Introduction
Human CD99 is a 32 kDa transmembrane glycoprotein
encoded by a pseudoautosomal MIC2 gene located in Xp22.33-pter and
Yp11-pter (1). It is expressed in
several types of cells and is involved in several cellular events
such as intercellular adhesion between lymphocytes and endothelial
cells (2), migration of immune
cells to inflammatory sites (3,4) and
attenuation of graft-versus-host disease (5). Certain studies have found that CD99 is
expressed in tumors, including lymphoblastic lymphoma/leukaemia
(6), Ewing’s sarcoma/primitive
neuroectodermal tumors (ES/PNET) (7), dermatofibrosarcoma protuberans, as
well as giant cell fibroblastoma (8) and hepatoblastomas (9), while others found that CD99 is
expressed in benign pancreatic endocrine neoplasm and gastric
adenocarcinoma (10), but not in
malignant counterparts.
Mouse CD99 antigen-like2 (mCD99L2), a widely
expressed antigen of unknown function with moderate sequence
homology to CD99, was identified and cloned by Suh et
al(11). mCD99L2 mRNA is widely
distributed in neuronal cells, choroid plexus, sertoli cells and
ovarian granulosa and thecal cells. It acts during leukocyte
extravasation in mouse leukocytes and endothelial cells (12) to help neutrophils penetrate the
endothelial basement membrane (13). Amino acid sequence alignment showed
five putative functional regions highly conserved between mCD99L2
and human CD99, implying they may have similar functions. However,
to date, the majority of studies on mCD99L2 have focused mainly on
its roles in inflammation. Its effects on tumor cells have yet to
be reported.
A20 is a murine cell line derived from a
spontaneously arising tumor in an aged BALB/c mouse (14). It pathologically mimics the
characteristics of human diffuse large B cell lymphoma (15). An A20-related animal model may be
used in studies on the association between tumors and hosts
(16). We successfully established
a disseminated A20 animal model and described its
immuno-characteristics (17). We
also observed that mCD99L2 was expressed in A20 cells (18). To investigate the role of mCD99L2 in
A20 cells, four pLenti6/mCD99L2 expression vectors containing the
mCD99L2 shRNA-expressing cassette were constructed in the present
study, transfected into A20 cells using DMRIE-C2, and the stable
mCD99L2-downregulated A20 cell line, termed A20-mCD99L2−
cells, was established and identified using quantitative PCR and
western blot analysis. The effect of shRNA targeting mCD99L2 during
continuous culturing was observed. The morphological, biological
and phenotypic characteristics of the A20-mCD99L2− cells
were extensively investigated in vitro and in vivo,
to provide data for additional functional studies of mCD99L2 in
associated tumors.
Using CD99-deficient IM9 and BJAB B cell lines,
investigators have confirmed that the downregulation of CD99 is a
primary requirement for generating ‘Hodgkin and Reed-Sternberg’
(H/RS) cells (19,20). A previous study of ours suggested
that the CD99-upregulated H/RS cell line (L428) lost its nature as
H/RS cells (21). In this study, we
investigated whether mCD99L2 downregulation can induce cells with
an H/RS morphology and phenotypes in murine B lymphoma cells, to
determine whether there is a functional similarity between human
CD99 and murine CD99 genes.
Materials and methods
Cell lines and morphology
observation
The BALB/c-derived mouse B lymphoma A20 cell line
was kindly provided by Professor Chan of the Nebraska Medical
Center, Omaha, NE, USA. The subclones of A20 cells transfected with
shRNAs targeting mCD99L2 or negative control vectors were
constructed and cultured in RPMI-1640 medium supplemented with 10%
heat-inactivated fetal bovine serum (FBS) (Gibco, Grand Island, NY,
USA).
Morphological observation of live cells was
conducted with an inverted microscope. Transmission electron
microscopy (TEM) was performed according to the manufacturer’s
instructions. Hematoxylin and eosin (H&E) staining was applied
on the fixed cells previously dripped onto slides and preserved
under −80°C.
Preparation of lentiviral vectors and RNA
interference (RNAi)
Four different sequences targeting the mCD99L2 gene
were selected by BLOCK-iT™ RNAi Designer (Invitrogen, Carlsbad, CA,
USA). The preparation of lentiviral vectors expressing mCD99L2
short hairpin RNA (shRNA) was performed using the BLOCK-iT
Lentiviral RNAi Expression System (catalog no. K4944-00;
Invitrogen). Four pLenti6/mCD99L2 expression vectors containing the
mCD99L2 shRNA-expressing cassette were constructed. The lentiviral
vectors containing the human Lamin A/C shRNA-expressing cassette
(sequence 5′-CTGGACTTCCAGAAGAACA-3′) were used as the positive
control and the pLenti6/U6 mock vector was used as the negative
control. A20 cells were transfected with specific or negative
control lentiviral vectors using DMRIE-C2 at suitable ratios and
selected for stable integrants by culturing in complete medium
containing blasticidin (Invitrogen). Several single
blasticidin-resistant colonies were isolated using the soft agar
clone formation protocol, expanded into sub-cell lines by 96-well
plate limiting dilution assay.
RNA isolation and quantitative PCR
Total RNA was extracted using the Takara RNAiso plus
kit and cDNA was prepared from 2 μg total RNA by PrimeScript
Reverse Transcriptase (Takara Bio Co., Ltd., Shiga, Japan).
Real-time PCR was performed on a 7500/7500 Fast Real-Time PCR
System (Applied Biosystem, Foster City, CA, USA) using a SYBR-Green
Premix Ex Taq™ kit (Takara), following the manufacturer’s
instructions under the conditions of 95°C for 30 sec, followed by
40 cycles of 95°C for 5 sec and 58°C for 34 sec. PCR primers were
purchased from Invitrogen and were as follows: mCD99L2, forward,
5′-GCCCAGCAACAAGCAAAGCACAT-3′ and reverse,
5′-CCCAACCACCCTAGTTCCTCCG-3′; GAPDH, forward,
5′-ACAGTCAGCCGCATCTTCTT-3′ and reverse, 5′-GACAA
GCTTCCCGTTCTCAG-3′. The results were analyzed using the software
installed in the 7500/7500 Fast Real-Time PCR System (Applied
Biosystems) and the relative expression ratio was determined using
the formula 2−ΔΔCt.
Cell proliferation
Cell proliferation was analyzed using the MTT assay
(Sigma, St. Louis, MO, USA). Briefly, 1×103 cells were
seeded into each well of a 96-well plate with quadruplicate repeats
for each condition. After 24 h of incubation, cells were mixed with
MTT reagent and incubated for 4 h. The formazan crystals formed by
viable cells were then solubilized in dimethyl sulfoxide (DMSO) and
measured at 490 nm. Each experiment was performed in
triplicate.
Flow cytometry
The cultured cells were harvested at the exponential
growth phase and prepared as single cell suspensions. Cells
(1×106) were fixed in 70% ethanol in phosphate-buffered
saline (PBS) on ice, pelleted, incubated with RNase A (0.1 μg/ml)
for 30 min at 37°C and stained with propidium iodide (PI) (40
μg/ml) for cell cycle analysis.
Cells were stained with panels containing
fluorescein isothiocyanate (FITC)-conjugated anti-mouse antibodies
against CD19 and CD20, R-Phycoerythrin (PE)-conjugated anti-mouse
antibodies against CD30 and CD15, and control FITC- or
PE-conjugated mouse IgG1 (BD Pharmingen, San Diego, CA, USA), as
indicated. CD antigen expressions were analyzed on a FACSCalibur
machine (ELITE; Beckman-Coulter, Fullerton, CA, USA).
