Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignant diseases with 600,000 new cases reported each year
worldwide, and is the third leading cause of cancer-related
mortality (1). Although aggressive
surgery offers significant rates of cure, only 15% of patients are
eligible for optimal resection at diagnosis. The efficacy of
chemotherapy and radiotherapy for HCC remain disappointing. Gene
therapy may be a promising approach for the treatment of HCC.
The tumor suppressor gene p53 plays a key role in
cell cycle control, apoptosis and inhibition of tumor cell
proliferation. Several human tumors carry mutations in the p53 and
mutation or deletion of p53 is associated with poor prognosis and
resistance to chemotherapy and radiotherapy (2,3). In
HCC, absent p53 status is correlated with carcinogenesis, and p53
is frequently mutated in HCC and its presence indicates a poorer
prognosis (4,5). A number of groups have reported the
clinical responses to adenovirus p53 (Ad-p53) as a single agent or
combined with radiotherapy or chemotherapy, including head and neck
squamous cell carcinoma, breast cancer, non-small cell lung cancer,
glioma, bladder and esophageal cancer (6–10).Recent studies revealed that combined
gene therapy is more effective in treating the murine model of HCC
than therapy with one gene alone (11,12).
Human apurinic/apyrimidinic endonuclease (APE1) is a
dual-function protein, which has both DNA repair activity and redox
regulatory activity (13). As a
redox factor, APE1 maintains a number of transcriptional factors
including p53 in their reduced and active state, thereby regulating
their DNA-binding activity (14,15).
Several studies demonstrated that APE1 was overexpressed in several
human tumors and elevated APE1 level was associated with poor
clinical outcome (16–21). In a previous study, we constructed
chimeric adenoviral vector Ad5/F35 carrying human APE1 siRNA
(Ad5/F35-siAPE1), which inhibited APE1 expression and enhanced the
sensitivity to radiotherapy in colorectal cancer (22). Moreover, we found that
Ad5/F35-siAPE1 enhanced sensitivity to cisplatin, including ovarian
cancer (23) and non-small cell
lung cancer (24). Since silencing
of APE1 enhanced the sensitivity to radiotherapy and chemotherapy,
inhibition of APE1 protein by Ad5/F35-siAPE1 may be a promising
approach against cancer.
In the present study, we examined the therapeutic
effectiveness of combined Ad5/F35-siAPE1 and Ad-p53 in vitro
and in a murine model of HCC. Our data demonstrate that combined
gene therapy with Ad5/F35-siAPE1 and Ad-p53 was more effective than
therapy with either agent alone in HCC cells in vitro and
in vivo.
Materials and methods
Materials
Adenovirus vector Ad5/F35-siAPE1 carrying human APE1
siRNA sequence was constructed and purified as previously described
(22). Ad-p53 was obtained from
Shenzhen SiBiono GeneTech Co., Ltd. (Shenzhen, China). The control
adenovirus, Ad-EGFP and Ad5/F35-EGFP, was purchased from Vector
Gene Technology Co., Ltd. (Beijing, China). The monoclonal antibody
against hAPE1 was from Novus Biological (Littleton, CO, USA). The
antibodies directed against p53 (DO-1) and β-actin were purchased
from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The human
hepatoma cell line SMMC-7721 (carrying wild-type p53) was obtained
from the Cell Institute of Shanghai (Academia Sinica, Shanghai,
China). Cells were maintained at 37°C in a humidified incubator
under 5% CO2 and grown in Dulbecco’s modified Eagle’s
medium (DMEM) supplemented with 10% fetal bovine serum, 50 mg/ml
penicillin/streptomycin. Specific pathogen-free female athymic nude
mice, 4–6 weeks old, were purchased from Shanghai SLAC Laboratory
Animal Co., Ltd. (Shanghai, China). All experiments were carried
out in accordance with the China Animal Welfare Legislation and
were approved by the Third Military Medical University Committee on
Ethics in the Care and Use of Laboratory Animals.
Infection with adenoviruses and flow
cytometry
SMMC-7721 cells were infected with Ad5-EGFP or
Ad5/F35-EGFP with increasing multiplicities of infection (MOI) for
2 h and were then washed to remove the adenoviruses. They were
cultured for another 48 h and then analyzed for their EGFP
intensity using a FACScan (Becton-Dickinson, Mountain View, CA,
USA) or directly observed with a fluorescence microscope (1200 ECM;
Nikon, Düsseldorf, Germany).
Western blot analysis
Ten million cells were supplemented with 100 μl cell
lysis solution precooled to 0°C left on ice for 30 min, centrifuged
at 12,000 rpm and placed at room temperature for 10 min.
Supernatants were supplemented with 2X sodium dodecyl sulfate (SDS)
gel loading buffer and denatured at 100°C for 5 min. Then, 20 μg of
protein from nuclear, cytosolic, or mitochondrial fractions was
applied to 10% SDS-polyacrylamide gels and electrophoresed to
resolve proteins. The proteins were then transferred to
polyvinylidene difluoride (PVDF) membranes and blocked in
Tris-buffered saline and Tween-20 (TBST) [50 mM Tris-HCl, pH 7.5,
150 mM NaCl and 0.1% (v/v) Tween-20] containing 5% (w/v) defatted
milk and incubated with the specific primary antibody. The
membranes were then washed three times in TBST and incubated with a
horseradish peroxidase-conjugated secondary antibody (1:2,000)
(Pierce) for 1 h at 37°C. The membranes were then washed three
times with TBST and the blots were reacted with chemiluminescence
reagents and revealed with BioMax Light film (Kodak, Rochester, NY,
USA). Band intensities were analyzed using the Gel Doc 2000
apparatus and software (Quantity One; Bio-Rad Laboratories,
Hercules, CA, USA). Suppliers of incubation conditions for
antibodies used for western blot analysis were as follows:
anti-APE1 monoclonal (Novus), 1 h at 37°C, dilution 1:5,000;
anti-p53 monoclonal antibody (DO-1), overnight at 4°C, dilution
1:500; anti-β-actin monoclonal (Santa Cruz Biotechnology), 1 h at
37°C, dilution 1:2,000.
MTT assay
Cells (1×105 cells/ml) were immediately
inoculated into 96-well plates (200 μl/well) in triplicate
post-irradiation. After 72 h, 15 μl MTT (5 mg/ml) was added to each
well and incubated for 4 h in a humidified atmosphere (37°C, 5%
CO2). The culture medium was removed and 200 μl of DMSO
was added into each well. The plates were shaken on a swing bed for
10 min and the OD value at 492 nm was determined using a microplate
reader.
