Introduction
Gastric cancer, a malignant epithelial tumor, is one
of the most common cancers and is the second most common cause of
cancer-related mortality in the world (1,2).
Recently employed therapies for gastric cancer during the past few
years have included chemotherapy, surgery and radiation (3–5). It is
generally accepted that systemic chemotherapy is the main option.
Although these therapies have led to considerable improvements in
gastric cancer treatment, the occurrence of drug resistance, which
is associated with poor prognosis, remains a dire issue (6,7). Thus,
there is a pressing need for the development of effective drugs for
the treatment of gastric cancer.
The regulation of the cell cycle is a series of
events leading to the maintenance of cell proliferation in
multicellular organisms. Regulation consists of four distinct
sequential phases: the G1 phase, the S phase (synthesis), the G2
phase, and the M phase (mitosis) (8). The major factors involved in this
process are cyclins and cyclin-dependent kinases (CDKs), which
regulate the progression of the cell cycle (8–10).
Progression from the G1 to the S phase of the cell cycle
predominantly requires the activation of cyclin D1-CDK4 and cyclin
E-CDK2 (8). The kinase activity of
these cyclins/CDK complexes can be blocked by CDK-inhibitory
proteins, including p21WAF1 and p27KIP, which subsequently obstruct
G1 cell cycle progression (11). In
addition, previous results have suggested that activation of the
mitogen-activated protein kinase (MAPK) signaling transduction
pathway, such as extracellular signaling-regulated kinase 1/2
(ERK1/2), c-Jun N-terminal kinase (JNK) and p38 MAPK are known to
be involved in the inhibition of cell growth (12–14).
Accumulative evidence has shown that the p38MAPK signaling
transduction pathway may be associated with the process of cell
cycle arrest (13,14).
Many studies have found that MMPs, particularly
MMP-2 (gelatinase A, 72-kDa gelatinase) and MMP-9 (gelatinase B,
92-kDa gelatinase), promote the degradation of the extracellular
matrix (ECM), which is a process that is involved in tumor invasion
and metastasis (15,16). Previous studies have shown that
higher levels of MMP-9 are associated with metastatic tumors such
as gastric cancer (17–19). Previous reports have indicated that
TNF-α induces MMP-9 expression in cancer cells (20–23).
The expression of MMP-9 by TNF-α is mediated via activation of the
transcription factors NF-κB and AP-1 in several tumor cell types
(20–23).
Gleditsia sinensis has been used mainly in
Oriental countries for years as a traditional medicine for the
treatment of swelling, suppuration, carbuncle and skin diseases
(24). The main constituents of
Gleditsia sinensis include stigmasterol, ellagic acid
glycoside and lupine acid (25–27).
Gleditsia sinensis exhibits a number of biological
activities, including the promotion of anti-allergenic,
anti-inflammatory and antitumor effects (29,30).
Although noticeable progress has been made toward our understanding
of the mechanisms of the Gleditsia sinensis-induced
antitumor effect, the sequence of events leading to cell growth
inhibition in cancer cells treated with Gleditsia sinensis
thorns remains unclear. The present study demonstrated that the
antitumor effect of an extract of Gleditsia sinensis thorns
on the p38MAPK signaling pathway involved cell cycle modulation and
growth inhibition in human gastric cancer cells in vitro.
Moreover, we examined MMP-9 regulation in gastric cancer SNU-5
cells following treatment with Gleditsia sinensis
thorns.
Materials and methods
Materials
Polyclonal antibodies to cyclin E, CDK2 and CDK4
were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). Polyclonal antibodies to cyclin D1, p21WAF1, p53, p27KIP,
ERK1/2, phospho-ERK, p38 MAP kinase, phospho-p38 MAP kinase, JNK
and phospho-JNK were obtained from New England Biolabs (Beverly,
MA, USA). U0126 and SB203580 were obtained from Calbiochem (San
Diego, CA, USA). A polyclonal antibody to MMP-9 was obtained from
Chemicon (Temecula, CA, USA).
Preparation of the extract
Air-dried and crushed Gleditsia sinensis
thorns (100 g) were added to ethanol, and extraction was performed
by heating at 100°C. The extract was then concentrated with a
rotary evaporator and lyophilized. The final extract weighed 10 g
(a collection rate of 10%), and was diluted with saline
solution.
Cell cultures
The human SNU-5 gastric cancer cell line was
obtained from the American Type Culture Collection (Manassas, VA,
USA). The cells were maintained in DMEM (4.5 g glucose/liter)
supplemented with 10% fetal calf serum, L-glutamine, and
antibiotics (Biological Industries, Beit Haemek, Israel) at 37°C in
a 5% CO2 humidified incubator.
Cell viability assay
Subconfluent, exponentially growing SNU-5 cells in
96-well plates, were incubated with the ethanol extract of
Gleditsia sinensis thorns (EEGS) for various periods of
time. Cell viability was determined using a modification of a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay, which was based on the conversion of the tetrazolium salt
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2-tetrazolium
to a formazan product by mitochondrial dehydrogenase (31). The formazan product was quantified
by measuring the absorbance at 490 nm.
Apoptosis detection by ELISA
Detection of cell apoptosis was based on the
quantification of the enrichment of mono- and oligo-nucleosomes in
the cytoplasm using a Cell Death Detection ELISA kit (Roche,
Mannheim, Germany). Briefly, after treatment of cells with EEGS,
the cells were lysed and centrifuged. The supernatant containing
the cytoplasmic histone-associated DNA fragments was transferred to
a microplate coated with streptavidin, and was then reacted with a
mixture of the anti-histone antibodies labeled with biotin and
anti-DNA antibodies coupled with peroxidase. Peroxidase was
thereafter added as a substrate, and the development of the color
was read photometrically at 405 nm with 490 nm as the background.
The specific enrichment of mono- and oligo-nucleosomes released
into the cytoplasm was expressed as an enrichment factor compared
with the control.
Cell cycle analysis (FACS)
Cells were harvested, fixed in 70% ethanol, and
stored at −20°C. Cells then were washed twice with ice-cold PBS and
incubated with RNase and a DNA intercalating dye, propidium iodide.
Cell cycle phase analysis was performed using a Becton Dickinson
FACStar flow cytometer equipped with Becton Dickinson Cell Fit
software.
Immunoprecipitation and
immunoblotting
Growth-arrested cells were treated with EEGS in the
presence of 10% FBS for various time periods at 37°C. Cell lysates
were prepared, and immunoprecipitation and immunoblotting were
performed as previously described (31,36).
