Introduction
Bone is the most common site of prostate cancer
(PCa) metastasis and once tumors metastasize to bone, they are
essentially incurable and result in significant morbidity prior to
patient mortality (1,2). Therefore, to develop more effective
therapeutics, a more thorough understanding of the key complex
processes that are central to metastasis may reveal more robust and
less redundant therapeutic targets.
Neovascularization is essential for tumor growth and
it also facilitates tumor invasion and metastasis (3). Furthermore, in cancer patients with
metastasis, neovascularization-mediated progression of
micrometastasis to lethal macrometastasis is the major cause of
mortality (4). Emerging evidence
implicates hypoxia as a key inducer of neovascularization in tumors
(5). Moreover, hypoxia-induced
pathological neovascularization mediates tumor cell dissemination,
invasion and metastasis in an animal model (6). However, the mechanisms underlying
pathological neovascularization in relation to hypoxia in tumor
invasion and metastasis remain elusive.
The process of hypoxia-induced neovascularization
was formerly attributed to the migration and proliferation of
pre-existing, fully differentiated endothelial cells (ECs), known
as angiogenesis (7). However, it is
now well established that tumors can acquire their vasculature by
various mechanisms including postnatal vasculogenesis, a process
during which circulating bone-marrow-derived endothelial progenitor
cells (BM-EPCs) home to sites of neovascularization and
differentiate into ECs (8).
Although there has been controversy concerning the time during
neovascularization at which the proposed contribution of EPCs to
neovascular vessels might occur (9), it has been proposed that the
recruitment of BM-EPCs is pivotal for tumor vasculogenesis
(10–12). Notably, recent studies have shown
that BM-EPCs are recruited to the angiogenic switch in tumor growth
and metastatic progression (4,13,14)
and blood levels of EPCs tend to increase in cancer patients and
correlate with the stage of the malignant disease (15). In prostate cancer, although
microvessel density is related to clinical stage, progression,
metastasis and survival (16–18),
it is not known whether EPCs contribute to the bone metastasis of
PCa.
The mobilization, homing and incorporation of EPCs
into tumors are multi-step and multi-factor events during tumor
vasculogenesis. This complex process requires the participation of
numerous cytokines. A series of studies have shown that several
factors, such as VEGF (20), SDF-1
(20), G-CSF (21), GM-CSF (22), PIGF (23), IGF2 (24), E-selectin (25) and angiopoietin-2 (26), play an important role in inducing
EPC homing and vasculogenesis. Although it is not very clear how
circulating EPC home from the bone marrow specifically into the
tumor microenvironment, the mobilization of EPC from the bone
marrow into the bloodstream, migration to the tumor site, and
subsequent integration into the vascular network, are all
presumably controlled by tumor-derived factors (11). However, in PCa progression, it is
not clear whether the above-mentioned tumor-derived factors are
involved in the regulation of EPC migration and vasculogenesis in
hypoxia, and whether other cytokines play roles in these processes.
Since a recent study found that PCa cells which metastasize to bone
competed with haematopoietic stem cells for the niche and colonize
in the niche (27) and
haematopoietic stem cell niches within the bone marrow are hypoxic
(28,29), we speculated that PCa cells in the
hypoxic microenvironment could recruit BM-EPCs and induce
vasculogenesis by releasing hypoxia-induced cytokines.
In this in vitro study, we found that
conditioned medium (CM) of PC-3 cells, derived from bone metastasis
of PCa, by hypoxia for 24 h promoted proliferation and migration,
and augmented the vasculogenesis capacity of BM-EPCs. Human
cytokine antibody array revealed that under early hypoxia, the PC-3
cells secreted several cytokines involved in BM-EPC functions,
including proliferation, migration and vasculogenesis. These
findings suggest that PCa cells may have the potential to modulate
their microenvironment and facilitate BM-EPC migration and
vasculogenesis in the early stage of hypoxia.
Materials and methods
Cell culture and hypoxic treatment
PC-3 cells were purchased from the American Type
Culture Collection (ATCC, Manassas, VA, USA) and maintained in F-12
culture medium (HyClone) supplemented with 10% fetal bovine serum
(HyClone). For normoxia experiments, the PC-3 cells were cultured
at 37°C with 5% CO2/95% air in a humidified incubator,
and for hypoxia experiments, the PC-3 cells were cultured at 37°C
with 5% CO2, 94% N2, and 1% O2 in
a multigas incubator (Juji Field, Tokyo, Japan).
Conditioned medium collection
PC-3 cells were cultured in hypoxia and normoxia for
24 and 48 h and CM was collected. Briefly, PC-3 cells were seeded
at a density of 2.5×105 cells/ml in F12 (HyClone)
complete media into a 10-cm cell culture dish and grown to ~80%
confluency. Cells were washed twice with phosphate-buffered saline
(PBS) and incubated in serum-free medium for 24 and 48 h. CM was
collected and centrifuged at 2,000 × g at 4°C for 10 min to remove
cell debris and the supernatant was used.
Isolation and cultivation of BM-EPCs
Bone marrow was collected from male patients with
lumbar degenerative diseases (age range, 27–72 years; mean age
52.4±13.9 years; disc herniation with lumbar spondylolisthesis,
degenerative lumbar spondylolisthesis and lumbar spinal stenosis
with instability) at the time of operation. Informed consent was
obtained from the patients for bone marrow blood collection, and
all procedures were performed in accordance with the guidance and
approval of a research Ethics Committee at the First Affiliated
Hospital of Sun Yat-sen University.
Mononuclear cells were collected by density gradient
centrifugation using Ficoll-Paque™ Premium (1.077; GE Healthcare,
San Francisco, CA, USA) according to the manufacturer’s
instructions (30). Briefly, the
isolated cells were cultivated in dishes coated with fibronectin
and induced by EGM-2 MV Single Quots (Cambrex) at 37°C with 5%
CO2/95% air in a humidified incubator at a density of
3–5×106/cm2. After 3 days in culture,
non-adherent cells were removed by washing with new medium and
changed. After 7 days in culture, BM-EPCs were identified by
immunofluorescence staining and flow cytometry. Quantitative
fluorescence-activated cell sorting (FACS) was performed on a
FACSVantage SE flow cytometer (Becton-Dickinson, San Jose, CA,
USA). The Weibel-Palade body in cultivation cells was visualized by
transmission electron microscope (TEM).
BM-EPC proliferation assay
EPC proliferation assay was performed to assess the
viability of BM-EPCs treated with CM of PC-3 cells. The effect of
CM on BM-EPC proliferation was determined by
2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium,
monosodium salt (WST-8) assay kit (CCK-8, Dojindo, Tokyo, Japan).
Briefly, BM-EPCs were plated in 96-well plates at a concentration
of 5×103 cells/well in 100 μl CM of PC-3 cells.
