Introduction
Malignant pleural mesothelioma (MPM) is a deadly
disease that arises from the mesothelial layer of the pleural
cavity and afflicts 2,000 to 3,000 individuals every year in the US
(1,2). MPM has been linked to asbestos, iron
and Simian virus 40 exposure (3–5) and
represents a worldwide concern since its incidence has greatly
increased during the last three decades (6). MPM progresses rapidly and is
extraordinarily delibitating, particularly in the later stages. In
addition, its poor prognosis stems from an overall survival period
of only 9–17 months from the initial diagnosis (2). An aggressive multimodality therapy for
MPM can offer survival benefits not only for a small subset of
patients in the early stages of the disease but also for terminal
patients with unresectable tumors who are not responsive to current
therapeutic approaches (2,7). This suggests an urgent need for new
therapeutic targets and effective therapies for improving the
prognosis of MPM. Several studies have reported that novel
plant-derived compounds act as antitumor agents through the
modulation of biological pathways (8). This suggests that chemotherapeutic
agents used in the treatment of MPM may activate apoptosis
pathways.
The present study focused on honokiol (HNK) as this
anticancer agent is widely used as the drug of choice in treating
cancer. HNK, a biologically active biphenolic compound isolated
from the Magnolia species, has multiple medicinal uses
against oxidative stress, microbial infection and anxiety, among
others (9). Noteworthy, HNK
demonstrates antineoplastic properties in vitro against
various types of cancers and is effective in vivo in
treating angiosarcoma (10),
colorectal carcinoma (11), breast
cancer (12) and gastric cancer
(13). Several studies have
suggested that the cytotoxic effect of HNK is caused by the
predominant activation of distinct apoptotic mechanisms in tumor
cells (10,11,14–16).
Apart from apoptosis, the suppression of cancer cell proliferation
is one of the most important strategies for antitumorigenesis.
Specificity protein 1 (Sp1), a constitutive
transcription factor, plays an important role in various
physiological processes such as cell cycle regulation, apoptosis
and angiogenesis (17,18). In this regard, many studies have
documented that Sp1 activity and expression levels are increased in
various types of human cancers including prostate cancer,
pancreatic adenocarcinoma, thyroid cancer, hepatocellular
carcinomas, lung cancer, colorectal cancer, gastric cancer and
breast cancer, but not in normal tissues (19,20).
Therefore, the present study examined the regulatory
effects of HNK on the growth and apoptosis of MSTO-211H
mesothelioma cells and investigated its anticancer mechanism in
relation to Sp1.
Materials and methods
Cell culture and transfection
MSTO-211H cells obtained from the American Tissue
Culture Collection (Manassas, VA) were maintained at 37°C in
RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS) and
penicillin/streptomycin in the presence of 5% CO2. For
transient transfection, 1.4×106 cells were plated onto a
60-mm cell culture dish, grown overnight, and transfected with DNA
using Lipofectamine (Invitrogen).
MTS assay
The effects of HNK on cell viability were determined
using the CellTiter 96 AQueous One Solution cell proliferation
assay kit (Promega, Madison, WI) according to the manufacturer’s
instructions. MSTO-211H cells were seeded onto a 96-well plate for
24 h and then treated with HNK for 24 and 48 h. The MTS solution
was analyzed using the GloMax-Multi Microplate Mulimode reader
(Promega) at 482 and 690 nm.
DAPI staining
The number of cells undergoing apoptosis following
treatment with HNK was quantified using DAPI staining. Cells with
nuclear condensation and fragmentation were determined using the
nucleic acid stain 4′-6-diamidino-2-phenylindole (DAPI)
(Sigma-Aldrich). Following a 48-h treatment of HNK at different
concentrations (2, 4 and 6 μg/ml), MSTO-211H cells were harvested
and fixed in 100% methanol at room temperature for 20 min. The
cells were seeded on slides, stained with DAPI (2 mg/ml), and then
monitored using a FluoView confocal laser microscope (FluoView
FV10i, Olympus Corp., Tokyo, Japan).
Propidium iodide staining
Following HNK treatment of the MSTO-211H cells,
detached cells were collected separately, and adherent cells were
dissociated by trypsin-EDTA. The cells were washed with cold PBS
and then pooled and centrifuged before being fixed in 70% ethanol
overnight at −20°C. Prior to the flow cytometric analysis, the
cells were centrifuged and incubated for 30 min at 37°C in PBS to
allow for the release of low-molecular-weight DNA. After
centrifugation, cell pellets were resuspended and treated with 150
mg/ml RNase A and 20 mg/ml propidium iodide (PI) using a MACSQuant
Analyzer (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany).
Western blot analysis
MSTO-211H cells treated with HNK were washed three
times with ice-cold phosphate-buffered saline (PBS) and harvested
in an ice-cold PRO-PREP™ protein extraction solution (Intron
Biotechnology, Inc., Korea) containing a protease inhibitor.
Protein concentrations were measured using the Bradford protein
assay. Protein samples were separated on an SDS-PAGE gel and
transferred to a PVDF membrane (Immobilon-P, Millipore) using a
semi-dry blotting apparatus. ECL western blotting was performed
according to the manufacturer’s instructions (Thermo Scientific,
Rockford, IL).
In vitro pull-down assays
Cell lysates were incubated with HNK-Sepharose 4B
(or Sepharose 4B only as a control) beads (50 μl, 50% slurry) in
reaction buffer (50 mM Tris, pH 7.5, 5 mM EDTA, 150 mM NaCl, 1 mM
dithiothreitol, 0.01% Nonidet P-40, 2 μg/ml bovine serum albumin,
0.02 mM phenylmethylsulfonyl fluoride and 1X protease inhibitor
cocktail). After incubation with gentle rocking overnight at 4°C,
the beads were washed five times with a buffer solution (50 mM
Tris, pH 7.5, 5 mM EDTA, 150 mM NaCl, 1 mM dithiothreitol, 0.01%
Nonidet P-40 and 0.02 mM phenylmethylsulfonyl fluoride) and
proteins bound to the beads were analyzed by western blotting using
the anti-Sp1 antibody.