Zymography
The conditioned medium was electrophoresed in a
polyacrylamide gel containing gelatin at a concentration of 1
mg/ml. The gel was washed at room temperature for 2 h with 2.5%
Triton X-100 and then at 37°C overnight in a buffer containing 10
mM CaCl2, 150 mM NaCl and 50 mM Tris-HCl (pH 7.5). The
gel was stained with 0.2% Coomassie blue and photographed on a
light box. Proteolysis was detected as a white zone in a dark blue
field.
Nuclear extracts and electrophoretic
mobility shift assay
Nuclear extracts were essentially prepared as
described elsewhere (31,36). Cultured cells were collected by
centrifugation, washed and suspended in a buffer containing 10 mM
HEPES (pH 7.9), 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM DTT and
0.5 mM PMSF. After 15 min on ice, the cells were vortexed in the
presence of 0.5% Nonidet NP-40. The nuclear pellet was then
collected by centrifugation for 15 min at 4°C and extracted in a
buffer containing 20 mM HEPES (pH 7.9), 0.4 M NaCl, 1 mM EDTA, 1 mM
EGTA, 1 mM DTT and 1 mM PMSF.
The nuclear extract (10–20 μg) was preincubated at
4°C for 30 min with a 100-fold excess of an unlabeled
oligonucleotide spanning the -79 MMP-9 cis element of
interest. The sequences were: AP-1, CTGACCCCTGAGTCAGCACTT; NF-κB,
CAG TGGAATTCCCCAGCC; Sp-1, GCCCATTCCTTCCGCC CCCAGATGAAGCAG. The
reaction mixture was then incubated at 4°C for 20 min in a buffer
(25 mM HEPES buffer pH 7.9, 0.5 mM EDTA, 0.5 mM DTT, 0.05 M NaCl
and 2.5% glycerol) with 2 μg of poly dI/dC and 5 fmol
(2×104 cpm) of a Klenow end-labeled (32P-ATP)
30-mer oligonucleotide, spanning the DNA-binding site of the MMP-9
promoter. The reaction mixture was electrophoresed at 4°C in a 6%
polyacrylamide gel using a TBE (89 mM Tris, 89 mM boric acid and 1
mM EDTA) running buffer. The gel was rinsed with water, dried and
exposed overnight to X-ray film.
Statistical analysis
Where appropriate, data were expressed as the means
± SE. Data were analyzed using factorial ANOVA and a Fisher's least
significant differences test where appropriate. Statistical
significance was set at P<0.05.
Results
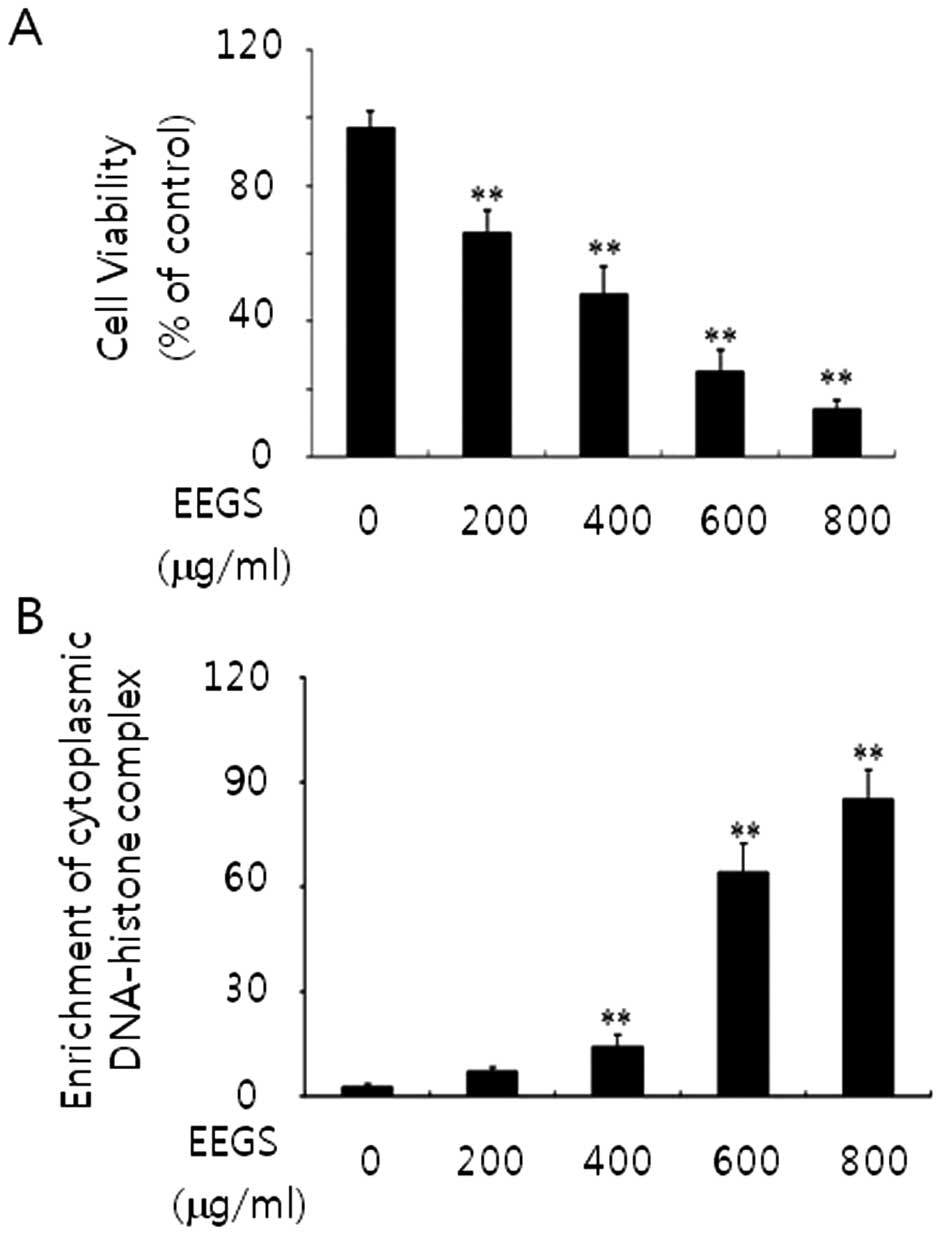
EEGS treatment reduces the proliferation
of human SNU-5 gastric cancer cells
To assess the effect of EEGS on cell proliferation
and cell death, SNU-5 cells were treated with 200, 400, 600 and 800
μg/ml doses of EEGS for 24 h. Cells treated with EEGS demonstrated
a concentration-dependent inhibition of cell growth (Fig. 1A). In addition, as shown in Fig. 1B, using an ELISA-based assay, we
observed an increase in cytoplasmic DNA-histone complexes (>600
μg/ml), which is associated with apoptosis, in the EEGS-treated
cells. Cells treated with the vehicle (ethanol) showed no changes
in the basal levels of cell growth and cell death (data not shown).