In vivo experiments
SMMC-7721 cells (5×106) in 100 μl
phosphate-buffered saline (PBS) were injected subcutaneously into
the right flank of nude mice. When the tumors grew to ~100
mm3 on Day 12 after cell injection, 16 tumor-bearing
mice were randomized into the following four treatment groups (n=4
animals per group): i) Ad5/F35-EGFP+Ad-EGFP; ii)
Ad5/F35-siAPE1+Ad-EGFP; iii) Ad5/F35-EGFP+Ad-p53; iv)
Ad5/F35-siAPE1+Ad-p53. Tumor-bearing mice were injected with the
aforementioned agents directly into the tumors every 3 days. On Day
18, all nude mice were sacrificed, and each tumor was isolated and
measured. The maximum diameters (Dmax) and minimum diameters
(Dmin) of xenografts were measured before each treatment and
after mice were sacrificed, and tumor size was calculated according
to the following formula: tumor size (mm3) =
(Dmax × Dmin2)/2.
Immunohistochemical analysis of tumors
for APE1 and p53
The expressions of APE1 and p53 protein were
analyzed using immunohistochemistry. Sections from
paraffin-embedded tumors were incubated overnight with mouse
anti-human APE1 monoclonal antibody (Novus) at a 1:2,000 dilution
or anti-p53 antibody (DO-1) (Santa Cruz Biotechnology) at a 1:500
dilution, and then incubated with goat anti-mouse secondary
antibody (Pierce, Rockford, IL, USA). Antigen-antibody complexes
were visualized by incubation with 3,3′-diaminobenzidine (DAB)
substrate and counterstained with diluted Harris hematoxylin.
TUNEL assay for apoptosis
In vitro, coverslips covered with SMMC-7721
cells were rinsed in PBS and fixed with 4% paraform, and then
measured by terminal dUTP nick end labeling (TUNEL) staining using
the ApopTag kit (Intergen, Purchase, NY, USA). The formalin-fixed
and paraffin-embedded 5 μm sections of all tumor samples were also
analyzed for apoptosis by TUNEL assay. The extent of apoptosis was
evaluated by counting the positive brown-stained cells as well as
the total number of cells at 10 arbitrarily selected ×100
microscope fields in a blinded manner.
Statistical analysis
All quantitative data were obtained from at least
three independent experiments and expressed as the means ± SD. The
statistical significance of differences was determined by the
Student’s two tailed t-test in two groups and by one-way analysis
of the variance (ANOVA) using computer SPSS software SPSS 10.0
(SPSS Inc., Chicago, IL, USA). P<0.05 was considered to indicate
a statistically significant difference.
Results
Infectivity of adenovirus vectors Ad5/F35
and Ad5 to human hepatoma cells
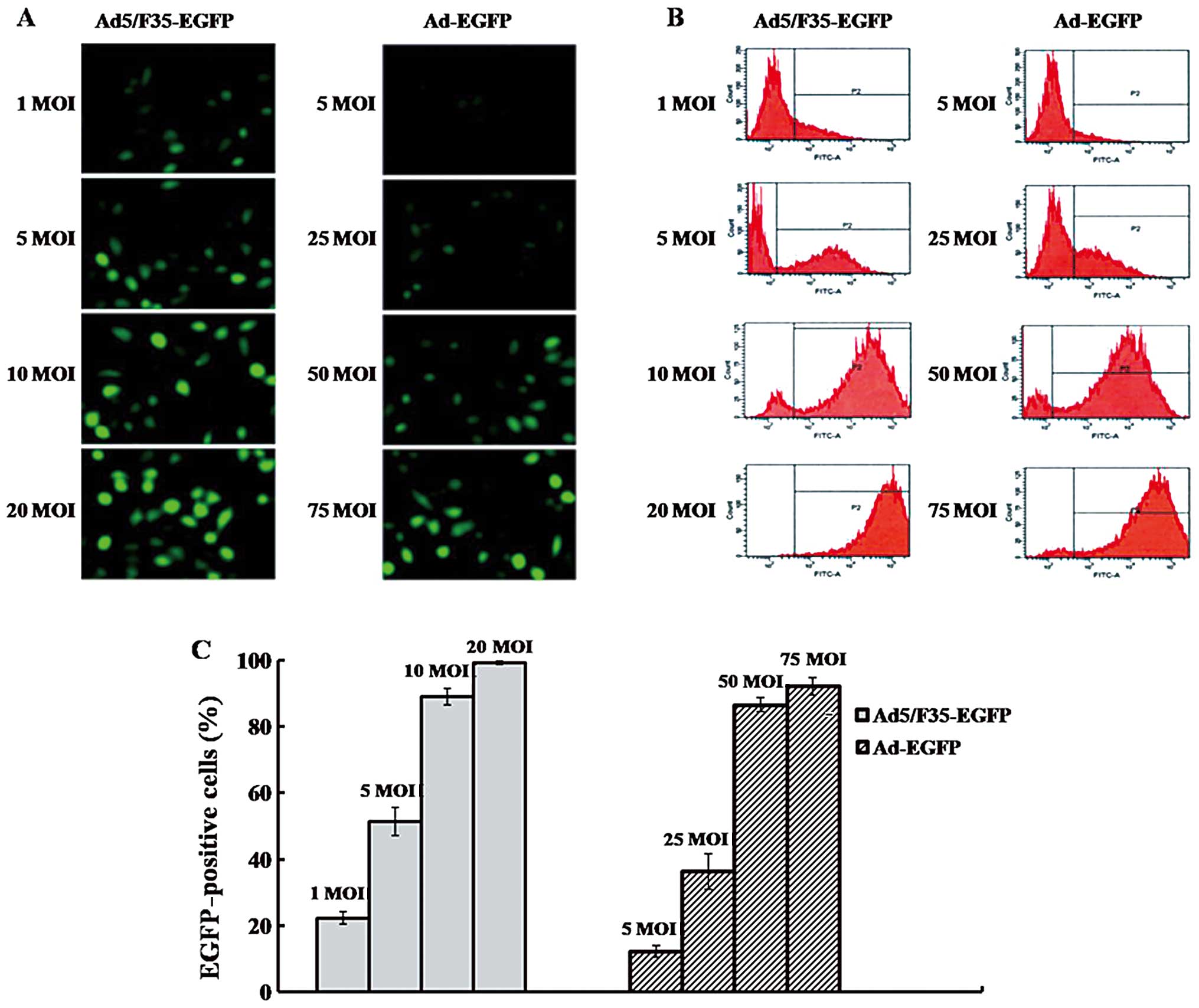
We examined the transduction efficiency of
Ad5/F35-EGFP and Ad5-EGFP to SMMC-7721 cells. As the same promoter
was used to transcribe the EGFP gene in all vectors, the
EGFP-positive population was primarily determined by the adenovirus
infectivity. We thereby regarded the percentage of positive EGFP
cells as putative infectivity of adenoviruses in the present study.
The infectivity of Ad5/F35-EGFP and Ad5-EGFP increased in a
dose-dependent manner (Fig. 1A and
B). The infectivity with 10 MOI Ad5/F35-EGFP following
transduction was 89%, and increased to 99.1% with 20 MOI
Ad5/F35-EGFP. Moreover, the infectivity of 50 MOI Ad5-EGFP was
86.5%, and reached 92.1% with 75 MOI Ad5-EGFP (Fig. 1C).