Immunofluorescence
Cells (2.0×105/ml) were inoculated into
each well of 6-well plates (Costar, Corning, NY, USA) and cultured
in complete medium for 48 h followed by in serum-free medium for
another 24 h. After deposition, fixation and permeabilization, the
cells were labeled with rabbit anti-mouse CD30 mAb (2 μg/ml; Abcam)
followed with PE-conjugated goat anti-rabbit IgG (15 μg/ml,
ZF-0311; ZSGB-BIO, Beijing, China). Negative controls were
performed by replacing the primary antibodies with PBS. The cells
were observed under a fluorescence microscope (Nikon, Tokyo,
Japan).
Mouse cytokine antibody arrays
RayBio® Mouse Cytokine Antibody Arrays
(RayBiotech, Inc., Norcross, GA, USA) were used to investigate the
expression of 62 cytokines in the different cell groups, according
to the manufacturer’s instructions. In brief, proteins were
extracted, quantified and transferred onto membranes. The membranes
were then sealed and incubated with antibodies against cytokines.
The differences in cytokine expression were visualized,
photographed and analyzed.
Western blot analysis
Cell lysates were prepared, and equal amounts of
protein (50 μg) were separated on 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and
transferred onto polyvinylidene difluoride (PVDF) membranes
(Bio-Rad Laboratories, Hercules, CA, USA). Membranes were incubated
with 5% skim milk in TBS-0.1% Tween-20 for 2 h to block the
residual binding sites followed by immunoblotting overnight at 4°C
with appropriately diluted rabbit anti-human p-IκBα antibody
(1:500; Bioworld Technology, Inc., St. Louis Park, MN, USA), rabbit
mCD99L2 antibody (Abcam, Cambridge, MA, USA) and rabbit β-actin
antibody (ZhongShan Golden Bridge Biotechnology, Bejing, China).
Specific binding was revealed by mouse HRP-conjugated anti-rabbit
IgG (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and an
enhanced chemiluminescence system (ECL-Plus; Amersham Biosciences
Inc., Piscataway, NJ, USA).
Animals and in vivo tests
Twelve nude mice and 84 BALB/c mice (six to
eight-week-old female/male) were purchased from the Central
Laboratory of Animal Science of the Southern Medical University
(Guangzhou, China) and randomized into an A20-mCD99L2−
group and an A20-empty group of nude and BALB/c mice, respectively
(Table I). Tumor cells
(2×106 to 2×107) in 0.1–0.4 ml growth medium
were injected into the mice using various methods. Tumor growth was
observed by calculating the tumor volume. Mice were sacrificed when
exhibiting external signs of suffering (such as reduced mobility
and altered behavior). The procedures were conducted under sterile
conditions. The animal protocol for this experiment was approved by
the Animal Care and Use Committee of the Southern Medical
University.
 | Table ISequences of shRNA targeting
mCD99L2. |
Table I
Sequences of shRNA targeting
mCD99L2.
| No. | | Sequences |
|---|
| 1 | Top strand |
5′-CACCGCCTTGTCCAGAGAGGATATCGAAATATCCTCTCTGGACAAGG-3′ |
| Bottom strand | 5′-AAAACCTTGTCCAGAGAGGATATTTCGATATCCTCTCTGGACAAGGC-3′ |
| 2 | Top strand |
5′-CACCGCCACTACTACAACTAGAACGAATTCTAGTTGTAGTAGTGGC-3′ |
| Bottom strand | 5′-AAAAGCCACTACTACAACTAGAATTCGTTCTAGTTGTAGTAGTGGC-3′ |
| 3 | Top strand |
5′-CACCGGAAGATGCCTTGGATGATCGAAATCATCCAAGGCATCTTCC-3′ |
| Bottom strand | 5′-AAAAGGAAGATGCCTTGGATGATTTCGATCATCCAAGGCATCTTCC-3′ |
| 4 | Top strand |
5′-CACCGCCTTGCTATGGCCCTGATTCGAAAATCAGGGCCATAGCAAGG-3′ |
| Bottom strand | 5′-AAAACCTTGCTATGGCCCTGATTTTCGAATCAGGGCCATAGCAAGGC-3′ |
Immunohistochemistry
Tissue specimens were collected from tumors in the
mice and fixed in formalin and embedded in paraffin. The tissues
were then cut into 2-μm sections and dried on capillary-gap glass
slides. Immunochemistry of the paraffin sections was carried out
using the ChemMate™ EnVision™ Detection kit (Dako, Carpinteria, CA,
USA). The sections were dewaxed, dehydrated, subjected to antigen
retrieval and blocked for endogenous peroxidase activity. The
sections were then immunostained with CD3 antibody (brown color)
with DAB substrate (Dako) and then counterstained using
hematoxylin.
Statistical analysis
The SPSS 13.0 software was used for statistical
analysis. The results are expressed as the means ± standard
deviation (SD). Where indicated, differences were compared using
the Student’s t-test. Assay differences between in vitro
cell growth and in vivo tumor growth were examined for
statistical significance using analysis of variance (ANOVA) for
factorial design. Proliferation assay and FACS results of antigen
expression or lymphocyte percentages were examined using one-way
ANOVA. Statistical analysis of tumor growth was carried out using
Dunnett’s multiple comparison tests. P<0.05 was considered to
indicate a statistically significant difference.
Results
Establishment of A20 subclones using
shRNA targeting mCD99L2 gene
Four primers were designed based on the RNAi
technique, shRNA was constructed and sequencing was confirmed
followed by transient transfection into A20 cells (Table II). Interference efficacy was
examined using real-time RT-PCR until targeting sequences were
screened. No. 2 lentiviral vector was most effective at blocking
mCD99L2 expression (Table
II, no. 2 shRNA). Subsequently, the pLenti6/mCD99L2 (no.
2) and pLenti6/U6 mock vector were transfected into the A20 cells
and blasticidin-resistant single clones were selected, achieving
permanent transfection to gain stable integrants. The transfection
rate was 57% at 16 days assayed by PI staining flow cytometry. For
the sake of convenience, clones transfected using
pLenti6/mCD99L2 or pLenti6/U6 mock vector were termed
A20-mCD99L2− cells and A20-empty cells, respectively.
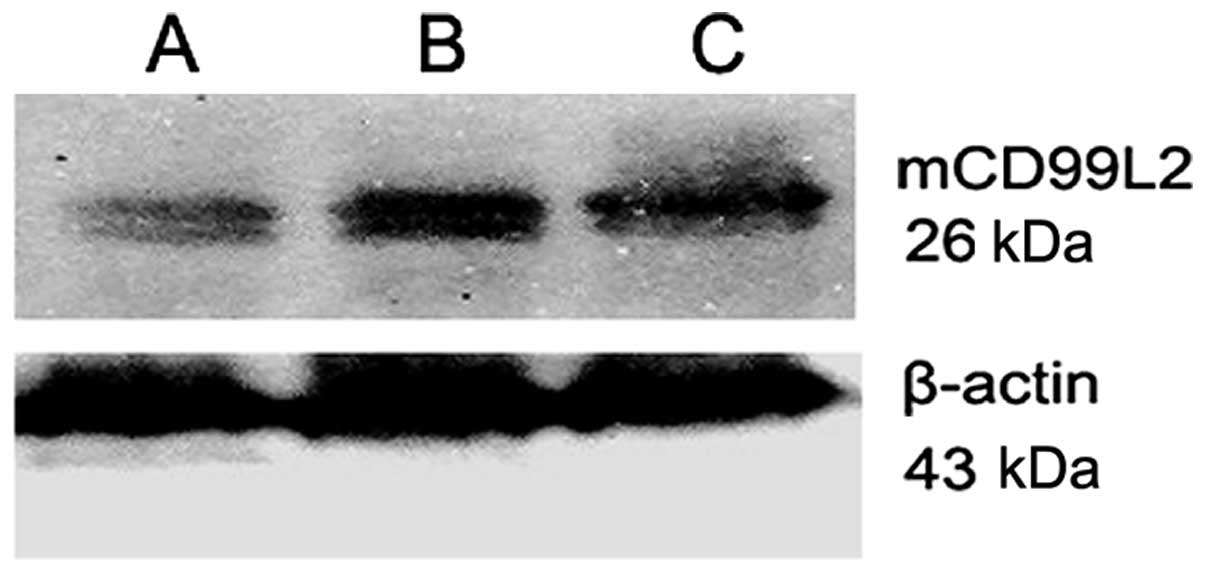
Stable integrants of low mCD99L2 expression of the A20 cells were
achieved (A20-mCD99L2− cells) and confirmed in various
clones. mCD99L2 expression was significantly lower in the
A20-mCD99L2− cell group compared with the A20 and
A20-empty groups; it decreased by 50%, as indicated by real-time
PCR and western blot analysis (Fig.