These data suggest a strong growth inhibitory effect and an
apoptotic effect in EEGS-treated SNU-5 cells.
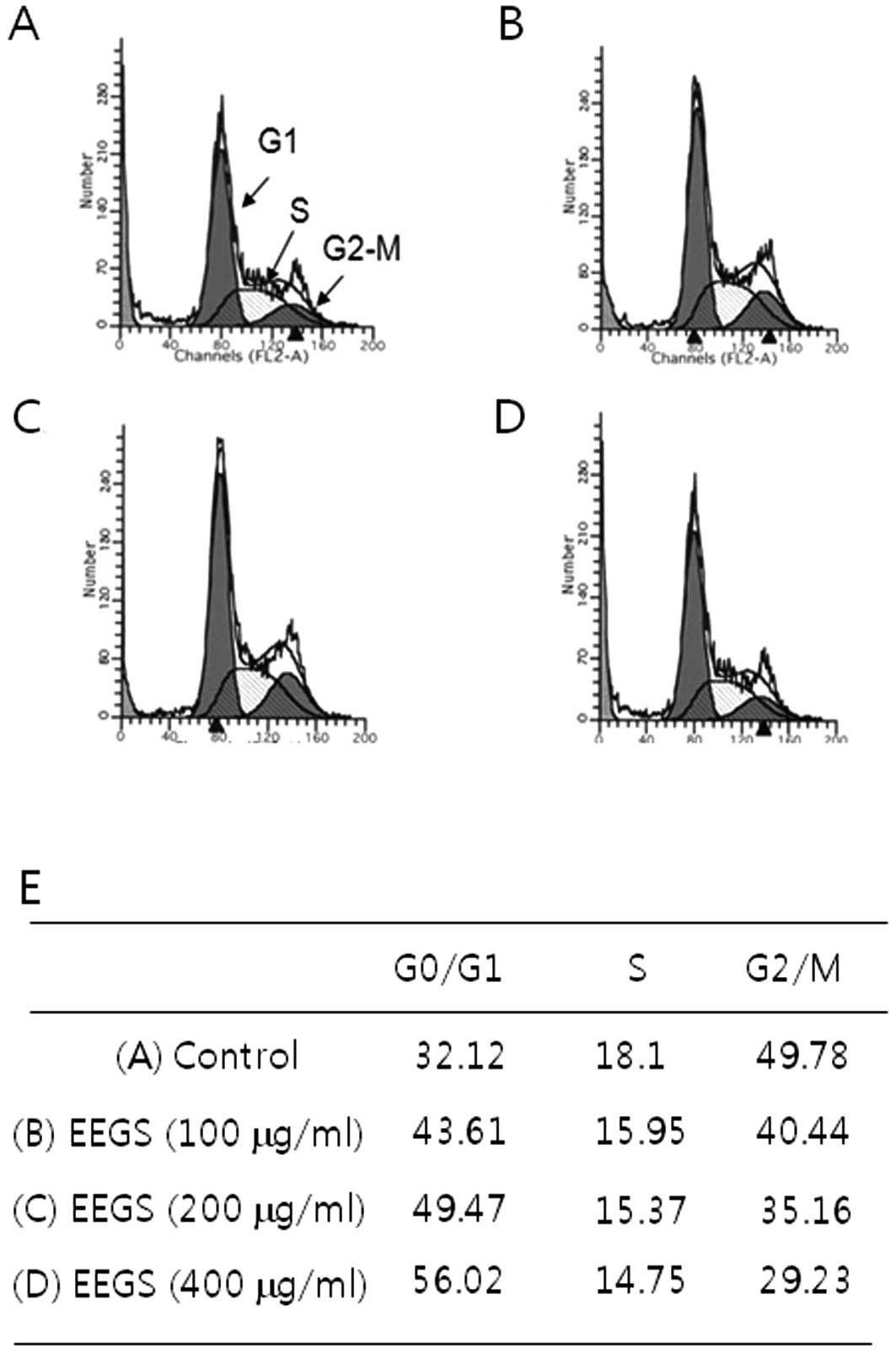
EEGS treatment induces G1 phase cell
cycle arrest
Since EEGS treatment resulted in a strong inhibitory
effect on cell growth of SNU-5 cells, we analyzed the cell cycle
distribution using flow cytometry. SNU-5 cells exhibited an
accumulation of the DNA content characteristic of the G1 phase cell
cycle following treatment with EEGS (400 μg/ml), based on a
comparison with the control (Fig.
2). To investigate the mechanism controlling the G1 phase of
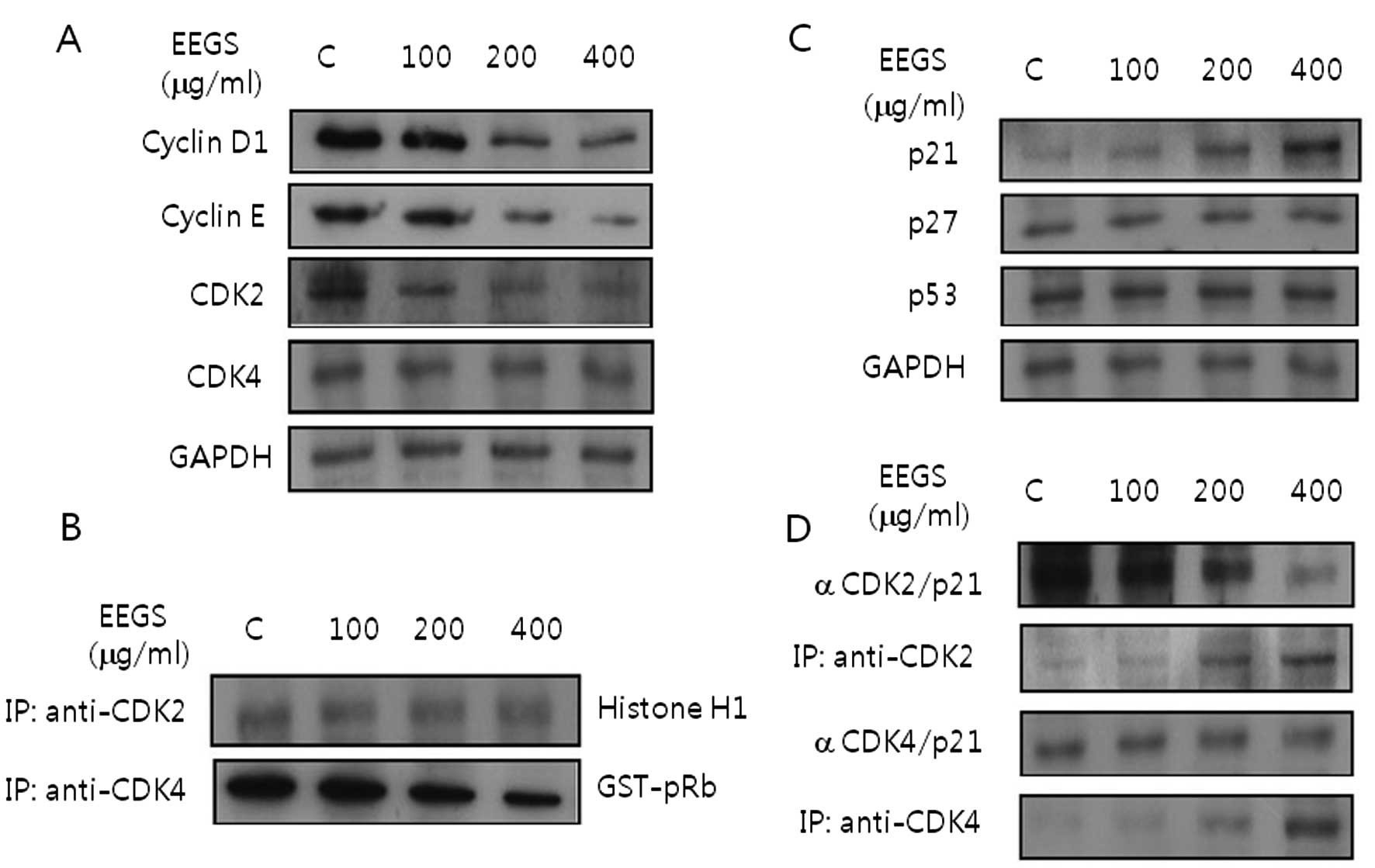
the cell cycle, we further examined the effects of EEGS treatment
on the levels of cyclins and CDKs, which are associated with the G1
phase of the cell cycle. EEGS treatment for 24 h resulted in
complete inhibition of the expression of cyclin D1 and cyclin E, as
well as a decrease in CDK2 and CDK4 proteins (Fig. 3A). Cell lysates were next examined
for kinase activity of CDK2 and CDK4 immunoprecipitates in the
EEGS-treated cells. EEGS treatment inhibited CDK2- and
CDK4-associated kinase activities in the SNU-5 cells in a
dose-dependent manner (Fig.
3B).
p21WAF1 is associated with EEGS-induced
G1 phase cell cycle arrest
Cyclin-dependent kinase inhibitors (CDKIs) are
negative regulatory proteins that bind to CDK/cyclin complexes and
inhibit kinase activities (11).
Based on our results demonstrating the cell cycle arrest effect of
EEGS, immunoblotting was performed to determine whether EEGS