Luciferase assay
MSTO-211H cells were grown to 50–80% confluence on a
60-mm cell dish and were transfected with Lipofectamine
(Invitrogen) with 1 μg each of cyclin D1-luc, Mcl-1-luc and
survivin-luc and 0.5 μg of pCMV/β-gal plasmids. The luciferase
reporter assay kit from Promega was used following the
manufacturer’s protocol. Luciferase activity was measured with the
Lumat LB 9507 luminometer (EG&G Berthold). The values shown
represent an average of three experiments for each sample.
Statistical analysis
The statistical significance of differences was
assessed using the Student’s t-test. A P-value <0.05 relative to
the control was considered to indicate a statistically significant
result.
Results
Growth inhibitory effect of HNK on a
human mesothelioma cell line
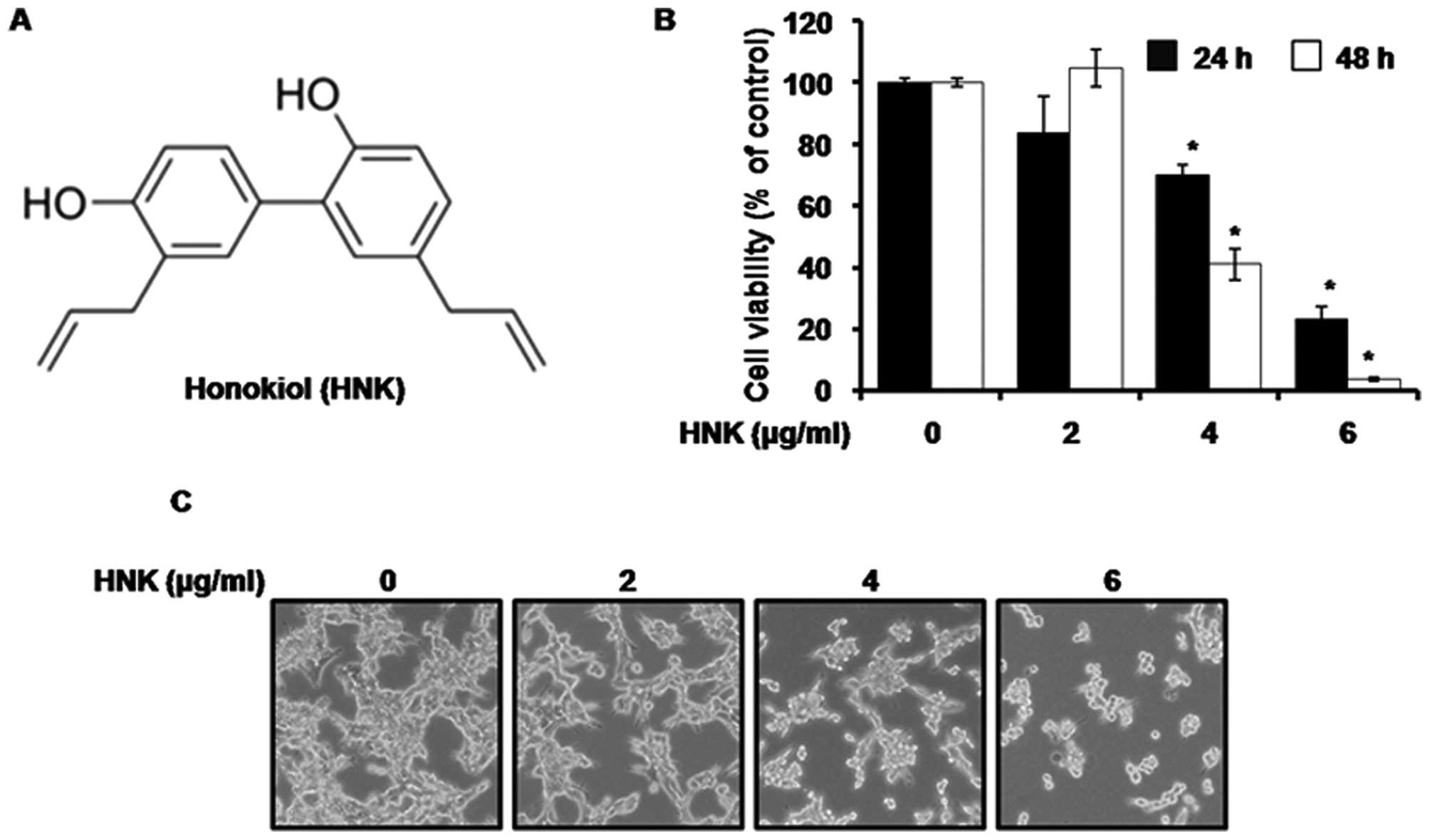
We investigated the effects of HNK on a human
mesothelioma cell line. The structure of HNK is shown in Fig. 1A. To ascertain the efficiency of HNK
as an anticancer drug, the effects of HNK were tested using an MTS
assay using MSTO-211H cells. As shown in Fig. 1B, the efficiency of HNK in altering
the cell viability of MSTO-211H cells was assayed after 24 and 48 h
of incubation in HNK-containing medium at different concentrations
(2, 4 or 6 μg/ml). The viability curves revealed that HNK reduced
MSTO-211H cell viability at 24 and 48 h, in a
concentration-dependent manner (P<0.05). In particular, a
maximum reduction in cell viability was observed at 48 h.
Apoptosis-induced changes in the cell morphology in the
HNK-containing medium were observed. After 48 h, the apoptotic
phenotype showed cell rounding, cytoplasmic blebbing and
irregularities in shape, indicating a sharp increase in the
apoptosis of HNK-treated MSTO-211H cells in a
concentration-dependent manner (Fig.
