Introduction
Hepatocellular carcinoma (HCC) is one of the most
common types cancer in the world, accounting for 85–90% of the
total primary liver cancer burden worldwide (1); it is also the third most frequent
cause of cancer-related mortality (2). In China, HCC was projected to
contribute more than 400,000 new cases and caused 370,000 deaths
during 2008; in the same year, the incidence of HCC in American men
and women was approximately 15,000 and 6,000 respectively
(International Agency for Research on Cancer: http://www-dep.iarc.fr/). Therefore, HCC represents an
international public health concern as one of the most common and
aggressive types of cancer worldwide.
There are multiple etiological factors that are
associated with the development of HCC, the most prevalent being
chronic hepatitis B (HBV) and hepatitis C virus (HCV) infections
(3); other non-viral causes of HCC
include alcohol abuse, long-term exposure to aflatoxin B1 (AFB),
iron overload syndromes, and non-alcoholic steatohepatitis
(NASH).
The majority of HCC cases occur in
fibrotic/cirrhotic livers, and the common mechanism for
hepatocarcinogenesis is chronic inflammation associated with severe
oxidative stress (4). There is a
large body of evidence indicating that oxidative stress plays a
common pathogenetic role in different agent-mediated
hepatocarcinogeneses, including HBV (5,6), HCV
(7,8), alcohol (9), AFB (10), NASH (11).
Oxidative stress results from a disruption of redox
homeostasis due either to an elevation of reactive oxygen species
(ROS) production or a decline of ROS-scavenging capacity (12). ROS are defined as molecules derived
from oxygen with characteristic instability and chemical reactivity
and include both free radicals, such as superoxide
(O2−) and hydroxyl radical (OH-),
and non-radicals, such as hydrogen peroxide
(H2O2) and singlet oxygen
(1O2). Cells are protected against ROS by
scavenger enzymes such as superoxide dismutases (Mn-SOD and
CuZn-SOD), and non-enzymatic antioxidants, predominantly
glutathione (GSH). ROS can react with polyunsaturated fatty acids
derived from membrane phospholipids or from dietary intake,
resulting in the generation of lipid peroxidation (LPO) products
such as trans-4-hydroxy-2-nonenal (4-HNE) and
malondialdehyde (MDA) (13). 4-HNE
can react with DNA bases such as deoxyadenosine and deoxycytidine
to form the exocyclic etheno-DNA adducts including 1,
N6-ethenodeoxyadenosine (ɛ-dA) and 3,
N4-ethenodeoxycytidine (ɛ-dC) (14). These adducts are highly mutagenic;
for example one leads to the missense mutations in the TP53 gene
(which encodes p53) (15,16). Previous studies provided evidence
that ɛ-DNA adducts may act as a driving force towards malignancy in
cancer-prone liver diseases (9,17–19).
However, detection of ɛ-DNA adducts in HCC and adjacent non-tumor
liver tissues has not been reported.
Therefore, the aim of the present study was to: i)
evaluate the status of oxidative stress in HCC and the surrounding
non-tumor liver tissues; ii) determine the occurrence of ɛ-dA in
these tissues; iii) determine if the formation of ɛ-dA was
associated with liver inflammatory activity and fibrosis
development; iv) assess if a correlation exists between the
formation of ɛ-dA and mutant p53 expression in HCC tissue to
elucidate its potential as a contributor to human
hepatocarcinogenesis.
Materials and methods
Patients and liver specimens
We obtained tumor and (or) adjacent non-malignant
liver tissues from 32 HCC patients who underwent hepatectomies at
our hospital between 2010 and 2011. None of the patients had
received chemotherapy or radiotherapy prior to surgery. Informed
consent was obtained from all patients for subsequent use of their
resected liver tissues. Tissue samples were collected immediately
after liver resection. The non-malignant liver tissues were ≥2 cm
in distance from the tumor margin. The paired tumors and adjacent
non-malignant specimens were not always available for HCC. Half of
the tissue was fixed in 4% paraformaldehyde and embedded in
paraffin (30 patients had paired tumors and adjacent non-malignant
tissues, from 2 patients only tumors were obtained). Serial
sections (5 μm) were prepared for histopathological diagnosis and
immunohistochemical staining. The other half of the tissue was
washed extensively with 0.01 M phosphate-buffered saline (PBS; pH
7.4) solution to remove erythrocytes, and was then blotted on
filter paper and stored at −80°C for further testing (28 patients
had paired tumors and adjacent non-malignant tissues, from 2
patients only tumors were obtained and from 2 patients only
non-malignant tissues). Additionally, control liver sections were
obtained from 8 patients with benign space-occupying diseases of
the liver who underwent surgery (5 liver hemangiomas, 1
hepatolithiasis, 1 focal nodular hyperplasia, 1 chronic
granulomatous inflammation of the liver). The diagnoses were
confirmed by histopathological study. Tumor staging was determined
by the Tumor-Node-Metastasis (TNM) Classification (6th edit)of the
International Union Against Cancer. Table I shows the general
clinicopathological features of these 32 patients with HCC. The
evidence of metastasis included vascular invasion, particularly to
portal vein invasion and/or intrahepatic dissemination. Thirty HCC
patients showed markers of hepatitis B virus infection, and 11 HCC
patients had a history of alcohol abuse (mean alcohol consumption
of 204.5±163.5 g/d; range 50–500 g/d). In 2 HCC cases, the
underlying cause of the liver disease remained unknown. No
HCV-related HCC was found in this study. The present study was
performed according to the guidelines of the Ethics Committee of
Tongji Hospital and was in accordance with the ethical standards of
the World Medical Association Declaration of Helsinki.
 | Table IClinicopathological characteristics
of the 32 HCC patients. |
Table I
Clinicopathological characteristics
of the 32 HCC patients.
| Characteristic | Results |
|---|
| Gender |
| Male | 29 |
| Female | 3 |
| Age (year) |
| ≤45 | 18 |
| >45 | 14 |
| Etiology |
| HBV | 19 |
| HBV + alcohol | 11 |
| Unknown | 2 |
| Aflatoxin B1 |
| Normal | 3 |
| High | 29 |
| Tumor diameter
(cm) |
| ≤5 | 5 |
| >5 | 27 |
| No. of tumors |
| Single | 26 |
| Multiple | 6 |
| Pathological
grade |
| Well
differentiated | 1 |
| Well-moderately
differentiated | 3 |
| Moderately
differentiated | 16 |
| Moderately-poorly
differentiated | 5 |
| Poorly
differentiated | 7 |
| Metastasis |
| Yes | 9 |
| No | 23 |
| TNM |
| I | 18 |
| II + II | 14 |
Histopathological examinations
Histopathological analyses were performed by H&E
and Masson staining. Based on histological grade, tumors were
subdivided into group I (well, well-moderately and moderately
differentiated) and group II (moderately-poorly and poorly
differentiated). The non-cancerous liver tissues were scored for
the grade of necroinflammatory activity and stage of fibrosis
according to the criteria of Desmet et al(20). Necroinflammatory activity (A) was
graded according to the intensity of necroinflammatory lesions: 0,
no histological activity; 1, mild activity; 2, moderate activity;
3, severe activity. The stage of fibrosis (F) was scored: 0, no
fibrosis; 1, portal fibrosis without septa; 2, portal fibrosis with
few septa; 3, numerous septa without cirrhosis; 4, cirrhosis. All
the sections were analyzed without knowledge of clinical data.
Analysis of oxidative stress-related
parameters
Frozen tissue samples (100 mg) were mixed with 1.0
ml 0.01 M PBS (pH 7.4), incubated on ice and then homogenized.