modulates the expression levels of CDKIs. Our results indicated
that treatment of SNU-5 cells with EEGS for 24 h induced the
expression levels of p21WAF1 in a dose-dependent manner compared
with untreated cells (Fig. 3C).
However, EEGS treatment resulted in no noticeable change in the
induction of p27KIP1 and p53 tumor-suppressor proteins in the SNU-5
cells (Fig. 3C). These results
clearly showed that p21WAF1 is involved in EEGS-induced G1 phase
cell cycle arrest. Next, we determined the effect of EEGS on the
interaction between p21WAF1 and CDKs. The cell lysates from control
and EEGS-treated cells were immunoprecipitated using anti-CDK2 or
anti-CDK4 antibody, respectively. In addition, the immune complex
was analyzed for the presence of p21WAF1 by immunoblotting. As
shown in Fig. 3D, EEGS increased
the association of CDK2 with p21WAF1. In addition, the interaction
of p21WAF1/CDK4 complexes was maintained at high levels in SNU-5
cells 24 h after EEGS treatment (Fig.
3D). These results suggest that the increased association
between p21WAF1 and CDKs plays an important role in inhibiting CDK
kinase activity, accompanied by G1 phase cell cycle arrest
following EEGS treatment in SNU-5 cells.
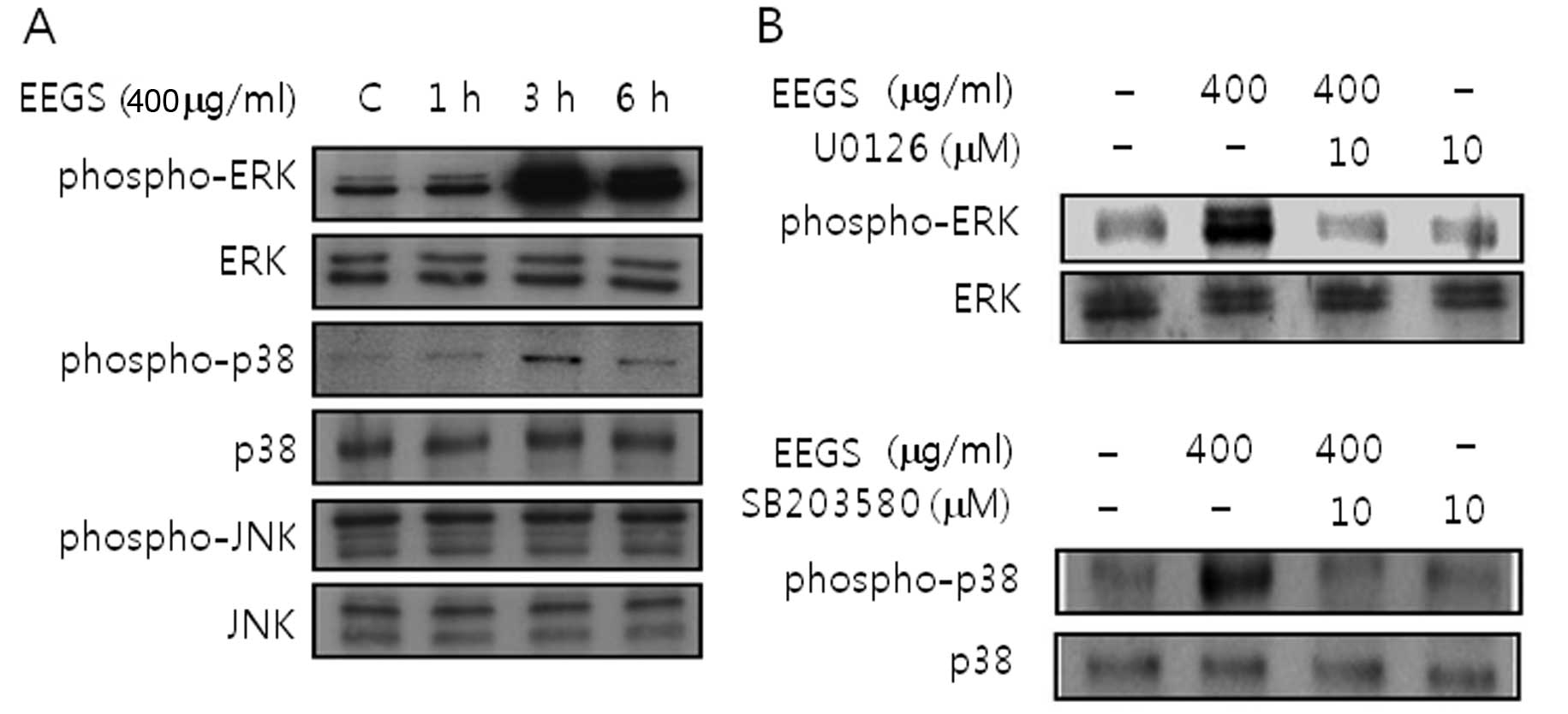
EEGS treatment activates ERK1/2 and p38
MAP kinase in SNU-5 cells
To examine whether MAPK signaling pathways are
involved in the inhibition of cell growth induced by EEGS,
immunoblotting was carried out. EEGS treatment induced activation
of ERK1/2 and p38 MAP kinase (Fig.