Following incubation for 36 h by CM, CCK-8 was used according to
the manufacturer’s instructions. WST-8 was added into each well for
4 h prior to the measurement. The absorbance at 450 nm was measured
using a microplate reader.
BM-EPC migration assay
Cell migration was evaluated using the Transwell
system (Corning Costar, Acton, MA, USA) with 6.5-mm diameter
polycarbonate filters (8-μm pore size). Briefly, BM-EPCs were
seeded onto chemotaxis filters in 100 μl F-12 medium. CM of PC-3
cells cultured under normoxia and hypoxia for 24 and 48 h was
collected and then added to the lower chamber with a normal medium
(NM; F-12) as control medium. After the 24-h migration period,
non-migrating cells were completely removed from the top surface of
the membrane and attached cells were fixed with 95% ethanol for 10
min and stained with 0.1% crystal violet for 25 min. The plate was
immersed in fresh tap water to remove excess dye. The adherent
cells were visualized under light microscopy at ×200 magnification
and quantified in triplicate by counting adherent cells in 5
randomly selected fields per well (Axiovert 100; Carl Zeiss
Microimaging, Thornwood, NY, USA). The results are representative
of 4 independent experiments.
In vitro BM-EPC vasculogenesis assay
In vitro EPC vasculogenesis assay was
performed with In Vitro Angiogenesis Assay kit (Chemicon, Temecula,
CA, USA) according to the manufacturer’s instructions. Briefly, EC
Matrix™ solution was thawed on ice overnight, mixed with 10X EC
Matrix diluents and placed in a 96-well tissue culture plate at
37°C for 1 h to allow the matrix solution to solidify. BM-EPCs were
plated in normoxic or hypoxic CM at a concentration of
1×104 cells/well on the top of the solidified matrix
solution (1% FBS was added to CM and NM). Tube formation was
inspected under an inverted light microscope at ×200 magnification.
Tube formation was defined as a structure exhibiting a length four
times its width. Five independent fields were assessed for each
well, and the average number of tubes/x200 field was
determined.
Human cytokine antibody array
Cytokine concentrations in CM were quantified by
using human cytokine antibody arrays. RayBio Human Cytokine
Antibody Array C Series 2000 kit was purchased from RayBiotech
(Norcross, GA, USA). This array consisted of 174 different
antibodies spotted in duplicate onto three membranes. CM was
collected from normoxic or hypoxic PC-3 cells for 24 and 48 h with
an NM (F-12) as control medium. Experiments were performed by
KangChen Bio-tech (Shanghai, China) as recommended by the
manufacturer. The signal intensities can be quantified by
densitometry, and relative expression levels of cytokines can be
acquired by comparing them. Positive control can be used to
normalize the results from different membranes being compared.
Gene Ontology analysis
The Gene Ontology (GO) project provides a controlled
vocabulary to describe gene and gene product attributes in any
organism (http://www.geneontology.org). The
ontology covers three domains: biological process, cellular
component and molecular function. Fisher’s exact test is used to
find if there is more overlap between the DE list and the GO
annotation list than would be expected by chance. The P-value
denotes the significance of GO terms enrichment in the DE genes.
The lower the P-value, the more significant the GO Term (P-value
≤0.05 is recommended). GOID, the ID of the Gene Ontology term used
in Gene Ontology Project. Term, the name of the gene Ontology Term.
Ontology, the ontology the GOID belongs to, i.e., ‘biological
process’, ‘cellular component’ or ‘molecular function’.
Statistical analysis
The experimental data are presented as the means of
each condition ± SEM. One-way ANOVA was performed for comparing
more than two groups, and paired Student’s t-test was performed
when comparing two conditions (SPSS for Windows® version
16.0). P<0.05 (*P<0.05, **P<0.01)
was considered to indicate a statistically significant
difference.
Results
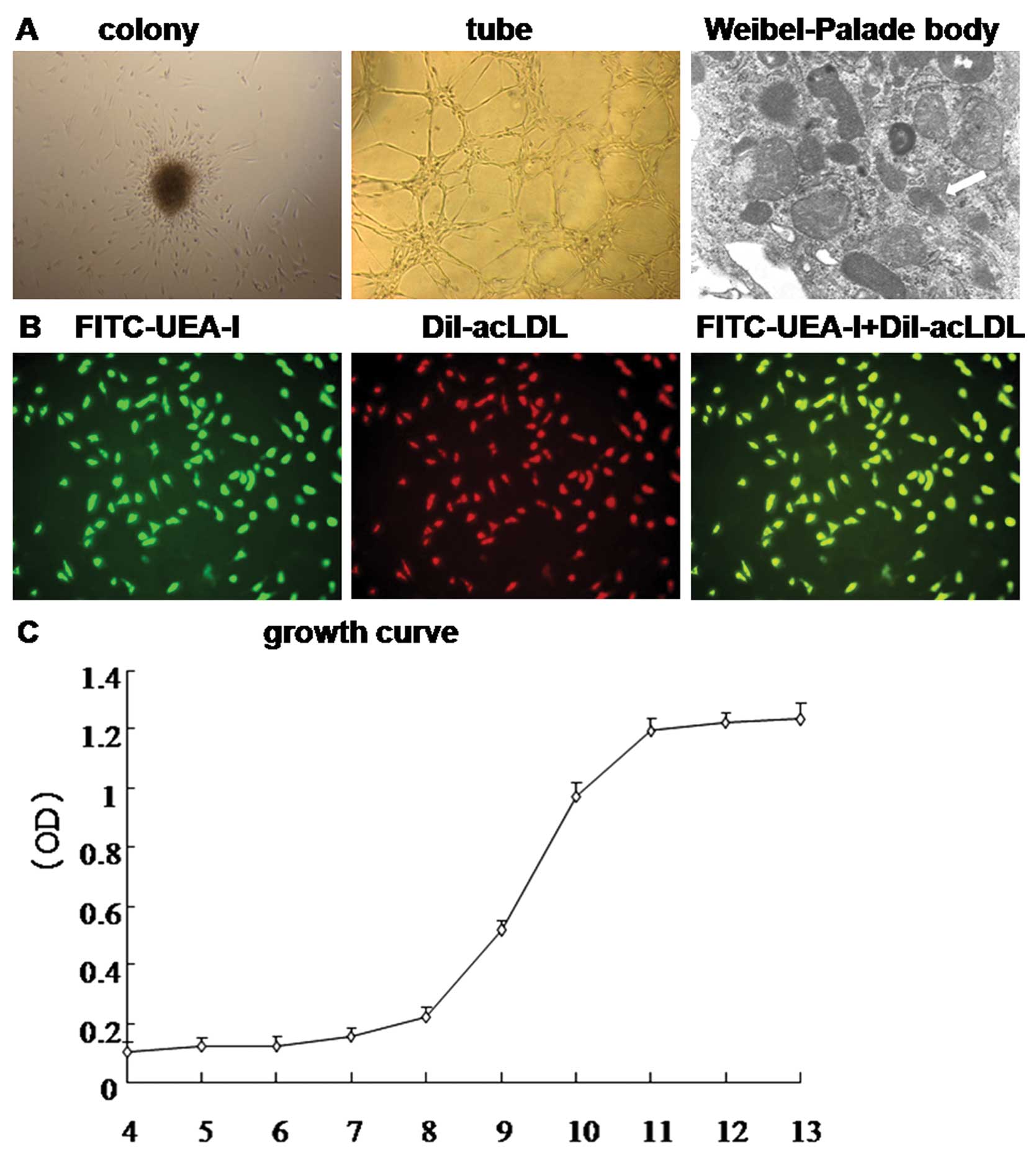
The growth processes of BM-EPCs
Mononuclear cells were isolated from human bone
marrow blood and cultivated in EGM2 on fibronectin-coated dishes.