After centrifugation for 20 min at 12,000 rpm at 4°C, the
supernatants were transferred to fresh tubes. After the protein
concentrations were determined, GSH, total superoxide dismutase
(SOD) activity, total antioxidant capacity (T-AOC) and MDA were
assayed. GSH, total SOD activity and T-AOC of the liver tissue were
detected with dithiobis-2-nitrobenzoic acid reactivity assay,
xanthine oxidase-hydroxylamine method and ferric reducing ability
of plasma method separately by using commercially available kits
according to the manufacturer’s instructions (Jiancheng Biological
Technical Institute, Nanjing, China). MDA levels were estimated
with the thiobarbituric acid reactivity assay by using a
commercially available kit according to the manufacturer’s
instructions (Beyotime Biotech Inc., Haimen, China).
Immunohistochemical detection of
ɛ-dA
This was performed as previously described (18) with some modifications in our
laboratory. The paraffin-embedded sections were dewaxed using
xylene and hydrated in descending gradations of alcohol solutions,
then washed in 0.01 M PBS (pH 7.4). The sections were treated in 3%
H2O2 in absolute methanol for 10 min to block
endogenous peroxidase activity. To enhance the immunostaining,
sections were placed in citrate buffer (0.01 M, pH 6.0) and
microwaved intermittently for up to 3 min at 800 W, 7 min at 640 W
and 3 min at 480 W for antigen unmasking. Slides were slowly cooled
down to room temperature. Slides were incubated with proteinase K
(10 μg/ml) (Roche, Mannheim, Germany) to remove histone and
non-histone proteins from DNA, increasing antibody accessibility.
After washing with PBS, slides were treated with 20 μg/ml
ribonuclease (RNAse; Solarbio Science & Technology Co., Ltd.,
Beijing, China) (heated for 10 min at 80°C to inactivate DNAse) at
37°C for 1 h to prevent antibody binding on RNA adducts, and then
washed in PBS for 10 min. DNA was denatured by treatment with 4 N
HCl for 5 min at room temperature and subsequently rinsed in PBS
and double distilled water. The pH was neutralized with 50 mM
Tris-base buffer, pH 7.4 for 5 min at room temperature. After
washing with double distilled water and PBS, these sections were
treated with 8% bovine serum albumin (BSA), 2% horse serum, 0.05%
Tween-20 and 0.05% Triton X for 20 min at room temperature to block
nonspecific binding sites. Slides were incubated with the primary
monoclonal antibody EM-A-1 against ɛ-dA (diluted to 1:20, provided
by Dr P. Lorenz and Dr M. Rajewsky, University of Essen, Germany)
at 4°C overnight. After washing the slides with PBS for 10 min,
antibody detection was performed using the Vectastain Elite ABC kit
(Vector Laboratories, Burlingame, CA, USA), according to the
manufacturer’s instructions. The slides were washed again and then
the reaction products were visualized with diaminobenzidine (DAB;
Dako, Glostrup, Denmark) as the chromogen. After stopping the
reaction in H2O for 5 min, sections were counterstained
with haematoxylin (Dingguo Changsheng Biotechnology Co., Ltd,
Beijing, China) and covered with neutral balsam (Shanghai specimen
and model factory, Shanghai, China). All sections were subjected to
the same procedure under standardized conditions. Negative controls
were performed by omitting the primary antibody.
Immunohistochemical staining of p53, PCNA
and α-SMA
Serial sections (5 μm) were prepared from paraffin
blocks. Sections were deparaffinized and hydrated by sequential
immersion in xylene and graded alcohol solutions, and finally
washed in PBS (0.01 M, pH 7.4). The sections were then incubated in
3% H2O2 in absolute methanol for 10 min to
quench endogenous peroxidase. Antigen retrieval was performed by
microwaving the slides in citrate buffer (0.01 M, pH 6.0) for 3 min
at 800 W, for 7 min at 640 W and for 3 min at 480 W. Slides were
slowly cooled down to room temperature. After washing with PBS,
these sections were treated with 8% BSA and 2% horse serum for 20
min at room temperature to block nonspecific binding sites. Slides
were incubated with the primary antibodies overnight at 4°C. A
mouse anti-p53 monoclonal antibody (diluted to 1:100; Boster
Bioengineering Co., Ltd., Wuhan, China) was used to detect mutant
p53 protein-positive cells. We used a mouse anti-PCNA monoclonal
antibody (diluted to 1:400; Boster Bioengineering Co.) as the
marker of cell proliferation. Immunohistochemical staining of
α-smooth muscle actin (α-SMA) with a mouse monoclonal antibody
(diluted to 1:150; Boster Bioengineering Co.) was performed to
detect activated hepatic stellate cells (HSCs). After washing the
slides with PBS for 10 min, antibody detection was performed using
the Vectastain Elite ABC kit (Vector Laboratories), according to
the manufacturer’s instructions. The slides were washed again and
the reaction products were then visualized with DAB (Dako) as the
chromogen. After stopping the reaction in H2O for 5 min,
sections were counterstained with haematoxylin (Dingguo Changsheng
Biotechnology Co.) and covered with neutral balsam. All sections
were subjected to the same procedure under standardized conditions.
Negative controls were performed by omitting the primary
antibody.
Imaging and semi-quantitative analysis of
immunohistochemical staining
The frequency of ɛ-dA, mutant p53 and PCNA
positively stained nuclei was expressed as % of stained cell nuclei
over total number of cells counted, and 1,000 cells were observed
in five or more random fields at a magnification of ×200 to
calculate the percentage of positive cells.
Statistical analysis
Data are expressed as the means ± standard
deviation. Significant differences were calculated using the
Student’s t-test or Mann-Whitney U test. Correlations were
calculated by Spearman’s rank analysis. P<0.05 was considered to
indicate statistically significant differences. All statistical
analyses were performed using SPSS version 13.0 and GraphPad Prism
version 5.0.