4A). In addition, the activation of ERK1/2 and p38 MAP kinase
was inhibited by the presence of specific kinase inhibitors such as
U0126 (ERK1/2) and SB203580 (p38 MAP kinase), respectively
(Fig. 4B). However, EEGS had no
effect on JNK activation (Fig. 4A).
These results suggest that EEGS treatment could be used to activate
the ERK1/2 and p38 MAP kinase signaling pathways in SNU-5
cells.
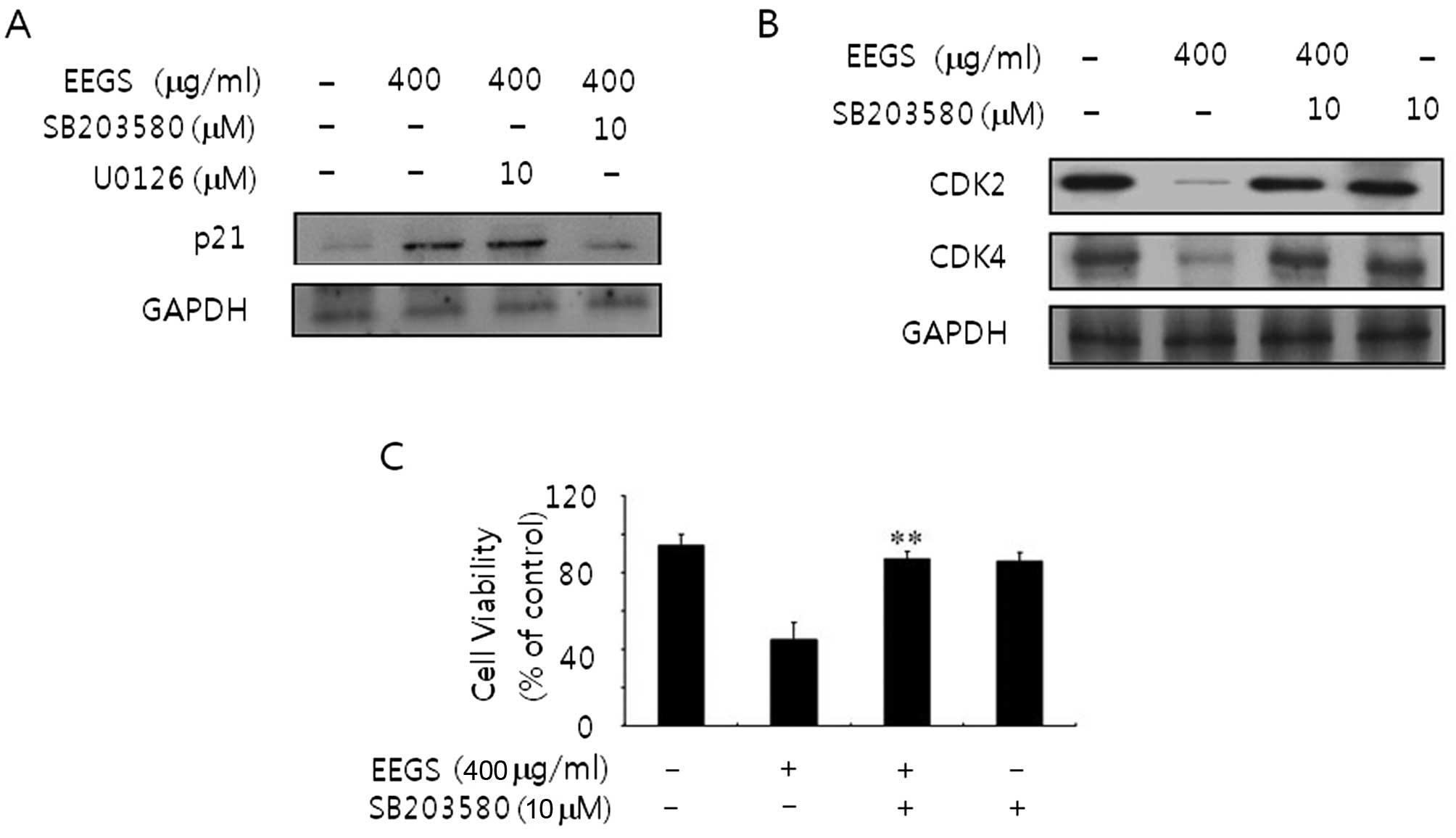
SB203580, a specific inhibitor of p38 MAP
kinase, reverses increased p21WAF1 expression and decreased CDK
levels in EEGS-treated SNU-5 cells
To confirm whether MAPK is associated with
EEGS-induced G1 phase cell cycle arrest, we next examined the
p21WAF1 expression and CDK levels using immunoblotting following
pretreatment with MAP kinase-specific kinase inhibitors. As shown
in Fig. 5A, the p21WAF1 expression
induced by EEGS was inhibited in the presence of SB203580. However,
U0126 treatment had no effect on EEGS-induced p21WAF1 expression
(Fig. 5A). In addition, the
decreased expression of CDK2 and CDK4 at the protein levels was
also reversed by pretreatment with SB203580 (Fig. 5B). These results indicate that the
p38 MAP kinase signaling pathway is involved in G1 phase cell cycle
arrest via expression of p21WAF1.
Inhibition of ERK recovers the inhibitory
growth effects in EEGS-treated SNU-5 cells
To further investigate the potential role of p38 MAP
kinase on the EEGS-induced inhibition of cell growth, an MTT assay
was performed following pretreatment with SB203580. To accomplish
this experiment, SNU-5 cells were untreated or treated with EEGS in
the absence or presence, respectively, of SB203580. Incubation of
cells with SB203580 blocked the decrease in cell growth in
EEGS-treated SNU-5 cells (Fig. 5C),
as compared with that in cells treated with EEGS alone. Our results
suggest that the p38 MAP kinase signaling pathway in SNU-5 cells is
associated with the cell growth inhibition that is induced by
EEGS.