Cells demonstrated round morphology with different sizes as
assessed by light microscopy during the first 3 days. With a
tendency of ~20% of the mononuclear cells transferred to culture
plates grew as adherent cells during 7 days of cultivation and
clusters or colonies formed. Furthermore, in endochylema under TEM,
the Weibel-Palade body was also observed and BM-EPCs had active
angiogenic potential (Fig. 1A).
Following cultivation for 7 days, the adherent cells exhibited
strong ability to uptake Dil-acLDL, and FITC-UEA-I and the double
positive rate was (94.3±4.1%) (Fig.
1B), which were identified as differentiating EPCs. The growth
curve is shown in Fig. 1C. The
percentage of expression of CD133, CD34, KDR, VE-cadherin,
E-selectin and vWF was determined by flow cytometry at Day 7 and
Day 14 of culture. The expression pattern of surface markers in
BM-EPCs changed toward a more mature endothelial cell phenotype
from Day 7 to Day 14 (Table I).
 | Table IThe expression pattern of surface
markers of BM-EPCs at Day 7 and Day 14. |
Table I
The expression pattern of surface
markers of BM-EPCs at Day 7 and Day 14.
| Time | CD133 | CD34 | KDR | VE-cadherin | E-selectin | vWF |
|---|
| Day 7 | 19.23±5.0 | 41.49±9.01 | 65.97±7.2 | 21.27±6.9 | 8.02±5.53 | 5.15±2.72 |
| Day14 | 4.04±3.52 | 32.94±8.37 | 82.57±6.2 | 25.23±4.4 | 45.08±7.77 | 30.3±7.81 |
| P-value | 0.013 | 0.295 | 0.039 | 0.448 | 0.003 | 0.002 |
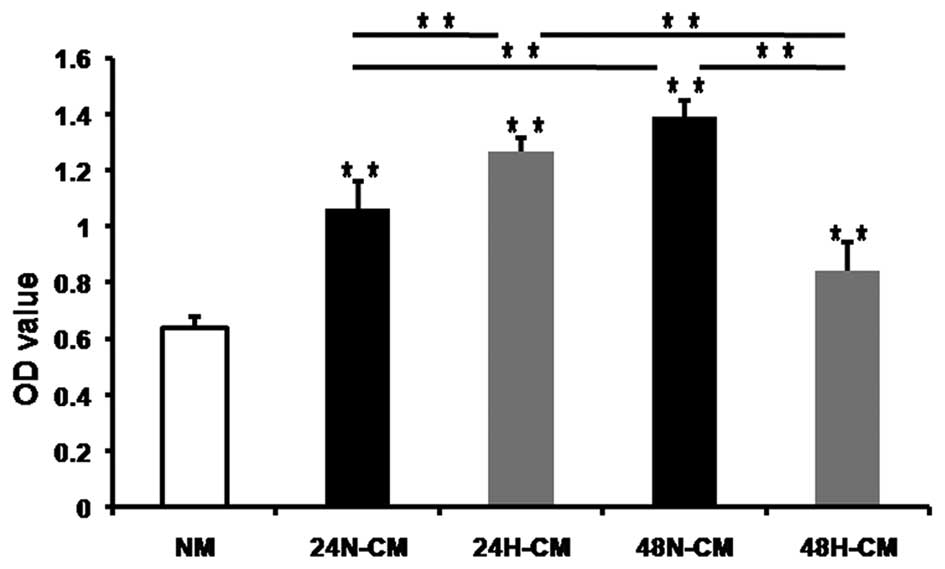
H-CM of PC-3 cells promotes proliferation
of BM-EPCs
The proliferation of EPCs, as a crucial step of
vasculogenesis, is triggered and modulated by a variety of stimuli
including hypoxic microenvironment. To demonstrate whether H-CM for
24 and 48 h promoted proliferation of BM-EPCs, the proliferation
assay was performed to assess the viability of BM-EPCs treated with
CM of tumor cells. WST-8 assay was used to examine the changes in
cell proliferation. We exposed BM-EPCs to 24 and 48 H-CM, 24/48
N-CM and NM as a control medium for 36 h. All CM induced BM-EPC
proliferation and 24H-CM promoted proliferation of BM-EPCs more
evidently compared to 24N-CM (Fig.
2). However, 48N-CM promoted proliferation of BM-EPCs more
apparently compared to 48H-CM. Furthermore, 24H-CM had stronger
effects on enhancing BM-EPC proliferation than 48H-CM. These
results suggested that hypoxia induced PC-3 cells to release some
factors which promoted the proliferation of BM-EPCs.