1).
| Table IIAssignment of animals groups. |
Table II
Assignment of animals groups.
| Group | Animal (no.) | Methods | Cells inoculated
(/mice) | Inoculation
site |
|---|
| A | Nude mouse (6) | Subcutaneous
inoculation |
2×107/0.2 ml | Axillary fossa |
| B1 | BALB/c mouse
(7) | Subcutaneous
inoculation |
2×106/0.1 ml | Left axillary
fossa |
| B2 | BALB/c mouse
(7) | Subcutaneous
inoculation |
2×107/0.1 ml | Left axillary
fossa |
| C1 | BALB/c mouse
(7) | Subcutaneous
transplantation | Tumor tissue | Left axillary
fossa |
| C2 | BALB/c mouse
(7) | Subcutaneous
transplantation | Tumor tissue | Right axillary
fossa |
| D1 | BALB/c mouse
(7) | Subcutaneous
transplantation |
5×106/0.2 ml | Caudal vein |
| D2 | BALB/c mouse
(7) | Subcutaneous
transplantation |
2×107/0.4 ml | Caudal vein |
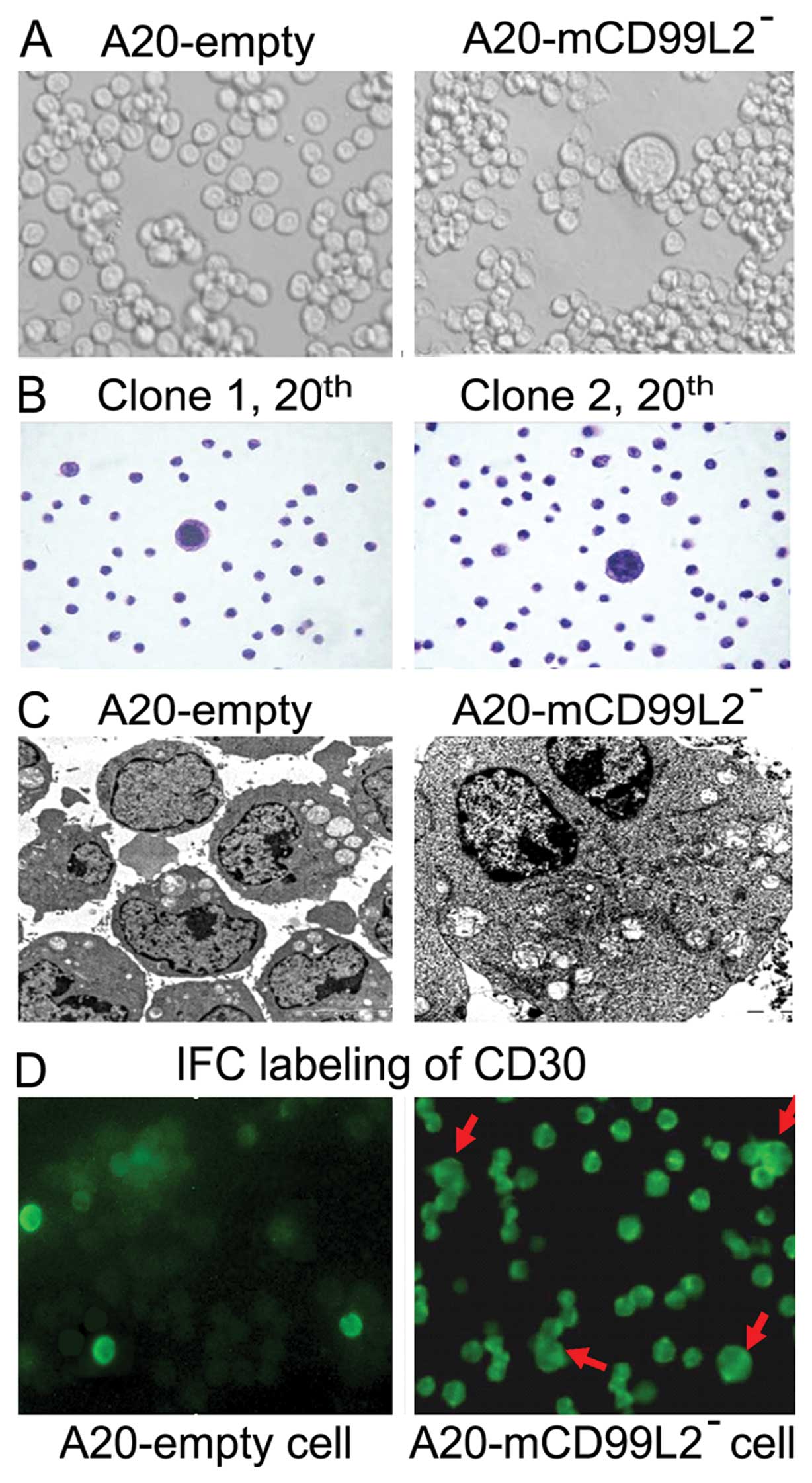
Morphological changes by downregulating
mCD99L2 expression in vitro
Morphology was extremely significant in the present
study. Some cells in the A20-mCD99L2− group demonstrated
morphological changes including larger volume, abundant cytoplasm,
marked pleomorphism, large and deeply stained nucleus, while some
were binucleated or polynucleated (Fig.
2A). Even when cells were continuously cultured to the 20th
passages (Fig. 2B), these changes
in various clones of the A20-mCD99L2− group remained.
TEM observation indicated that controls were naïve cells with less
cytoplasm and organelles and relatively larger nucleus, while the
giant cells in the A20-mCD99L2− group were much larger,
with abundant cytoplasm and organelles, particularly mitochondria
and endoplasmic reticulum (Fig.
2C). The unique morphology of binucleated or polynucleated
appearance with larger nucleus and nucleoli exhibited by some
A20-mCD99L2− cells partly mimic that of human H/RS
cells.