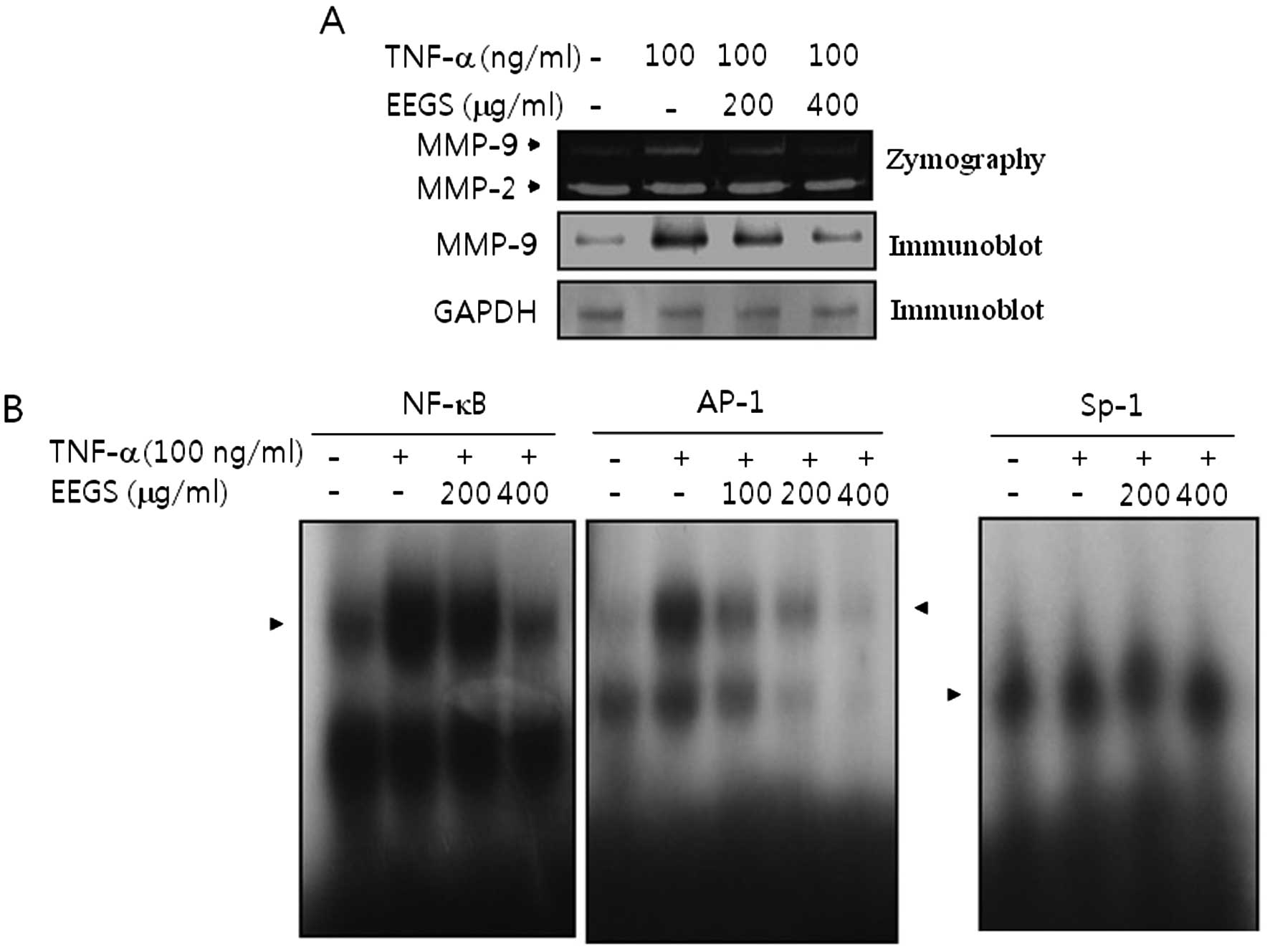
EEGS abolishes TNF-α-induced MMP-9
expression via a reduction in NF-κB and AP-1 binding activity
To determine the effect of EEGS on MMP-9 expression
in TNF-α-treated SNU-5 cells, gelatin zymography assay was
performed. The results showed an increase in MMP-9 expression
following treatment with TNF-α (Fig.
6A). TNF-α-induced MMP-9 secretion was inhibited by
pre-incubation of SNU-5 cells with EEGS (Fig. 6A). By contrast, there was no effect
on constitutive MMP-2 expression in the presence of either TNF-α or
EEGS (Fig. 6A). Similar results
were observed in the immunoblot analysis (Fig. 6A). Next, gel-shift assays were
performed to identify the potential transcription factors by which
TNF-α regulates MMP-9 expression. As shown in Fig. 6B, TNF-α increased the binding for
both NF-κB and AP-1 motifs in the SNU-5 cells. However, Sp-1
binding activity was not stimulated in response to TNF-α (Fig. 6B). Finally, we investigated the
possible implications of transcription factors NF-κB and AP-1 in
the regulation of MMP-9 in response to TNF-α by EEGS. As shown in
Fig. 6B, both NF-κB and AP-1 DNA
binding activities were almost abolished by pretreatment with EEGS
in TNF-α-treated SNU-5 cells. These results suggest that EEGS
inhibited TNF-α-induced MMP-9 expression via a decrease in the
activation of NF-κB and AP-1 motifs in SNU-5 cells.
Discussion
Many types of antitumor agents have been effective
against tumor initiation and the promotion of tumorigenesis
(3–5). However, crucial problems, such as
side-effects and the occurrence of drug resistance, must be
overcome for the effective treatment of gastric cancer (6,7,30).
Recently, use of the biological properties of natural plants to
develop antitumor agents has attracted increased interest. In the
present study, we evaluated the mechanisms of an ethanol extract of
Gleditsia sinensis thorns (EEGS) in the potential
therapeutic effects underlying cell cycle control, signal
transduction pathways and MMP regulation.