Cell survival following adenovirus
infection in SMMC-7721 cells
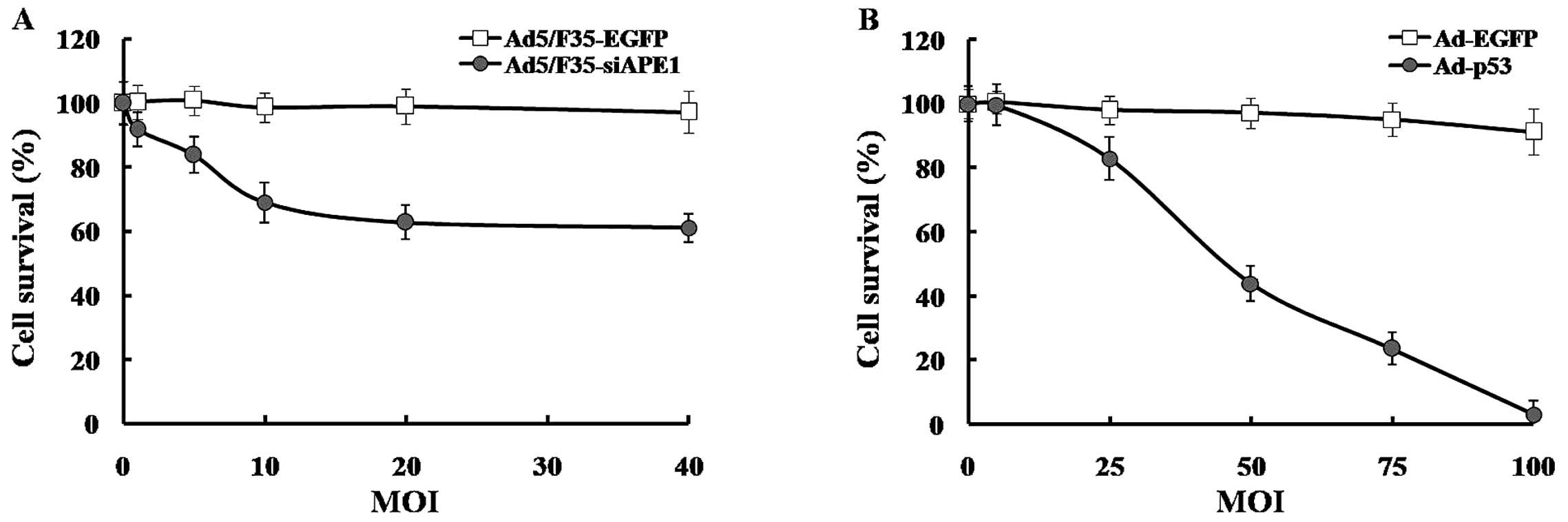
To investigate the cell survival of SMMC-7721 cells
following adenovirus infectivity, MTT assays were performed. As
shown in Fig. 2A, Ad5/F35-siAPE1
inhibited cell growth in a dose-dependent manner, compared with the
Ad5/F35-EGFP group. Although the cell survival decreased to 61.06%
after 40 MOI Ad5/F35-siAPE1, there was no significant difference
compared with that after 20 MOI Ad5/F35-siAPE1, which had an
inhibition rate of 63%. Since 99.1% SMMC-7721 cells showed
EGFP-positive cells (Fig. 1C) and
the cell survival reached in 98.91% after 20 MOI Ad5/F35-EGFP
infection (Fig. 2A), the dose of 20
MOI was used in following assays.
Subsequently, we showed that the cell proliferation
of SMMC-7721 cells was inhibited by Ad-p53 in a dose-dependent
manner, and 100 MOI Ad-p53 almost completely suppressed the cell
growth. At lower doses, Ad-EGFP caused slight damage to cells, but
the cell survival declined to 91.31% after a higher 100 MOI Ad-EGFP
transfection (Fig. 2B). Due to the
high adenovirus infectivity of SMMC-7721 cells at 50 MOI Ad-EGFP
and significant inhibition caused by 50 MOI Ad-p53, the dose of 50
MOI was used in following assays.
Time course and dose-dependent expression
of APE1 and p53 proteins following adenovirus transfection
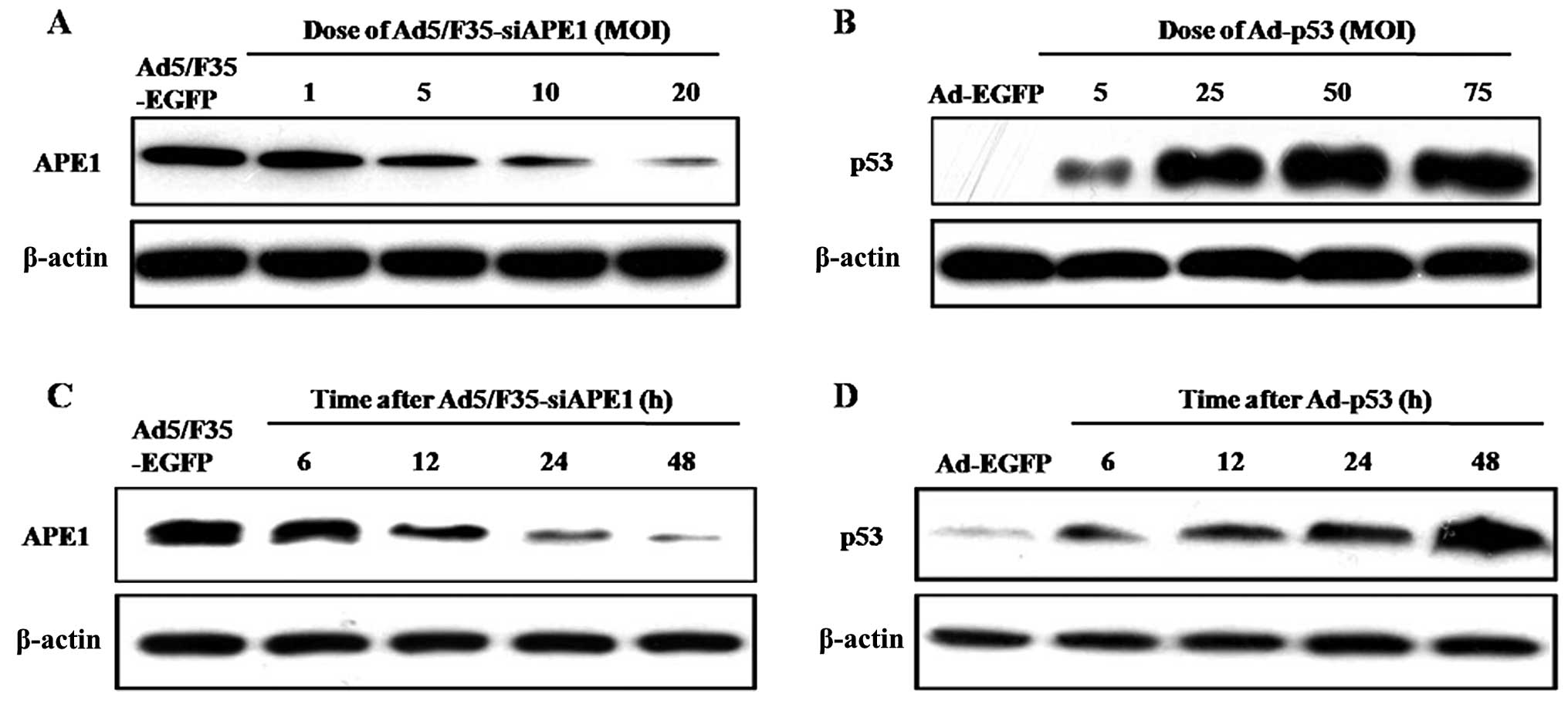
We examined the expression of APE1 protein in
SMMC-7721 cells following Ad5/F35-siAPE1 treatment. Fig. 3A shows that Ad5/F35-siAPE1 inhibited
APE1 protein expression levels in a dose-dependent manner, and the
inhibition rate of APE1 reached >90% with 20 MOI of
Ad5/F35-siAPE1. Then, we analyzed the expression of p53 protein
following Ad-p53 treatment in SMMC-7721 cells, and found that p53
protein increased in a dose-dependent manner following infection
with Ad-p53 (Fig. 3B).