Results
Comparison of oxidative stress parameters
in HCC and non-tumor liver tissues
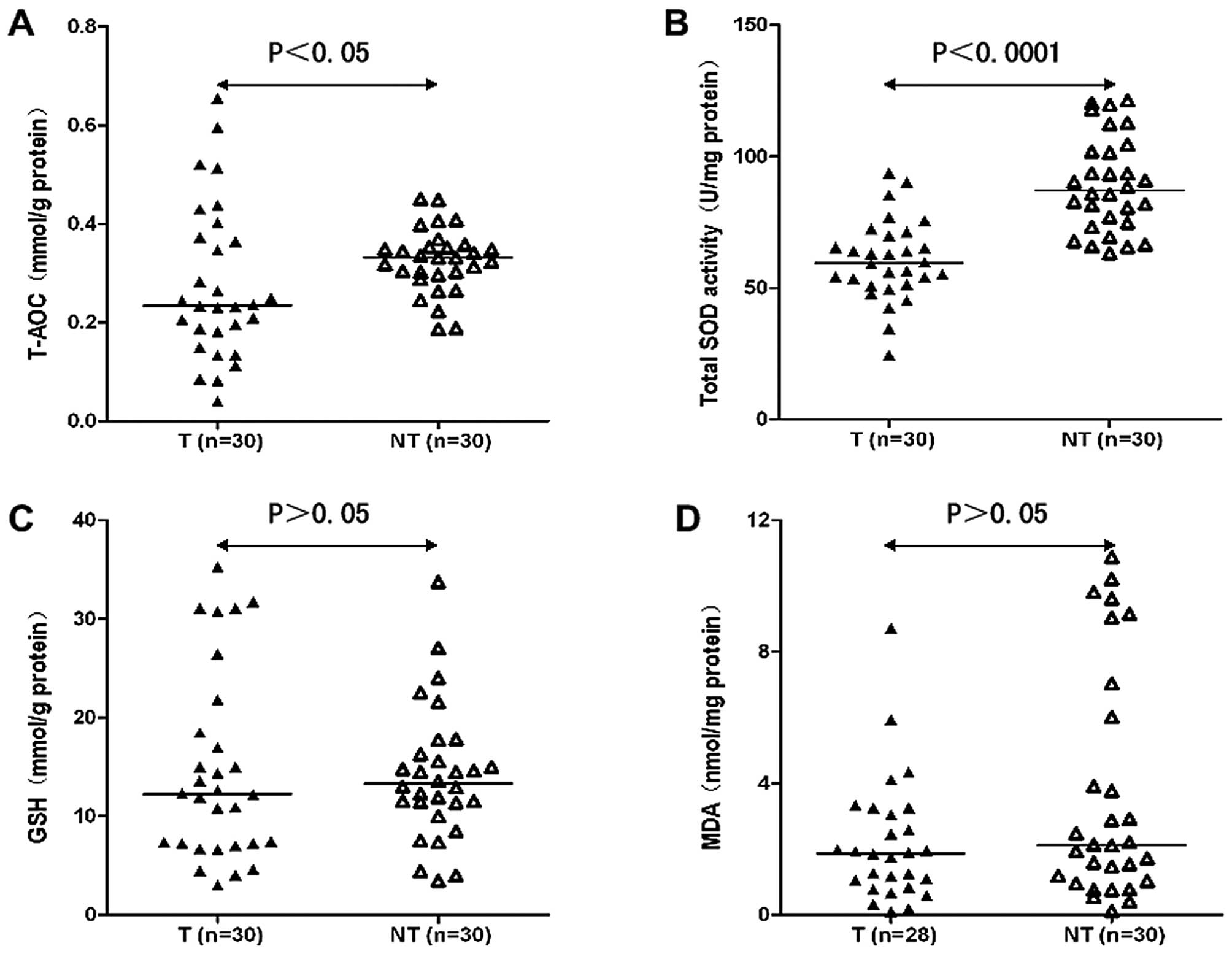
The T-AOC of HCC tissues (T) was lower compared to
that of non-tumor liver tissues (NT) (0.277±0.156 mmol/g protein in
T vs. 0.325±0.065 mmol/g protein in NT; P<0.05) (Fig. 1A), and the activity of total SOD
also tended to be significantly lower in T when compared with the
NT (60.420±15.222 U/mg protein in T vs. 89.381±18.064 U/mg protein
in NT; P<0.0001) (Fig. 1B). The
GSH content was not different between T and NT (14.595±9.531 vs.
14.140±6.699 mg/g protein; P>0.05) (Fig. 1C). The level of MDA was 2.200±1.884
nmol/mg protein in T, while it was 3.882±3.641 nmol/mg protein in
NT, and the difference was not significant between them (P>0.05)
(Fig. 1D).
Relationship between T-AOC and total SOD
activity in HCC tissues with clinicopathological features
As shown in Table
II, to elucidate the biological significance of T-AOC and total
SOD activity in HCC, we compared the levels of T-AOC and total SOD
activity with the clinicopathological features of HCC patients.
With respect to differentiation, we found that T-AOC and total SOD
activity were significantly lower in group I than in group II
(P<0.01 and P<0.05, respectively). On the other hand, it
should be noted that the level of T-AOC was significantly higher in
HCC cases with metastasis than without (P<0.05). Total SOD
activity was not different between them (P<0.05). There were no
significant differences regarding age, etiology, tumor number, TNM
