, Villablanca JG
and Maurer BJ: Retinoid therapy of high-risk neuroblastoma. Cancer
Lett. 197:185–192. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kanemaru KK, Tuthill MC, Takeuchi KK,
Sidell N and Wada RK: Retinoic acid induced downregulation of MYCN
is not mediated through changes in Sp1/Sp3. Pediatr Blood Cancer.
50:806–811. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Tanaka K, Tamiya-Koizumi K, Hagiwara K,
Ito H, Takagi A, Kojima T, Suzuki M, Iwaki S, Fujii S, Nakamura M,
Banno Y, Kannagi R, Tsurumi T, Kyogashima M and Murate T: Role of
down-regulated neutral ceramidase during all-trans retinoic
acid-induced neuronal differentiation in SH-SY5Y neuroblastoma
cells. J Biochem. 151:611–620. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Cetinkaya C, Hultquist A, Su Y, Wu S,
Bahram F, Påhlman S, Guzhova I and Larsson LG: Combined IFN-gamma
and retinoic acid treatment targets the N-Myc/Max/Mad1 network
resulting in repression of N-Myc target genes in MYCN-amplified
neuroblastoma cells. Mol Cancer Ther. 6:2634–2641. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Masiá S, Alvarez S, de Lera AR and
Barettino D: Rapid, nongenomic actions of retinoic acid on
phosphatidylinositol-3-kinase signaling pathway mediated by the
retinoic acid receptor. Mol Endocrinol. 21:2391–2402.
2007.PubMed/NCBI
|
|
69
|
Wegert J, Bausenwein S, Kneitz S, Roth S,
Graf N, Geissinger E and Gessler M: Retinoic acid pathway activity
in Wilms tumors and characterization of biological responses in
vitro. Mol Cancer. 10:1362011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Jiao RQ, Li G and Chiu JF: Comparative
proteomic analysis of differentiation of mouse F9 embryonic
carcinoma cells induced by retinoic acid. J Cell Biochem.
113:1811–1819. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li X, Li H, Bi J, Chen Y, Jain S and Zhao
Y: Human cord blood-derived multipotent stem cells (CB-SCs) treated
with all-trans-retinoic acid (ATRA) give rise to dopamine neurons.
Biochem Biophys Res Commun. 419:110–116. 2012.
|
|
72
|
Marzinke MA and Clagett-Dame M: The
all-trans retinoic acid (atRA)-regulated gene Calmin (Clmn)
regulates cell cycle exit and neurite outgrowth in murine
neuroblastoma (Neuro2a) cells. Exp Cell Res. 318:85–93. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Marcantonio P, Del Re B, Franceschini A,
Capri M, Lukas S, Bersani F and Giorgi G: Synergic effect of
retinoic acid and extremely low frequency magnetic field exposure
on human neuroblastoma cell line BE(2)C. Bioelectromagnetics.
31:425–433. 2010.PubMed/NCBI
|
|
74
|
Lin H, Head M, Blank M, Han L, Jin M and
Goodman R: Myc-mediated transactivation of HSP70 expression
following exposure to magnetic fields. J Cell Biochem. 69:181–188.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Shaul YD and Seger R: The MEK/ERK cascade:
from signaling specificity to diverse functions. Biochim Biophys
Acta. 1773:1213–1226. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Karsy M, Albert L, Tobias ME, Murali R and
Jhanwar-Uniyal M: All-trans retinoic acid modulates cancer stem
cells of glioblastoma multiforme in an MAPK-dependent manner.
Anticancer Res. 30:4915–4920. 2010.PubMed/NCBI
|
|
77
|
De Melo M, Gerbase MW, Curran J and Pache
JC: Phosphorylated extracellular signal-regulated kinases are
significantly increased in malignant mesothelioma. J Histochem
Cytochem. 54:855–861. 2006.PubMed/NCBI
|
|
78
|
Menakongka A and Suthiphongchai T:
Involvement of PI3K and ERK1/2 pathways in hepatocyte growth
factor-induced cholangiocarcinoma cell invasion. World J
Gastroenterol. 16:713–722. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Webster B, Hansen L, Adame A, Crews L,
Torrance M, Thal L and Masliah E: Astroglial activation of
extracellular-regulated kinase in early stages of Alzheimer
disease. J Neuropathol Exp Neurol. 65:142–151. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Dagda RK, Zhu J, Kulich SM and Chu CT:
Mitochondrially localized ERK2 regulates mitophagy and autophagic
cell stress: implications for Parkinson’s disease. Autophagy.
4:770–782. 2008.PubMed/NCBI
|
|
81
|
Kawamata J and Shimohama S: Stimulating
nicotinic receptors trigger multiple pathways attenuating
cytotoxicity in models of Alzheimer’s and Parkinson’s diseases. J
Alzheimers Dis. 24(Suppl 2): 95–109. 2011.PubMed/NCBI
|
|
82
|
Jin M, Blank M and Goodman R: ERK1/2
phosphorylation, induced by electromagnetic fields, diminishes
during neoplastic transformation. J Cell Biochem. 78:371–379. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Friedman J, Kraus S, Hauptman Y, Schiff Y
and Seger R: Mechanism of short-term ERK activation by
electromagnetic fields at mobile phone frequencies. Biochem J.
405:559–568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Schmidt-Ullrich RK, Contessa JN, Lammering
G, Amorino G and Lin PS: ERBB receptor tyrosine kinases and
cellular radiation responses. Oncogene. 22:5855–5865. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Winker R, Ivancsits S, Pilger A, Adlkofer
F and Rudiger HW: Chromosomal damage in human diploid fibroblasts
by intermittent exposure to extremely low-frequency electromagnetic
fields. Mutat Res. 585:43–49. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wahab MA, Podd JV, Rapley BI and Rowland
RE: Elevated sister chromatid exchange frequencies in dividing
human peripheral blood lymphocytes exposed to 50 Hz magnetic
fields. Bioelectromagnetics. 28:281–288. 2007. View Article : Google Scholar
|
|
87
|
Stronati L, Testa A, Villani P, Marino C,
Lovisolo GA, Conti D, Russo F, Fresegna AM and Cordelli E: Absence
of genotoxicity in human blood cells exposed to 50 Hz magnetic
fields as assessed by comet assay, chromosome aberration,
micronucleus, and sister chromatid exchange analyses.
Bioelectromagnetics. 25:41–48. 2004. View Article : Google Scholar
|
|
88
|
Scarfi MR, Sannino A, Perrotta A, Sarti M,
Mesirca P and Bersani F: Evaluation of genotoxic effects in human
fibroblasts after intermittent exposure to 50 Hz electromagnetic
fields: a confirmatory study. Radiat Res. 164:270–276. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Adair RK: Extremely low frequency
electromagnetic fields do not interact directly with DNA.
Bioelectromagnetics. 19:136–138. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wan C, Fiebig T, Schiemann O, Barton JK
and Zewail AH: Femtosecond direct observation of charge transfer
between bases in DNA. Proc Natl Acad Sci USA. 97:14052–14055. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Porath D, Bezryadin A, De Vries S and
Dekker C: Direct measurement of electrical transport through DNA
molecules. Nature. 403:635–638. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Giese B: Electron transfer through DNA and
peptides. Bioorg Med Chem. 14:6139–6143. 2006. View Article : Google Scholar : PubMed/NCBI
|