We further investigated the time-dependent effect of
APE1 following Ad5/F35-siAPE1 transfection, and found that the APE1
expression level was markedly decreased in a time-dependent manner
in 20 MOI Ad5/F35-siAPE1-transfected SMMC-7721 cells, and the
suppression rate of APE1 reached ~90% at 48 h after infection
(Fig. 3C). We also observed that
there was a time-dependent increase of p53 protein in 50 MOI
Ad-p53-transfected SMMC-7721 cells (Fig. 3D).
Combined Ad5/F35-siAPE1 and Ad-p53
potentiates cell growth inhibition and apoptosis induction in
vitro
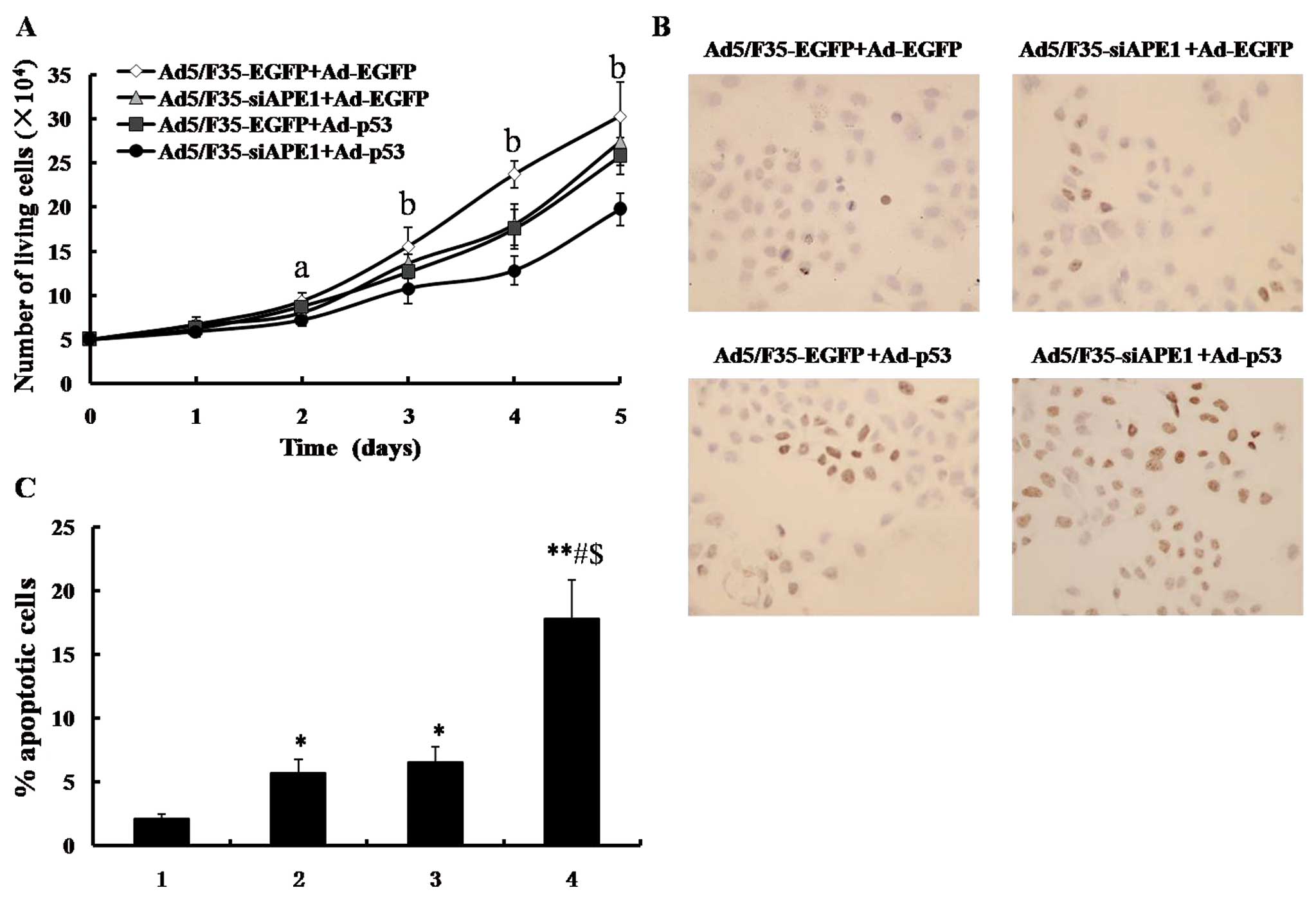
To examine the suppression of Ad5/F35-siAPE1 in
combination with Ad-p53, the cellular proliferation capacity was
detected by MTT assay in the SMMC-7721 cell line. As shown in
Fig. 4A, a significant cell
proliferation inhibition was observed in the Ad5/F35-EGFP+Ad-p53,
Ad5/F35-siAPE1+Ad-EGFP or Ad5/F35-siAPE1+Ad-p53 group, compared
with the Ad5/F35-EGFP+Ad-EGFP control group. The combined
Ad5/F35-siAPE1 and Ad-p53 group caused a significant inhibition of
cell growth compared with the Ad5/F35-siAPE1 or the Ad-p53
treatment group alone. However, no statistical differences were
found between the Ad5/F35-EGFP+Ad-p53 and Ad5/F35-siAPE1+Ad-EGFP
groups.
In another series of experiments, cells were
collected at 48 h following adenovirus treatment, and apoptotic
cells were measured by TUNEL assay. As shown in Fig. 4B and C, the apoptotic rates of the
Ad5/F35-EGFP+Ad-EGFP control group, Ad5/F35-siAPE1+Ad-EGFP,
Ad5/F35-EGFP+Ad-p53 and Ad5/F35-siAPE1+Ad-p53 were 2.10±0.41,
5.73±1.02, 6.57±1.19 and 17.82±3.02%, respectively. Ad5/F35-siAPE1
or Ad-p53 alone induced a slight increase in apoptotic cells
compared with the Ad5/F35-EGFP+Ad-EGFP control group, while
Ad5/F35-siAPE1 combined with Ad-p53 significantly increased cell
apoptosis induction. Collectively, our data demonstrate that
combined Ad5/F35-siAPE1 and Ad-p53 enhance cell growth inhibition
and apoptosis induction in the human SMMC-7721 cell line.