stage.
| Table IIRelationship between T-AOC and total
SOD activity in HCC tissues with clinicopathological
features.a |
Table II
Relationship between T-AOC and total
SOD activity in HCC tissues with clinicopathological
features.a
| Factor | No. | T-AOC (mmol/g
protein) | P-value | Total SOD activity
(U/mg protein) | P-value |
|---|
| Age (years) | | | 0.093 | | 0.355 |
| ≤45 | 16 | 0.226±0.116 | | 57.968±15.069 | |
| >45 | 14 | 0.337±0.177 | | 63.222±15.460 | |
| Etiology | | | 0.981 | | 0.366 |
| HBV | 18 | 0.280±0.161 | | 57.840±14.336 | |
| HBV + alcohol | 10 | 0.301±0.156 | | 63.523±17.861 | |
| Tumor no. | | | 0.604 | | 0.500 |
| Single | 24 | 0.290±0.172 | | 59.843±16.676 | |
| Multiple | 6 | 0.225±0.035 | | 62.728±7.526 | |
|
Differentiationb | | | 0.007 | | 0.013 |
| Group I | 19 | 0.324±0.134 | | 65.531±13.298 | |
| Group II | 11 | 0.197±0.163 | | 51.592±14.773 | |
| Metastasis | | | 0.018 | | 0.909 |
| Yes | 8 | 0.388±0.176 | | 59.898±6.588 | |
| No | 22 | 0.237±0.130 | | 60.609±17.474 | |
| TNM stage | | | 0.098 | | 0.786 |
| I | 17 | 0.241±0.147 | | 60.342±19.438 | |
| II + II | 13 | 0.324±0.160 | | 60.522±7.491 | |
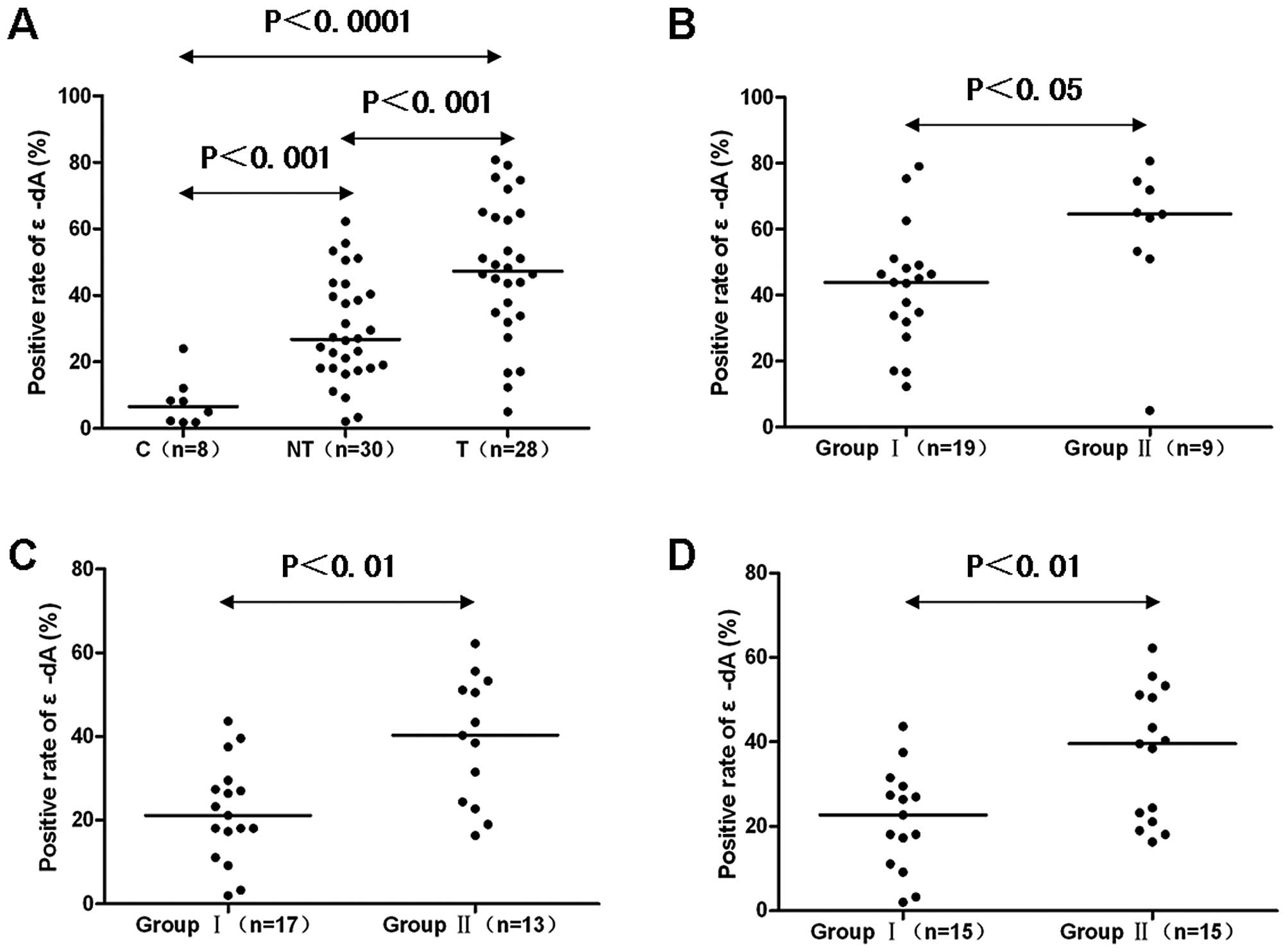
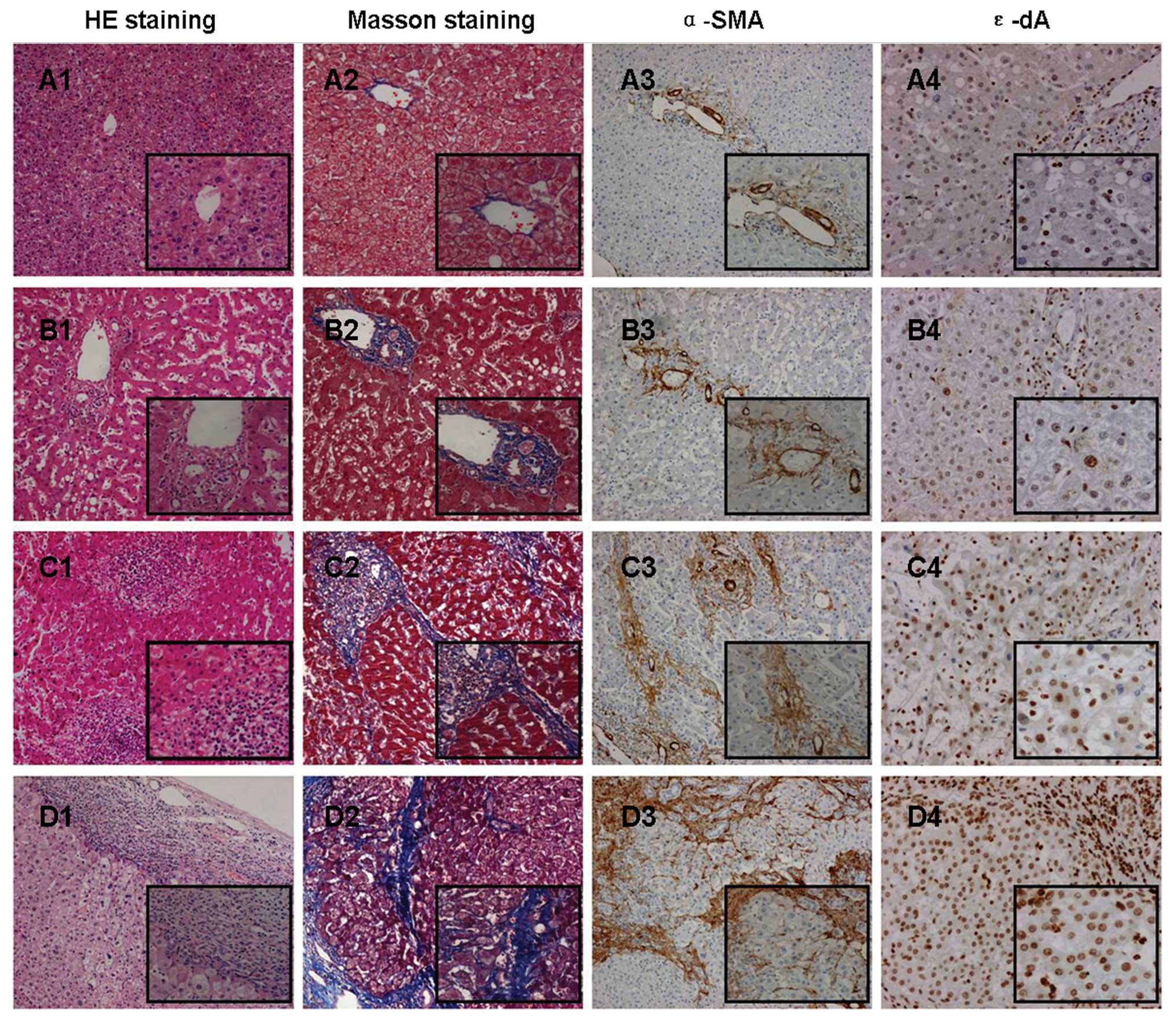
Comparison of ɛ-dA prevalence in control
liver, non-tumor liver and HCC tissues
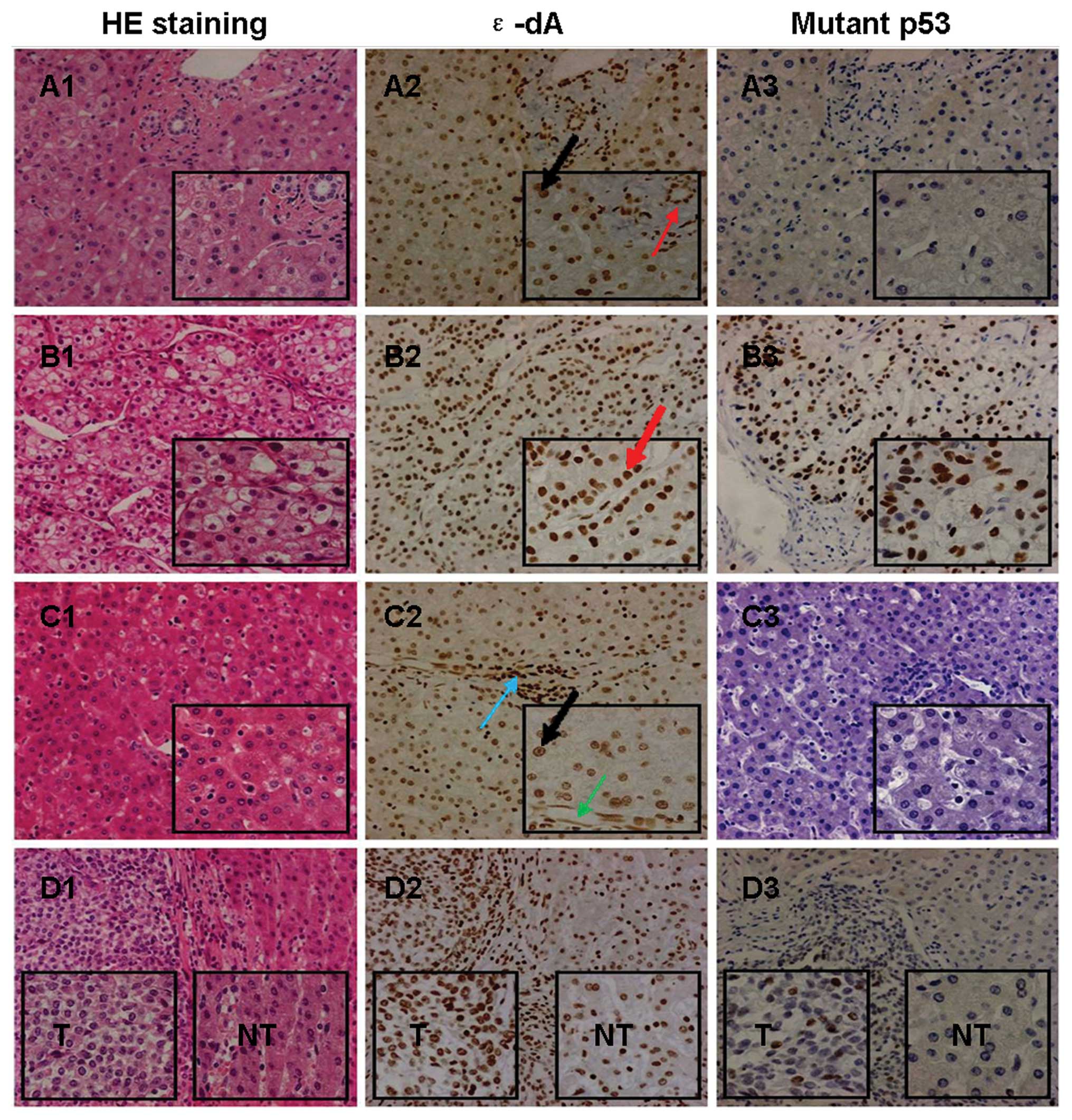
The prevalence of nuclei positive for ɛ-dA in
control livers was (7.9±7.5%), which was significantly increased to
(29.4±15.8%) in NT and (47.6±20.6%) in T (P<0.001 and
P<0.0001, respectively) (Figs.