Biological characteristics of
A20-mCD99L2− cells
The growth of cultured A20-mCD99L2− cells
was investigated using the MTT method. Slower kinetics of cell
proliferation compared with the controls were observed in the
A20-mCD99L2 group (Table III)
(P<0.05), which indicated that mCD99L2 downregulation induces a
weaker proliferative ability.
| Table IIIProliferation of various cell groups
in vitro by MTT (n=6). |
Table III
Proliferation of various cell groups
in vitro by MTT (n=6).
| Day | A20 | A20-empty |
A20-mCD99L2− |
|---|
| 1 | 0.43±0.02 | 0.43±0.02 | 0.43±0.02 |
| 2 | 0.53±0.02 | 0.53±0.03 | 0.47±0.02a,b |
| 3 | 0.65±0.02 | 0.64±0.03 | 0.55±0.04a,b |
| 4 | 0.84±0.02 | 0.83±0.03 | 0.65±0.03a,b |
| 5 | 0.95±0.02 | 0.93±0.03 | 0.73±0.05a,b |
| 6 | 0.94±0.03 | 0.92±0.01 | 0.71±0.03a,b |
| 7 | 0.92±0.02 | 0.92±0.02 | 0.70±0.02a,b |
To evaluate the cell cycle distribution, the DNA
contents of asynchronous cultures of various cell groups were
measured. The S phase of each group showed no significant
difference (P>0.05, n=4), while the G2 phase was significantly
prolonged in the A20-mCD99L2− cells compared to the A20
and A20-empty cells (Table IV)
(P<0.05), which indicated that A20-mCD99L2− cells may
be defective in cytokinesis.
| Table IVCell cycle analysis of various cell
groups (n=4). |
Table IV
Cell cycle analysis of various cell
groups (n=4).
| Cell groups | G1 (%) | S (%) | G2 (%) |
|---|
| A20 | 42.60±4.23 | 54.05±4.86 | 3.33±1.31a |
| A20-empty | 41.63±3.93 | 55.03±4.10 | 3.32±1.30b |
|
A20-mCD99L2− | 35.90±3.13 | 53.55±5.50 | 10.58±4.97a,b |
Immunophenotypes of
A20-mCD99L2− cells in vitro
As H/RS cells are characterized by a high expression
of CD15 and CD30 (22) and previous
findings have demonstrated that upregulated CD99 markedly
downregulates the expression of CD30 and CD15 (19–21),
flow cytometry was applied to examine the changes in CD antigen
expression by downregulating mCD99L2. The results indicated that
when compared with the controls, A20-mCD99L2− cells
exhibited significantly higher CD30 and CD15 levels (Table V) (P<0.01, n=3) and moderately
decreased CD19 and CD20 levels (P<0.05, n=3). The enhanced
expression of CD30 was also confirmed by immunofluoresence (IFC)
labeling using mouse CD30 antibodies and the giant cells as well as
some transformed A20-mCD99L2− cells were
CD30+ (Fig. 2D).
| Table VAnalysis of the antigen expression of
each group using FACS (n=3). |
Table V
Analysis of the antigen expression of
each group using FACS (n=3).
| Group | CD19 | CD20 | CD30 | CD15 |
|---|
| A20 | 91.57±2.11 | 41.33±2.25 | 41.70±2.60 | 51.2±2.60 |
| A20-empty | 90.90±4.41 | 40.80±3.90 | 41.87±2.06 | 49.9±2.92 |
|
A20-mCD99L2− | 75.70±3.2a | 13.10±5.16a | 76.0±2.44a | 71.6±3.60a |
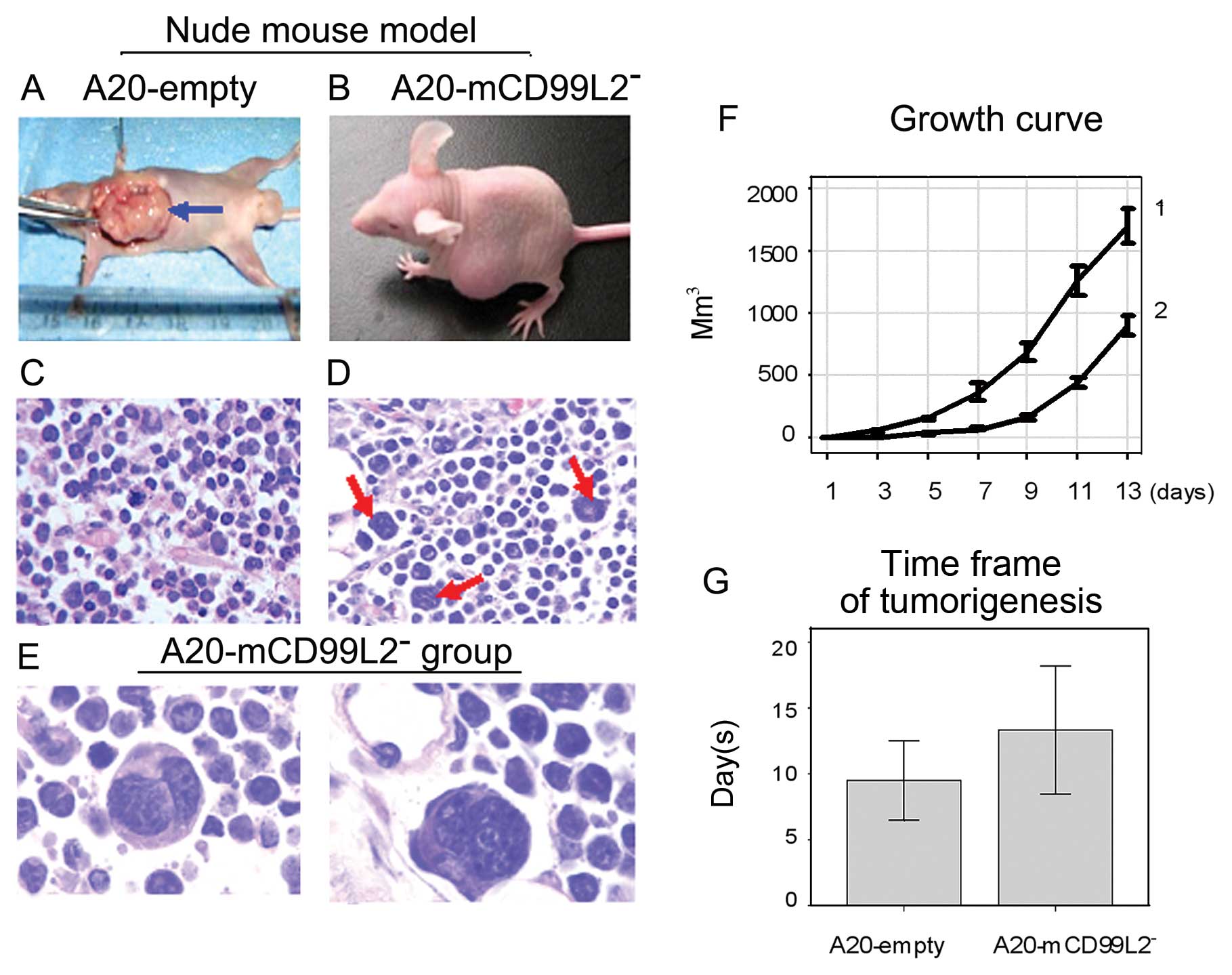
Histology, biology and phenotypes of
tumors in nude mice
Subcutaneous tumor models in nude mice were
successfully established (Table
VI, Fig. 3A–E). Tumor growth of
the A20-mCD99L2− group was much slower than the
A20-empty group (Fig. 3F). The time
frame of tumorigenesis in the A20-mCD99L2− group was
much longer compared with the control group (Fig. 3G), which indicated that the
proliferative ability of the A20-mCD99L2− cells was
weaker due to the downregulation of mCD99L2.
| Table VITumorigenesis in each mouse group
inoculated with various tumor cells. |
Table VI
Tumorigenesis in each mouse group
inoculated with various tumor cells.