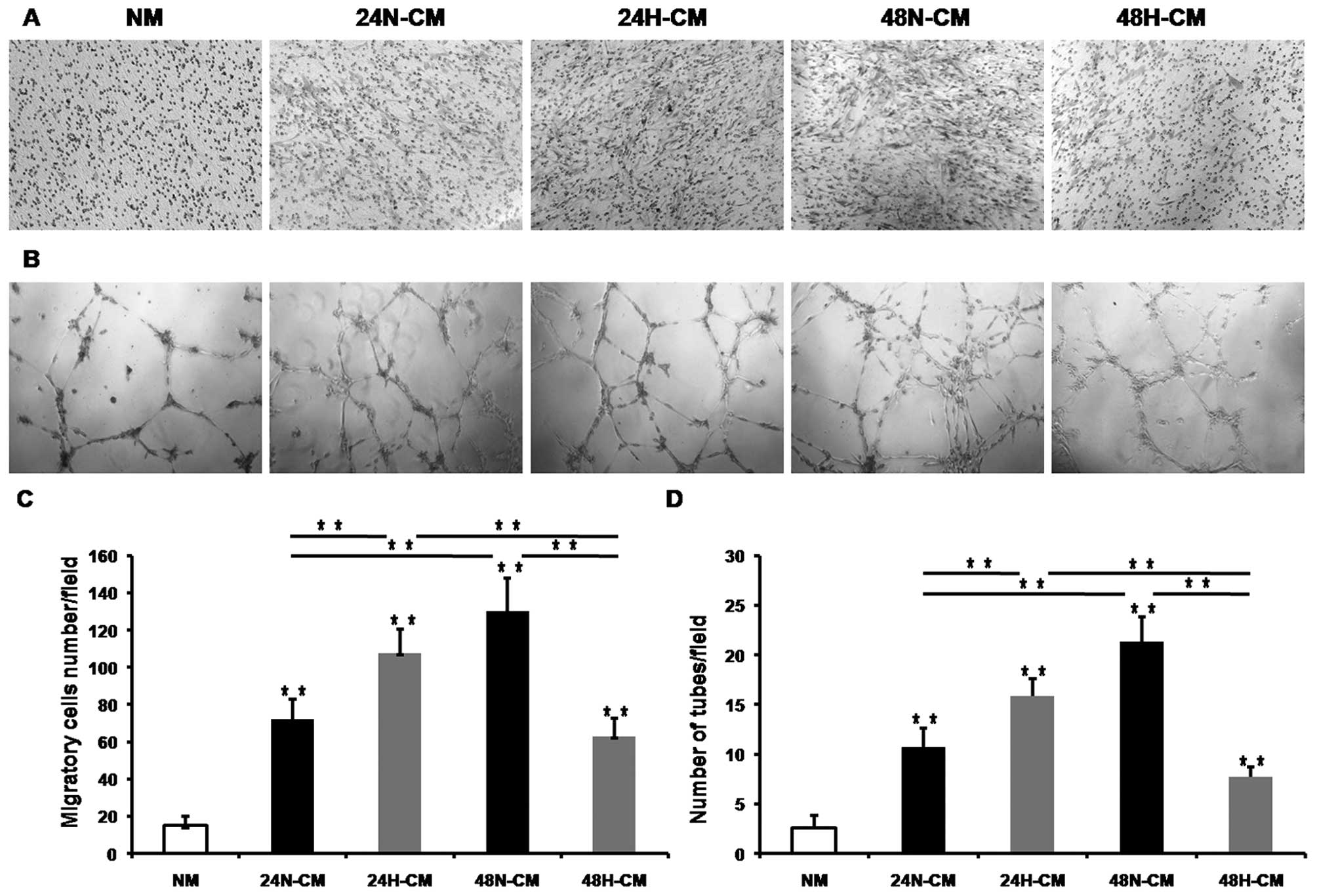
H-CM of PC-3 cells promotes migration of
BM-EPCs
The mobilization of EPC from the bone marrow into
the bloodstream, migration to the tumor site, and subsequent
integration into the vascular network are all presumably controlled
by tumor-derived factors and those factors are released to the
tumor microenvironment that is CM of tumor cells in vitro.
To determine if CM collected from hypoxia-treated PC-3 cells (H-CM)
for 24/48 h increased migration of BM-EPCs compared to CM collected
from normoxia-treated PC-3 cells (N-CM), we used a Transwell assay
to estimate the ability of EPCs to migrate. All CM enhanced BM-EPC
migration compared with NM as a control (Fig. 3A and C). Also, 24H-CM enhanced
migration of BM-EPCs more apparently than 24N-CM. However, 48N-CM
increased migration of BM-EPCs more evidently than 48H-CM.
Moreover, 24H-CM enhanced migration of BM-EPCs more apparently than
48H-CM. These observations suggested that hypoxia induced PC-3
cells to release some factors, which promoted the migration of
BM-EPCs.
H-CM of PC-3 cells augments in vitro
vasculogenesis of BM-EPCs
One of the most prominent effects of hypoxia is the
induction of tumor neovascularization (5). To determine whether CM of PC-3 cells
augmented the vasculogenesis of BM-EPCs in vitro, we exposed
BM-EPCs to 24/48H-CM, 24/48N-CM and NM as a control medium for 18 h
on Matrigel. All CM induced BM-EPC tube formation (Fig. 3B and D). The 24H-CM augmented the
tube formation of BM-EPCs more evidently than 24N-CM. However,
48N-CM augmented the tube formation of BM-EPCs more apparently than
48H-CM. At the same time, 24H-CM had stronger effects on enhancing
BM-EPC tube formation than 48H-CM. These observations indicated
that hypoxia induced PC-3 cells to release some factors which
augmented the in vitro vasculogenesis capacity of
BM-EPCs.
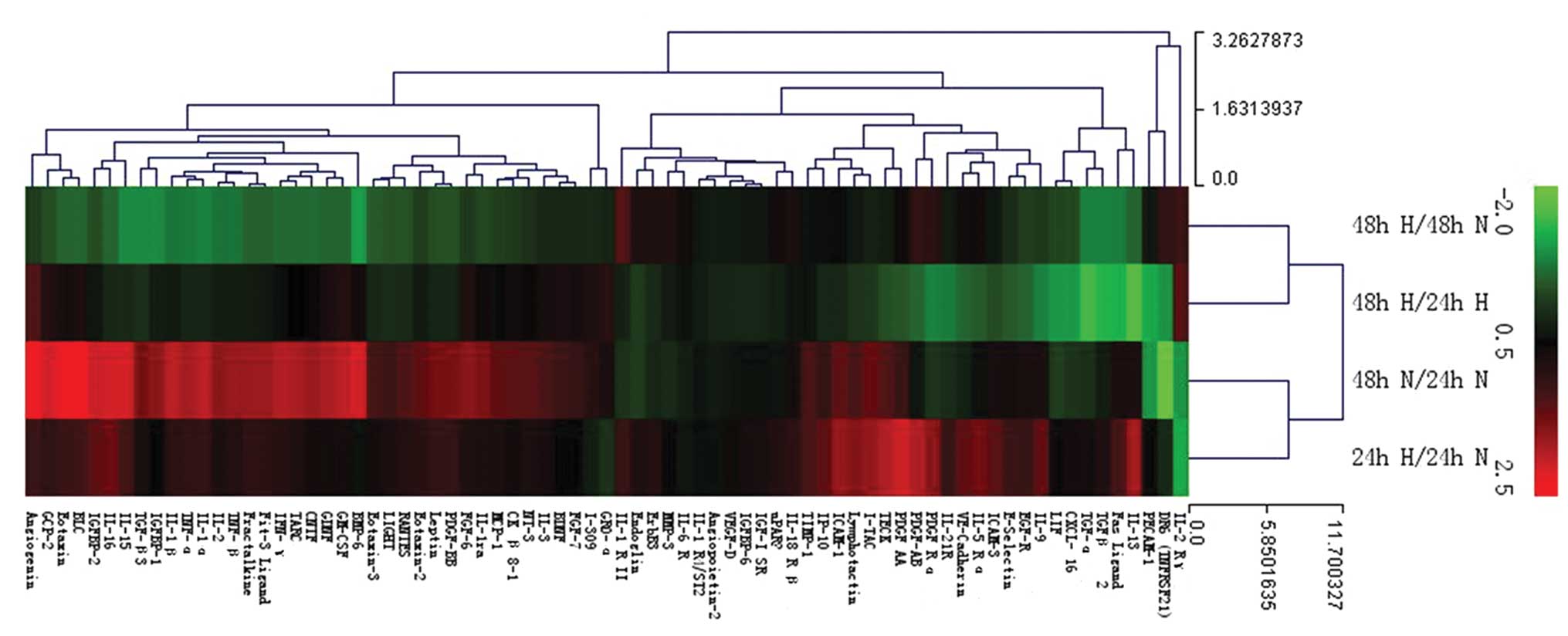
Significant alterations of the cytokines
in H-CM
In order to investigate tumor-derived cytokines of
PC-3 cells which effect the functions of BM-EPCs, we used human
cytokine antibody array to reveal the changes of cytokines in H-CM
compared with N-CM. The cytokine profile for each of the CM was
performed using three independent biological samples on protein
arrays printed with 174 anti-cytokine antibodies in replicate.