APE1 and p53 protein expression in
vivo
To investigate the expression levels of APE1 and p53
protein with or without Ad5/F35-siAPE1 and/or Ad-p53 treatment,
immunohistochemistry was performed in human SMMC-7721 xenografts.
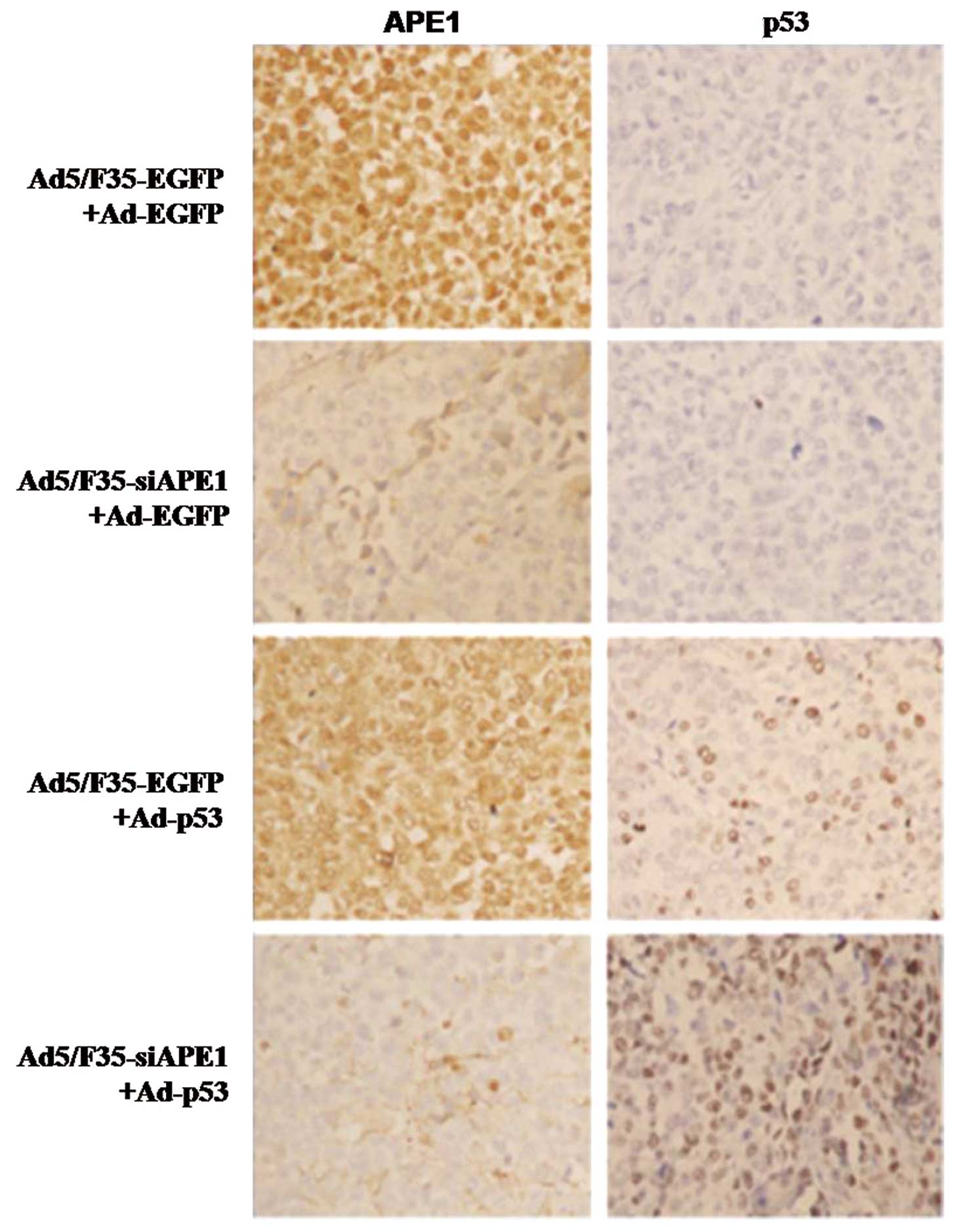
In the Ad5/F35-EGFP+Ad-EGFP control group, APE1 protein was
predominantly localized in the nucleus of tumor cells, and the
expression level of APE1 was revealed to be the same in the
Ad5/F35-EGFP+Ad-EGFP and Ad5/F35-EGFP+Ad-p53 group, while the
expression level of APE1 significantly decreased in the
Ad5/F35-siAPE1+Ad-EGFP and Ad5/F35-siAPE1+Ad-p53 groups (Fig. 5). Moreover, no p53 protein
expression was observed in the Ad5/F35-EGFP+Ad-EGFP and
Ad5/F35-siAPE1+Ad-EGFP groups, whereas the p53 protein level
significantly increased in the Ad5/F35-EGFP+Ad-p53 and
Ad5/F35-siAPE1+Ad-p53 groups (Fig.
5).
Combined treatment with Ad5/F35-siAPE1
and Ad-p53 inhibits tumor growth
We showed that the expression of APE1 protein in
SMMC-7721 xenografts was inhibited by Ad5/F35-siAPE1, and the p53
protein expression was potentiated by Ad-p53. To investigate
whether the combination of Ad5/F35-siAPE1 and Ad-p53 could enhance
the inhibition of tumor growth in vivo, tumor-bearing mice
were injected intratumorally with or without Ad5/F35-siAPE1 and/or
Ad-p53 every three days. We initiated in vivo tumor therapy
on Day 0, which corresponded to 12 days following SMMC-7721 cell
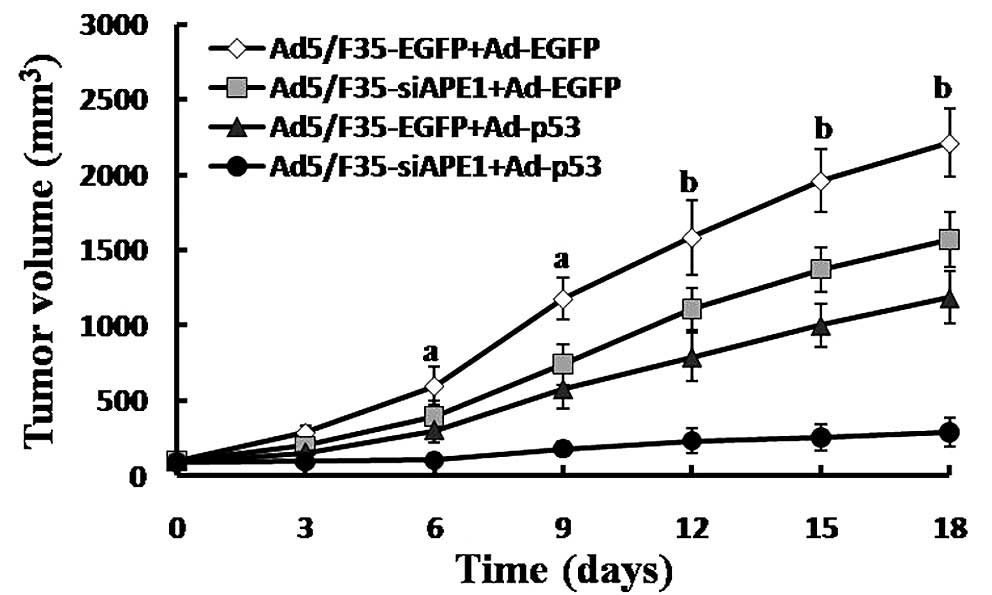
injection. As shown in Fig. 6, we
noted an inhibition of tumor growth in groups of mice treated with
Ad5/F35-siAPE1+Ad-p53, Ad5/F35-siAPE1+Ad-EGFP and
Ad5/F35-EGFP+Ad-p53 vs. the control group. Furthermore,
Ad5/F35-siAPE1 in combination with Ad-p53 caused a significant
inhibition of tumor growth compared with the Ad5/F35-siAPE1+Ad-EGFP
or the Ad5/F35-EGFP+Ad-p53 group. On Day 18, the tumor-inhibition
rates of the Ad5/F35-siAPE1+Ad-EGFP group, the Ad5/F35-EGFP+Ad-p53
group and the Ad5/F35-siAPE1+Ad-p53 group were 29.0, 46.47 and
86.87%, respectively (P<0.05).