1C).
HNK induces cell cycle arrest at the G1
phase and apoptosis in a human mesothelioma cell line
Cancer cell growth can be suppressed by cell cycle
arrest or apoptosis induction or both (8). Morphological changes induced by HNK
were examined following treatment of MSTO-211H cells with HNK at a
concentration of 2, 4 and 6 μg/ml for 48 h. In the DAPI staining
analysis, MSTO-211H cells were observed via fluorescence microscopy
following exposure to DAPI, which specifically stains nuclei. The
results indicated the presence of nuclear condensation and
perinuclear apoptotic bodies in the MSTO-211H cells following HNK
treatment at concentrations of 4 and 6 μg/ml for 48 h (Fig. 2A). The percentage of apoptosis was
determined to be 6±1.6, 26±11.3 and 20±20.3% at concentrations of
2, 4 and 6 ng/ml HNK compared with the untreated control cells.
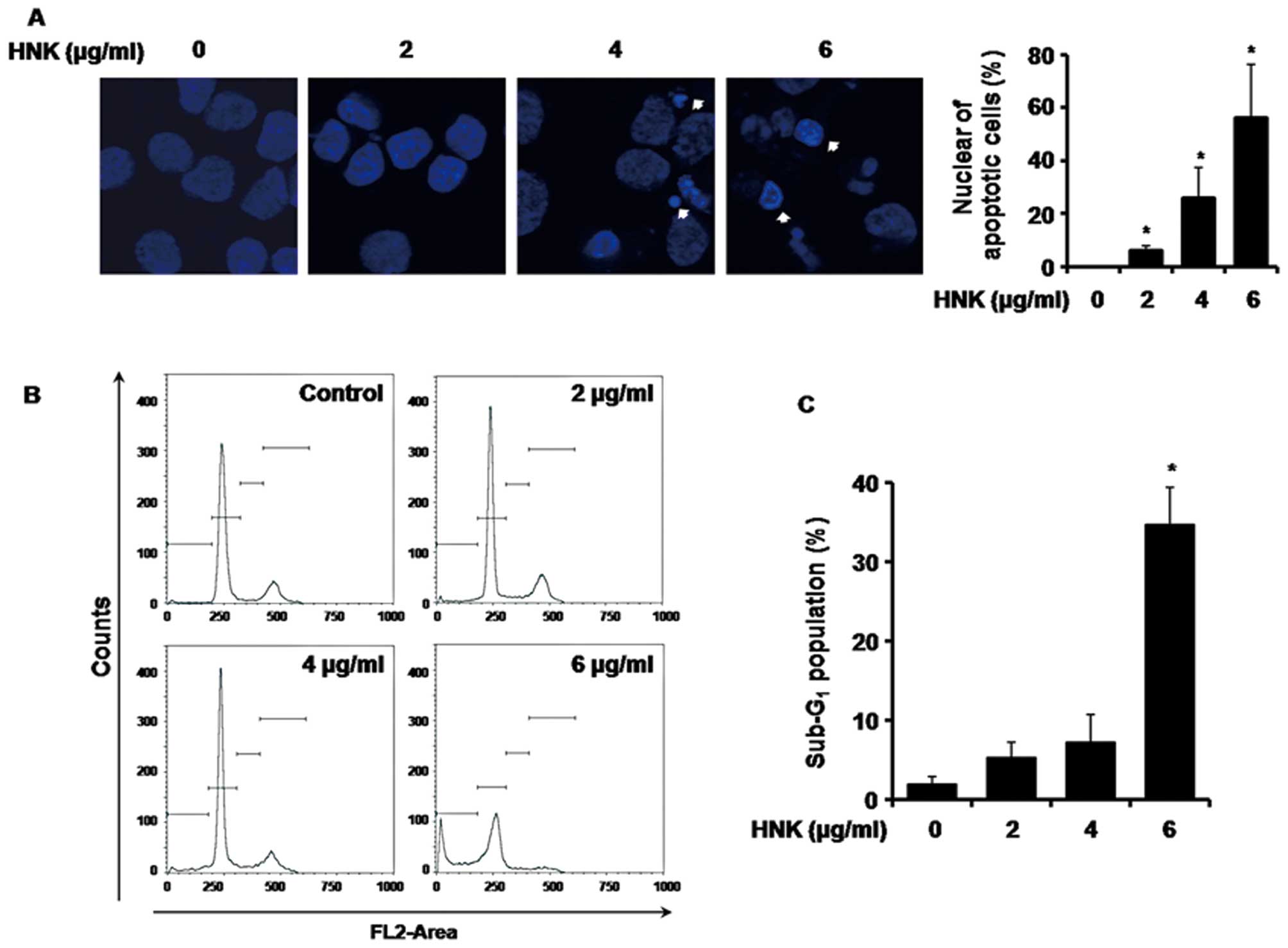 | Figure 2Effects of HNK on the proliferation of
MSTO-211H cells. (A, left) MOTO-211H cells were cultured without
(control) or with HNK (2, 4 and 6 μg/ml) for 48 h in each case.
Images of DAPI-stained MSTO-211H cells are based on fluorescence
microscopy (magnification, ×600). White arrows indicate condensed
nuclei. (A, right) DNA fragmentation and nuclear condensation were
quantified, and the data represent the mean percentage levels ± SD
(n=3; *P<0.05). (B) MSTO-211H cell cultures were
treated with 2, 4 and 6 μg/ml HNK or PBS (vehicle). The cells were
then washed, fixed and stained with PI and analyzed for DNA content
through FACS analysis 48 h after treatment. Ratios of apoptotic
cells were measured by FACS analysis after PI staining. (C) The
data represent the percentage of the G0–G1
population in MSTO-211H cells following a 48-h treatment with DMSO
(control) or 2, 4, and 6 μg/ml HNK. *P<0.05,
significantly different compared with DMSO-treated control by
paired t-test. Repeated experiments produced similar results
(n=3). |
For the determination of whether the HNK-mediated
growth inhibition of MSTO-211H cells was attributable to cell cycle
arrest, cell cycle distribution was analyzed by FACS analysis. As
shown in Fig. 2B, a significant
increase in the number of sub-G1 cells was noted when
compared with the control (5.3±1.9, 7.2±3.5 and 34.7±4.6% in the
presence of 2, 4 and 6 μg/ml HNK, respectively, in MSTO-211H
cells). The histogram in Fig. 2C
shows the quantification of FACS data.