| | Percentage of
tumorigenesis (%) (n) | Time frame of
tumorigenesis (days) |
|---|
| |
|
|
|---|
| Group | N (total) | A20-empty |
A20-mCD99L2− | A20-empty |
A20-mCD99L2− |
|---|
| A | 12 | 100 (6/6) | 100 (6/6) | 9.5±2.9 | 13.33±4.63 |
| B1 | 14 | 0 (0/7) | 0 (0/7) | - | - |
| B2 | 14 | 100 (7/7) | 14.3 (1/7) | 15.29±3.2 | 10 |
| C1 | 14 | 100 (7/7) | 14.3 (1/7) | 7.0±0.82 | 6 |
| C2 | 14 | 100 (7/7) | 0 (0/7) | 6.29±0.49 | - |
| D1 | 14 | 71.4 (5/7) | 0 (0/7) | 76.8±12.0 | >3 months |
| D2 | 14 | 100 (7/7) | 0 (0/7) | 26.1±7.9 | >3 months |
Histologically, the A20-empty tumors were
characterized by a diffuse homogeneous infiltrate consisting of
large and cohesive tumor cells with moderate cytoplasm and
pleomorphic nuclei (Fig. 3C), while
the A20-mCD99L2− tumor cells showed marked pleomorphism,
a diffuse distribution pattern, and some had two or more large and
deeply stained nuclei (Fig. 3D and
E).
The immunophenotypes of the primary tumor cells
dissociated from the xenotransplanted tumors were examined using
flow cytometry. CD30 expression was significantly upregulated,
whereas CD19 expression was lower in the A20-mCD99L2−
compared with the A20-empty group.
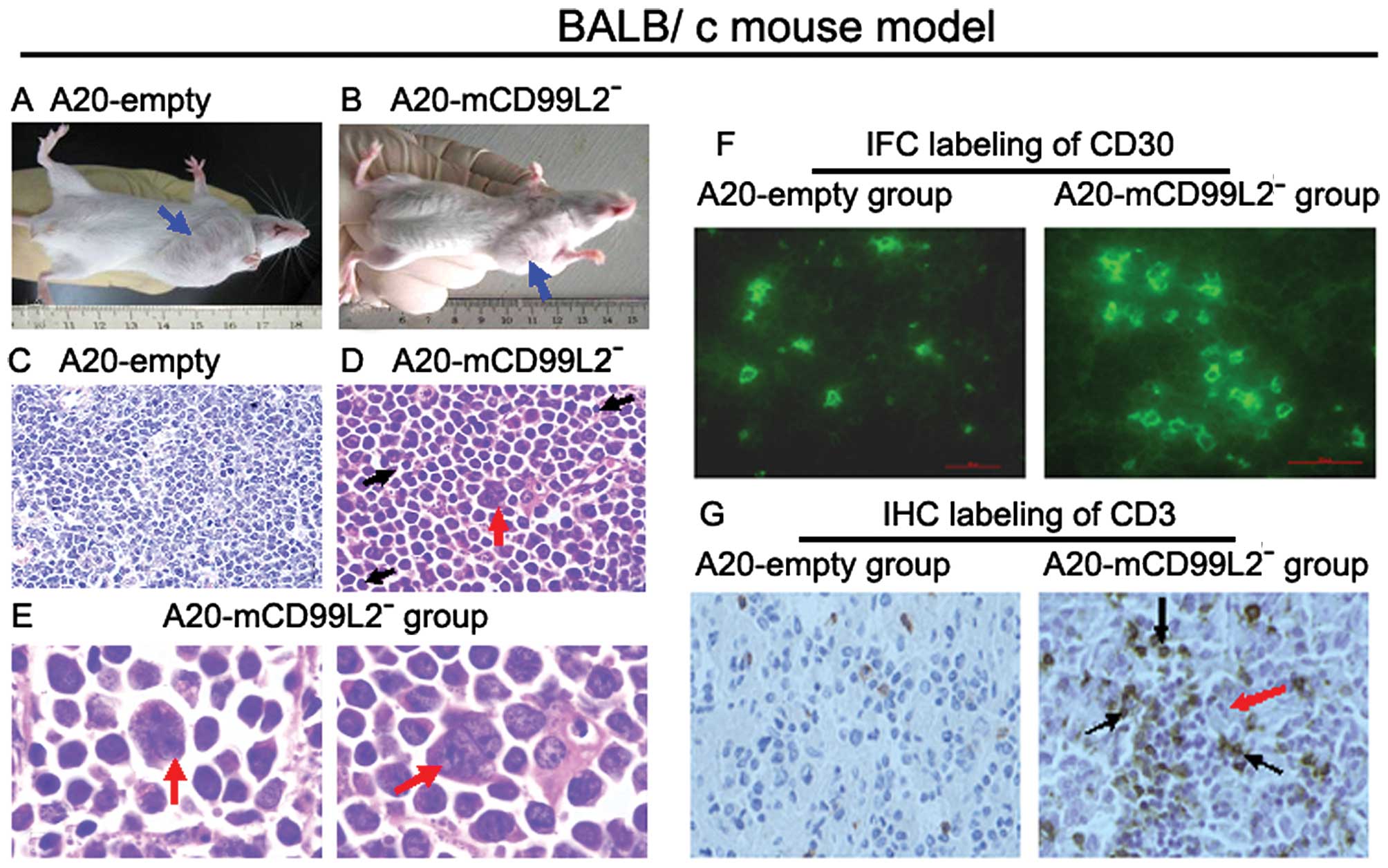
Effect of downregulation of mCD99L2 in
BALB/c mice
Inoculating BALB/c mice with A20-mCD99L2−
cells and A20-empty cells was conducted using various methods
(Table II, Fig. 4A and B). It was difficult for the
A20-mCD99L2− cells to form tumors in BALB/c mice
irrespective of the methods applied (Table VI), indicating that the
proliferative ability of the A20 cells with decreased mCD99L2
expression was significantly impaired in the immunocompetent BALB/c
mice.
The histological characteristics of the A20-empty
cell-induced tumor tissues demonstrated uniform B lymphoid cells,
which were most consistent with those in human diffuse large B cell
lymphoma (DLBCL) (Fig. 4C). Cells
in A20-mCD99L2− tumor tissues exhibited marked
pleomorphism and large and deeply stained nuclei, some were
binucleated or polynucleated (Fig. 4D
and E, red arrow). In addition, some lymphocytes were observed
in the tumor tissues (Fig. 4D,
black arrow).
CD30 was positive in A20-mCD99L2−
cell-induced tumors as observed using IFC labeling of mouse CD30,
particularly in the giant cells (Fig.
4F). As in Hodgkin’s lymphoma, H/RS cells were accompanied by a
number of background cells, including T lymphocytes.
Immunohistochemistry was applied to detect the expression of
CD3+ T lymphocytes in tumor tissues. The results
indicated that more CD3+ lymphocytes infiltrated into
the A20-mCD99L2− cell-induced tumor tissues compared
with the control group (Fig. 4G),
which suggested that various immune reactions were induced in
vivo.