Combination of Ad5/F35-siAPE1 and Ad-p53
enhances apoptosis induction in vivo
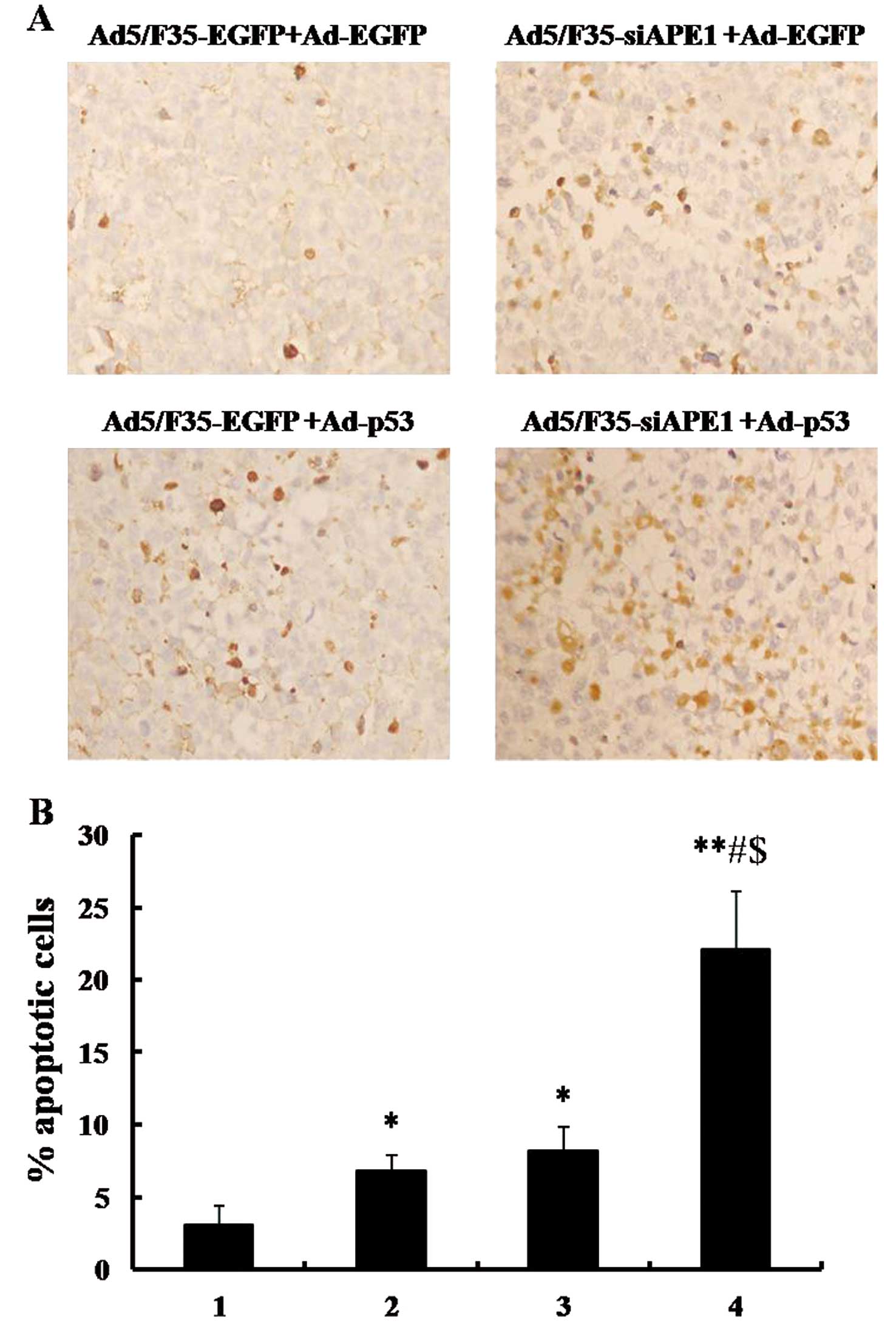
To investigate the effects of combined
Ad5/F35-siAPE1 and Ad-p53 on apoptosis induction in vivo,
the apoptosis index was calculated using an in situ
apoptosis detection assay. As shown in Fig. 7A and B, apoptosis index in
Ad5/F35-siAPE1+Ad-p53, Ad5/F35-EGFP+Ad-p53, Ad5/F35-siAPE1+Ad-EGFP
and Ad5/F35-EGFP+Ad-EGFP was 22.12±3.99, 8.23±1.60, 6.85±1.11 and
3.06±1.35%, respectively. The Ad5/F35-siAPE1+Ad-EGFP and Ad5/F35-
EGFP+Ad-p53 groups induced a slightly higher apoptosis index than
the Ad5/F35-EGFP+Ad-EGFP control group, whereas Ad5/F35-siAPE1 in
combination with Ad-p53 caused a significantly higher apoptosis
index.
Discussion
The p53 gene, regarded as the genome guardian of
cells, plays an important role in cell cycle control, apoptosis and
tumor growth inhibition. It is absent or mutated in approximately
half of all types of human cancer (25,26).
The incidence of the p53 mutation was 61% in HCC and its presence
indicates a poorer prognosis (4,5).
Wild-type p53 promotes cell cycle arrest and apoptosis of tumor
cells following radiation or chemotherapy, but p53 loss abrogates
the effective apoptotic response and induces radio- and
chemoresistance (2,3). As a potentially effective approach,
clinical trials of Ad-p53 as a single agent or combined with radio-
or chemotherapy are ongoing in patients with head and neck squamous
cell carcinoma, non-small cell lung cancer, glioma, breast cancer,
bladder and esophageal cancer (7–10,27).