HNK suppresses Sp 1 expression in a human
mesothelioma cell line
The potential interaction between HNK and
apoptosis-related proteins was examined since HNK has been found to
have marked effects on apoptosis and cell cycle arrest in various
cancer cell lines. Therefore, an in vitro pull-down assay
was performed using HNK-conjugated Sepharose 4B beads. Sp1 was
found to bind to HNK-conjugated Sepharose beads but not to
Sepharose beads alone (Fig. 3A),
providing support for the direct interaction between HNK and Sp1
and suggesting that Sp1 may be a possible target of HNK in human
MSTO-211H cells. The effects of HNK treatment on the levels of Sp1
were examined by western blotting. As shown in Fig. 3B, HNK treatment led to a sharp
decrease in the level of Sp1 in MSTO-211H cells at 48 h after
treatment.
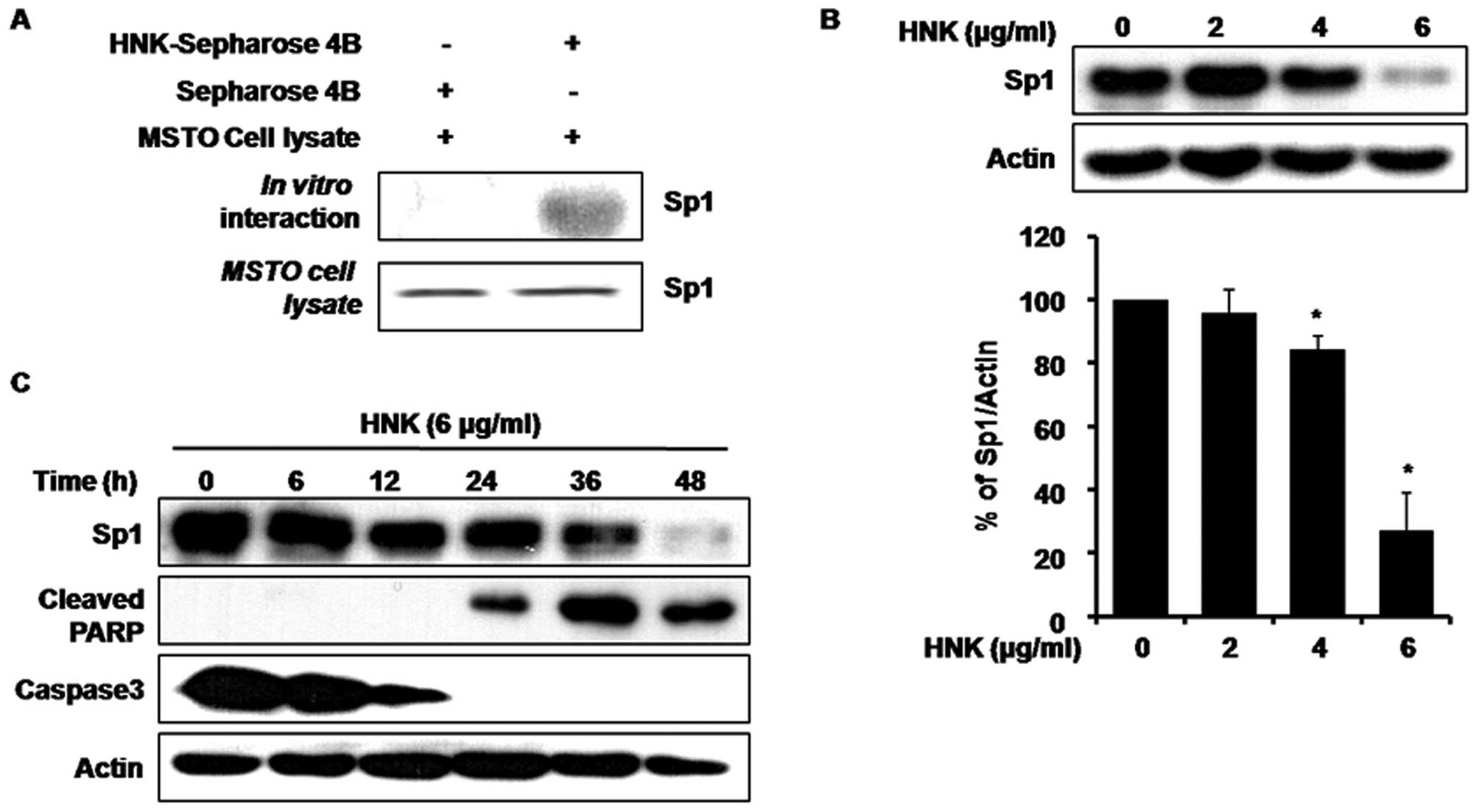 | Figure 3HNK binds to Sp1 and suppresses Sp1
levels via apoptosis. (A) MSTO-211H cell lysates were prepared,
subjected to a pull-down assay using HNK-Sepharose 4B beads (or
only Sepharose 4B as a control), and subjected to western blot
analysis. (B) MSTO-211H cells were treated with 2, 4 and 6 μg/ml
HNK for 48 h, and whole-cell extracts were prepared and separated
on SDS-PAGE and then subjected to western blotting with an anti-Sp1
antibody. Actin was used as a loading control. The graphs indicate
the ratio of Sp1 to actin expression. (C) Experiments to assess the
time-dependent effects of HNK on Sp1, cleaved PARP, and caspase-3
expression were performed using MSTO-211H cells treated with 6
μg/ml HNK for 0, 6, 12, 24, 36 and 48 h. |
In order to better characterize the apoptotic action
of HNK, the cleavage of the DNA repair enzyme poly(adenosine
diphosphate-ribose) polymerase (PARP) as a substrate for activated
caspase-3 and caspase-3 expression levels were confirmed by western
blotting (Fig. 3C). There were
increases in cleaved PARP and activated caspase-3 following HNK (6
μg/ml) treatment of MSTO-211H cells in a time-dependent manner.