Treatment of SNU-5 cells with EEGS (200–800 μg/ml)
resulted in inhibition of cell growth. Apoptotic cells were
observed following high-dose treatments (600–800 μg/ml). The
potential of chemopreventive or chemotherapeutic agents to suppress
the growth of cancer cells is associated with blocking the G1 to S
transition checkpoint. In general, G1 to S cell cycle progression
is regulated by activation of CDK regulatory proteins and cyclin
complexs (8–10). Therefore, the induction of G1 phase
cell cycle arrest in cancer cells is proposed as a critical
therapeutic approach, which inhibits cell cycle progression and
tumor growth. EEGS treatment induced G1 phase cell cycle arrest,
with a concomitant decrease in cyclin D1/CDK4, and cyclin E/CDK2
complexes in SNU-5 cells. These results are inconsistent with those
of previous studies indicating that EEGS induced G2/M phase cell
cycle arrest in colon cancer cells (31). The results of the present study
demonstrated that EEGS caused G1 phase cell cycle arrest via a
reduction in cyclins and CDKs, which are regulatory molecules
involved in G1 to S cell cycle progression in SNU-5 cells.
p21WAF1, one of the CDKI family proteins (CDK
inhibitors), is a universal inhibitor of CDKs, the expression
levels of which are normally regulated by the kinase activity of
CDK/cyclin complexes through either a p53-dependent or a
p53-independent mechanism (11,32).
In addition, p21WAF1 plays an essential role in the cellular stress
induced by anti-proliferative signals, which inhibit the G1 to S
phase cell cycle progression (11).
Previous studies have shown that accumulation of p27KIP1 is
associated with G2/M phase cell cycle arrest in colon cancer cells
(31). Our data revealed that EEGS
increases p21WAF1 protein expression during G1 phase cell cycle
arrest in SNU-5 cells. However, the expression levels of other
CDKIs, including p27KIP1 and p53, remained essentially unchanged.
These results suggest that induction of p21WAF1 may also be
responsible for G1 phase arrest in EEGS-treated SNU-5 cells.
Collectively, the mechanistic investigation showed that EEGS causes
p21WAF1-mediated G1 phase cell cycle arrest via downregulation of
cyclin/CDK complexes in SNU-5 cells.
Regulation of cell growth inhibition occurs through
various mechanisms including the MAPK signaling pathway (12–14).
In the present study, we examined the potential involvement of MAPK
pathways, such as ERK, p38 MAP kinase and JNK, in the EEGS-induced
inhibition of cell growth. Treatment of cells with EEGS induced
activation of ERK1/2 and p38 MAP kinase. Previous studies have
demonstrated that the MAPK pathway is accompanied by change in the
cell cycle regulation (13,14,33,34).
Since EEGS induced p21WAF1 expression, the involvement of MAPK
signaling pathways in EEGS-induced p21WAF1 expression was
investigated. Inhibition of p38 MAP kinase, using a pharmacological
inhibitor (SB203580), reversed p21WAF1 expression induced by EEGS.
However, pretreatment of cells with U0126 (an ERK1/2 inhibitor) did
not affect the EEGS-mediated p21WAF1 expression. These data suggest
that ERK1/2 activation is not required for p21WAF1 expression in
response to EEGS, although EEGS induces activation of ERK1/2. The
results of the present study showed that the activation of p38 MAP
kinase is a main factor in the regulation of EEGS-induced p21WAF1
expression in SNU-5 cells. In addition, the inhibition of p38 MAP
kinase by pre-incubation with SB203580 restored the cell growth
inhibition and downregulation of G1 phase cell cycle-associated
proteins, CDK2 and CDK4. Our findings obtained by the inhibition of
p38 MAP kinase indicate that p38 MAP kinase is involved in the
inhibition of cell proliferation by suppressing cell cycle
regulatory proteins, the CDKs. The present study is the first to
show that the p38 MAP kinase signaling pathway may be responsible
for p21WAF1-mediated G1 phase cell cycle arrest in the
EEGS-mediated inhibition of cell growth.
Matrix metalloproteinase-9 (MMP-9) is a major
component involved in the degradation of the extracellular matrix
(ECM) and participates in the metastatic progression of gastric
cancer (17–19). A previous report showed that TNF-α
enhances MMP-9 expression in gastric cancer cell lines (35). Based on these studies, we further
investigated the effects of EEGS on the regulatory mechanism of
MMP-9 in TNF-α-treated SNU-5 cells. In TNF-α-treated cells, using
both zymographic and immunoblot analyses, TNF-α-induced MMP-9
expression was inhibited by treatment with EEGS at the protein
level, without altering the level of MMP-2. It is well known that
transcription factors NF-κB, AP-1, and Sp-1 are involved in MMP-9
expression in response to TNF-α in several cell lines (20–23,36).
However, the identification of cis-elements in the induction
of MMP-9 by TNF-α in gastric cancer has not been addressed to date.
Our data from the present study represent the first evidence that
TNF-α effectively enhances MMP-9 expression via increasing both
AP-1 and NF-κB binding activities in a gastric cancer cell line.
However, unexpectedly, we did not observe the Sp-1 binding activity
in TNF-α-treated cells. In addition, we found that treatment of
cells with EEGS showed significantly decreased binding activities
in both AP-1 and NF-κB motifs in response to TNF-α. Our results
suggest that NF-κB and AP-1 sites are important for the inhibition
of EEGS-mediated MMP-9 expression in TNF-α-treated SNU-5 cells.
In conclusion, the results of the present study
revealed that EEGS-induced inhibition of cell growth was associated
with p38 MAP kinase activation via p21WAF1-mediated G1 phase cell
cycle arrest, which involved a decrease in the cyclin and CDK
complexes in gastric cancer SNU-5 cells. In addition, EEGS
significantly inhibited MMP-9 expression via the suppression of the
binding activities of NF-κB and AP-1 cis-elements in
TNF-α-treated cells. The results of the present study warrant the
development of EEGS as a novel anticancer agent for the effective
therapy of human gastric cancer. However, further study is required
to elucidate the effects of the EEGS compound in regards to its
molecular mechanisms, which are responsible for its in vivo
efficacy.
Acknowledgements
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korean government
(MEST) (no. 2012-0000482) and by a grant from the Next-Generation
BioGreen 21 Program (no. PJ0081952011), Rural Development
Administration, Republic of Korea.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics. CA Cancer J Clin. 59:225–249. 2009.
|
|
2
|
Lau M, Le A and El-Serag HB: Non-cardia
gastric adenocarcinoma remains an important and deadly cancer in
the United States: secular trends in incidence and survival. Am J
Gastroenterol. 101:2485–2492. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cervantes A, Roselló S, Roda D and
Rodríguez-Braun E: The treatment of advanced gastric cancer:
current strategies and future perspectives. Ann Oncol. 19:103–107.
2008. View Article : Google Scholar
|
|
4
|
Tham CK, Choo SP, Poon DYH, et al:
Capecitabine with radiation is an effective adjuvant therapy in
gastric cancers. World J Gastroenterol. 16:3709–3715. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Watanabe T, Kume K, Taip M, et al: Gastric
mucosal cancer smaller than 7 mm can be treated with conventional
endoscopic mucosal resection as effectively as with endoscopic
submucosal dissection. Hepatogastroenterology. 57:668–673.