Relative expression levels of cytokines could be acquired by
comparing the signal intensities. Significantly differentially
expressed cytokines were selected as following standards:
fold-change >1.3-fold and P<0.05 in H-CM compared with N-CM,
and 24H-CM compared with 48H-CM. The results of cluster analysis
are shown in Fig. 4. Compared with
24N-CM, most cytokines in 24H-CM were upregulated, but compared
with 48N-CM, most cytokines in 48H-CM were downregulated. Compared
with 24N-CM, most cytokines in 48N-CM were upregulated, however,
compared with 24H-CM, most cytokines in 48H-CM were downregulated.
These data indicated that in the early stage, hypoxia induced
expression of most cytokines in PC-3 cells, but in the later stage,
hypoxia repressed expression of several cytokines.
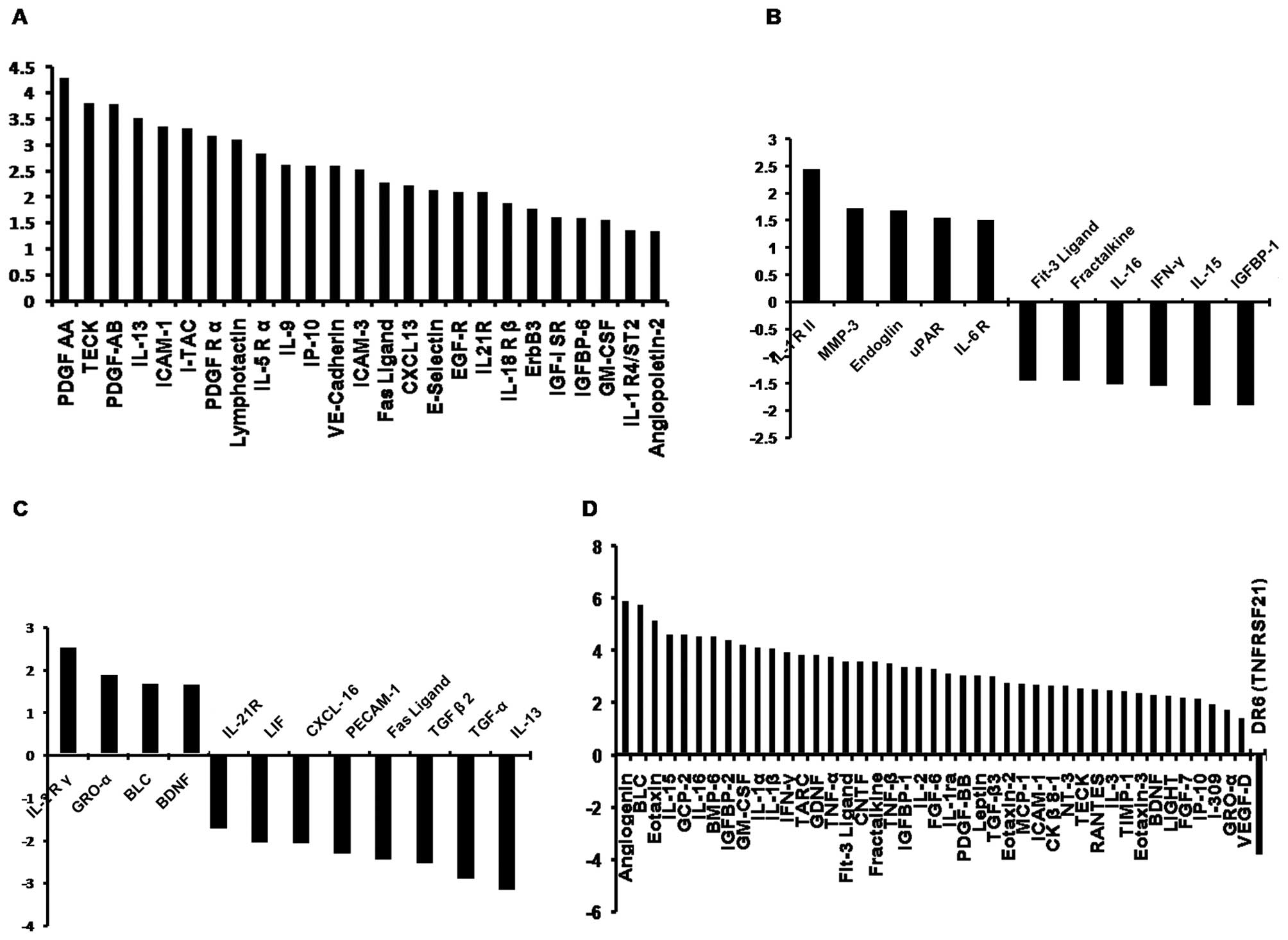
Twenty-five types of cytokines were significantly
different between 24H-CM and 24N-CM, and all these factors
increased in 24H-CM (Fig. 5A).
Eleven types of cytokines were significantly different between
48H-CM and 48N-CM (Fig. 5B), and,
among them, only 5 factors increased and 6 decreased in 48H-CM.
Twelve types of cytokines were significantly different between
48H-CM and 24H-CM, and, among them, 4 factors increased and 8
decreased in 48H-CM (Fig. 5C).
Forty-four types of cytokines were significantly different between
24N-CM and 48N-CM, and, among them, 43 factors increased in 48N-CM
(Fig. 5D).
Some cytokines increased the expression in 24H-CM
compared with that in 24N-CM, but these changes were not
statistically significant, which play an important role in inducing
EPCs homing and vasculogenesis according to the results in
literature (20,23,24),
including VEGF (fold-change=2.47, P=0.083), SDF-1
(fold-change=1.43, P=0.429), IGF-II (fold-change=1.84, P=0.053),
PIGF (fold-change=1.79, P=0.125).