2A and 3A2, B2 and C2). Also,
the percentage of positively stained nuclei was significantly
higher in T than in NT (P<0.001) (Figs. 2A and 3B2 and C2).
Relationship between positive rate of
ɛ-dA and clinicopathological features of HCC
We analyzed the relationship between positive rate
of ɛ-dA and clinical features of HCC patients. There was no
significant association between ɛ-dA positive expression and age,
etiology, tumor number, metastasis, TNM stage (data not shown). On
the other hand, the prevalence of ɛ-dA was positively correlated
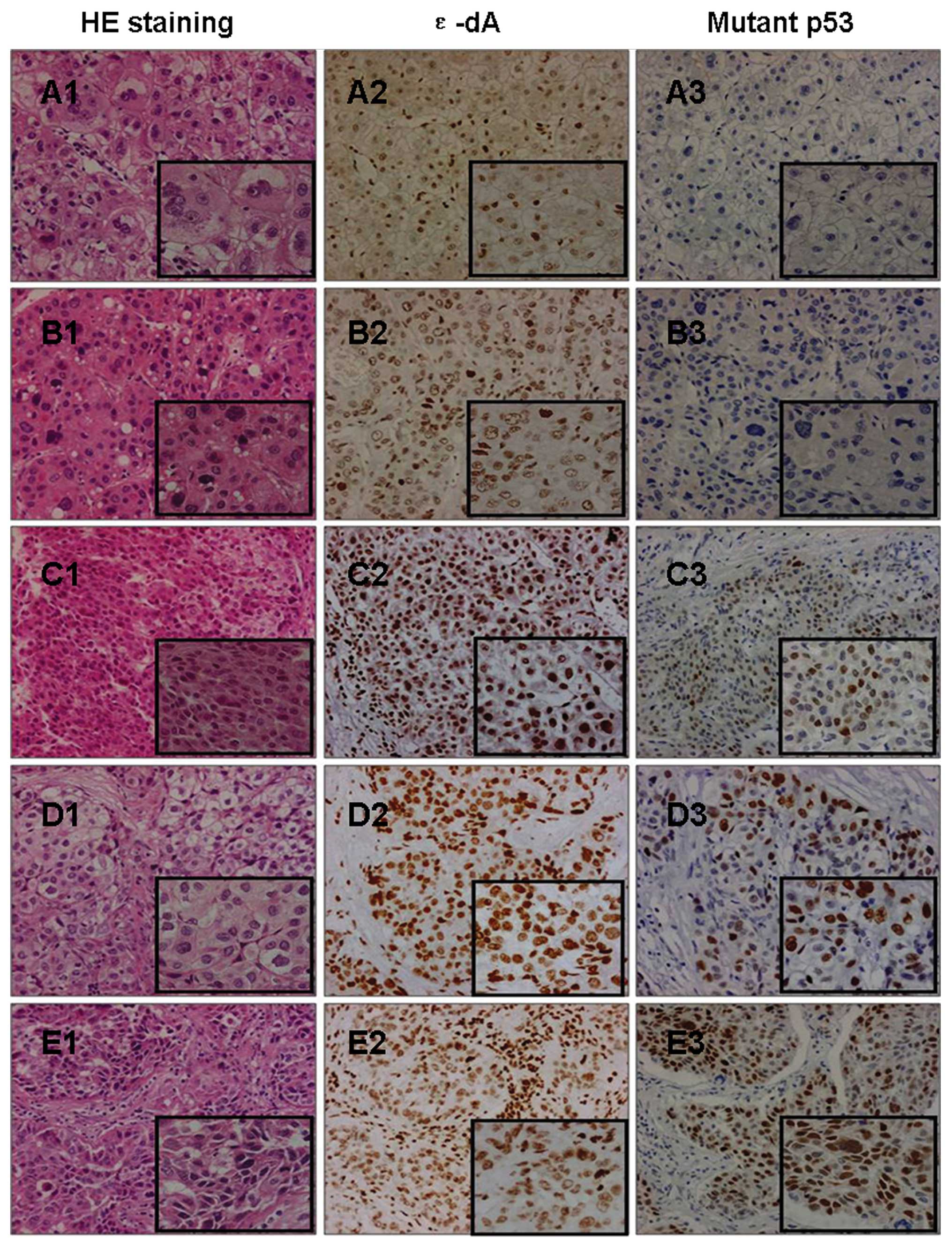
with differentiation. Group II showed a higher percentage of ɛ-dA
positivity than group I (58.8±22.3 vs. 42.2±17.9%; P<0.05)
(Figs. 2B and 4A2, B2, C2, D2 and E2).
Relationship between positive rate of
ɛ-dA and necroinflammatory activity and fibrosis extent of
non-tumor liver tissues
Based on the grade of necroinflammatory activity
(A), NT were subdivided into group I (A0 + A1) and group II (A2 +
A3). We found the positive percentage of ɛ-dA in group I was
significantly lower than in group II (21.9±11.9 vs. 39.1±15.2%;
P<0.01) (Figs. 2C and 5A4, B4, C4, and D4). Based on the stage of
fibrosis (F), NT were subdivided into group I (F0 + F1 + F2) and
group II (F3 + F4). The prevalence of nuclei positive for ɛ-dA in
group I was 21.7±12.0%, which was significantly increased to
37.1±15.6% in group II (P<0.01) (Figs. 2D and 5A4, B4, C4 and D4).