Differential cytokine profile of cells
and tissues
The weak proliferative ability and pathological
features exhibited in the A20-mCD99L2− groups, which
partly mimicked those of Hodgkin’s lymphoma, encouraged us to
investigate the differential expression of cytokines/chemokines
(Fig. 5A and B) induced by the
downregulation of mCD99L2, as several cytokines are involved in the
cross-talk between H/RS cells and the background inflammatory
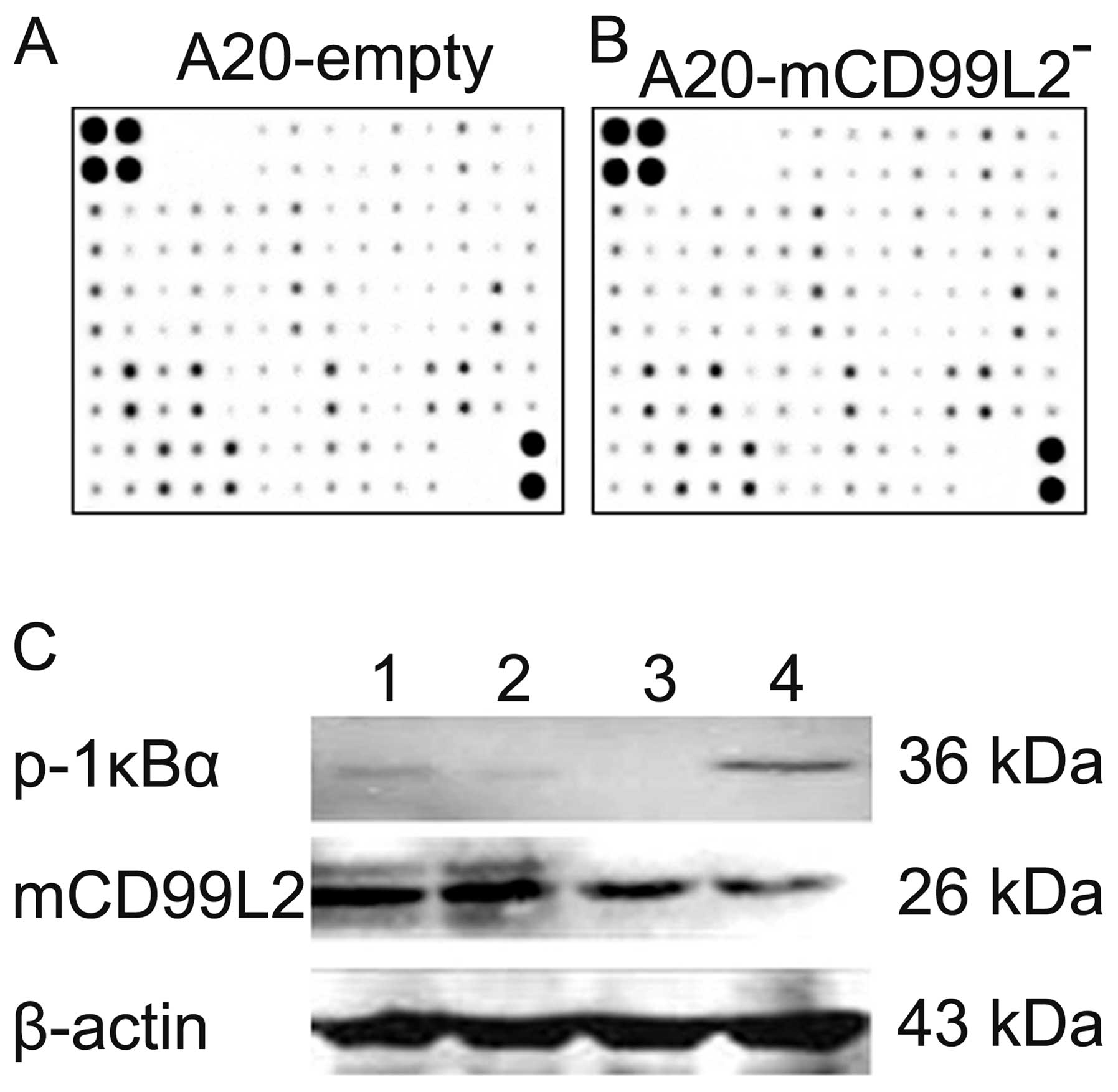
environment. Cytokine protein arrays showed that several cytokines
including CD30T, IL-12p40/p70, IL-3, IFN-γ, CXCL16, MIP-1α
and CD40, were upregulated (≥1.5-fold) in the
A20-mCD99L2− cells and no cytokine was downregulated in
excess of 1.5-fold (Table VII).
The changes in morphology, biology and phenotypes may be directly
or indirectly associated with these differentially expressed
cytokines/chemokines.
| Table VIIUpregulated cytokines in excess of
1.5-fold in A20-mCD99L2− cells compared with A20-empty
cells. |
Table VII
Upregulated cytokines in excess of
1.5-fold in A20-mCD99L2− cells compared with A20-empty
cells.
| Row | Col | Col | Name | 1 Primary | 2 Primary | 1 Standard | 2 Standard | No. 2/1 |
|---|
| 1, 2 | 9 | i | CD30T | 7199 | 12042 | 0.0564 | 0.164 | 2.914 |
| 1, 2 | 10 | j | CD40 | 12714 | 16692 | 0.173 | 0.261 | 1.519 |
| 1, 2 | 13 | m | CXCL16 | 11067.5 | 14359.5 | 0.137 | 0.212 | 1.548 |
| 3, 4 | 6 | f | IFN-γ | 11977.5 | 15866.5 | 0.156 | 0.243 | 1.5598 |
| 3, 4 | 13 | m | IL-3 | 8380.5 | 10564 | 0.081 | 0.134 | 1.650 |
| 5, 6 | 6 | f | IL-12 p40/p70 | 9328.5 | 12161 | 0.101 | 0.1677 | 1.654 |
| 7, 8 | 6 | f | MIP-1α | 10985 | 14135.5 | 0.135 | 0.208 | 1.533 |
mCD99L2 downregulation correlates with
the activated nuclear factor-κB (NF-κB) pathway
As regards the mecharnism involved, the NF-κB
pathway plays a crucial role in the pathology of Hodgkin’s lymphoma
and is a significant factor affecting cytokines/chemokines. NF-κB
is retained in the cytoplasm of inactivated cells through the
interaction with members of the IκB inhibitor family, including
IκBα. The phosphorylation and subsequent degradation of IκB lead to
the release of NF-κB, allowing it to translocate to the nucleus and
activate transcription. The level of p-IκBα, which could represent
the activation of NF-κB pathway, was examined in various cell
groups using western blot analysis (Fig. 5C).
The results indicated that the expression of the
mCD99L2 protein in the A20-mCD99L2− cells (Fig. 5C, lane 4) was weaker compared with
that in the A20 and A20-empty cells (Fig. 5C, lanes 1 and 2), which was an
evidence of the effect of shRNA targeting mCD99L2. The expression
of p-IκBα protein was stronger in the A20-mCD99L2− cells
(Fig. 5C, lane 4) compared with
that in the A20 and A20-empty cells (Fig. 5C, lanes 1 and 2), suggesting that
NF-κB activity was elevated in the A20-mCD99L2−
cells.
When the cells were treated with BAY, an inhibitor
of the NF-κB signaling pathway, p-IκBα protein expression was
significantly decreased in the A20-mCD99L2− cells, while
mCD99L2 protein expression was not affected (Fig. 5C, lane 3), which suggests a
potential correlation between the downregulation of mCD99L2 and the
NF-κB pathway.
Discussion
Effect of shRNA targeting mCD99L2 on B
cell lymphoma
Previous reports have indicated that the
overexpression of the full-length CD99 isoform (CD99wt), one of the
two distinct proteins produced by the alternative splicing of the
CD99 gene transcript, dramatically inhibits cancer cell
proliferation, migration and metastasis, whereas the overexpression
of the short CD99 isoform (CD99sh) remarkably favors these
phenomena (23), while the effects
of mCD99L2 on tumor cells have not yet been reported. Our
previous study confirmed that mCD99L2 is expressed in the
A20 cell line (18). In the present
study, the effective shRNA sequence targeting mCD99L2 was selected
and identified, the mCD99L2-downregulated A20 subclones
(A20-mCD99L2− cell) were established and the
morphological, biological and phenotypic characteristics of the
A20-mCD99L2− cells were investigated in vitro and
in vivo for the first time.