HNK modulates regulators of cell cycle
arrest and apoptosis
Sp1 activation has been shown to regulate the
expression of various gene products involved in cell survival and
proliferation (18,21). Based on the results for DNA
fragmentation and cell cycle distribution, the potential
relationship between the induction of apoptosis and cell cycle
arrest by HNK and the transcriptional activity of cell cycle
regulatory proteins and apoptosis-related transcription factors was
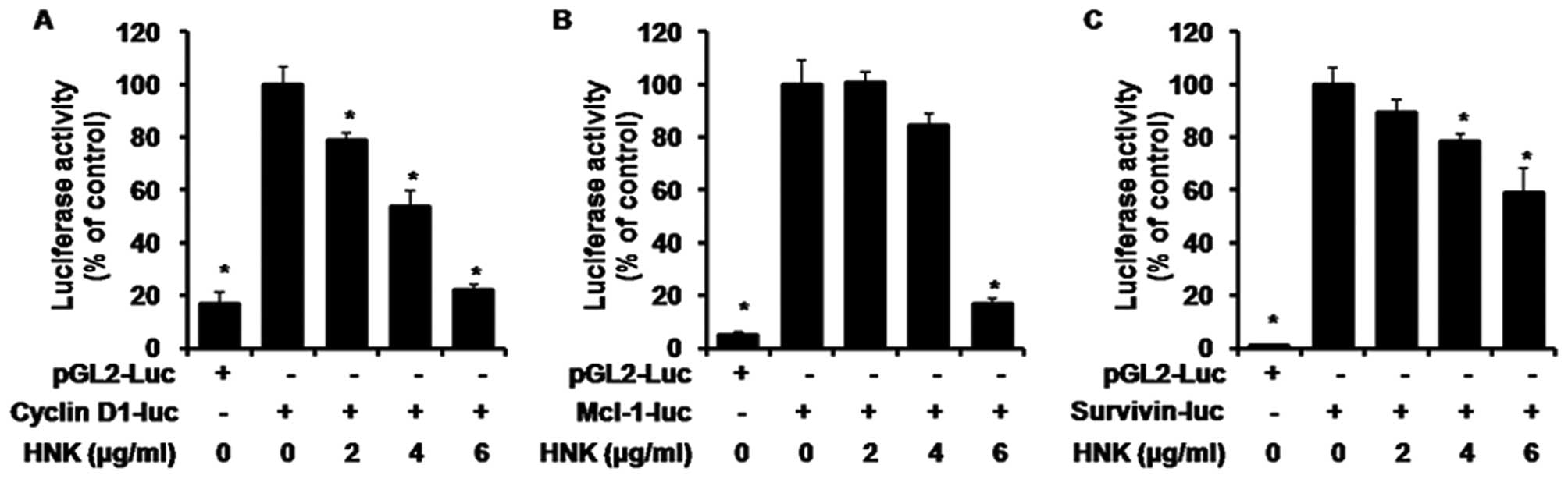
examined. We investigated the effects of HNK on cell cycle
regulator cyclin D1, anti-apoptotic proteins Mcl-1 and survivin
dependent gene transcriptions. When the MSTO-211H cells transiently
transfected with cyclin D1-luc (Fig.
4A), Mcl-1-luc (Fig. 4B), and
survivin-luc (Fig. 4C) constructs
were treated with 2, 4 and 6 μg/ml HNK, significant decreases in
transcriptional activity mediated by these proteins were observed
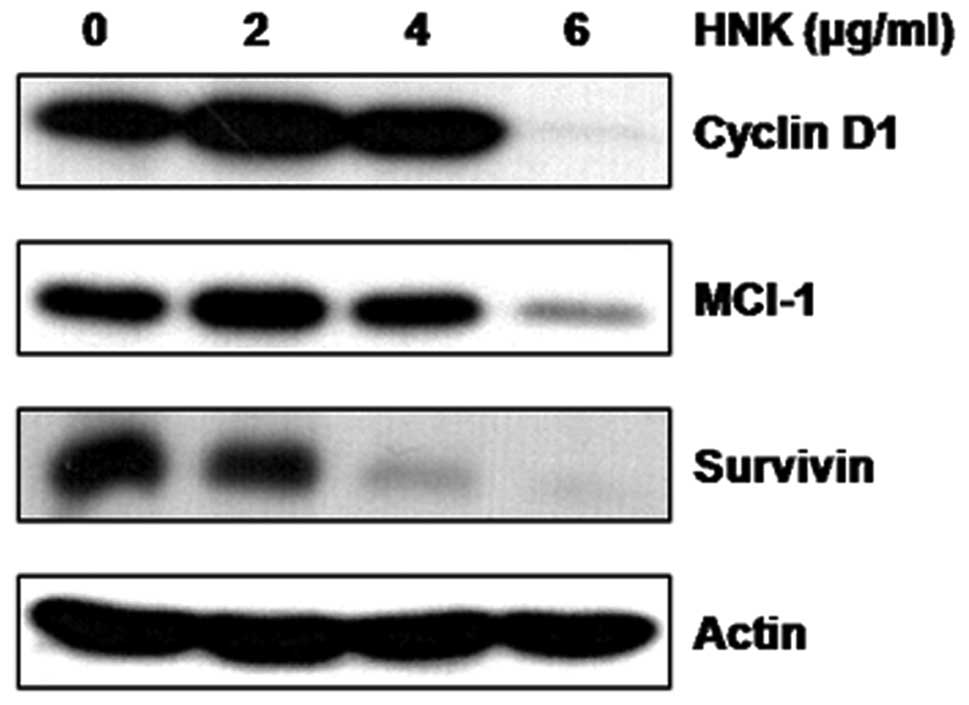
in a dose-dependent manner. The cyclin D1, Mcl-1 and survivin
protein expression levels were examined using western blotting. The
results indicated that these levels decreased gradually after HNK
treatment (Fig. 5).