Nuclear expression of mutant p53 protein
in liver tissues
The nuclear expression of mutant p53 protein was
detectable in 26 of 30 T (86.7%) whereas this was not found in any
control (Fig. 3A3) or NT (Fig. 3C3). Only well (n=1) (Fig. 4A3), well-moderately (n=1) (Fig. 4B3), and moderately (n=2)
differentiated HCC showed negative expression of the mutant p53
protein. The expression levels of the mutant p53 protein tended to
be higher in group II compared to group I tumors (34.4±15.1 vs.
21.8±17.5%; P=0.062) (Fig. 4A3, B3, C3,
D3 and E3). No significant differences in mutant p53 expression
in association with age, etiology, tumor number, metastasis, TNM
stage were observed (data not shown). However, the positive rate of
ɛ-dA showed a significant correlation with the expression rate of
the mutant p53 protein in T (n=24, r=0.5162, P<0.01) (Figs. 4 and 6).
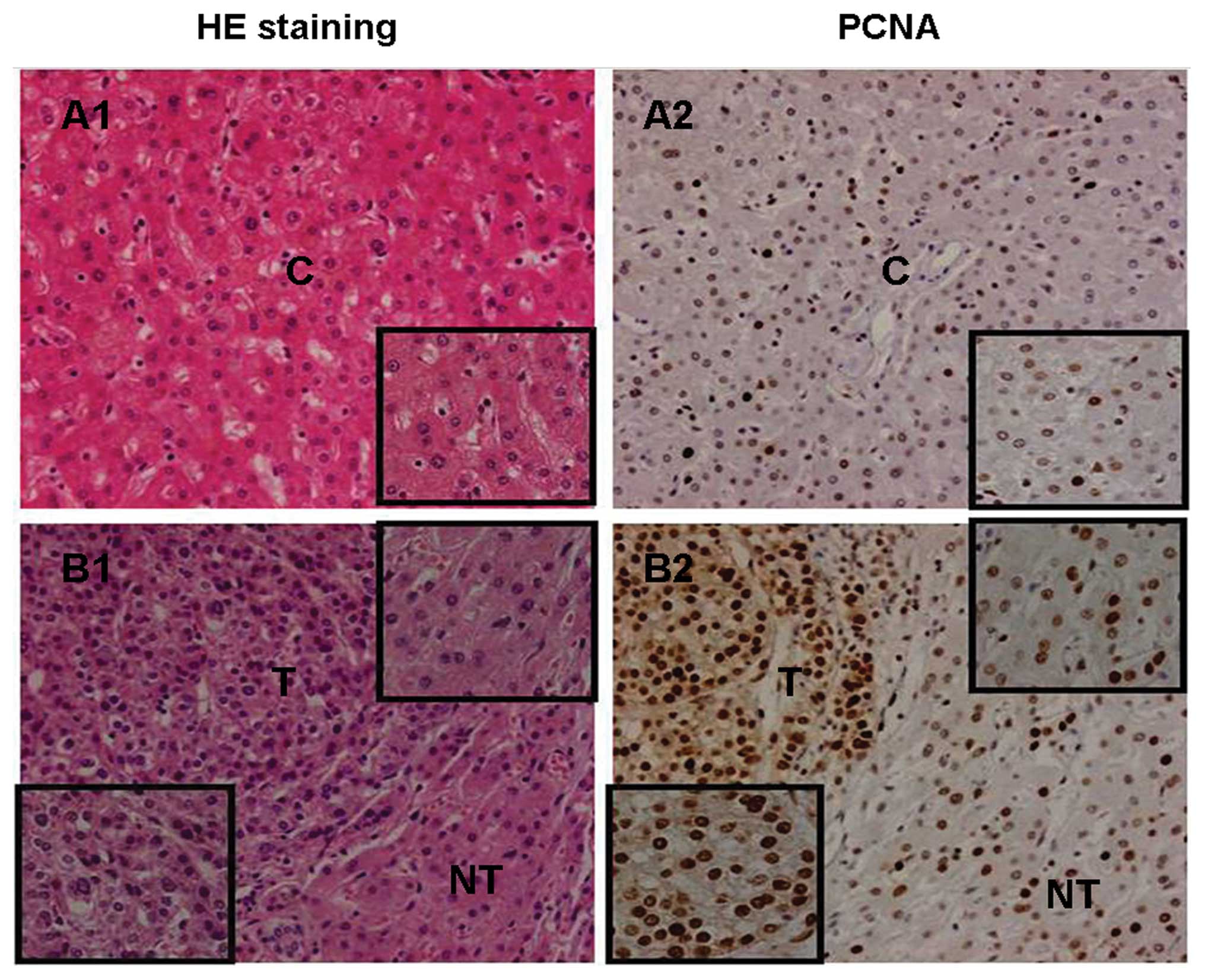
Positive expression of PCNA in liver
tissues
The positive rate of PCNA in T (47.6±10.9%) was
significantly higher than in the adjacent NT (34.1±8.2%,
P<0.0001) (Fig. 7B2) and control
livers (25.8±7.7%; P<0.0001) (Fig.
7A2). The positive rate of PCNA in the NT was higher compared
to the control livers (P<0.05) (Fig.
7A2 and B2). There was no significant association between PCNA
positive expression and any clinicopathological feature in T or NT
(data not shown).
Discussion
In the present study, the significant decrease in
the levels of T-AOC and total SOD activity in HCC tissue samples
provides evidence that the antioxidant system of HCC tissue was
severely impaired, and HCC tissue underwent serious oxidative
stress. The prevalence of nuclei positive for ɛ-dA in HCC tissues
was significantly higher compared to control and non-tumor liver
tissues; it is suggested that oxidative stress could induce the
excessive formation of ɛ-dA in HCC and ɛ-dA might be implicated in
hepatocarcinogenesis. The poorer differentiated HCC had the lower
levels of T-AOC and total SOD activity. On the contrary, the poorer
differentiated HCC had the higher prevalence of ɛ-dA. Our findings
are similar to a previous report that HCC tissues showed stronger
immunoreactivity of 8-hydroxy-2′-deoxyguanosine (8-OHdG) which
reflects oxidative DNA damage by oxidative stress than non-tumor
counterparts, and the number and positive rates of 8-OHdG-stained
hepatocytes was greater in poorly differentiated HCC than in well
and moderately differentiated HCC (21). These findings indicated that
oxidative DNA damage may be closely associated with the development
of HCC.
A previous report showed that the characteristic
patterns of p53 mutations were observed in liver tumors (including
HCC and liver angiosarcoma) associated with vinyl chloride exposure
in rats and humans. These mutations are consistent with the
pro-mutagenic properties of ɛ-DNA adducts by vinyl chloride in the
liver, that may be compatible with the hypothesis indicating
pro-mutagenic ɛ-DNA adducts as the initiating lesions (15). One study identified the mutations of
p53 in tumors induced by urethane in A/J mice, that also suggested
they could arise from ɛ-DNA adducts (16). In our study, we found the positive
prevalence of ɛ-dA and mutant p53 protein in HCC and hepatosarcoma
tissues (Fig. 3D1-3), and the
positive rate of ɛ-dA was significantly correlated with the
expression rate of the mutant p53 protein in HCC tissues. This
suggests that ɛ-dA may increase the incidence of p53 gene mutation
in HCC tissues. Moreover, we noted that the levels of T-AOC and
total SOD activity tended to be lower, but the expression levels of
ɛ-dA and mutant p53 tended to be higher in poorly differentiated
tumors compared to well and moderately differentiated tumors.