However, gene transfer of p53 alone does not always have a positive
therapeutic outcome in all human types of cancer (7,28,29),
thus, combined gene therapy is urgently required. In the present
study, Ad-p53 alone inhibited cell proliferation of the SMMC-7721
cell line, increased p53 expression levels and induced partial
tumor regression in an HCC murine model. These data are in line
with previous reports (11,12), suggesting the potential role of
Ad-p53 in the treatment of HCC.
APE1, with dual functions of DNA repair and redox
regulation activity of transcription factors, is generally
abundantly expressed in most cells and is mainly localized in
nuclei of normal cells (13). APE1
is the major apurinic/apyrimidinic (AP) endonuclease in the DNA
base excision repair (BER) pathway, which plays a critical role in
repairing DNA damage (30). In
addition to its DNA repair function, APE1 is also a multifunctional
protein that is involved in other crucial cellular processes,
including the response to oxidative stress, regulation of
transcription factors, cell cycle control and apoptosis (31). As a redox factor, APE1 controls the
redox status of a number of transcription factors, including
hypoxia inducible factor-α (HIF-α), nuclear factor-κB (NF-κB), p53,
cAMP response element binding protein (CREB), thyroid transcription
factor 1 (TTF-1), paired box 5 (Pax5) and 8 (Pax8) (31).
The functional importance of APE1 is underscored by
the embryonic lethality of APE1 murine knockouts at very early
stages (E3.5-E9.5) and the lack of viable cell lines completely
deficient for the APE1 gene (32,33).
Several studies demonstrated that APE1 was highly expressed in
several human tumors, and increased APE1 expression has been shown
to be associated with resistance to radiotherapy and chemotherapy
(16–21). Previous studies using DNA antisense
methodology implicated APE1 in cellular resistance to a variety of
agents that cause alkylation and oxidative DNA damage. Targeted
reduction of APE1 by specific antisense oligonucleotides in human
HeLa, rat glioma, or human lung carcinoma cells renders mammalian
cells hypersensitive to alkylating and oxidative agents, as well as
ionizing radiation (31,34–36).
Therefore, APE1 has been approved to be a viable target for cancer
therapeutics. In the present study, we used Ad5/F35-si APE1 in a
murine model of HCC using the SMMC-7721 cell line. Data presented
here show that intratumoral injection of Ad5/F35-siAPE1 was able to
suppress APE1 expression and tumor growth and increase apoptosis.
These data are in accordance with a report by Xiang et
al(22), indicating the
effective role of Ad5/F35-si APE1 in the treatment of cancer.
As the therapeutic efficacy of monogene therapy is
disappointing, combined multi-targeting gene therapy is urgently
required to alleviate the suffering caused by cancer and to
minimize the mortality rate. The multi-targeting gene therapy by
combination of epidermal growth factor receptor ligand epiregulin
(EREG), cyclooxygenase 2 (COX2), and matrix metalloproteinases 1
and 2 (MMP1 and MMP2) produced a significant inhibition of
pulmonary metastasis of human breast cancer compared with either
therapy alone (37). Compared with
the suicide gene thymidine kinase and interleukin 2 (IL-12) gene
therapy alone, the combination resulted in inhibited tumor growth
and prolonged animal survival in a murine HCC model (11,12).
Combined B7.1 and angiostatin completely eradicated large (0.4 cm
in diameter) EL-4 lymphomas which were established in syngeneic
C57BL/6 mice, whereas B7.1 and angiostatin monotherapies were
ineffective (38). These studies
indicate that combined gene therapy may be an effective approach
against cancer. In our study, we combined Ad5/F35-siAPE1 and
Ad-p53. Our data demonstrate that combination therapy is more
effective in vitro and in treating an HCC murine model than
therapy with a single vector, which is in accordance with the
previous studies in HCC (11,12).
In the HCC murine model here, we found that the tumor-inhibition
rate and apoptosis index in the combined Ad5/F35-siAPE1 plus Ad-p53
group significantly increased. However, treatment with either
Ad5/F35-siAPE1 or Ad-p53 induced a slight increase in
tumor-inhibition rate and apoptosis index.
In conclusion, our data demonstrate that combination
therapy of Ad5/F35-siAPE1 and Ad-p53 was more efficient than
therapy with either vector alone in vitro and in a murine
HCC model. Ad-p53, as a gene therapy agent, can be combined with
Ad5/F35-siAPE1 due to its chemo- and radiosensitized efficacy in
cancer, and represents a potential therapeutic approach for
patients with cancer. Furthermore, the clinical use of
Ad5/F35-siAPE1 in combination with Ad-p53 has yet to be explored
and warrants further investigations in human HCC patients.
Acknowledgements
The study was financially supported by the National
Natural Science Foundation of China (no. 30872975 to D.W.).
References
|
1
|
Bosch FX, Ribes J, Cleries R and Diaz M:
Epidemiology of hepatocellular carcinoma. Clin Liver Dis.
9:191–211. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lowe SW, Ruley HE, Jacks T and Housman DE:
p53-dependent apoptosis modulates the cytotoxicity of anticancer
agents. Cell. 74:957–967. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eisold S, Linnebacher M, Ryschich E, et
al: The effect of adenovirus expressing wild-type p53 on
5-fluorouracil chemosensitivity is related to p53 status in
pancreatic cancer cell lines. World J Gastroenterol. 10:3583–3589.
2004.PubMed/NCBI
|
|
4
|
Honda K, Sbisa E, Tullo A, et al: p53
mutation is a poor prognostic indicator for survival in patients
with hepatocellular carcinoma undergoing surgical tumour ablation.
Br J Cancer. 77:776–782. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hsia CC, Nakashima Y, Thorgeirsson SS, et
al: Correlation of immunohistochemical staining and mutations of
p53 in human hepatocellular carcinoma. Oncol Rep. 7:353–356.
2000.PubMed/NCBI
|
|
6
|
Clayman GL, Frank DK, Bruso PA and
Goepfert H: Adenovirus-mediated wild-type p53 gene transfer as a
surgical adjuvant in advanced head and neck cancers. Clin Cancer
Res. 5:1715–1722. 1999.PubMed/NCBI
|
|
7
|
Nemunaitis J, Swisher SG, Timmons T, et
al: Adenovirus-mediated p53 gene transfer in sequence with
cisplatin to tumors of patients with non-small-cell lung cancer. J
Clin Oncol. 18:609–622. 2000.PubMed/NCBI
|
|
8
|
Swisher SG, Roth JA, Komaki R, et al:
Induction of p53-regulated genes and tumor regression in lung
cancer patients after intratumoral delivery of adenoviral p53 (INGN
201) and radiation therapy. Clin Cancer Res. 9:93–101.
2003.PubMed/NCBI
|
|
9
|
Lang FF, Bruner JM, Fuller GN, et al:
Phase I trial of adenovirus-mediated p53 gene therapy for recurrent
glioma: biological and clinical results. J Clin Oncol.