Functions of the cytokines with
significant alterations in H-CM are involved in proliferation,
migration and vasculogenesis
Differentially expressed cytokines selected as
standards were analyzed by GO analysis. The GO project provides a
controlled vocabulary to describe gene and gene product attributes
in any organism (http://www.geneontology.org, GO Result, Fisher’s exact
method, P-value ≤0.05). Therefore, we know the cytokines which may
be involved in the change on functions of BM-EPCs by PC-3 CM
between hypoxia and normoxia. ‘Biological process’ and ‘molecular
function’ of those cytokines involved were analyzed and the terms
were found, including positive and negative regulation of cell
proliferation, chemotaxis, cell mobility, cell migration,
vasculogenesis, positive and negative regulation of angiogenesis,
angiogenesis, which are involved in the proliferation, migration
and tube formation in vitro. GO analysis results showed that
several cytokines are involved in proliferation, migration,
however, a few cytokines were involved in vasculogenesis and
angiogenesis (Table II).
| Table IIGene Ontology analysis results of
differentially expressed cytokines in conditioned media which
related with proliferation, migration and angiogenesis. |
Table II
Gene Ontology analysis results of
differentially expressed cytokines in conditioned media which
related with proliferation, migration and angiogenesis.
| 24H/24N | 48H/48N | 48H/24H | 48N/24N |
|---|
| Positive regulation
of cell proliferation (GO:0008284) | Upregulated CSF2,
CXCL10, EGFR, FASLG, IGF1, IL9, IL13, PDGFA, PDGFB, PDGFRA | Upregulated
IL6R
Downregulated IFNG, IL15 | Downregulated
FASLG, IL13, LIF, TGFA, TGFB2 | Upregulated ANG,
CCL2, CCL5, CCL11, CCL24, CCL26, CNTF, CSF2, CXCL10, FGF7, FGF6,
FIGF, IFNG, IL1B, IL2, IL3, IL15, PDGFB, TIMP1, TNF |
| Negative regulation
of cell proliferation (GO:0008285) | | Upregulated
ENG
Downregulated FNG, IL15 | Upregulated
CXCL1
Downregulated LIF, TGFB2 | Upregulated ANG,
CCL23, CXCL1, IFNG, IL1A IL1B, IL2, IL15, LTA, TGFB3, TNF |
| Chemotaxis
(GO:0006935) | Upregulated ANGPT2,
CCL25, CXCL10, PDGFA, PDGFB, XCL1 | Upregulated ENG,
IL6R, PLAUR
Downregulated IL16, CX3CL1 | Upregulated CXCL1,
CXCL13
Downregulated CXCL16, TGFB2 | Upregulated CCL1,
CCL2, CCL5, CCL11, CCL17, CXCL11, CXCL13, EGFR, CCL23, CCL24, CL25,
CCL26, CXCL1, CXCL6, CXCL10, CXCL13, CX3CL1, FGF7, FIGF, GDNF, IL1B
IL16, NTF3, PDGFB |
Cell migration
(GO:0016477)
Cell motility (GO:0048870) | Upregulated ANGPT2,
CCL25, EGFR, ICAM1, IGF1, PDGFA, PDGFB, PDGFRA, SELE | Upregulated ENG,
IL6R
Downregulated CX3CL1, IL16 | Downregulated
CXCL16, ECAM1, TGFB2 | Upregulated ANG,
CCL2, CCL5, CCL11, CCL24, CCL25, CCL26, CX3CL1, FGF7, FIGF, GDNF,
ICAM1, IL1B, IL16, PDGFB, TNF |
| Vasculogenesis
(GO:0001570) | | Upregulated
ENG | | |
| Positive regulation
of angiogenesis (GO:0045766) | | Downregulated
CX3CL1 | | Upregulated IL1A,
IL1B, CCL11, CCL24, CX3CL1 |
| Negative regulation
of angiogenesis (GO:0016525) | | | Downregulated
FASLG, LIF | |
| Angiogenesis
(GO:0001525) | | Upregulated
ENG
Downregulated CX3CL1 | Downregulated
FASLG, LIF, TGFB2 | Upregulated ANG,
CCL2, CCL11, CCL24, CX3CL1, FGF6, FIGF, IL1A, IL1B |
Discussion
In the present study, we demonstrated that 24H-CM of
PC-3 cells of PCa presented stronger abilities of promoting
proliferation and migration, and augmenting in vitro
vasculogenesis capacity of BM-EPCs compared to 24N-CM. Moreover,
24H-CM had stronger effects than 48H-CM. At the same time, our
results also demonstrated that hypoxia for 24 h induced several
types of cytokines significantly increasing expression in CM of
PC-3 cells. However, downregulated factors were more than
upregulated factors in 48H-CM compared with 48N-CM or 24H-CM.
Furthermore, according to the Gene Ontology analysis, all those
changed cytokines were involved in the regulation of cell
proliferation, chemotaxis, cell motility, cell migration,
vasculogenesis and angiogenesis. The changed regularity of
cytokines in the 24H-CM and 48H-CM of PC-3 cells was in concert
with the functional changes of BM-EPCs treated with different CM of
PC-3 cells in enhancing the proliferation, migration and
vasculogenesis potential of BM-EPCs. These data suggest that PCa
cells may have the potential to modulate their microenvironment and
facilitate BM-EPC migration and vasculogenesis in the early stage
of hypoxia.
Our results demonstrated that the function of the
majority of the hypoxia-induced cytokines released by PCa cells
from bone metastasis in early stage was to promote chemotaxis, cell
motility and cell migration. In vivo, as PCa cells colonize
in the haematopoietic stem cell niches which are hypoxic (27–29),
once into the niches, PCa cells may meet up with the hypoxic bone
microenvironment. Those cytokines released by PCa cells in the
hypoxic bone microenvironment may recruit BM-EPCs to the
parasitizing focus and induce vasculogenesis. Gao et
al(4) used mouse models of
pulmonary metastasis and identified BM-EPCs as critical regulators
of this angiogenic switch. Animal studies showed that BM-EPCs
played a role in vasculogenesis within 48 h after tumor
transplantation (31,32). A kinetic analysis of EPC
contribution as a function of tumor growth showed that EPCs are
recruited to early tumors preceding vessel formation, followed by
differentiation into endothelial cells and luminal incorporation
into a subset of sprouting tumor neovessels (12). These findings suggested that BM-EPCs
might contribute to the development of micrometastasis of PCa in
bone marrow in the early stage of colonization of metastatic PCa
cells.
The migration of EPCs induced by cytokines released
from tumor cells is one of the key mechanisms in the process of
vascular development of hypoxia-induced vasculogenesis and one of
the rate-limiting steps in the homing of EPCs (10–12).