HNK alters the expression of proteins
associated with the cell cycle and survival
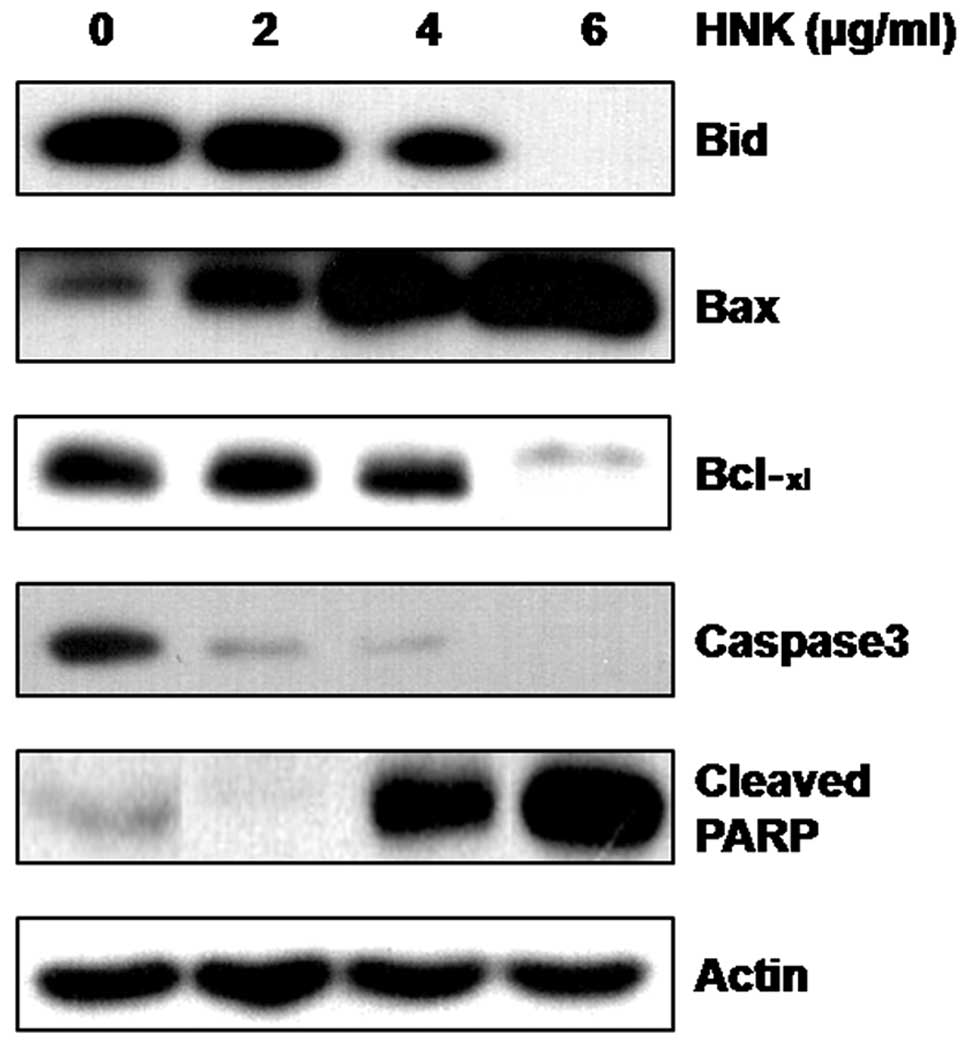
The expression of genes involved in the regulation
of apoptosis was analyzed. The apoptotic cell death by HNK was well
correlated with the activation of caspase-3 through the cleavage of
the pro-form during HNK treatment. The upregulation and
downregulation of Bax and Bid, respectively, and Bcl-xL expression
appeared to be involved in apoptotic cell death. In addition, PARP
cleavage was induced by HNK (Fig.
6). These results indicate that downregulation of cyclin D1,
Mcl-1 and survivin and activation of caspase-3 were involved in the
HNK-mediated growth arrest and apoptotic cell death in MSTO-211H
cells.
Discussion
Compounds of natural origin such as topotecan,
irinotecan, etoposide, teniposide and paclitaxil are widely
employed in clinical trials as chemotherapeutic agents (22). Therefore, the evaluation of natural
compounds with anticancer properties has been considered to play an
important role in the development of anticancer drug. The present
study focused on the anticancer effects of HNK, a pharmacologically
active component found in the traditional Chinese medicinal herb,
Magnolia species. Previous studies have demonstrated the
anticancer properties of HNK, including cell cycle arrest,
apoptosis and necrosis in a variety of cancer cell lines such as
human breast cancer (23), human
colorectal carcinoma RKO (24),
human squamous lung cancer CH27 (15), human pancreatic cancer (25), adult T-cell leukemia (26) and hepatocellular carcinoma cells
(27). This suggests that HNK may
be involved in the apoptotic pathway. However, the exact mechanism
underlying the anticancer activity of HNK remains unclear.