Collectively, these findings could be explained by the observation
that ROS can cause oxidative DNA damage, gene mutation or deletion,
a loss of p53 function, and a defect in DNA repair capacity. This
would promote genomic instability leading to oncogene activation,
aberrant metabolism, mitochondrial dysfunction and a deficit in
antioxidants. All these events can further increase ROS levels,
leading to more DNA damage and genomic instability. Such a vicious
cycle can effectively amplify oxidative stress and promote cancer
development and progression (22).
The wild-type p53 protein has a crucial role in
preventing oxidative damage to DNA, genetic instability, and
carcinogenesis. The p53 gene is one of the most common targets for
genetic alterations in HCC, being mutated and accumulated in tumor
tissues. Studies in vitro and in vivo confirm that
the expression of the mutant p53 protein can have a positive effect
on cell growth and drive the development of different types of
tumors (23,24). In our study, we found that the
mutant p53 protein was expressed in most HCCs, and the positive
rate of PCNA in the HCC was significantly higher than in the
adjacent non-tumor and control liver tissues, indicating that the
expression of mutant p53 protein has a positive effect on cell
proliferation and contributes to the development of HCC. Therefore,
the elevated accumulation of ɛ-dA in liver tissues may stimulate
cell proliferation and promote hepatocarcinogenesis by inducing the
overexpression of mutant p53 protein.
Chronic hepatitis and fibrosis/cirrhosis are the
preneoplastic liver lesions that characterize the setting in which
HCC most often develops. Kitada et al reported that 8-OHdG
was observed by immunohistochemistry in patients with various forms
of chronic liver diseases, and the number of positive hepatocytes
was significantly correlated with the severity of chronic hepatitis
activity (25). In our study, we
found that ɛ-dA levels were significantly higher in non-tumor liver
tissues with moderate and severe inflammation compared to those
with absent and mild inflammation, and many ɛ-dA-positive
hepatocytes were localized in contact with infiltrating lymphocytes
in non-tumor liver tissues (Fig.
5D4). These findings can be explained by the observation that
activated inflammatory cells represent a significant endogenous
source of ROS that is capable of inducing DNA damage and genomic
instability, and inflammatory cells may use cytokines as TNF-α to
stimulate ROS accumulation in neighboring cells (26). We also found that more severe
fibrosis development tended to have higher ɛ-dA levels in non-tumor
liver tissues. This finding is consistent with the fact that ROS
and other oxidative stress-related intermediates, released by
damaged hepatocytes or activated inflammatory cells, can activate
hepatic stellate cells (HSCs), which are collagen-producing cells
in the liver (27). Moreover, it
has been reported that activated HSCs are also an important source
of endogenous ROS in liver fibrogenesis, and this is mainly
associated with NADPH oxidase which is expressed in activated HSCs
(28). Taken together, these
findings provide evidence of a possible link between ROS-generated
ɛ-dA and persistent inflammation and fibrosis in chronic liver
diseases. It is possible to hypothesize that chronic inflammation
and fibrosis could facilitate continuous accumulation of oxidative
ɛ-DNA damage, which may partly drive chronic inflammatory and
fibrotic/cirrhotic liver tissues to HCC. Therefore, measurement of
ɛ-dA in liver tissue could be explored as a potential biomarker for
disease progression and cancer risk assessment in patients with
cancer-prone liver diseases.
Excess levels of ROS-derived ɛ-DNA adducts have been
measured in the patients with inflammatory cancer-prone liver
diseases. Using a sensitive
immunoaffinity/32P-postlabeling method, elevated levels
of ɛ-DNA adducts were detected in the liver of patients with
primary hemochromatosis and Wilson’s disease (17). Excess ɛ-dA levels were detected in
liver biopsy specimens obtained from European alcoholic liver
disease (ALD) patients diagnosed with alcohol-related hepatitis,
fibrosis and cirrhosis by immunohistochemistry (18). Furthermore, using an ultrasensitive
and specific immunoprecipitation/high performance liquid
chromatography (HPLC)-fluorescence detection method for quantifying
ɛ-dA excreted in urine, significantly higher urinary ɛ-dA levels
were also found in HBV and HCV-related liver disease and ALD
patients compared with controls (19). These findings show that ɛ-dA may
play a driving role towards hepatocarcinogenesis in cancer-prone
liver diseases.
The promutagenic properties of ROS-derived ɛ-DNA
adducts were established by studies in E. coli and in
mammalian cells (29). ɛ-DNA
adducts produced mainly base pair substitution mutations. ɛ-dA can
lead to AT→GC transitions, to AT→TA and AT→CG transversions
(30,31). ɛ-dC can cause CG→TA transitions and
CG→AT transversions (32,33). N2,
3-ethenodeoxyguanosine, that is also formed in vivo from LPO
products, can lead to GC→AT transitions (33). These miscoding properties of ɛ-bases
strongly implicate them in the initiation of carcinogenesis by
vinyl chloride (34), urethane
(35,36) and other ɛ-adduct forming chemicals.
Incorporation of a single ɛ-dA in either DNA strand of HeLa cells
showed a similar miscoding frequency and was more mutagenic than
8-oxo-deoxyguanosine (37).
In conclusion, we have demonstrated that ROS-induced
ɛ-dA lesions may be relevant to the severity of inflammation and
fibrosis in chronic liver diseases. The elevated accumulation of
ɛ-dA partly contributes to the mutant p53 protein overexpression
and excessive cell proliferation, eventually leading to
hepatocarcinogenesis.
Acknowledgements
The authors thank the patients who participated in
this study. We thank Dr Cuihuan Wu, Department of Pathology, Tongji
Hospital for assistance with histopathological examination and Dr
Zhiwei Zhang, Department of Liver Surgery, Tongji Hospital for
assistance with liver sample collection. This study was funded by
the National Natural Science Foundation of China (grant no.
81071999).
References
|
1
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
3
|
Donato F, Boffetta P and Puoti M: A
meta-analysis of epidemiological studies on the combined effect of
hepatitis B and C virus infections in causing hepatocellular
carcinoma. Int J Cancer. 75:347–354. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Seitz HK and Stickel F: Risk factors and
mechanisms of hepatocarcinogenesis with special emphasis on alcohol
and oxidative stress. Biol Chem. 387:349–360. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Severi T, Ying C, Vermeesch JR, Cassiman
D, Cnops L, Verslype C, Fevery J, Arckens L, Neyts J and van Pelt
JF: Hepatitis B virus replication causes oxidative stress in
HepAD38 liver cells. Mol Cell Biochem. 290:79–85. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hsieh YH, Su IJ, Wang HC, Chang WW, Lei
HY, Lai MD, Chang WT and Huang W: Pre-S mutant surface antigens in
chronic hepatitis B virus infection induce oxidative stress and DNA
damage. Carcinogenesis. 25:2023–2032. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qadri I, Iwahashi M, Capasso JM, Hopken
MW, Flores S, Schaack J and Simon FR: Induced oxidative stress and
activated expression of manganese superoxide dismutase during
hepatitis C virus replication: role of JNK, p38 MAPK and AP-1.
Biochem J. 378:919–928. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moriya K, Nakagawa K, Santa T, Shintani Y,
Fujie H, Miyoshi H, Tsutsumi T, Miyazawa T, Ishibashi K, Horie T,
Imai K, Todoroki T, Kimura S and Koike K: Oxidative stress in the
absence of inflammation in a mouse model for hepatitis C
virus-associated hepatocarcinogenesis. Cancer Res. 61:4365–4370.