Several studies have demonstrated that factors such as VEGF
(20), SDF-1 (20), G-CSF (21), PIGF (23) and IGF2 (24) are pivotal in the regulation of
homing of EPCs. In the present study, the results showed that
hypoxia did not induce statistically significant high expressions
of the above-mentioned cytokines in PC-3 cells. However, the
expression levels of GM-CSF, angiopoietin-2, PDGF-AB, and
E-selectin were induced significantly by 24-h hypoxia and 2 types
of cytokines, endoglin (ENG) and IL6, in 48-h hypoxia. Previous
studies have demonstrated that GM-CSF (22), E-selectin (25), angiopoietin-2 (26), PDGF-AB (33), ENG (34) and IL6 (5) are the important factors for the
hypoxia-induced homing of BM-EPCs in the vasculogenesis process.
Therefore, we speculated that these cytokines might play a more
important role in the hypoxia-induced homing of EPCs in PCa bone
metastasis compared to VEGF, SDF-1, G-CSF, PIGF and IGF2. We also
found that the expression levels of several cytokines significantly
upregulated, including CXCL13 (BLC), CXCL10 (IP-10), CCL25 (TECK),
CXCL11 (I-TAC), lymphotactin (XCL1), ICAM1, EGFR, IGF1R in CM of
PC-3 cells by 24-h hypoxia. These cytokines are involved in
chemotaxis, cell motility and cell migration according to the gene
ontology analysis. Some of them have also been proved to be
involved in EPC homing, such as ICAM1 (36,37),
IGF1R (38). Those cytokines may be
other factors that recruit EPCs to the tumor. Therefore, EPC homing
might be a combined effect of several cytokines in the
vasculogenesis process of bone metastasis of PCa.
EPCs that are incorporated into the tumor
vasculature could constitute as much as 50% (39), but it is not known if the
proliferating capabilities of EPCs are controlled by the tumor
microenvironment factors. In the present study, the results showed
that CM of PC-3 cells by 24-h hypoxia promoted EPC proliferation
and hypoxia significantly induced PC-3 cells releasing several
cytokines of positive regulation of cell proliferation. According
to the Gene Ontology analysis, those cytokines included GM-CSF,
IL5, IL13, EGFR, IGF1R, IL9, CXCL10 in 24-h hypoxia, and IFNG,
IL15, IL6 in 48-h hypoxia, among them, GM-CSF (40) was proved to have the function of
promoting proliferation of EPCs. Some of those cytokines have also
been reported to be involved in EPC homing, such as GM-CSF
(22), IL6 (35) and IGF1R (38). Therefore, the proliferating
capabilities of EPCs may be controlled by the tumor
microenvironment.
Our results showed that 24H-CM of PC-3 cells
augmented the in vitro vasculogenesis capacity of BM-EPCs
and hypoxia significantly induced PC-3 cells releasing
angiopoietin-2. Since angiopoietin-2 was proved to promote
vasculogenesis of EPC (41), it was
suggested that angiopoietin-2 might play an important role in the
vasculogenesis of BM-EPCs in PCa progression. The results also
showed that ENG and IL6 significantly increased in 48H-CM of PC-3
cells and ENG (34) and IL6
(35) have been proved to be
involved in vasculogenesis. It was suggested that ENG and IL6 might
play a role in vasculogenesis in 48-h hypoxia.
In conclusion, our study detected 174 cytokines in
CM of PC-3 cells from bone metastasis of PCa and found that 24-h
hypoxia significantly induced the secretion of several cytokines.
Those released cytokines in the CM of PC-3 cells by 24-h hypoxia
promoted proliferation and migration and augmented vasculogenesis
in vitro of BM-EPCs. Our data suggested that PCa cells may
have the potential to modulate their microenvironment and
facilitate BM-EPC migration and vasculogenesis in the early stage
of hypoxia and BM-EPCs might contribute to the development of
micrometastasis of PCa in bone marrow in the early stage of
colonization of metastatic PCa cells. However, aside from the
cytokines found in this study, other cytokines may be involved in
these processes. Furthermore, it also is necessary to detect the
functions of these cytokines in future studies. The
characterization of tumor-associated BM-EPCs may provide valuable
insight for more specific anti-angiogenesis therapy and/or tumor
diagnosis.
Acknowledgements
The authors thank Dr Wenjian Wang and Dr Wen Li from
the Surgical Laboratory at the First Affiliated Hospital of Sun
Yat-sen University for their help. We also thank the NSFC-Guangdong
Joint funding, China (no. u0732001); the Science and Technology
planning project of Guangdong Province, China (no. 2008B030301037),
the Science and Technology Planning Project of Guangzhou, China
(11C22060772).
Abbreviations:
|
PCa
|
prostate cancer
|
|
Hx
|
hypoxia
|
|
Nx
|
normoxia
|
|
BM-EPC
|
bone marrow-derived endothelial
progenitor cell
|
|
CM
|
conditioned medium
|
|
H-CM
|
hypoxic conditioned medium
|
|
N-CM
|
normoxic conditioned medium
|
|
NM
|
normal medium
|
|
24N
|
24-h normoxia
|
|
24H
|
24-h hypoxia
|
|
48N
|
48-h normoxia
|
|
48H
|
48-h hypoxia
|
References
|
1
|
Roodman GD: Mechanisms of bone metastasis.
N Engl J Med. 350:1655–1664. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mundy GR: Metastasis to bone: causes,
consequences and therapeutic opportunities. Nat Rev Cancer.
2:584–593. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gao D, Nolan DJ, Mellick AS, Bambino K,
McDonnell K and Mittal V: Endothelial progenitor cells control the
angiogenic switch in mouse lung metastasis. Science. 319:195–198.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liao D and Johnson RS: Hypoxia: a key
regulator of angiogenesis in cancer. Cancer Metastasis Rev.
26:281–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee SL, Rouhi P, Dahl Jensen L, et al:
Hypoxia-induced pathological angiogenesis mediates tumor cell
dissemination, invasion, and metastasis in a zebrafish tumor model.
Proc Natl Acad Sci USA. 106:19485–19490. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Senger DR and Davis GE: Angiogenesis. Cold
Spring Harb Perspect Biol. 3:a0050902011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Asahara T and Kawamoto A: Endothelial
progenitor cells for postnatal vasculogenesis. Am J Physiol Cell
Physiol. 287:C572–C579. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wickersheim A, Kerber M, de Miguel LS,
Plate KH and Machein MR: Endothelial progenitor cells do not
contribute to tumor endothelium in primary and metastatic tumors.
Int J Cancer. 125:1771–1777. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
De Palma M and Naldini L: Role of
haematopoietic cells and endothelial progenitors in tumour
angiogenesis. Biochim Biophys Acta. 1766:159–166. 2006.PubMed/NCBI
|
|
11
|
Spring H, Schüler T, Arnold B, Hämmerling
GJ and Ganss R: Chemokines direct endothelial progenitors into
tumor neovessels. Proc Natl Acad Sci USA. 102:18111–18116. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nolan DJ, Ciarrocchi A, Mellick AS, et al:
Bone marrow-derived endothelial progenitor cells are a major
determinant of nascent tumor neovascularization. Genes Dev.