In this study, our findings along with the findings
of previous research, suggest that HNK may induce apoptosis in
human MPM cells. In this study, the apoptotic effects of HNK on
MSTO-211H cells were examined through an MTS assay. This assay, a
traditional method for discovering anticancer drugs, can be used to
determine the cytotoxic effect and proliferation of cell lines
(28,29). In this study, this assay was used to
elucidate the apoptotic effects of HNK on MSTO-211H cells (Fig. 1).
As shown in Fig. 2,
HNK inhibited the proliferation of MSTO-211H cells through cell
cycle arrest at G0/G1 and induction of
apoptosis.
Sp1, a transcription factor, is overexpressed in
various human cancer cell lines (30–34)
and plays a role in the regulation of genes involved in most
cellular processes (35). In this
regard, Sp1 may exhibit transcriptional activity for promoters of
genes involved in the progression, differentiation and oncogenesis
of the cell cycle (19). In
addition, Sp1 may be inhibited by compounds of natural origin with
anticancer properties. For example, recent studies have
demonstrated that flavonoids suppress Sp1 expression (36) and, thus, inhibit the known Sp1
target genes cyclin D1, Mcl-1 and survivin in a selective manner
(37–41). The results of the present study
demonstrated the potential chemotherapeutic properties of HNK. That
is, HNK suppressed Sp1 expression via direct interactions with Sp1,
preventing Sp1 from binding to G-C-rich promoters. In addition, HNK
inhibited the transcriptional activity and expression of Sp1
downstream proteins, including cyclin D1, Mcl-1 and survivin, in a
dose-dependent manner (Figs. 4 and
5). HNK reduced Bid and Bcl-xL,
increased Bax, and activated caspase-3 and PARP, suggesting that
HNK regulated Sp1 and ultimately led to apoptotic cell death
(Fig. 6).
The results regarding the chemopreventive effect of
HNK as a compound of natural origin on MPM cells revealed that HNK
downregulated Sp1 expression and Sp1 target transcription factors,
including cyclin D1, Mcl-1 and survivin, thereby inducing the
apoptosis of MSTO-211H cells. To conclude, this study is the first
to suggest that the apoptosis induced by HNK in MSTO-211H cells may
be mediated by the downregulation of Sp1. The results suggest that
the antiproliferative and apoptotic effects of HNK are mediated
through the suppression of Sp1-regulated gene products.
Acknowledgements
This research was supported by the Basic Science
Research Program of the National Research Foundation Korea (NRF),
funded by the Ministry of Education, Science and Technology
(2011-0008463); the Next-Generation BioGreen 21 Program
(PJ008116062011) of the Rural Development Administration of the
Republic of Korea.
References
|
1
|
Cugell DW and Kamp DW: Asbestos and the
pleura: a review. Chest. 125:1103–1117. 2004. View Article : Google Scholar
|
|
2
|
Tsao AS, Wistuba I, Roth JA and Kindler
HL: Malignant pleural mesothelioma. J Clin Oncol. 27:2081–2090.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Broaddus VC: Asbestos, the mesothelial
cell and malignancy: a matter of life or death. Am J Respir Cell
Mol Biol. 17:657–659. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morinaga K, Kishimoto T, Sakatani M, Akira
M, Yokoyama K and Sera Y: Asbestos-related lung cancer and
mesothelioma in Japan. Ind Health. 39:65–74. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dufresne A, Begin R, Churg A and Masse S:
Mineral fiber content of lungs in patients with mesothelioma
seeking compensation in Quebec. Am J Respir Crit Care Med.
153:711–718. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Britton M: The epidemiology of
mesothelioma. Semin Oncol. 29:18–25. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zellos L and Sugarbaker DJ: Current
surgical management of malignant pleural mesothelioma. Curr Oncol
Rep. 4:354–360. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gupta SC, Kim JH, Prasad S and Aggarwal
BB: Regulation of survival, proliferation, invasion, angiogenesis,
and metastasis of tumor cells through modulation of inflammatory
pathways by nutraceuticals. Cancer Metastasis Rev. 29:405–434.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liou KT, Shen YC, Chen CF, Tsao CM and
Tsai SK: Honokiol protects rat brain from focal cerebral
ischemia-reperfusion injury by inhibiting neutrophil infiltration
and reactive oxygen species production. Brain Res. 992:159–166.
2003. View Article : Google Scholar
|
|
10
|
Bai X, Cerimele F, Ushio-Fukai M, et al:
Honokiol, a small molecular weight natural product, inhibits
angiogenesis in vitro and tumor growth in vivo. J Biol Chem.
278:35501–35507. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen F, Wang T, Wu YF, et al: Honokiol: a
potent chemotherapy candidate for human colorectal carcinoma. World
J Gastroenterol. 10:3459–3463. 2004.PubMed/NCBI
|
|
12
|
Wolf I, O’Kelly J, Wakimoto N, et al:
Honokiol, a natural biphenyl, inhibits in vitro and in vivo growth
of breast cancer through induction of apoptosis and cell cycle
arrest. Int J Oncol. 30:1529–1537. 2007.PubMed/NCBI
|
|
13
|
Sheu ML, Liu SH and Lan KH: Honokiol
induces calpain-mediated glucose-regulated protein-94 cleavage and
apoptosis in human gastric cancer cells and reduces tumor growth.