2001.PubMed/NCBI
|
|
9
|
Wang Y, Millonig G, Nair J, Patsenker E,
Stickel F, Mueller S, Bartsch H and Seitz HK: Ethanol-induced
cytochrome P4502E1 causes carcinogenic etheno-DNA lesions in
alcoholic liver disease. Hepatology. 50:453–461. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu HC, Wang Q, Wang LW, Yang HI, Ahsan H,
Tsai WY, Wang LY, Chen SY, Chen CJ and Santella RM: Urinary
8-oxodeoxyguanosine, aflatoxin B1 exposure and hepatitis B virus
infection and hepatocellular carcinoma in Taiwan. Carcinogenesis.
28:995–999. 2007.PubMed/NCBI
|
|
11
|
Starley BQ, Calcagno CJ and Harrison SA:
Nonalcoholic fatty liver disease and hepatocellular carcinoma: a
weighty connection. Hepatology. 51:1820–1832. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Toyokuni S, Okamoto K, Yodoi J and Hiai H:
Persistent oxidative stress in cancer. FEBS Lett. 358:1–3. 1995.
View Article : Google Scholar
|
|
13
|
Bartsch H and Nair J: Chronic inflammation
and oxidative stress in the genesis and perpetuation of cancer:
role of lipid peroxidation, DNA damage, and repair. Langenbecks
Arch Surg. 391:499–510. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
el Ghissassi F, Barbin A, Nair J and
Bartsch H: Formation of 1,
N6-ethenoadenine and 3,
N4-ethenocytosine by lipid peroxidation
products and nucleic acid bases. Chem Res Toxicol. 8:278–283.
1995.
|
|
15
|
Barbin A, Froment O, Boivin S, Marion MJ,
Belpoggi F, Maltoni C and Montesano R: p53 gene mutation pattern in
rat liver tumors induced by vinyl chloride. Cancer Res.
57:1695–1698. 1997.PubMed/NCBI
|
|
16
|
Horio Y, Chen A, Rice P, Roth JA,
Malkinson AM and Schrump DS: Ki-ras and p53 mutations are early and
late events, respectively, in urethane-induced pulmonary
carcinogenesis in A/J mice. Mol Carcinog. 17:217–223. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nair J, Carmichael PL, Fernando RC,
Phillips DH, Strain AJ and Bartsch H: Lipid peroxidation-induced
etheno-DNA adducts in the liver of patients with the genetic metal
storage disorders Wilson’s disease and primary hemochromatosis.
Cancer Epidemiol Biomarkers Prev. 7:435–440. 1998.PubMed/NCBI
|
|
18
|
Frank A, Seitz HK, Bartsch H, Frank N and
Nair J: Immunohistochemical detection of 1,
N6-ethenodeoxyadenosine in nuclei of human
liver affected by diseases predisposing to hepatocarcinogenesis.
Carcinogenesis. 25:1027–1031. 2004.
|
|
19
|
Nair J, Srivatanakul P, Haas C,
Jedpiyawongse A, Khuhaprema T, Seitz HK and Bartsch H: High urinary
excretion of lipid peroxidation-derived DNA damage in patients with
cancer-prone liver diseases. Mutat Res. 683:23–28. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Desmet VJ, Gerber M, Hoofnagle JH, Manns M
and Scheuer PJ: Classification of chronic hepatitis: diagnosis,
grading and staging. Hepatology. 19:1513–1520. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ichiba M, Maeta Y, Mukoyama T, Saeki T,
Yasui S, Kanbe T, Okano J, Tanabe Y, Hirooka Y, Yamada S, Kurimasa
A, Murawaki Y and Shiota G: Expression of
8-hydroxy-2′-deoxyguanosine in chronic liver disease and
hepatocellular carcinoma. Liver Int. 23:338–345. 2003.
|
|
22
|
Trachootham D, Alexandre J and Huang P:
Targeting cancer cells by ROS-mediated mechanisms: a radical
therapeutic approach? Nat Rev Drug Discov. 8:579–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bossi G, Lapi E, Strano S, Rinaldo C,
Blandino G and Sacchi A: Mutant p53 gain of function: reduction of
tumor malignancy of human cancer cell lines through abrogation of
mutant p53 expression. Oncogene. 25:304–309. 2006.PubMed/NCBI
|
|
24
|
Shaulsky G, Goldfinger N and Rotter V:
Alterations in tumor development in vivo mediated by expression of
wild type or mutant p53 proteins. Cancer Res. 51:5232–5237.
1991.PubMed/NCBI
|
|
25
|
Kitada T, Seki S, Iwai S, Yamada T,
Sakaguchi H and Wakasa K: In situ detection of oxidative DNA
damage, 8-hydroxydeoxyguanosine, in chronic human liver disease. J
Hepatol. 35:613–618. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar
|
|
27
|
Novo E and Parola M: Redox mechanisms in
hepatic chronic wound healing and fibrogenesis. Fibrogenesis Tissue
Repair. 1:52008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bataller R, Schwabe RF, Choi YH, Yang L,
Paik YH, Lindquist J, Qian T, Schoonhoven R, Hagedorn CH, Lemasters
JJ and Brenner DA: NADPH oxidase signal transduces angiotensin II
in hepatic stellate cells and is critical in hepatic fibrosis. J
Clin Invest. 112:1383–1394. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bartsch H, Barbin A, Marion MJ, Nair J and
Guichard Y: Formation, detection, and role in carcinogenesis of
ethenobases in DNA. Drug Metab Rev. 26:349–371. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Basu AK, Wood ML, Niedernhofer LJ, Ramos
LA and Essigmann JM: Mutagenic and genotoxic effects of three vinyl
chloride-induced DNA lesions: 1,
N6-ethenoadenine, 3,
N4-ethenocytosine, and
4-amino-5-(imidazol-2-yl)imidazole. Biochemistry. 32:12793–12801.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pandya GA and Moriya M: 1,
N6-ethenodeoxyadenosine, a DNA adduct
highly mutagenic in mammalian cells. Biochemistry. 35:11487–11492.
1996.
|
|
32
|
Palejwala VA, Rzepka RW, Simha D and
Humayun MZ: Quantitative multiplex sequence analysis of mutational
hot spots. Frequency and specificity of mutations induced by a
site-specific ethenocytosine in M13 viral DNA. Biochemistry.
32:4105–4111. 1993. View Article : Google Scholar
|
|
33
|
Moriya M, Zhang W, Johnson F and Grollman
AP: Mutagenic potency of exocyclic DNA adducts: marked differences
between Escherichia coli and simian kidney cells. Proc Natl
Acad Sci USA. 91:11899–11903. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng KC, Preston BD, Cahill DS, Dosanjh
MK, Singer B and Loeb LA: The vinyl chloride DNA derivative
N2, 3-ethenoguanine produces G→A
transitions in Escherichia coli. Proc Natl Acad Sci USA.
88:9974–9978. 1991.PubMed/NCBI
|
|
35
|
Barbin A, Bartsch H, Leconte P and Radman
M: Studies on the miscoding properties 1,
N6-ethenoadenine and 3,
N4-ethenocytosine, DNA reaction products
of vinyl chloride metabolites, during in vitro DNA synthesis.
Nucleic Acids Res. 9:375–387. 1981.
|
|
36
|
Miller JA and Miller EC: The metabolic
activation and nucleic acid adducts of naturally-occurring
carcinogens: recent results with ethyl carbamate and the spice
flavors safrole and estragole. Br J Cancer. 48:1–15. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Levine RL, Yang IY, Hossain M, Pandya GA,
Grollman AP and Moriya M: Muta-genesis induced by a single 1,
N6-ethenodeoxyadenosine adduct in human
cells. Cancer Res. 60:4098–4104. 2000.PubMed/NCBI
|