Our results strongly suggest that suppressing
mCD99L2 may impair the proliferative ability of murine B cell
lymphoma. As the A20 cell-induced tumors were pathologically
described as DLBCL, which is a type of lymphoma with poor
prognosis, our investigation on mCD99L2 may provide a potential
target of CD99-related antigens for the clinical therapy of B cell
lymphoma.
Downregulation of mCD99L2 leads to the
transformation of some A20 cells into H/RS-like cells
Using CD99-deficient IM9 and BJAB B cell lines,
investigators have confirmed that the downregulation of CD99 is a
primary requirement for the generation of H/RS cells (19,20). A
previous study of ours suggested that the CD99-upregulated H/RS
cell line (L428) lost its nature as an H/RS cell line (21). Although a functional similarity
between CD99 and mCD99L2 in lymphoma cells has yet to be confirmed,
unique morphological changes were observed in the transformed
A20-mCD99L2− cells; some giant cells similar to human
H/RS cells were observed in the cultured cells, as well as in nude
and BALB/c mice, which suggests that the downregulation of mCD99L2
led to the transformation of some A20 cells into H/RS-like
cells.
Biologically, the suppressive effect of the
downregulation of mCD99L2 on the proliferative ability of
mouse B lymphoma cells was observed in the cultured cells and tumor
tissues. Although the A20-mCD99L2− cells grew at a
slower rate compared with the A20 cells, cell cycle analysis showed
that the S phase of each group did not vary significantly. However,
the G2 phase of the A20-mCD99L2− cells lasted for a
longer period of time. The fact that some A20-mCD99L2−
cells stay in the G2 phase and exhibit difficulty in entering the M
phase may be due to the fact that giant cells with two or more
nuclei were induced in the transformed A20-mCD99L2−
cells. The weaker proliferative ability and prolonged G2 phase
imply that A20-mCD99L2− cells are similar to H/RS cells
to a certain extent; thus defects were noted in the cell cycle
regulation as one of several anti-apoptotic mechanisms (24,25).
The vast majority of classical Hodgkin’s lymphomas
are thought to arise from transformed germinal center B cells due
to the loss of B cell characteristics during antigen selection
(26). The H/RS cells possess
unique morphological and biological features and phenotypic
characteristics, thus various phenotypic changes were detected in
these cells. The significantly increased CD30 and CD15 expression
and moderately decreased CD19 and CD20 expression indicate that the
A20-mCD99L2− cells tend to lose part of their B cell
characteristics and gain some phenotypic features of H/RS cells,
which are characterized by the high expression of CD15 and CD30
(22). Moreover, in the BALB/c
mouse tumor tissues of the A20-mCD99L2− group, typical
H/RS-like cells were detected, the CD30 antigen was highly
expressed and more CD3+ T lymphocytes were observed.
Our results in vitro and in vivo
encouraged us to investigate the cytokine expression in various
cell groups for the interaction of cytokines/chemokines, which may
lead to an environment in which H/RS cells are able to proliferate,
escape from apoptosis and survive host antitumor defense (27–29).
The results show that several cytokines, such as CD30T,
IL-12p40/p70, IL-3, IFN-γ, CXCL16, MIP-1α and CD40 were
upregulated, a number of which have been associated with Hodgkin’s
lymphomas or H/RS cells in previous reports (30,31).
For instance, compared with other types of lymphoma, the expression
of CD30 and CD40 in Hodgkin’s lymphoma is highly expressed
(32). IL-3 is overexpressed in
H/RS cell lines (30). CXCL16 has
also been reported to be expressed in some H/RS cell lines
(33). Thus, the cytokine
expression profile in the A20-mCD99L2− cells exhibited
some similarities to that of H/RS cells. Combining our in
vitro and in vivo observation, A20-mCD99L2−
cells partly mimic the characteristics of human H/RS cells.
Effect of shRNA targeting mCD99L2 may
involve NF-κB pathway
The A20-mCD99L2− cells were observed to
have difficulty in forming tumors in mice with normal immune
functions. More CD3+ lymphocytes infiltrated into the
A20-mCD99L2− cell-induced tumor tissues compared with
the control group in the BALB/c mice, which suggested that certain
immune reactions were induced by certain cell groups. Of the
upregulated cytokines in the A20-mCD99L2− cells in the
cytokine profile, IFN-γ, IL-12p40/p70 and MIP-1α have been reported
to be involved in regulating immune functions between tumor cells
and lymphocytes (34). CXCL16 has
also been reported to be involved in the tumor anti-host reactions
(35). Although details are yet to
be elucidated, the differentially expressed cytokines may elucidate
some of the mechanisms involved in the effect of the downregulation
of mCD99L2 in A20 cells and may provide clues for further
study.
As regards the pathways, the constitutive NF-κB
activation is a striking feature and the major pathogenetic
mechanism in H/RS cells (36–38);
therefore, we focused on the NF-κB pathway. Our finding suggesting
that p-IκBα, an indicator of the activation of the NF-κB pathway,
was enhanced in the A20-mCD99L2− cells, suggested the
elevated NF-κB activity by the downregulation of mCD99L2. Treatment
with BAY significantly decreased the p-IκBα level in
A20-mCD99L2− cells without affecting mCD99L2 protein
expression, suggesting a potential regulatory role of mCD99L2 in
the NF-κB pathway. Although the downregulation of mCD99L2 may
trigger various pathways to regulate several cytokines, our results
suggested that the downregulation of mCD99L2 in the A20 cells may
correlated with the activated NF-κB pathway, which may partly
contribute to the morphological, biological and phenotypic changes
induced by shRNA targeting mCD99L2 in A20 cells. The regulatory
mechanisms between mCD99L2 and NF-κB require extensive
investigation in the future.
In conclusion, the stable mCD99L2-downregulated A20
cell line was established and identified. The effect of shRNA
targeting mCD99L2 in vitro and in vivo was observed.
The downregulation of mCD99L2 led to the transformation of some A20
cells into H/RS-like cells, impaired the proliferative ability of
murine B cell lymphoma, changed the immunophenotypes, led to
differentially expressed cytokines and suggested the involvement of
the activated NF-κB pathway. Our study provides experimental data
for additional studies on the mCD99L2 gene and protein in
lymphomas.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant nos. 81071941, 81071659 and
81101537).
References
|
1
|
Levy R, Dilley J, Fox RI, et al: A human
thymusleukemia antigen defined by hybridoma monoclonal antibodies.
Proc Natl Acad Sci USA. 76:6552–6556. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bernard G, Raimondi V, Alberti I, et al:
CD99 (E2) up-regulates alpha4beta1-dependent T cell adhesion to
inflamed vascular endothelium under flow conditions. Eur J Immunol.
30:3061–3065. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schenkel AR, Mamdouh Z, Chen X, et al:
CD99 plays a major role in the migration of monocytes through
endothelial junctions. Nature Immunol. 3:143–150. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tato CM, Joyce-Shaikh B, Banerjee A, et
al: The myeloid receptor PILRβ mediates the balance of inflammatory
responses through regulation of IL-27 production. PLoS One.
7:e316802012.
|
|
5
|
Park HJ, Byun D, Lee AH, et al:
CD99-dependent expansion of myeloid-derived suppressor cells and
attenuation of graft-versus-host disease. Mol Cells. 33:259–267.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dworzak MN, Froschl G, Printz D, et al:
CD99 expression in T-lineage ALL: implications for flow cytometric
detection of minimal residual disease. Leukemia. 18:703–708. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Maitra A, Hansel DE, Argani P, et al:
Global expression analysis of well-differentiated pancreatic
endocrine neoplasms using oligonucleotide microarrays. Clin Cancer
Res. 9:5988–5995. 2003.
|
|
8
|
Kang LC and Dunphy CH: Immunoreactivity of
MIC2 (CD99) and terminal deoxynucleotidyl transferase in bone
marrow clot and core specimens of acute myeloid leukemias and
myelodysplastic syndromes. Arch Pathol Lab Med. 130:153–157.