21:2508–2518. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
INGN 201. Ad-p53, Ad5CMV-p53, adenoviral
p53, p53 gene therapy - introgen, RPR/INGN 201. Drugs R D.
8:176–187. 2007.PubMed/NCBI
|
|
11
|
Drozdzik M, Qian C, Xie X, et al: Combined
gene therapy with suicide gene and interleukin-12 is more efficient
than therapy with one gene alone in a murine model of
hepatocellular carcinoma. J Hepatol. 32:279–286. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Su H, Lu R, Ding R and Kan YW:
Adeno-associated viral-mediated gene transfer to hepatoma:
thymidine kinase/interleukin 2 is more effective in tumor killing
in non-ganciclovir (GCV)-treated than in GCV-treated animals. Mol
Ther. 1:509–515. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Duguid JR, Eble JN, Wilson TM and Kelley
MR: Differential cellular and subcellular expression of the human
multifunctional apurinic/apyrimidinic endonuclease (APE/ref-1) DNA
repair enzyme. Cancer Res. 55:6097–6102. 1995.PubMed/NCBI
|
|
14
|
Hanson S, Kim E and Deppert W: Redox
factor 1 (Ref-1) enhances specific DNA binding of p53 by promoting
p53 tetramerization. Oncogene. 24:1641–1647. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jayaraman L, Murthy KG, Zhu C, Curran T,
Xanthoudakis S and Prives C: Identification of redox/repair protein
Ref-1 as a potent activator of p53. Genes Dev. 11:558–570. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Herring CJ, West CM, Wilks DP, et al:
Levels of the DNA repair enzyme human apurinic/apyrimidinic
endonuclease (APE1, APEX, Ref-1) are associated with the intrinsic
radiosensitivity of cervical cancers. Br J Cancer. 78:1128–1133.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Moore DH, Michael H, Tritt R, Parsons SH
and Kelley MR: Alterations in the expression of the DNA
repair/redox enzyme APE/ref-1 in epithelial ovarian cancers. Clin
Cancer Res. 6:602–609. 2000.PubMed/NCBI
|
|
18
|
Robertson KA, Bullock HA, Xu Y, et al:
Altered expression of Ape1/ref-1 in germ cell tumors and
overexpression in NT2 cells confers resistance to bleomycin and
radiation. Cancer Res. 61:2220–2225. 2001.PubMed/NCBI
|
|
19
|
Bobola MS, Blank A, Berger MS, Stevens BA
and Silber JR: Apurinic/apyrimidinic endonuclease activity is
elevated in human adult gliomas. Clin Cancer Res. 7:3510–3518.
2001.PubMed/NCBI
|
|
20
|
Koukourakis MI, Giatromanolaki A,
Kakolyris S, et al: Nuclear expression of human
apurinic/apyrimidinic endonuclease (HAP1/Ref-1) in head-and-neck
cancer is associated with resistance to chemoradiotherapy and poor
outcome. Int J Radiat Oncol Biol Phys. 50:27–36. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang D, Luo M and Kelley MR: Human
apurinic endonuclease 1 (APE1) expression and prognostic
significance in osteosarcoma: enhanced sensitivity of osteosarcoma
to DNA damaging agents using silencing RNA APE1 expression
inhibition. Mol Cancer Ther. 3:679–686. 2004.
|
|
22
|
Xiang DB, Chen ZT, Wang D, et al: Chimeric
adenoviral vector Ad5/F35-mediated APE1 siRNA enhances sensitivity
of human colorectal cancer cells to radiotherapy in vitro and in
vivo. Cancer Gene Ther. 15:625–635. 2008. View Article : Google Scholar
|
|
23
|
Zhang Y, Wang J, Xiang D, Wang D and Xin
X: Alterations in the expression of the apurinic/apyrimidinic
endonuclease-1/redox factor-1 (APE1/Ref-1) in human ovarian cancer
and indentification of the therapeutic potential of APE1/Ref-1
inhibitor. Int J Oncol. 35:1069–1079. 2009.PubMed/NCBI
|
|
24
|
Wang D, Xiang DB, Yang XQ, et al: APE1
overexpression is associated with cisplatin resistance in non-small
cell lung cancer and targeted inhibition of APE1 enhances the
activity of cisplatin in A549 cells. Lung Cancer. 66:298–304. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Levine AJ, Momand J and Finlay CA: The p53
tumour suppressor gene. Nature. 351:453–456. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hollstein M, Sidransky D, Vogelstein B and
Harris CC: p53 mutations in human cancers. Science. 253:49–53.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Clayman GL, el-Naggar AK, Lippman SM, et
al: Adenovirus-mediated p53 gene transfer in patients with advanced
recurrent head and neck squamous cell carcinoma. J Clin Oncol.
16:2221–2232. 1998.PubMed/NCBI
|
|
28
|
Roth JA, Nguyen D, Lawrence DD, et al:
Retrovirus-mediated wild-type p53 gene transfer to tumors of
patients with lung cancer. Nat Med. 2:985–991. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Swisher SG, Roth JA, Nemunaitis J, et al:
Adenovirus-mediated p53 gene transfer in advanced non-small-cell
lung cancer. J Natl Cancer Inst. 91:763–771. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fleck O and Nielsen O: DNA repair. J Cell
Sci. 117:515–517. 2004. View Article : Google Scholar
|
|
31
|
Evans AR, Limp-Foster M and Kelley MR:
Going APE over ref-1. Mutat Res. 461:83–108. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xanthoudakis S, Smeyne RJ, Wallace JD and
Curran T: The redox/DNA repair protein, Ref-1, is essential for
early embryonic development in mice. Proc Natl Acad Sci USA.
93:8919–8923. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Larsen E, Meza TJ, Kleppa L and Klungland
A: Organ and cell specificity of base excision repair mutants in
mice. Mutat Res. 614:56–68. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Walker LJ, Craig RB, Harris AL and Hickson
ID: A role for the human DNA repair enzyme HAP1 in cellular
protection against DNA damaging agents and hypoxic stress. Nucleic
Acids Res. 22:4884–4889. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ono Y, Furuta T, Ohmoto T, Akiyama K and
Seki S: Stable expression in rat glioma cells of sense and
antisense nucleic acids to a human multifunctional DNA repair
enzyme, APEX nuclease. Mutat Res. 315:55–63. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen DS and Olkowski ZL: Biological
responses of human apurinic endonuclease to radiation-induced DNA
damage. Ann NY Acad Sci. 726:306–308. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gupta GP, Nguyen DX, Chiang AC, et al:
Mediators of vascular remodelling co-opted for sequential steps in
lung metastasis. Nature. 446:765–770. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun X, Kanwar JR, Leung E, Lehnert K, Wang
D and Krissansen GW: Angiostatin enhances B7.1-mediated cancer
immunotherapy independently of effects on vascular endothelial
growth factor expression. Cancer Gene Ther. 8:719–727. 2001.
View Article : Google Scholar : PubMed/NCBI
|