21:1546–1558. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gao D, Nolan D, McDonnell K, Vahdat L,
Benezra R, Altorki N and Mittal V: Bone marrow-derived endothelial
progenitor cells contribute to the angiogenic switch in tumor
growth and metastatic progression. Biochim Biophys Acta.
1796:33–40. 2009.PubMed/NCBI
|
|
14
|
Martin-Padura I, Gregato G, Marighetti P,
et al: The white adipose tissue used in lipotransfer procedures is
a rich reservoir of CD34+ progenitors able to promote
cancer progression. Cancer Res. 72:325–334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Döme B, Hendrix MJ, Paku S, Tóvári J and
Tímár J: Alternative vascularization mechanisms in cancer:
pathology and therapeutic implications. Am J Pathol. 170:1–15.
2007.PubMed/NCBI
|
|
16
|
Strohmeyer D, Rössing C, Strauss F,
Bauerfeind A, Kaufmann O and Loening S: Tumor angiogenesis is
associated with progression after radical prostatectomy in pT2/pT3
prostate cancer. Prostate. 42:26–33. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Revelos K, Petraki C, Scorilas A, et al:
Correlation of androgen receptor status, neuroendocrine
differentiation and angiogenesis with time-to-biochemical failure
after radical prostatectomy in clinically localized prostate
cancer. Anticancer Res. 27:3651–3660. 2007.
|
|
18
|
Lissbrant IF, Stattin P, Damber JE and
Bergh A: Vascular density is a predictor of cancer-specific
survival in prostatic carcinoma. Prostate. 33:38–45. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Asahara T, Takahashi T, Masuda H, et al:
VEGF contributes to postnatal neovascularization by mobilizing bone
marrow-derived endothelial progenitor cells. EMBO J. 18:3964–3972.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Orimo A, Gupta PB, Sgroi DC, et al:
Stromal fibroblasts present in invasive human breast carcinomas
promote tumor growth and angiogenesis through elevated SDF-1/CXCL12
secretion. Cell. 121:335–348. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Powell TM, Paul JD, Hill JM, et al:
Granulocyte colony-stimulating factor mobilizes functional
endothelial progenitor cells in patients with coronary artery
disease. Arterioscler Thromb Vasc Biol. 25:296–301. 2005.
View Article : Google Scholar
|
|
22
|
Takahashi T, Kalka C, Masuda H, et al:
Ischemia- and cytokine-induced mobilization of bone marrow-derived
endothelial progenitor cells for neovascularization. Nat Med.
5:434–438. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li B, Sharpe EE, Maupin AB, Teleron AA,
Pyle AL, Carmeliet P and Young PP: VEGF and PlGF promote adult
vasculogenesis by enhancing EPC recruitment and vessel formation at
the site of tumor neovascularization. FASEB J. 20:1495–1497. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maeng YS, Choi HJ, Kwon JY, et al:
Endothelial progenitor cell homing: prominent role of the
IGF2-IGF2R-PLCβ2 axis. Blood. 113:233–243. 2009.PubMed/NCBI
|
|
25
|
Liu ZJ, Tian R, Li Y, An W, Zhuge Y,
Livingstone AS and Velazquez OC: Inhibition of tumor angiogenesis
and melanoma growth by targeting vascular E-selectin. Ann Surg.
254:450–457. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gill KA and Brindle NP: Angiopoietin-2
stimulates migration of endothelial progenitors and their
interaction with endothelium. Biochem Biophys Res Commun.
336:392–396. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shiozawa Y, Pedersen EA, Havens AM, et al:
Human prostate cancer metastases target the hematopoietic stem cell
niche to establish footholds in mouse bone marrow. J Clin Invest.
121:1298–1312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chow DC, Wenning LA, Miller WM and
Papoutsakis ET: Modeling pO(2) distributions in the bone marrow
hematopoietic compartment. II Modified Kroghian models. Biophys J.
81:685–696. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Harrison JS, Rameshwar P, Chang V and
Bandari P: Oxygen saturation in the bone marrow of healthy
volunteers. Blood. 99:3942002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang Y, Huang B, Sun L, Peng X, Chen X and
Zou X: Ginkgolide B promotes proliferation and functional
activities of bone marrow-derived endothelial progenitor cells:
involvement of Akt/eNOS and MAPK/p38 signaling pathways. Eur Cell
Mater. 21:459–469. 2011.PubMed/NCBI
|
|
31
|
Asahara T, Masuda H, Takahashi T, et al:
Bone marrow origin of endothelial progenitor cells responsible for
postnatal vasculogenesis in physiological and pathological
neovascularization. Circ Res. 85:221–228. 1999. View Article : Google Scholar
|
|
32
|
Lyden D, Hattori K, Dias S, et al:
Impaired recruitment of bone-marrow-derived endothelial and
hematopoietic precursor cells blocks tumor angiogenesis and growth.
Nat Med. 7:1194–1201. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Langer HF, May AE, Vestweber D, De Boer
HC, Hatzopoulos AK and Gawaz M: Platelet-induced differentiation of
endothelial progenitor cells. Semin Thromb Hemost. 33:136–143.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nassiri F, Cusimano MD, Scheithauer BW, et
al: Rotondo F Endoglin (CD105): a review of its role in
angiogenesis and tumor diagnosis, progression and therapy.
Anticancer Res. 31:2283–2290. 2011.PubMed/NCBI
|
|
35
|
Fan Y, Ye J, Shen F, et al: Interleukin-6
stimulates circulating blood-derived endothelial progenitor cell
angiogenesis in vitro. J Cereb Blood Flow Metab. 28:90–98. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Silverman MD, Haas CS, Rad AM, Arbab AS
and Koch AE: The role of vascular cell adhesion molecule 1/very
late activation antigen 4 in endothelial progenitor cell
recruitment to rheumatoid arthritis synovium. Arthritis Rheum.
56:1817–1826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu Y, Ip JE, Huang J, et al: Essential
role of ICAM-1/CD18 in mediating EPC recruitment, angiogenesis, and
repair to the infarcted myocardium. Circ Res. 99:315–322. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Humpert PM, Djuric Z, Zeuge U, et al:
Insulin stimulates the clonogenic potential of angiogenic
endothelial progenitor cells by IGF-1 receptor-dependent signaling.
Mol Med. 14:301–308. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Garcia-Barros M, Paris F, Cordon-Cardo C,
et al: Tumor response to radiotherapy regulated by endothelial cell
apoptosis. Science. 300:1155–1159. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang QR, Wang F, Zhu WB, Lei J, Huang YH,
Wang BH and Yan Q: GM-CSF accelerates proliferation of endothelial
progenitor cells from murine bone marrow mononuclear cells in
vitro. Cytokine. 45:174–178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim KL, Shin IS, Kim JM, et al:
Interaction between Tie receptors modulates angiogenic activity of
angiopoietin2 in endothelial progenitor cells. Cardiovasc Res.
72:394–402. 2006. View Article : Google Scholar : PubMed/NCBI
|