2006.
|
|
9
|
Diwan AH, Skelton HG III, Horenstein MG,
et al: Dermatofibrosarcoma protuberans and giant cell fibroblastoma
exhibit CD99 positivity. J Cutan Pathol. 35:647–650. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ramsay AD, Bates AW, Williams S, et al:
Variable antigen expression in hepatoblastomas. Appl
Immunohistochem Mol Morphol. 16:140–147. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Suh YH, Shin YK, Kook MC, et al: Cloning,
genomic organization, alternative transcripts and expression
analysis of CD99L2, a novel paralog of human CD99, and
identification of evolutionary conserved motifs. Gene. 307:63–76.
2003. View Article : Google Scholar
|
|
12
|
Bixel MG, Petri B, Khandoga AG, et al: A
CD99-related antigen on endothelial cells mediates neutrophil but
not lymphocyte extravasation in vivo. Blood. 109:5327–5336. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bixel MG, Li H, Petri B, et al: CD99 and
CD99L2 act at the same site as, but independently of, PECAM-1
during leukocyte diapedesis. Blood. 116:1172–1184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim KJ, Kanellopoulos-Langevin C, Merwin
RM, et al: Establishment and characterization of BALB/c lymphoma
lines with B cell properties. J Immunol. 122:549–554.
1979.PubMed/NCBI
|
|
15
|
Passineau MJ, Siegal GP, Everts M, et al:
The natural history of a novel, systemic, disseminated model of
syngeneic mouse B-cell lymphoma. Leuk Lymphoma. 46:1627–1638.
2005.PubMed/NCBI
|
|
16
|
Warncke M, Buchner M, Thaller G, et al:
Control of the specificity of T cell-mediated anti-idiotype
immunity by natural regulatory T cells. Cancer Immunol Immunother.
60:49–60. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu F, Zhang G, Zhou XH, et al:
Immuno-characterization of mouse model similar to human diffuse
large B cell lymphoma. Zhongguo Shi Yan Xue Ye Xue Za Zhi.
18:655–659. 2010.(In Chinese).
|
|
18
|
Shen LJ, Fang WY, Xie SM, et al:
Expression and cloning of mCD99L2 gene from mouse B lymphoma cell
line A20 and construction of its eukaryotic expression vector. Nan
Fang Yi Ke Da Xue Xue Bao. 26:144–149. 2006.(In Chinese).
|
|
19
|
Kim SH, Choi EY, Shin YK, et al:
Generation of cells with Hodgkin’s and Reed-Sternberg phenotype
through downregulation of CD99 (Mic2). Blood. 92:4287–4295.
1998.
|
|
20
|
Kim SH, Shin YK, Lee IS, et al: Viral
latent membrane protein 1 (LMP-1)-induced CD99 down-regulation in B
cells leads to the generation of cells with Hodgkin’s and
Reed-Sternberg phenotype. Blood. 95:294–300. 2000.PubMed/NCBI
|
|
21
|
Huang X, Zhou X, Wang Z, et al: CD99
triggers upregulation of miR-9-modulated PRDM1/BLIMP1 in
Hodgkin/Reed-Sternberg cells and induces redifferentiation. Int J
Cancer. 131:E382–E394. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Burgess SC, Young JR, Baaten BJ, et al:
Marek’s disease is a natural model for lymphomas overexpressing
Hodgkin’s disease antigen (CD30). Proc Natl Acad Sci USA.
101:13879–13884. 2004.
|
|
23
|
Scotlandi K, Zuntini M, Manara MC, et al:
CD99 isoforms dictate opposite functions in tumour malignancy and
metastases by activating or repressing c-Src kinase activity.
Oncogene. 26:6604–6618. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Garcia JF, Camacho FI, Morente M, et al:
Hodgkin and Reed-Sternberg cells harbor alterations in the major
tumor suppressor pathways and cell-cycle checkpoints: analyses
using tissue microarrays. Blood. 101:681–689. 2003. View Article : Google Scholar
|
|
25
|
Sánchez-Aguilera A, Montalbán C, de la
Cueva P, et al: Tumor microenvironment and mitotic checkpoint are
key factors in the outcome of classic Hodgkin lymphoma. Blood.
108:662–668. 2006.PubMed/NCBI
|
|
26
|
Marafioti T, Hummel M, Foss HD, et al:
Hodgkin and Reed-Sternberg cells represent an expansion of a single
clone originating from a germinal center B-cell with functional
immunoglobulin gene rearrangements but defective immunoglobulin
transcription. Blood. 95:1443–1450. 2000.
|
|
27
|
Maggio E, van den Berg A, Diepstra A, et
al: Chemokines, cytokines and their receptors in Hodgkin’s lymphoma
cell lines and tissues. Ann Oncol. 13(Suppl 1): 52–56. 2002.
|
|
28
|
Skinnider BF and Mak TW: The role of
cytokines in classical Hodgkin lymphoma. Blood. 99:4283–4297. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Steidl C, Connors JM and Gascoyne RD:
Molecular pathogenesis of Hodgkin’s lymphoma: increasing evidence
of the importance of the microenvironment. J Clin Oncol.
29:1812–1826. 2011.
|
|
30
|
Aldinucci D, Olivo K, Lorenzon D, et al:
The role of interleukin-3 in classical Hodgkin’s disease. Leuk
Lymphoma. 46:303–311. 2005.
|
|
31
|
Niederkorn JY: Emerging concepts in CD8(+)
T regulatory cells. Curr Opin Immunol. 20:327–331. 2008.
|
|
32
|
Lee IS, Kim SH, Song HG, et al: The
molecular basis for the generation of Hodgkin and Reed-Sternberg
cells in Hodgkin’s lymphoma. Int J Hematol. 77:330–335. 2003.
|
|
33
|
Hanamoto H, Nakayama T, Miyazato H, et al:
Expression of CCL28 by Reed-Sternberg cells defines a major subtype
of classical Hodgkin’s disease with frequent infiltration of
eosinophils and/or plasma cells. Am J Pathol. 164:997–1006.
2004.PubMed/NCBI
|
|
34
|
Luft T, Luetjens P, Hochrein H, et al:
IFN-alpha enhances CD40 ligand-mediated activation of immature
monocyte-derived dendritic cells. Int Immunol. 14:367–380. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Meijer J, Ogink J, Kreike B, et al: The
chemokine receptor CXCR6 and its ligand CXCL16 are expressed in
carcinomas and inhibit proliferation. Cancer Res. 68:4701–4708.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Horie R, Watanabe T, Morishita Y, et al:
Ligand-independent signaling by overexpressed CD30 drives NF-kappaB
activation in Hodgkin-Reed-Sternberg cells. Oncogene. 21:2493–2503.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aldinucci D, Lorenzon D, Cattaruzza L,
Pinto A, Gloghini A, Carbone A and Colombatti A: Expression of CCR5
receptors on Reed-Sternberg cells and Hodgkin lymphoma cell lines:
involvement of CCL5/Rantes in tumor cell growth and
microenvironmental interactions. Int J Cancer. 122:769–776. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Schwarzer R and Jundt F: Notch and NF-κB
signaling pathways in the biology of classical Hodgkin lymphoma.
Curr Mol Med. 11:236–245. 2011.
|