PLoS One. 2:e10962007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ishitsuka K, Hideshima T, Hamasaki M, et
al: Honokiol overcomes conventional drug resistance in human
multiple myeloma by induction of caspase-dependent and -independent
apoptosis. Blood. 106:1794–1800. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang SE, Hsieh MT, Tsai TH and Hsu SL:
Down-modulation of Bcl-XL, release of cytochrome c and sequential
activation of caspases during honokiol-induced apoptosis in human
squamous lung cancer CH27 cells. Biochem Pharmacol. 63:1641–1651.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shigemura K, Arbiser JL, Sun SY, et al:
Honokiol, a natural plant product, inhibits the bone metastatic
growth of human prostate cancer cells. Cancer. 109:1279–1289. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chu S and Ferro TJ: Sp1: regulation of
gene expression by phosphorylation. Gene. 348:1–11. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Deniaud E, Baguet J, Mathieu AL, Pages G,
Marvel J and Leverrier Y: Overexpression of Sp1 transcription
factor induces apoptosis. Oncogene. 25:7096–7105. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li L and Davie JR: The role of Sp1 and Sp3
in normal and cancer cell biology. Ann Anat. 192:275–283. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sankpal UT, Goodison S, Abdelrahim M and
Basha R: Targeting Sp1 transcription factors in prostate cancer
therapy. Med Chem. 7:518–525. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jutooru I, Chadalapaka G, Sreevalsan S, et
al: Arsenic trioxide downregulates specificity protein (Sp)
transcription factors and inhibits bladder cancer cell and tumor
growth. Exp Cell Res. 316:2174–2188. 2010. View Article : Google Scholar
|
|
22
|
Mann J: Natural products in cancer
chemotherapy: past, present and future. Nat Rev Cancer. 2:143–148.
2002. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park EJ, Min HY, Chung HJ, et al:
Down-regulation of c-Src/EGFR-mediated signaling activation is
involved in the honokiol-induced cell cycle arrest and apoptosis in
MDA-MB-231 human breast cancer cells. Cancer Lett. 277:133–140.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang T, Chen F, Chen Z, et al: Honokiol
induces apoptosis through p53-independent pathway in human
colorectal cell line RKO. World J Gastroenterol. 10:2205–2208.
2004.PubMed/NCBI
|
|
25
|
Arora S, Bhardwaj A, Srivastava SK, et al:
Honokiol arrests cell cycle, induces apoptosis, and potentiates the
cytotoxic effect of gemcitabine in human pancreatic cancer cells.
PLoS One. 6:e215732011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ishikawa C, Arbiser JL and Mori N:
Honokiol induces cell cycle arrest and apoptosis via inhibition of
survival signals in adult T-cell leukemia. Biochim Biophys Acta.
1820:879–887. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rajendran P, Li F, Shanmugam MK, et al:
Honokiol inhibits signal transducer and activator of
transcription-3 signaling, proliferation, and survival of
hepatocellular carcinoma cells via the protein tyrosine phosphatase
SHP-1. J Cell Physiol. 227:2184–2195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Riss TL and Moravec RA: Use of multiple
assay endpoints to investigate the effects of incubation time, dose
of toxin, and plating density in cell-based cytotoxicity assays.
Assay Drug Dev Technol. 2:51–62. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Y, Huang W, Huang S, Du J and Huang C:
Screening of anti-cancer agent using zebrafish: comparison with the
MTT assay. Biochem Biophys Res Commun. 422:85–90. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chiefari E, Brunetti A, Arturi F, et al:
Increased expression of AP2 and Sp1 transcription factors in human
thyroid tumors: a role in NIS expression regulation? BMC Cancer.
2:352002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hosoi Y, Watanabe T, Nakagawa K, et al:
Up-regulation of DNA-dependent protein kinase activity and Sp1 in
colorectal cancer. Int J Oncol. 25:461–468. 2004.PubMed/NCBI
|
|
32
|
Wang L, Wei D, Huang S, et al:
Transcription factor Sp1 expression is a significant predictor of
survival in human gastric cancer. Clin Cancer Res. 9:6371–6380.
2003.PubMed/NCBI
|
|
33
|
Yao JC, Wang L, Wei D, et al: Association
between expression of transcription factor Sp1 and increased
vascular endothelial growth factor expression, advanced stage, and
poor survival in patients with resected gastric cancer. Clin Cancer
Res. 10:4109–4117. 2004. View Article : Google Scholar
|
|
34
|
Zannetti A, Del Vecchio S, Carriero MV, et
al: Coordinate up-regulation of Sp1 DNA-binding activity and
urokinase receptor expression in breast carcinoma. Cancer Res.
60:1546–1551. 2000.PubMed/NCBI
|
|
35
|
Davie JR, He S, Li L, et al: Nuclear
organization and chromatin dynamics - Sp1, Sp3 and histone
deacetylases. Adv Enzyme Regul. 48:189–208. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee KA, Lee YJ, Ban JO, et al: The
flavonoid resveratrol suppresses growth of human malignant pleural
mesothelioma cells through direct inhibition of specificity protein
1. Int J Mol Med. 30:21–27. 2012.
|
|
37
|
Culver C, Melvin A, Mudie S and Rocha S:
HIF-1α depletion results in SP1-mediated cell cycle disruption and
alters the cellular response to chemotherapeutic drugs. Cell Cycle.
10:1249–1260. 2011.
|
|
38
|
Blume SW, Snyder RC, Ray R, Thomas S,
Koller CA and Miller DM: Mithramycin inhibits SP1 binding and
selectively inhibits transcriptional activity of the dihydrofolate
reductase gene in vitro and in vivo. J Clin Invest. 88:1613–1621.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chintharlapalli S, Papineni S, Lei P,
Pathi S and Safe S: Betulinic acid inhibits colon cancer cell and
tumor growth and induces proteasome-dependent and -independent
downregulation of specificity proteins (Sp) transcription factors.
BMC Cancer. 11:3712011. View Article : Google Scholar
|
|
40
|
Pietrzak M and Puzianowska-Kuznicka M:
p53-dependent repression of the human MCL-1 gene encoding an
anti-apoptotic member of the BCL-2 family: the role of Sp1 and of
basic transcription factor binding sites in the MCL-1 promoter.
Biol Chem. 389:383–393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tapias A, Ciudad CJ, Roninson IB and Noe
V: Regulation of Sp1 by cell cycle-related proteins. Cell Cycle.
7:2856–2867. 2008. View Article : Google Scholar : PubMed/NCBI
|