2010.
|
|
6
|
Kang HC, Kim IJ, Park HW, et al:
Regulation of MDK expression in human cancer cells modulates
sensitivities to various anticancer drugs: MDK overexpression
confers to a multi-drug resistance. Cancer Lett. 247:40–47. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yin F, Shi YQ, Zhao WP, Xiao B, Miao JY
and Fan DM: Suppression of P-gp induced multiple drug resistance in
a drug resistant gastric cancer cell line by over-expression of
Fas. World J Gastroenterol. 6:664–670. 2000.PubMed/NCBI
|
|
8
|
Sherr CJ: Cancer cell cycles. Science.
274:1672–1677. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jacks T and Weinberg RA: Cell-cycle
control and its watchman. Nature. 381:643–644. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Collins K, Jacks T and Pavletich NP: The
cell cycle and cancer. Proc Natl Acad Sci USA. 94:2776–2778. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Peter M and Herskowitz I: Joining the
complex: cyclin-dependent kinase inhibitory proteins and the cell
cycle. Cell. 79:181–184. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xia Z, Dickens M, Raingeaud J, Davis RJ
and Greenberg ME: Opposing effects of ERK and JNK-p38 MAP kinases
on apoptosis. Science. 270:1326–1331. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thornton TM and Rincon M: Non-classical
p38 map kinase functions: cell cycle checkpoints and survival. Int
J Biol Sci. 5:44–51. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han J and Sun P: The pathways to tumor
suppression via route p38. Trends Biochem Sci. 32:364–371. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liotta LA: Tumor invasion and
metastasis-role of extracellular matrix: Rhoads Memorial Award
Lecture. Cancer Res. 46:1–7. 1986.PubMed/NCBI
|
|
16
|
Matrisian LM: Metalloproteinases and their
inhibitors in matrix remodeling. Trends Genet. 6:121–125. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sampieri CL, de la Peña S, Ochoa-Lara M,
Zenteno-Cuevas R and León-Córdoba K: Expression of matrix
metalloproteinases 2 and 9 in human gastric cancer and superficial
gastritis. World J Gastroenterol. 16:1500–1505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng H, Takahashi H, Murai Y, et al:
Expressions of MMP-2, MMP-9 and VEGF are closely linked to growth,
invasion, metastasis and angiogenesis of gastric carcinoma.
Anticancer Res. 26:3579–3583. 2006.PubMed/NCBI
|
|
19
|
Yamanaka N, Morisaki T, Nakashima H, et
al: Interleukin 1beta enhances invasive ability of gastric
carcinoma through nuclear factor-kappaB activation. Clin Cancer
Res. 10:1853–1859. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bond M, Rosalind P, Fabunmi P, Baker AH
and Newby AC: Synergistic upregulation of metalloproteinase-9 by
growth factors and inflammatory cytokines: an absolute requirement
for transcription factor NF-kappa B. FEBS Lett. 435:29–34. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sato H, Kita M and Seiki M: v-Src
activates the expression of 92-kDa type IV collagenase gene through
the AP-1 site and the GT box homologous to retinoblastoma control
elements. A mechanism regulating gene expression independent of
that by inflammatory cytokines. J Biol Chem. 268:23460–23468.
1993.PubMed/NCBI
|
|
22
|
Sato H and Seiki M: Regulatory mechanism
of 92 kDa type IV collagenase gene expression which is associated
with invasiveness of tumor cells. Oncogene. 8:395–405.
1993.PubMed/NCBI
|
|
23
|
Farina AR, Tacconelli A, Vacca A, Maroder
M, Gulino A and Mackay AR: Transcriptional up-regulation of matrix
metalloproteinase-9 expression during spontaneous epithelial to
neuroblast phenotype conversion by SK-N-SH neuroblastoma cells,
involved in enhanced invasivity, depends upon GT-box and nuclear
factor kappaB elements. Cell Growth Differ. 10:353–367. 1999.
|
|
24
|
Ahn DK: Illustrated Book of Korean
Medicinal Herbs. Kyohak Publishing Co; Seoul: pp. 6282003
|
|
25
|
Lim JC, Park JH, Budesinsky M, et al:
Antimutagenic constituents from the thorns of Gleditsia
sinensis. Chem Pharm Bull. 53:561–564. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou L, Li D, Wang J, Liu Y and Wu J:
Antibacterial phenolic compounds from the spines of Gleditsia
sinensis Lam. Nat Prod Res. 21:283–291. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li WH, Zhang XM, Tian RR, Zheng YT, Zhao
WM and Qiu MH: A new anti-HIV lupane acid from Gleditsia
sinensis Lam. J Asian Nat Prod Res. 9:551–555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shin TY and Kim DK: Inhibitory effect of
mast cell-dependent anaphylaxis by Gleditsia sinensis. Arch
Pharm Res. 23:401–406. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Park E and Shin MJ: Anti-inflammatory
activity of aqueous extract from Gleditsiae Spina. Arch
Pharm Res. 37:124–128. 1993.
|
|
30
|
Panchal RG: Novel therapeutic strategies
to selectively kill cancer cells. Biochem Pharmacol. 55:247–252.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee SJ, Cho YH, Kim H, et al: Inhibitory
effects of the ethanol extract of Gleditsia sinensis thorns
on human colon cancer HCT116 cells in vitro and in
vivo. Oncol Rep. 22:1505–1512. 2009.PubMed/NCBI
|
|
32
|
Macleod KF, Sherry N, Hannon G, et al:
p53-dependent and independent expression of p21 during cell growth,
differentiation, and DNA damage. Genes Dev. 9:935–944. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chambard JC, Lefloch R, Pouyssegur J and
Lenormand P: ERK implication in cell cycle regulation. Biochim
Biophys Acta. 1773:1299–1310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Malumbres M, Pérez De Castro I, Hernández
MI, Jiménez M, Corral T and Pellicer A: Cellular response to
oncogenic ras involves induction of the Cdk4 and Cdk6 inhibitor
p15(INK4b). Mol Cell Biol. 20:2915–2925. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim S, Choi MG, Lee HS, et al: Silibinin
suppresses TNF-alpha-induced MMP-9 expression in gastric cancer
cells through inhibition of the MAPK pathway. Molecules.
14:4300–4311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moon SK, Cha BY and Kim CH: ERK1/2
mediates TNF-alpha-induced matrix metalloproteinase-9 expression in
human vascular smooth muscle cells via the regulation of NF-kappaB
and AP-1: involvement of the ras dependent pathway. J Cell Physiol.
198:417–427. 2004. View Article : Google Scholar
|